
Yeast Protein Kinase A Isoforms: A Means of Encoding Specificity in the Response to Diverse Stress Conditions?


1
Division of Molecular and Cellular Function, School of Biological Sciences, Faculty of Biology, Medicine and Health, The University of Manchester, Manchester M13 9PT, UK
2
Division of Evolution, Infection and Genomics, School of Biological Sciences, Faculty of Biology, Medicine and Health, The University of Manchester, Manchester M13 9PT, UK
*
Author to whom correspondence should be addressed.
Biomolecules 2022, 12(7), 958; https://doi.org/10.3390/biom12070958
Submission received: 10 June 2022
/
Revised: 5 July 2022
/
Accepted: 6 July 2022
/
Published: 8 July 2022
(This article belongs to the Topic Cell Signaling Pathways)
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Eukaryotic cells have developed a complex circuitry of signalling molecules which monitor changes in their intra- and extracellular environments. One of the most widely studied signalling pathways is the highly conserved cyclic AMP (cAMP)/protein kinase A (PKA) pathway, which is a major glucose sensing circuit in the yeast Saccharomyces cerevisiae. PKA activity regulates diverse targets in yeast, positively activating the processes that are associated with rapid cell growth (e.g., fermentative metabolism, ribosome biogenesis and cell division) and negatively regulating the processes that are associated with slow growth, such as respiratory growth, carbohydrate storage and entry into stationary phase. As in higher eukaryotes, yeast has evolved complexity at the level of the PKA catalytic subunit, and Saccharomyces cerevisiae expresses three isoforms, denoted Tpk1-3. Despite evidence for isoform differences in multiple biological processes, the molecular basis of PKA signalling specificity remains poorly defined, and many studies continue to assume redundancy with regards to PKA-mediated regulation. PKA has canonically been shown to play a key role in fine-tuning the cellular response to diverse stressors; however, recent studies have now begun to interrogate the requirement for individual PKA catalytic isoforms in coordinating distinct steps in stress response pathways. In this review, we discuss the known non-redundant functions of the Tpk catalytic subunits and the evolving picture of how these isoforms establish specificity in the response to different stress conditions.
1. Introduction
The ability of cells to sense and rapidly respond to the challenges that are posed by their external environment is vital for their survival. Through fitness that is conferred by natural selection, organisms have evolved mechanisms to sense changes in their environment, such as fluctuations in temperature and nutrient availability, and mount appropriate stress responses to maintain cellular homeostasis [1,2]. Signal transduction pathways play a major role in responding to stresses by controlling enzyme cascades that ultimately promote remodelling of the transcriptome and proteome in multi-stress and stress-specific manners [1,2,3,4]. These signalling cascades involve multiple steps, including the production of second messenger molecules e.g., cyclic nucleotides, the subsequent activation of kinases which phosphorylate downstream substrates, and the fine-tuning of these signals by protein phosphatases.
One key signalling pathway that is conserved among eukaryotes is the cyclic AMP (cAMP)/protein kinase A (PKA) pathway, which plays a major role in the control of cell growth and proliferation. PKA was first identified over 50 years ago as a cAMP-dependent heterotetrameric–holoenzyme complex, consisting of two catalytic (C) subunits and a regulatory (R) subunit dimer (R2C2) (reviewed in [5,6]). In S. cerevisiae, the C subunits are encoded by the genes TPK1, TPK2 and TPK3, promoting the expression of three independent isoforms, while the R subunit is encoded by a single gene, BCY1 [7]. In comparison, higher eukaryotes have evolved further complexity at the level of the PKA subunits, with several genes and alternative splicing events giving rise to multiple subunit variants [8,9]. S. cerevisiae is one of the few known fungal species to express more than two PKA catalytic subunits however [10], and has therefore been used as a model eukaryote to study the role of specificity in PKA signalling [11].
PKA activity is essential in S. cerevisiae since at least one catalytic isoform is required for viability [12]. The activity of these yeast Tpk isoforms is multifaceted and controls many developmental and adaptive processes, including cellular growth, carbohydrate metabolism, the cell cycle and the general response to stress [1,13,14,15]. Tpk1-3 are believed to be redundant in supporting many of these functions, although an increasing number of studies have highlighted various processes for which isoform specificity is a key component in achieving targeted signalling. Notably, isoform specificity has been shown as important in the response to different environmental stresses. This gives rise to key questions concerning how signalling specificity is achieved under different stress conditions if different PKA kinase isoforms are important in dictating the varied stress responses [2]. In this review, we focus on the identified functions of the individual Tpk isoforms as they exist in S. cerevisiae. We consider the strategies that are employed by yeast cells to direct and compartmentalise PKA signalling and discuss recent research that is aimed at unravelling the molecular mechanisms which encode Tpk specificity in the response to diverse stress conditions.
2. Glucose-Induced cAMP Signalling
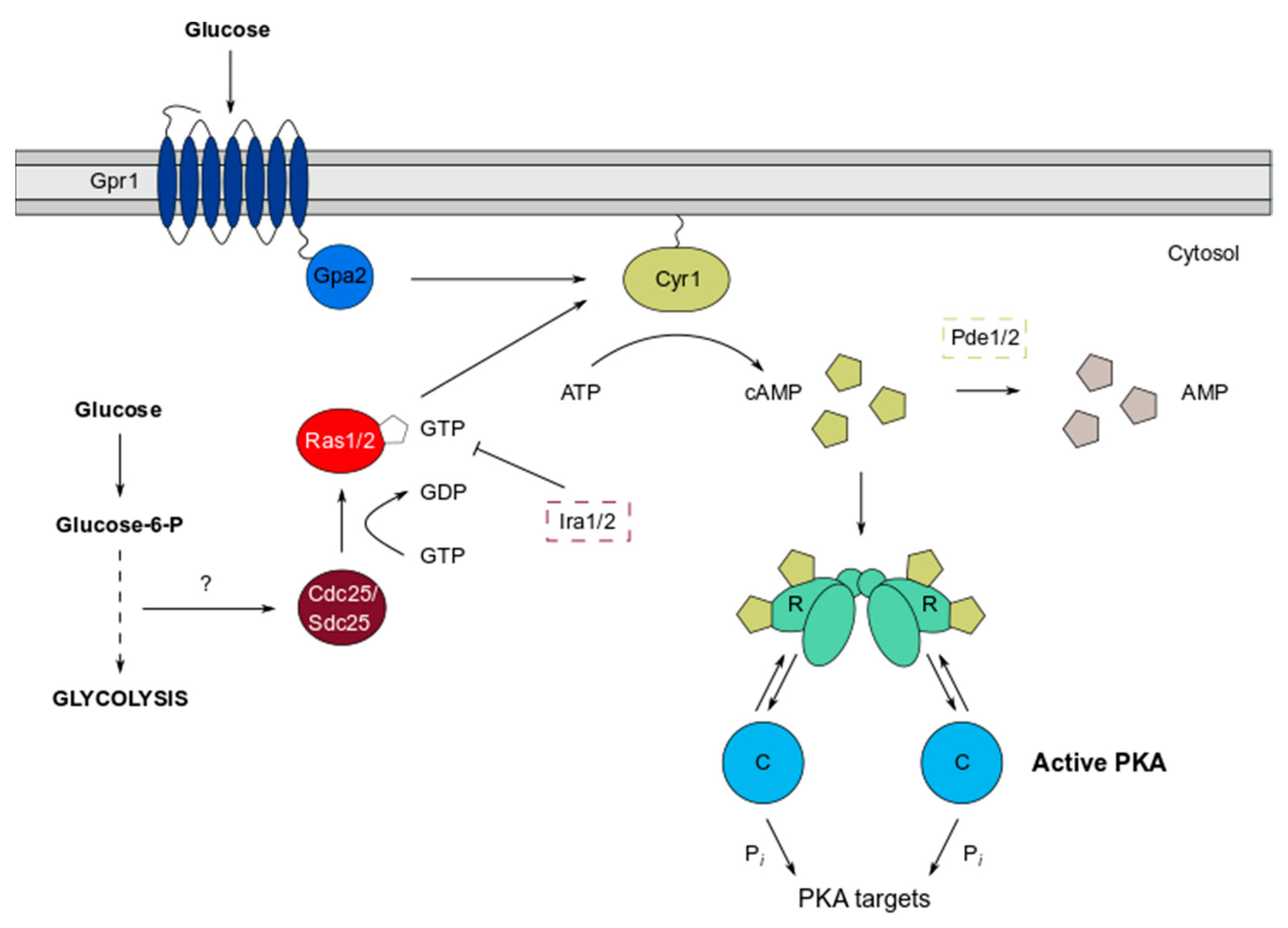
The PKA pathway in S. cerevisiae is a key mechanism by which cells detect nutrients such as glucose, facilitated by two systems functioning in concert to stimulate adenylate cyclase (Cyr1) activity (Figure 1). The transmembrane G-protein coupled receptor (GPCR) Gpr1 binds glucose as an extracellular ligand, and, acting via the G-alpha subunit Gpa2, signals to Cyr1, which triggers production of the second messenger cAMP from ATP [15,16,17,18,19,20]. However, for Cyr1 to be sensitive to the increase in glucose concentration that is signalled by Gpr1, it must first be potentiated by Ras1 and Ras2 [19]. Ras proteins are a widely studied family of small GTPases conserved in lower and higher eukaryotes and play crucial roles in multiple signalling pathways involved in cell growth and proliferation, including the cAMP/PKA and mitogen-activated protein (MAP) kinase pathways [21,22,23]. The Ras system also responds to glucose, since intracellular glucose that is metabolised by the glycolysis pathway activates a guanine nucleotide exchange factor (GEF), Cdc25, that acts on Ras1/2 [19,24,25]. GTP bound Ras1/2, anchored predominantly at the plasma membrane and organellar membranes, are activated, allowing full stimulation of Cyr1 [26,27]. Downregulation of the Ras proteins is controlled by the GTPase activating proteins (GAPs) Ira1/2, which stimulate the intrinsic GTPase activity of Ras1/2 to hydrolyse GTP to GDP, thus returning them to an inactive state [28,29]. Therefore, this complex circuitry leads to transient changes in cAMP levels, resulting in the modulation of PKA activity and the fine control of a broad range of cellular processes.
3. The Mechanics of PKA Enzymatic Control by cAMP and Regulatory Subunits
Constitutively active PKA activity is detrimental to yeast cells. For example, deletion of the BCY1 gene, resulting in a loss of PKA regulation, causes heat sensitivity, defects in sporulation and sensitivity to nutrient starvation [7]. In fact, sensitivity to multiple stresses including heat and oxidative stress broadly correlates with cellular cAMP levels, emphasising the importance of the cAMP/PKA pathway in stress tolerance [30].
Not surprisingly therefore, cAMP levels are themselves tightly regulated and can be decreased by the activities of cAMP phosphodiesterases (PDEs) which hydrolyse cAMP to AMP. S. cerevisiae has two cAMP phosphodiesterases, high affinity (Pde2) and low affinity (Pde1) enzymes [31] (Figure 1). Pde2 is thought to regulate basal cAMP levels, whereas Pde1 functions after PKA activation to dampen cAMP signalling in a negative feedback signalling loop [32]. In mammalian cells, it is well established that phosphodiesterase enzymes are compartmentalised at specific sites, which spatially constrain cAMP signalling and direct local PKA activation [33]. Evidence in yeast has shown that Pde2 localisation is altered in a PKA-dependent manner [34], suggesting that its cellular location might be important in regulating the feedback inhibition of PKA activity. However, it is currently unknown whether PDEs in yeast regulate cAMP levels and PKA activity via compartmentalisation, similar to those in mammals.
The PKA regulatory subunit (Bcy1) is the only cAMP receptor molecule that is expressed in yeast. In the absence of cAMP, Bcy1 interacts with the catalytic subunits, Tpk1-3, in an inactive R2C2 complex (reviewed in [11,16,31]). After a rise in intracellular cAMP levels, for instance after glucose detection, cAMP binds to two cyclic nucleotide binding (CNB) domains that are present on each Bcy1 monomer [6,11]. This induces allosteric changes in the holoenzyme structure, which have been proposed to result in the dissociation of the R2C2 complex and release of the catalytic subunits.
Recent studies in mammalian systems have suggested that full dissociation of the R subunits from the catalytic C subunits is not essential for PKA holoenzyme activity that is consistent with the more localised targeting of PKA substrates [35,36]. For example, the simultaneous addition of cAMP and substrate has been shown to differentially affect the extent of type I and type II PKA holoenzyme dissociation using in vitro purified proteins [37]. However, this ‘loosening’ model proposed by the Scott laboratory has been challenged, as it was later demonstrated that although the majority of C subunits are released from R subunits upon cAMP binding, the molar excess of R over C subunits likely limits their diffusion and promotes efficient recapture [38]. Interestingly, some studies have also shown a role for the PKA substrates themselves in Tpk enzyme activation [39]. Consequently, the mode of PKA activation in yeast remains poorly understood, and the classical model that the full dissociation of the R from the C subunits occurs in response to cAMP binding may represent an oversimplification of the activation process.
Tpk isoenzymes phosphorylate the same basophilic recognition sequence (RRXS*/T* motif) but phosphoproteomics has revealed that the isoforms target different substrates, raising the question as to how substrate specificity arises [40]. In mammalian cells, substrate specificity can be driven by scaffold proteins called A-kinase anchoring proteins (AKAPs), which are responsible for determining the subcellular localisation of the holoenzyme and subsequent access to its activators and substrates [41]. Unlike the single yeast R protein, there are four mammalian R protein isoforms which display differential affinities with AKAPs. A structural analysis of Bcy1 revealed that the N-terminal region that is responsible for dimerization and for docking to AKAPs (D/D domain) in higher eukaryotic R subunits is not well conserved in yeast Bcy1 [42]. The AKAP regulation of yeast PKA activity therefore remains ill-defined, although a screen for Bcy1-interacting proteins identified potential candidate proteins including the Hsp60 mitochondrial chaperone that might function in the localisation of Tpk1 to mitochondria [43].
Bcy1 is normally localised to the nucleus in glucose-grown cells but displays a broader nucleocytoplasmic localisation in carbon source-derepressed cells [44]. This carbon source-dependent localisation of Bcy1 depends on the phosphorylation of Bcy1 via the Yak1 kinase, although it is unclear whether the nucleocytoplasmic distribution of Bcy1 affects PKA activity in an isoform-specific manner. It is possible that moderating Bcy1 localisation and hence binding to the catalytic subunits provides a mechanism to globally alter PKA activity. Some preliminary data suggested that the interaction between Tpk1/2 and Bcy1 is stronger than that of Tpk3, suggesting differential regulation of the C subunits by Bcy1 [45]. However, little is known regarding individual catalytic subunit affinities with the single Bcy1 regulatory subunit and how this relates to their intracellular distribution during different growth conditions.
4. Unravelling Tpk Isoform Specificity
The three PKA catalytic subunits in S. cerevisiae were originally isolated using genetic techniques and shown to be homologous to the bovine cAMP-dependent protein kinase [12]. The deletion of all subunit genes rendered cells inviable, but the retention of any single TPK gene was sufficient to rescue this lethality, suggesting redundancy among the three subunits for essential functions [12]. Tpk1-3 are similar in terms of protein sequence and domain structure, possessing a conserved C-terminal kinase domain (~300 residues) and relatively short N- and C-terminal extensions (~40 residues) (Figure 2) [12,46]. They also share a 75% sequence similarity across the kinase domain overall, with Tpk1 and Tpk3 exhibiting a slightly higher sequence identity (88%) due to their paralogous nature [12,47]. The highest variability is present in the N-terminal domain (NTD) of each isoform, with this being consistent with the observation that the catalytic subunits of unicellular eukaryotes show considerable variation in their N-termini relative to one another [48]. Such variability is particularly evident in the NTD of Tpk2. This possesses glutamine-rich regions that are predicted to form a prion-like domain (PrLD), which are absent from Tpk1/3 [49].
Evidence that has been gathered over the last three decades has shown that catalytic subunit isoforms are not equivalent in all respects and exhibit specificity in a diverse range of processes such as filamentation, iron regulation and mitochondrial biogenesis [46,50,51,52]. It is thought that distinct PKA catalytic subunit roles might be defined by their divergent N- and C-terminal extensions, which likely facilitate interactions with different proteins and phosphorylation targets [9,46]. The concept of C-kinase anchoring proteins (CKAPs), which encompass all proteins that physically bind PKA catalytic subunits, has been suggested in mammalian cells as a means of localising C-subunits to specific subcellular compartments, thus creating ‘pools’ of isoform-specific phosphorylation [9]. For example, studies in renal cells that mutated in different PKA catalytic subunits have revealed large-scale phosphoproteomic differences, in spite of the isoforms phosphorylating nearly identical consensus motifs [53]. These differing phosphorylation patterns add weight to the idea that differences in substrate phosphorylation are mediated by differences in protein interactions. Although the regulation of mammalian PKA is more complex due to the existence of multiple catalytic subunit splice variants and regulatory subunit isoforms, these principles could be extended to other eukaryotes, including S. cerevisiae, where PKA subunits are observed at distinct subcellular locations that are dependent on nutritional status and stress exposure [54,55]. Identifying the interacting partners of Tpk1-3 could therefore serve as a way of uncovering the potential spatial regulation of the different isoforms. Alterations in the protein–protein interaction networks associated with the response to cellular stress are also likely key to the changing localisation that is observed for the Tpk isoforms during stress conditions, as discussed later.
4.1. PKA-Specific Signalling Mediates Pseudohyphal Growth
Early studies on filamentous growth in S. cerevisiae highlighted a role for Ras2 activation in stimulating the pseudohyphal response [57,58]. The RAS2val19 dominant active mutant was shown to induce filamentation via both the mitogen-activated protein kinase (MAPK) and cAMP/PKA pathways; however, it has become clear that they contribute differently to filamentous growth activation [46,59,60]. PKA signalling specificity was first described by Robertson and Fink (1998), who observed differential roles for the Tpk isoforms in supporting filamentous growth. The capacity to grow in a filamentous form is an important fungal developmental response to stress and has been suggested to enable yeast such as S. cerevisiae to forage for nutrients and/or escape accumulating toxins. The formation of chains of elongated cells that remain connected to one another and have the capacity to invade solid growth media involves alterations in the cell cycle, flocculence and budding pattern. Studies on the role of PKA showed that TPK2 deletion ameliorated the filamentous growth response, while tpk1 mutant cells showed a phenotype that was indistinguishable from the wild-type [46]. In contrast, the loss of TPK3 induced hyperactive filamentous growth, highlighting opposing roles for the Tpk2 and Tpk3 isoforms [46,61].
Mechanistically, the Tpk2-dependent regulation of filamentous growth is thought to involve the phosphorylation and targeting of two transcription factors, Sfl1 and Flo8. These factors act antagonistically in the transcriptional regulation of a key cell wall, flocculin Flo11, and FLO11 transcription represents a critical determinant of filamentous growth [46,62,63]. More specifically, Tpk2 inhibits the Sfl1 repressor, while acting as a positive regulator of Flo8 [61]. Interestingly, many standard laboratory yeast strains have lost their capacity to undergo filamentous growth through mutations in the Flo8 transcription factor [64] and strains that are mutant in FLO11 are deficient in invasion, filamentation and biofilm formation. Therefore, the overall Tpk2-dependent phosphorylation of key transcription factors is postulated to activate FLO11 transcription to promote the switch to a filamentous morphology [61].
Although earlier research considered Tpk1 to have no direct role in filamentous growth [46], more recent studies have identified Tpk1 as playing a regulatory role via the dual-specificity tyrosine kinase Yak1 [60,65,66]. Yak1 was found to be part of a regulatory cascade which induced FLO11 transcription through activation of the transcription factors Phd1 and Sok2 [60,66]. Yak1 phosphorylation by Tpk1 abolishes this activation to limit FLO11 induction [60,65]. These findings highlight how Tpk isoforms can have more discrete functions in cellular processes by acting through networks of connected signalling cascades.
4.2. Requirement for PKA Activity in Mitochondrial Function and Biogenesis
Early links with mitochondrial function came from a key study that identified functional signatures for each of the three catalytic subunits via a genome-wide microarray analysis of yeast mutants lacking the individual Tpk isoforms [50]. Using this transcriptomic analysis, loss of TPK1 was found to decrease the expression of the genes BAT1 and ILV5, both of which are involved in branched chain amino acid (BCAA) biosynthesis [67]. Removal of ILV5 causes the deletion of large portions of mitochondrial DNA (mtDNA) [68], while Bat1 also functions in mitochondrial iron homeostasis [67]. This study therefore suggests that Tpk1 is required for maintaining the integrity of the mitochondrial genome and in the derepression of BCAA biosynthesis. In this expression analysis, Tpk2 was also suggested to play a role in iron regulation, as genes that were implicated in high affinity iron uptake such as FTR1 were upregulated in a tpk2 mutant [50]. Ftr1, which encodes a plasma membrane-localised high affinity iron transporter, has since been shown to activate the PKA pathway upon addition of iron to iron-starved cells [69]. This evidence suggests that once PKA is active and the requirement for respiration and thus iron is reduced, Tpk2 could operate via a negative feedback loop to downregulate transceptor gene expression and prevent the uptake of additional iron.
Curiously, this array analysis found that no transcriptional changes were attributed to the loss of TPK3, aside from those that were described for pseudohyphal growth [50]. Tpk3 is considered to have the lowest catalytic activity of the three isoforms due to poor expression of the TPK3 gene [70]. However, later studies confirmed that Tpk3 has non-redundant roles in both respiration and mitochondrial regulation [51,52,71]. Mitochondrial PKA substrates have been described, including at least eight proteins which are differentially phosphorylated by Tpk1-3, albeit with significant overlap [72]. Experiments which first suggested that Tpk3 functions directly in mitochondrial regulation showed that tpk3 mutant cells have significantly reduced respiratory rates and possess modified mitochondria [51]. Indeed the mitochondria contain reduced levels of key respiratory enzymes such as cytochrome c, which has previously been proposed as a PKA target [51,73]. In addition to controlling enzyme content, Tpk3 was suggested to regulate elements of the mitochondrial biogenesis machinery, as in its absence, the levels of reactive oxygen species (ROS) that were generated by mitochondria were greatly increased [71]. Hyperactivation of the PKA pathway via PDE2 deletion has also been shown to produce effects on mitochondria. Mitochondria are irregularly shaped and genes that are involved in the electron transport chain and ROS detoxification are down regulated. These unusual mitochondrial phenotypes and transcriptional effects are dependent on the expression of TPK3 [52]. Taken together, these studies highlight the key role that is played by Tpk3 in regulating mitochondrial biogenesis and function.
5. Role of the cAMP/PKA Pathway in Stress Tolerance
The ability of organisms to adapt to and navigate the challenges that are posed by their external environments is vital for survival. This is of particular importance for non-motile, unicellular eukaryotes such as yeast, which are often exposed to fluctuations in nutrient availability, temperature, levels of ROS and pH, etc. One such stress response that is activated by multiple diverse stressors, including glucose starvation, heat and oxidative stress, is the environmental (or general) stress response (ESR) [1,74,75,76,77,78] (Figure 3). The ESR is the best-characterised example of a stress response programme that is regulated by PKA, as the activity of many factors which stimulate the transcription of ESR-responsive genes are regulated by PKA-mediated phosphorylation [76,79,80]. Broadly speaking, PKA is rapidly inactivated in response to various stresses, which downregulate processes that would otherwise be deleterious for cellular survival during exposure to environmental insults [81]. These include transcription, translation, glycogen synthesis, cell cycle progression and cell growth. The removal of PKA activity simultaneously derepresses other key signalling molecules including selected kinases and transcription factors (TFs), which are then free to induce a multitude of genes that promote stress resistance and survival [16].
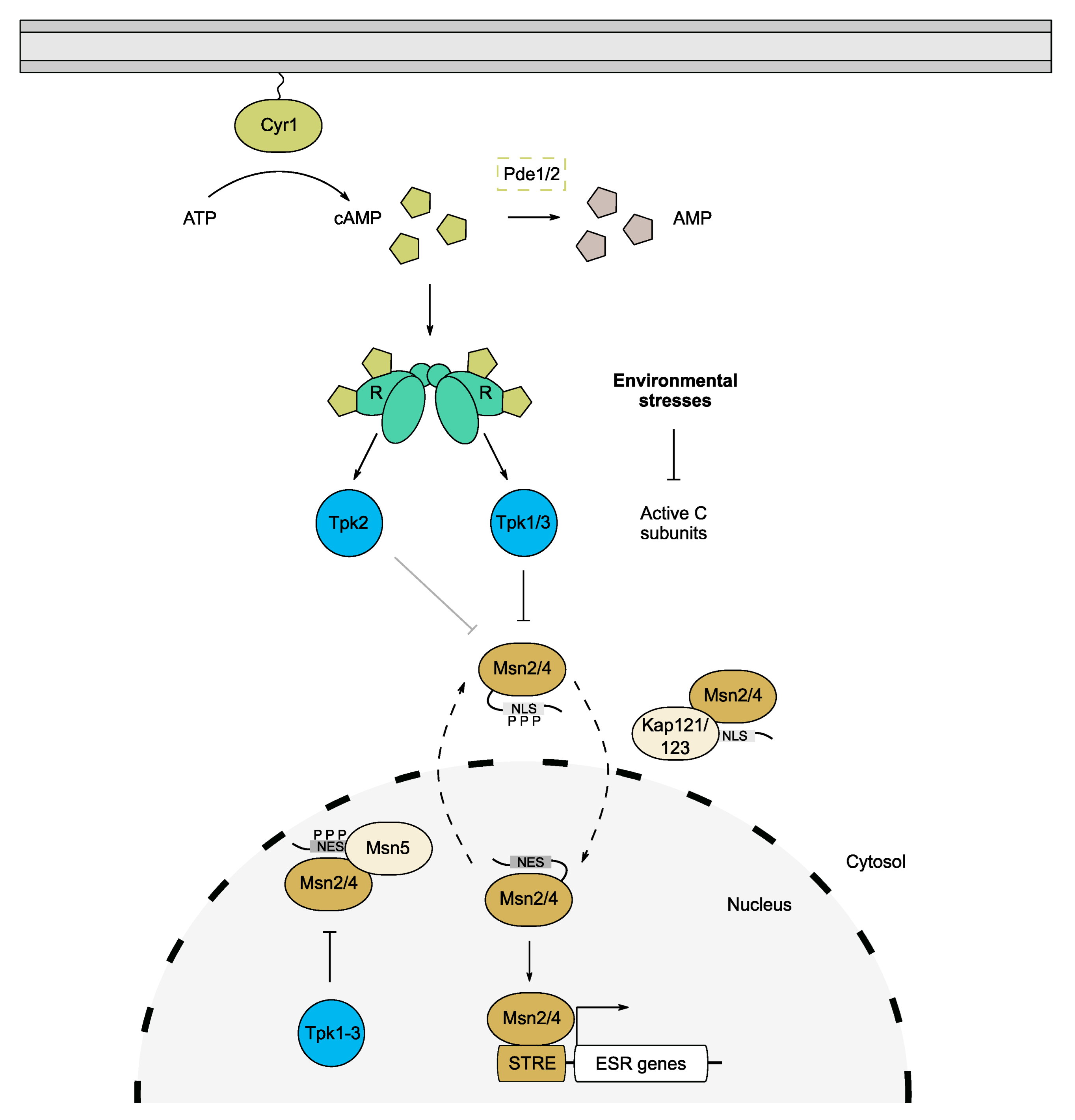
A large proportion of genes that are induced by the ESR contain a stress response element (STRE) in their promoter regions which acts as a binding site for the homologous zinc-finger TFs, Msn2 and Msn4 [16,76,82]. The STRE was originally identified as part of a Hsf1-independent transcription induction mechanism for the heat-induced genes, DDR2 and CTT1 [83,84]. Msn2/4 are among the most significant downstream targets of PKA, as their phosphorylation at functionally important motifs affords PKA control over the expression of a broad range of genes [1,80,85]. Additionally, the loss of MSN2 and MSN4 was found to rescue the lethality of a triple tpk mutant, confirming the key role that these transcription factors play in moderating the global effects of cAMP/PKA on cell growth [74,80]. A subsequent study revealed that Tpk1 and Tpk3 play more predominant roles in suppressing Msn2/4 activity, highlighting potential isoform specificity in controlling the ESR (Figure 3) [86].
Both Msn2 and Msn4 exhibit oscillatory behaviour and translocate between the nucleus and cytoplasm in a PKA- and karyopherin-dependent manner [76,80,87,88]. Under normal growth conditions, high PKA activity constrains the dynamics of Msn2/4 via increased phosphorylation of a conserved nuclear localisation signal (NLS), thus confining the majority of Msn2/4 molecules to the cytoplasm [88,89]. Conversely, after stress and the subsequent lowering of PKA activity, Msn2/4 become dephosphorylated and hence bind to nuclear import factors, leading to their accumulation in the nucleus [88]. In the case of Msn2, Kap121 and Kap123 are involved in its nuclear targeting [90], while its export is regulated by Msn5 via binding to its nuclear export sequence (NES) (Figure 3) [91]. The Msn2 NES is phosphorylated by nuclear PKA which promotes binding to Msn5 and increases the rate of its exit from the nucleus during non-stress conditions (Figure 3) [83,88]. This is abrogated under stress conditions, which retains Msn2 in the nucleus and allows binding to STRE-containing genes [87]. The retention of Msn2 in the nucleus can be detrimental however, and chronic nuclear accumulation results in degradation of the protein to avert a prolonged ESR [87]. The relative contributions of nuclear and cytosolic pools of Tpk proteins in regulating Msn2/4 oscillations are not well understood. It is therefore plausible that each isoform exhibits specificity for phosphorylating the NLS vs. NES, especially given that Tpk1 and Tpk3 are proposed to suppress Msn2, whereas Tpk2 is a partial activator [86].
6. Control of the Heat Shock Response by the cAMP/PKA Pathway
Fluctuations in temperature represent one of the most fundamental stresses that are experienced by yeast cells. It is widely accepted that heat stress can cause proteins to misfold, lose their native structures or adopt non-native states that are prone to aggregation. Stress-induced protein misfolding and aggregation can be cytotoxic, although this toxicity can be mitigated by the activity of molecular chaperones including members of the heat shock protein family [92,93]. Newly synthesised polypeptides are particularly susceptible to misfolding and so can require extensive remodelling via chaperones to assume their native states [92].
At suboptimal temperatures, yeast cells activate a stress response pathway that is known as the heat shock response (HSR) [2]. This transcriptional programme is triggered in response to diverse stress conditions including heat, mistranslation, osmotic and oxidative stresses, and is required to transiently regulate the expression of proteostatic genes such as those encoding chaperones [2,94]. The HSR is orchestrated in eukaryotes by the evolutionarily conserved transcription factor, heat shock factor 1 (Hsf1) [95]. Hsf1 is essential in yeast, suggesting that it is also required during normal, non-stress conditions [96]. It has been shown to function downstream of the cAMP/PKA pathway, and like the ESR it is negatively regulated by PKA activity [97]. Not surprisingly therefore, differences are observed in the cAMP/PKA targeted phosphoproteome upon heat shock [98]. These phosphoproteomic changes include the rapid dephosphorylation of the PKA phosphorylation sites within the Msn2/4 nuclear localization signal (NLS), as well as concomitant changes in the phosphorylation of other PKA substrate proteins. The Yak1 and Rim15 protein kinases phosphorylate and activate Hsf1 activity and are themselves inhibited by PKA [99]. Yak1 and Rim15 also phosphorylate Msn2, suggesting an overlapping mechanism moderating the ESR and HSR under the control of the cAMP/PKA pathway. Again, Tpk isoforms are not redundant in their capacity to moderate Hsf1 activity, since Tpk1 and Tpk3 play predominant roles in repressing the HSR [100]. The negative regulation of Hsf1, Yak1 and Rim15 by PKA is consistent with PKA activity being downregulated in response to heat stress, resulting in the dephosphorylation of proteins such as Msn2/4 and thus promoting transcriptional changes that lead to adaptation.
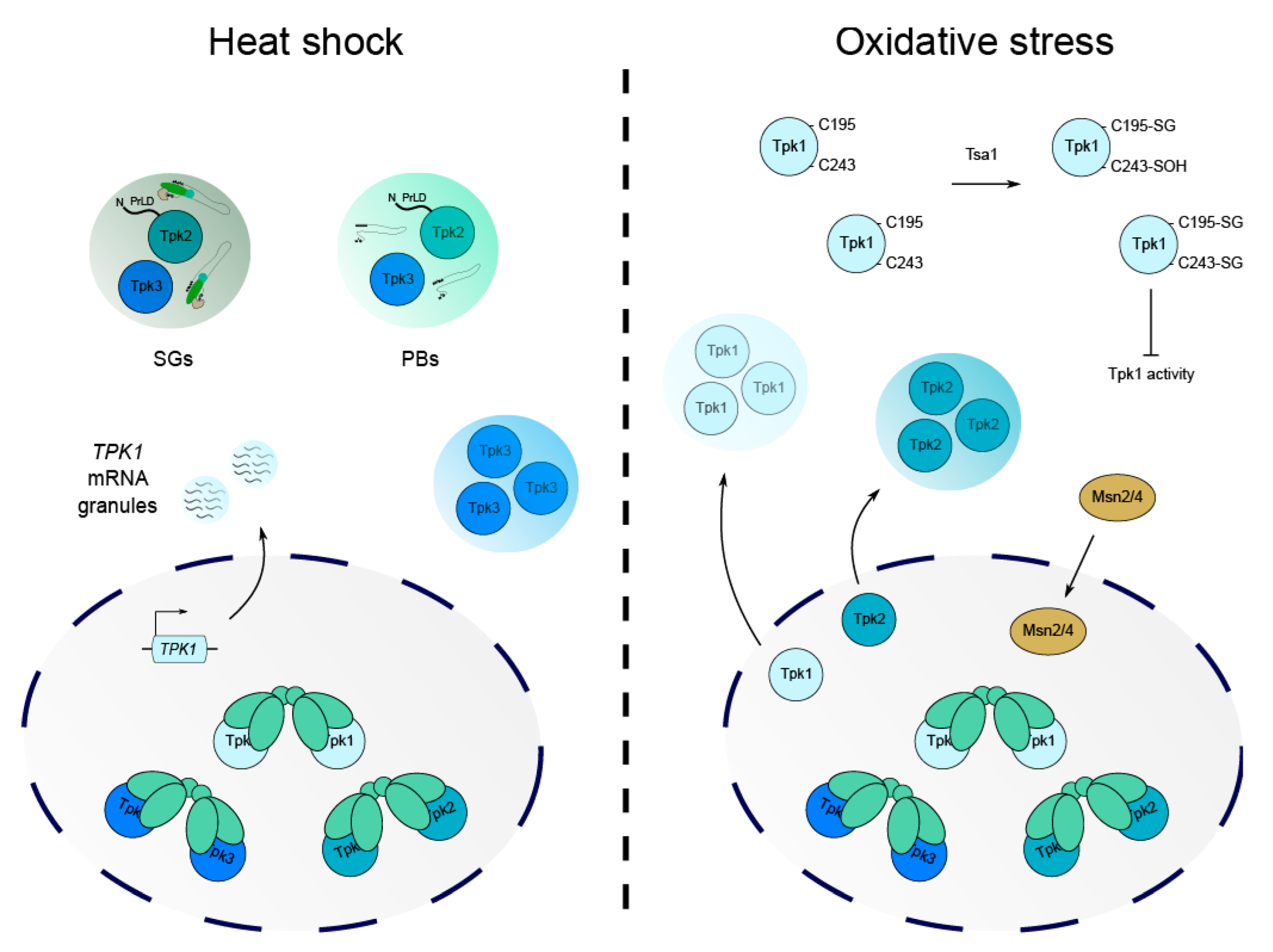
As high PKA activity is linked to lower stress tolerance, it seems plausible that regulation of PKA subunits at the transcriptional level might represent a mechanism to fine-tune PKA activity in response to stress. Some work has been carried out to study the regulation of TPK1-3 and BCY1 during heat shock and has identified that the TPK1 isoform is specifically upregulated, while TPK2 and TPK3 show no such increase in expression [101,102,103]. Curiously, a recent study of Tpk1 regulation during recurrent heat shock has shown that although TPK1 mRNA becomes more abundant, it is translationally inhibited and stored in mRNA granules until the stress is removed (Figure 4) [103]. During a recovery period, Tpk1 protein levels increase, suggesting that the transcriptional induction of TPK1 during the original stress serves as an adaptive measure to facilitate increased Tpk1-dependent phosphorylation upon recovery [103]. This emphasises an important role for the Tpk1 isoform and highlights how signalling specificity is required for stress adaptation such that cells are protected from future stressful events.
It is now well documented that heat shock causes translational inhibition and the formation of biomolecular condensates, termed mRNA processing bodies (PBs) and stress granules (SGs) [104,105,106]. These condensates represent specialised microenvironments containing mRNAs, components of the translational machinery and other proteins that are important for coordinating RNA fate. Temperature fluctuations have been found to cause isoform-specific alterations in Tpk catalytic subunit cellular localisation and their association with stress-induced condensates (Figure 4). For example, in response to severe heat stress, Tpk2 and Tpk3 proteins are targeted to PBs and SGs, while Tpk1 remains within the nucleus [107]. For Tpk2, the re-localisation is dependent on its kinase activity, whereas a catalytically inactive form of Tpk3 maintains its capacity to associate with granules [107]. Yeast strains that were deleted for TPK2 or TPK3 also exhibited differences in the expression of multiple mRNAs during heat stress, with Tpk2 seemingly responsible for the resumption of translation initiation after stress and Tpk3 promoting translational repression [107]. Further study has since revealed that a prion-like domain predicted in the N-terminus of Tpk2 is required for its re-localisation to PBs upon heat stress [49]. Tpk3 does not carry this N-terminal PrLD and so the mechanism by which this isoform associates with granules is clearly distinct to that of Tpk2 (Figure 4). This highlights how specific PKA structural domains are involved in condensate formation and how, as in mammalian systems, protein localisation likely drives targeted phosphorylation to modulate key processes such as translation and mRNA fate.
It is known that tolerance to other HSR-activating stress conditions which cause protein misfolding, such as mistranslation caused by the proline analogue L-azetidine-2-carboxylic acid (AZC), also require moderation of the cAMP/PKA pathway [108]. The downregulation of PKA activity in response to AZC stress is found to be dependent on the oxidation of the Bcy1 regulatory subunit that is mediated by the Tsa1 peroxiredoxin (Prx), which suppresses cAMP binding to Bcy1 and hence limits PKA activation [108]. It is interesting to note that despite the significant role of Tsa1 in the response to AZC stress, Bcy1 oxidation by Tsa1 was not observed in response to oxidant treatment [108]. This Tsa1 mechanism for preventing PKA activation was suggested as a global mechanism acting at the level of the holoenzyme; therefore, any Tpk isoform specificity in the response would appear to be unlikely.
7. Control of the Oxidative Stress Response by the cAMP/PKA Pathway
Crosstalk between oxidant dependent signalling and the PKA pathway is not limited to the effects of Tsa1 on Bcy1. The cAMP/PKA pathway has long been known to be required for the response to oxidative stress. Cells are continually exposed to ROS originating largely from mitochondrial metabolism. In excessive amounts, ROS can evade the coping defence systems that are evolved by cells, resulting in oxidative stress and damage to macromolecules, such as nucleic acids, proteins and lipids [2,109]. Part of the role of the cAMP/PKA pathway stems from its effects on the ESR via the Msn2/4 transcription factors. Additionally, the response to H2O2 stress is highly dependent on the Yap1 and Skn7 transcription factors, which mediate the major transcriptional response to oxidative stress in yeast [77,110]. Similar to the Msn2/4 mechanism, the Yap1/Skn7 transcription factors are negatively controlled via the cAMP/PKA pathway [111]. While work has been carried out on the relationship between general PKA signalling and hydrogen peroxide stress, little consideration has been given to the roles of individual Tpk isoforms in response to oxidative insults.
Distinct from changes in transcription, resistance to H2O2 stress has been attributed to the process of caloric restriction (CR). CR is a mechanism via which organisms from bacteria to mammals can prolong their lifespans, and in yeast, requires the downregulation of signalling pathways such as the cAMP/PKA pathway [112]. The downregulation of PKA in response to CR requires Tsa1, a key Prx with dual functionality as an antioxidant and molecular chaperone [113]. Tsa1 facilitates the reduction in harmful peroxide species but becomes hyperoxidised and retained in an inactive state when it is overwhelmed by ROS. This hyperoxidation is relieved by the activity of sulfiredoxin (Srx1) which reduces oxidised Cys residues in an ATP-dependent reaction [114]. In response to H2O2 or CR, low PKA activity stimulates the translational induction of Srx1, which increases oxidant tolerance and extends longevity [113]. As ROS production increases with age, the effects that are elicited by PKA downregulation are viewed as an intervention to mitigate oxidative stress.
Somewhat surprisingly, Msn2 activity can be regulated in response to blue light exposure in yeast in a mechanism that is mediated by hydrogen peroxide generated from a peroxisomal oxidase, and sensed by the Tsa1 peroxiredoxin [115]. Tsa1 was found to slow the rate at which Tpk isoforms exited the nucleus as a means of controlling Msn2 localisation (Figure 4). As mentioned previously, the exclusion of Msn2 from the nucleus reduces the expression of stress response genes and is associated with normal growth. Upon H2O2 treatment, Tpk1 and Tpk2 were found to rapidly exit the nucleus and form cytoplasmic foci, while Tpk3 and Bcy1 remained in the nucleus [115]. This indicates that Tsa1 only partially inhibits PKA, but that this is sufficient to allow nucleocytoplasmic oscillations of Msn2 (Figure 4). Moreover, it suggests that the Tsa1-mediated repression of PKA may act on Tpk1 and Tpk2 but not Tpk3, as Tpk3 localisation is unaffected by Tsa1 expression.
A more recent study investigating the relationship between PKA signalling and the response to hydrogen peroxide stress identified oxidative post-translational modifications (PTMs) in the Tpk1 catalytic subunit that require Tsa1 activity [116]. A model was proposed where the Tpk1 isoform was repressed via the sulfenylation of Cys243 and the glutathionylation of Cys195/243 when exposed to a bolus of hydrogen peroxide (Figure 4). Cys243 glutathionylation correlates with the increased dephosphorylation of Thr241 in the Tpk1 activation loop, which, when phosphorylated, prevents the autophosphorylation of Ser179 and PKA activation [116,117]. A reduction in Thr241 phosphorylation has been linked to a weakened Tpk1/Bcy1 interaction, which presumably increases the number of active Tpk1 subunits and thus PKA activity [117]. Upon H2O2 addition, Cys243 sulfenylation increases and there is an increase in the dephosphorylated state of Thr241, which is proposed to destabilise the activation loop and thus inhibit PKA activity. However, it is unknown how the interplay between these PTMs affects the stability of Tpk1-containing PKA complexes and whether these oxidative modifications exclusively inhibit Tpk1 subunits in a monomeric or Bcy1-bound state. Since Tpk1 Cys243 is conserved in Tpk2 and Tpk3 (Figure 2), there remains the strong possibility that Tpk2/3 may also be modified and regulated similarly to Tpk1 upon H2O2 stress, though this has not been explored. This is an interesting prospect, given that the redox modulation of cysteine residues under oxidative stress conditions appears to be evolutionary conserved in diverse eukaryotic protein kinases [118]. The presence of two additional cysteine residues in the N-terminal domain of Tpk3, which are not conserved in Tpk1/2, may also represent an additional means of regulating PKA during oxidising conditions (Figure 2).
8. Conclusions and Future Perspectives
In many eukaryotes, PKA activity is a composite of differentially regulated catalytic subunits and, although redundant for some functions, it is now clear that isoform specific activity is deployed for different biological processes. According to early data, yeast PKA activity was thought to be globally downregulated in response to a broad array of stress conditions to activate the environmental stress response [1,79,87,90,119]. However, more recent observations that are discussed in this review have challenged this concept and highlighted that PKA activity should no longer be considered as a single entity which is inhibited uniformly.
Studies in yeast and higher eukaryotes have shown that PKA is subject to several levels of regulation which modulate C subunit activity, including inhibition by the R subunit, phase separation, differential localisation and feedback control. As all PKA C subunits phosphorylate the RRXS*/T* consensus motif, it is likely that kinase-interacting proteins are key to directing substrate phosphorylation by anchoring the subunits to distinct subcellular sites. AKAPs are well characterised in mammalian cells; however, homologues in yeast are yet to be identified. CKAPs, which comprise all the proteins that interact with the C subunits, are therefore a likely determinant in targeting PKA isoforms to specific substrates. In response to stress, PKA has been shown to localise in an isoform specific manner to different cellular compartments, including stress-induced bodies such as PBs and SGs. It is likely that this re-localisation changes the targets of the Tpk isoforms, bringing them near to substrates that are important for stress tolerance. Stressors have also been shown to regulate PKA activity through post-translational modification of the C or R subunits, either directly inhibiting the Tpk subunits or affecting the stability of the holoenzyme complex. Alternatively, it is possible that phosphatases may regulate localised Tpk activity by compartmentalising PKA-dependent phosphorylation, as has been shown in mammalian cells [120,121]. Further careful study is therefore required to understand the heterogeneity of PKA signalling in terms of mapping isoform interaction networks and how the different holoenzyme subunits are regulated during changing cellular conditions. As the kinase activity of certain isoforms is required over others in response to distinct stresses, it will be interesting to dissect the mechanisms which modulate PKA isoforms in response to different conditions and how PTMs may contribute to this. Exciting new advances in the field, such as fluorescence resonance energy transfer (FRET) reporters to monitor cAMP and PKA activity, will also be powerful tools to study isoform specific signalling and its localisation in vivo [122]. A greater appreciation and understanding of the specific roles of kinase isoforms will hopefully facilitate studies to unravel the complexity of the kinome for the isoform specific branches of PKA signalling.
Author Contributions
Writing—original draft preparation, D.R.C.; writing—review and editing, D.R.C., S.J.H., M.P.A. and C.M.G. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by Studentship 2268014 from the UK Biotechnology and Biological Sciences Research Council (BBSRC) to D.R.C.
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Conflicts of Interest
The authors declare no conflict of interest. The funders had no role in the design of the study; in the collection, analyses, or interpretation of data; in the writing of the manuscript, or in the decision to publish the results.
References
- Gasch, A.P.; Spellman, P.T.; Kao, C.M.; Carmel-Harel, O.; Eisen, M.B.; Storz, G.; Botstein, D.; Brown, P.O. Genomic Expression Programs in the Response of Yeast Cells to Environmental Changes. Mol. Biol. Cell 2000, 11, 4241–4257. [Google Scholar] [CrossRef] [PubMed]
- Morano, K.A.; Grant, C.M.; Moye-Rowley, W.S. The Response to Heat Shock and Oxidative Stress in Saccharomyces cerevisiae. Genetics 2012, 190, 1157–1195. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Caza, M.; Kronstad, J.W. The cAMP/Protein Kinase A Pathway Regulates Virulence and Adaptation to Host Conditions in Cryptococcus neoformans. Front. Cell. Infect. Microbiol. 2019, 9, 212. [Google Scholar] [CrossRef]
- Portela, P.; Rossi, S. cAMP-PKA signal transduction specificity in Saccharomyces cerevisiae. Curr. Genet. 2020, 66, 1093–1099. [Google Scholar] [CrossRef]
- Taylor, S.S.; Wu, J.; Bruystens, J.G.; Del Rio, J.C.; Lu, T.-W.; Kornev, A.P.; Eyck, L.F.T. From structure to the dynamic regulation of a molecular switch: A journey over 3 decades. J. Biol. Chem. 2021, 296, 100746. [Google Scholar] [CrossRef]
- Taylor, S.S.; Buechler, J.A.; Yonemoto, W. Camp-dependent protein kinase: Framework fro a diverse family of regulatory enzymes. Annu. Rev. Biochem. 1990, 59, 971–1005. [Google Scholar] [CrossRef]
- Toda, T.; Cameron, S.; Sass, P.; Zöller, M.; Scott, J.D.; McMullen, B.; Hurwitz, M.; Krebs, E.G.; Wigler, M. Cloning and characterization of BCY1, a locus encoding a regulatory subunit of the cyclic AMP-dependent protein kinase in Saccharomyces cerevisiae. Mol. Cell. Biol. 1987, 7, 1371–1377. [Google Scholar] [CrossRef] [Green Version]
- Skalhegg, B.S.; Tasken, K. Specificity in the cAMP/PKA signaling pathway. differential expression, regulation, and subcellular localization of subunits of PKA. Front. Biosci. 2000, 5, D678–D693. [Google Scholar] [CrossRef] [Green Version]
- Søberg, K.; Skålhegg, B.S. The Molecular Basis for Specificity at the Level of the Protein Kinase a Catalytic Subunit. Front. Endocrinol. 2018, 9, 538. [Google Scholar] [CrossRef] [Green Version]
- Núñez, L.F.; Ocampo, J.; Gottlieb, A.M.; Rossi, S.; Moreno, S. Multiple isoforms for the catalytic subunit of PKA in the basal fungal lineage Mucor circinelloides. Fungal Biol. 2016, 120, 1493–1508. [Google Scholar] [CrossRef]
- Rinaldi, J.; Wu, J.; Yang, J.; Ralston, C.Y.; Sankaran, B.; Moreno, S.; Taylor, S.S. Structure of Yeast Regulatory Subunit: A Glimpse into the Evolution of PKA Signaling. Structure 2010, 18, 1471–1482. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Toda, T.; Cameron, S.; Sass, P.; Zoller, M.; Wigler, M. Three different genes in S. cerevisiae encode the catalytic subunits of the cAMP-dependent protein kinase. Cell 1987, 50, 277–287. [Google Scholar] [CrossRef]
- Gancedo, J.M. Control of pseudohyphae formation in Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2001, 25, 107–123. [Google Scholar] [CrossRef]
- Santangelo, G.M. Glucose Signaling in Saccharomyces cerevisiae. Microbiol. Mol. Biol. Rev. 2006, 70, 253–282. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thevelein, J.M.; de Winde, H. Novel sensing mechanisms and targets for the cAMP-protein kinase A pathway in the yeast Saccharomyces cerevisiae. Mol. Microbiol. 1999, 33, 904–918. [Google Scholar] [CrossRef]
- Conrad, M.; Schothorst, J.; Kankipati, H.N.; Van Zeebroeck, G.; Rubio-Texeira, M.; Thevelein, J.M. Nutrient sensing and signaling in the yeast Saccharomyces cerevisiae. FEMS Microbiol. Rev. 2014, 38, 254–299. [Google Scholar] [CrossRef] [Green Version]
- Colombo, S.; Ma, P.; Cauwenberg, L.; Winderickx, J.; Crauwels, M.; Teunissen, A.; Nauwelaers, D.; De Winde, J.H.; Gorwa, M.; Colavizza, D.; et al. Involvement of distinct G-proteins, Gpa2 and Ras, in glucose- and intracellular acidification-induced cAMP signalling in the yeast Saccharomyces cerevisiae. EMBO J. 1998, 17, 3326–3341. [Google Scholar] [CrossRef] [Green Version]
- Nakafuku, M.; Obara, T.; Kaibuchi, K.; Miyajima, I.; Itoh, H.; Nakamura, S.; Arai, K.; Matsumoto, K.; Kaziro, Y. Isolation of a second yeast Saccharomyces cerevisiae gene (GPA2) coding for guanine nucleotide-binding regulatory protein: Studies on its structure and possible functions. Proc. Natl. Acad. Sci. USA 1988, 85, 1374–1378. [Google Scholar] [CrossRef] [Green Version]
- Rolland, F.; de Winde, H.; Lemaire, K.; Boles, E.; Thevelein, J.; Winderickx, J. Glucose-induced cAMP signalling in yeast requires both a G-protein coupled receptor system for extracellular glucose detection and a separable hexose kinase-dependent sensing process. Mol. Microbiol. 2000, 38, 348–358. [Google Scholar] [CrossRef]
- Xue, Y.; Batlle, M.; Hirsch, J.P. GPR1 encodes a putative G protein-coupled receptor that associates with the Gpa2p Gα subunit and functions in a Ras-independent pathway. EMBO J. 1998, 17, 1996–2007. [Google Scholar] [CrossRef]
- Dautt-Castro, M.; Rosendo-Vargas, M.; Casas-Flores, S. The Small GTPases in Fungal Signaling Conservation and Function. Cells 2021, 10, 1039. [Google Scholar] [CrossRef] [PubMed]
- Kataoka, T.; Powers, S.; Cameron, S.; Fasano, O.; Goldfarb, M.; Broach, J.; Wigler, M. Functional homology of mammalian and yeast RAS genes. Cell 1985, 40, 19–26. [Google Scholar] [CrossRef]
- Tamanoi, F. Ras Signaling in Yeast. Genes Cancer 2011, 2, 210–215. [Google Scholar] [CrossRef]
- Gancedo, J.M. The early steps of glucose signalling in yeast. FEMS Microbiol. Rev. 2008, 32, 673–704. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broek, D.; Toda, T.; Michaeli, T.; Levin, L.; Birchmeier, C.; Zoller, M.; Powers, S.; Wigler, M. The S. cerevisiae CDC25 gene product regulates the RAS/adenylate cyclase pathway. Cell 1987, 48, 789–799. [Google Scholar] [CrossRef]
- Belotti, F.; Tisi, R.; Paiardi, C.; Groppi, S.; Martegani, E. PKA-dependent regulation of Cdc25 RasGEF localization in budding yeast. FEBS Lett. 2011, 585, 3914–3920. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Broggi, S.; Martegani, E.; Colombo, S. Live-cell imaging of endogenous Ras-GTP shows predominant Ras activation at the plasma membrane and in the nucleus in Saccharomyces cerevisiae. Int. J. Biochem. Cell Biol. 2013, 45, 384–394. [Google Scholar] [CrossRef]
- Tanaka, K.; Nakafuku, M.; Satoh, T.; Marshall, M.S.; Gibbs, J.B.; Matsumoto, K.; Kaziro, Y.; Toh-E, A. S. cerevisiae genes IRA1 and IRA2 encode proteins that may be functionally equivalent to mammalian ras GTPase activating protein. Cell 1990, 60, 803–807. [Google Scholar] [CrossRef]
- Tanaka, K.; Nakafuku, M.; Tamanoi, F.; Kaziro, Y.; Matsumoto, K.; Toh-e, A. IRA2, a second gene of Saccharomyces cerevisiae that encodes a protein with a domain homologous to mammalian ras GTPase-activating protein. Mol. Cell Biol. 1990, 10, 4303–4313. [Google Scholar]
- Park, J.-I.; Grant, C.M.; Dawes, I.W. The high-affinity cAMP phosphodiesterase of Saccharomyces cerevisiae is the major determinant of cAMP levels in stationary phase: Involvement of different branches of the Ras–cyclic AMP pathway in stress responses. Biochem. Biophys. Res. Commun. 2005, 327, 311–319. [Google Scholar] [CrossRef]
- Smets, B.; Ghillebert, R.; De Snijder, P.; Binda, M.; Swinnen, E.; De Virgilio, C.; Winderickx, J. Life in the midst of scarcity: Adaptations to nutrient availability in Saccharomyces cerevisiae. Curr. Genet. 2010, 56, 1–32. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ma, P.; Wera, S.; van Dijck, P.; Thevelein, J.M. The PDE1-encoded Low-Affinity Phosphodiesterase in the Yeast Saccha-romyces cerevisiae Has a Specific Function in Controlling Agonist-induced cAMP Signaling. Mol. Biol. Cell 1999, 10, 91–104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- McCormick, K.; Baillie, G.S. Compartmentalisation of second messenger signalling pathways. Curr. Opin. Genet. Dev. 2014, 27, 20–25. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.; Liu, E.; Bai, X.; Zhang, A. The localization and concentration of the PDE2-encoded high-affinity cAMP phosphodiesterase is regulated by cAMP-dependent protein kinase A in the yeast Saccharomyces cerevisiae. FEMS Yeast Res. 2010, 10, 177–187. [Google Scholar] [CrossRef] [Green Version]
- Smith, F.D.; Esseltine, J.L.; Nygren, P.J.; Veesler, D.; Byrne, D.P.; Vonderach, M.; Strashnov, I.; Eyers, C.E.; Eyers, P.A.; Langeberg, L.K.; et al. Local protein kinase A action proceeds through intact holoenzymes. Science 2017, 356, 1288–1293. [Google Scholar] [CrossRef] [Green Version]
- Smith, F.D.; Scott, J.D. Protein kinase A activation: Something new under the sun? J. Cell Biol. 2018, 217, 1895–1897. [Google Scholar] [CrossRef] [Green Version]
- Vigil, D.; Blumenthal, D.K.; Brown, S.; Taylor, A.S.S.; Trewhella, J. Differential Effects of Substrate on Type I and Type II PKA Holoenzyme Dissociation. Biochemistry 2004, 43, 5629–5636. [Google Scholar] [CrossRef]
- Walker-Gray, R.; Stengel, F.; Gold, M.G. Mechanisms for restraining cAMP-dependent protein kinase revealed by subunit quantitation and cross-linking approaches. Proc. Natl. Acad. Sci. USA 2017, 114, 10414–10419. [Google Scholar] [CrossRef] [Green Version]
- Galello, F.; Portela, P.; Moreno, S.; Rossi, S. Characterization of Substrates That Have a Differential Effect on Saccharomyces cerevisiae Protein Kinase A Holoenzyme Activation*. J. Biol. Chem. 2010, 285, 29770–29779. [Google Scholar] [CrossRef] [Green Version]
- Ptacek, J.; Devgan, G.; Michaud, G.; Zhu, H.; Zhu, X.; Fasolo, J.; Guo, H.; Jona, G.; Breitkreutz, A.; Sopko, R.; et al. Global analysis of protein phosphorylation in yeast. Nature 2005, 438, 679–684. [Google Scholar] [CrossRef]
- Dahlin, H.R.; Zheng, N.; Scott, J.D. Beyond PKA: Evolutionary and structural insights that define a docking and dimerization domain superfamily. J. Biol. Chem. 2021, 297, 100927. [Google Scholar] [CrossRef] [PubMed]
- Bardeci, N.G.; Caramelo, J.J.; Blumenthal, D.K.; Rinaldi, J.; Rossi, S.; Moreno, S. The PKA regulatory subunit from yeast forms a homotetramer: Low-resolution structure of the N-terminal oligomerization domain. J. Struct. Biol. 2015, 193, 141–154. [Google Scholar] [CrossRef] [PubMed]
- Galello, F.; Moreno, S.; Rossi, S. Interacting proteins of protein kinase A regulatory subunit in Saccharomyces cerevisiae. J. Proteom. 2014, 109, 261–275. [Google Scholar] [CrossRef] [PubMed]
- Griffioen, G.; Branduardi, P.; Ballarini, A.; Anghileri, P.; Norbeck, J.; Baroni, M.D.; Ruis, H. Nucleocytoplasmic Distribution of Budding Yeast Protein Kinase A Regulatory Subunit Bcy1 Requires Zds1 and Is Regulated by Yak1-Dependent Phosphorylation of Its Targeting Domain. Mol. Cell. Biol. 2001, 21, 511–523. [Google Scholar] [CrossRef] [Green Version]
- Haesendonckx, S.; Tudisca, V.; Voordeckers, K.; Moreno, S.; Thevelein, J.M.; Portela, P. The activation loop of PKA catalytic isoforms is differentially phosphorylated by Pkh protein kinases in Saccharomyces cerevisiae. Biochem. J. 2012, 448, 307–320. [Google Scholar] [CrossRef] [Green Version]
- Robertson, L.S.; Fink, G.R. The three yeast A kinases have specific signaling functions in pseudohyphal growth. Proc. Natl. Acad. Sci. USA 1998, 95, 13783–13787. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, R.; A Huynen, M.; Snel, B. Complex fate of paralogs. BMC Evol. Biol. 2008, 8, 337. [Google Scholar] [CrossRef] [Green Version]
- Mashhoon, N.; Carmel, G.; Pflugrath, J.W.; Kuret, J. Structure of the Unliganded cAMP-Dependent Protein Kinase Catalytic Subunit from Saccharomyces cerevisiae. Arch. Biochem. Biophys. 2001, 387, 11–19. [Google Scholar] [CrossRef]
- Barraza, C.E.; Solari, C.A.; Rinaldi, J.; Ojeda, L.; Rossi, S.; Ashe, M.P.; Portela, P. A prion-like domain of Tpk2 catalytic subunit of protein kinase A modulates P-body formation in response to stress in budding yeast. Biochim. Biophys. Acta 2020, 1868, 118884. [Google Scholar] [CrossRef]
- Robertson, L.S.; Causton, H.C.; Young, R.A.; Fink, G.R. The yeast A kinases differentially regulate iron uptake and respiratory function. Proc. Natl. Acad. Sci. USA 2000, 97, 5984–5988. [Google Scholar] [CrossRef] [Green Version]
- Chevtzoff, C.; Vallortigara, J.; Avéret, N.; Rigoulet, M.; Devin, A. The yeast cAMP protein kinase Tpk3p is involved in the regulation of mitochondrial enzymatic content during growth. Biochim. Biophys. Acta 2005, 1706, 117–125. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- E Leadsham, J.; Gourlay, C.W. cAMP/PKA signaling balances respiratory activity with mitochondria dependent apoptosis via transcriptional regulation. BMC Cell Biol. 2010, 11, 92. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raghuram, V.; Salhadar, K.; Limbutara, K.; Park, E.; Yang, C.-R.; Knepper, M.A. Protein kinase A catalytic-α and catalytic-β proteins have nonredundant regulatory functions. Am. J. Physiol. Physiol. 2020, 319, F848–F862. [Google Scholar] [CrossRef]
- Griffioen, G.; Anghileri, P.; Imre, E.; Baroni, M.D.; Ruis, H. Nutritional Control of Nucleocytoplasmic Localization of cAMP-dependent Protein Kinase Catalytic and Regulatory Subunits in Saccharomyces cerevisiae. J. Biol. Chem. 2000, 275, 1449–1456. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tudisca, V.; Recouvreux, V.; Moreno, S.; Boy-Marcotte, E.; Jacquet, M.; Portela, P. Differential localization to cytoplasm, nucleus or P-bodies of yeast PKA subunits under different growth conditions. Eur. J. Cell Biol. 2010, 89, 339–348. [Google Scholar] [CrossRef]
- Papadopoulos, J.S.; Agarwala, R. COBALT: Constraint-based alignment tool for multiple protein sequences. Bioinformatics 2007, 23, 1073–1079. [Google Scholar] [CrossRef] [Green Version]
- Mösch, H.U.; Roberts, R.L.; Fink, G.R. Ras2 signals via the Cdc42/Ste20/mitogen-activated protein kinase module to induce filamentous growth in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1996, 93, 5352–5356. [Google Scholar] [CrossRef] [Green Version]
- Gimeno, C.J.; Ljungdahl, P.O.; Styles, C.A.; Fink, G.R. Unipolar cell divisions in the yeast S. cerevisiae lead to filamentous growth: Regulation by starvation and RAS. Cell 1992, 68, 1077–1090. [Google Scholar] [CrossRef]
- Ward, M.P.; Gimeno, C.J.; Fink, G.R.; Garrett, S. SOK2 may regulate cyclic AMP-dependent protein kinase-stimulated growth and pseudohyphal development by repressing transcription. Mol. Cell. Biol. 1995, 15, 6854–6863. [Google Scholar] [CrossRef] [Green Version]
- Cullen, P.J.; Sprague, G.F. The Regulation of Filamentous Growth in Yeast. Genetics 2012, 190, 23–49. [Google Scholar] [CrossRef]
- Pan, X.; Heitman, J. Cyclic AMP-Dependent Protein Kinase Regulates Pseudohyphal Differentiation in Saccharomyces cerevisiae. Mol. Cell. Biol. 1999, 19, 4874–4887. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayikci, Ö.; Magwene, P.M. Divergent Roles for cAMP–PKA Signaling in the Regulation of Filamentous Growth in Saccharomyces cerevisiae and Saccharomyces bayanus. G3 Genes Genomes Genet. 2018, 8, 3529–3538. [Google Scholar] [CrossRef] [Green Version]
- Rupp, S.; Summers, E.; Lo, H.; Madhani, H.; Fink, G. MAP kinase and cAMP filamentation signaling pathways converge on the unusually large promoter of the yeast FLO11 gene. EMBO J. 1999, 18, 1257–1269. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, H.; A Styles, C.; Fink, G.R. Saccharomyces cerevisiae S288C has a mutation in FLO8, a gene required for filamentous growth. Genetics 1996, 144, 967–978. [Google Scholar] [CrossRef] [PubMed]
- Deminoff, S.J.; Howard, S.C.; Hester, A.; Warner, S.; Herman, P.K. Using Substrate-Binding Variants of the cAMP-Dependent Protein Kinase to Identify Novel Targets and a Kinase Domain Important for Substrate Interactions in Saccharomyces cerevisiae. Genetics 2006, 173, 1909–1917. [Google Scholar] [CrossRef] [Green Version]
- Malcher, M.; Schladebeck, S.; Mösch, H.-U. The Yak1 Protein Kinase Lies at the Center of a Regulatory Cascade Affecting Adhesive Growth and Stress Resistance in Saccharomyces cerevisiae. Genetics 2011, 187, 717–730. [Google Scholar] [CrossRef] [Green Version]
- Kispal, G.; Steiner, H.; Court, D.; Rolinski, B.; Lill, R. Mitochondrial and Cytosolic Branched-chain Amino Acid Transaminases from Yeast, Homologs of the myc Oncogene-regulated Eca39 Protein. J. Biol. Chem. 1996, 271, 24458–24464. [Google Scholar] [CrossRef] [Green Version]
- Zelenaya-Troitskaya, O.; Perlman, P.S.; Butow, R.A. An enzyme in yeast mitochondria that catalyzes a step in branched-chain amino acid biosynthesis also functions in mitochondrial DNA stability. EMBO J. 1995, 14, 3268–3276. [Google Scholar] [CrossRef]
- Schothorst, J.; Van Zeebroeck, G.; Thevelein, J.M. Identification of Ftr1 and Zrt1 as iron and zinc micronutrient transceptors for activation of the PKA pathway in Saccharomyces cerevisiae. Microb. Cell 2017, 4, 74–89. [Google Scholar] [CrossRef] [Green Version]
- Mazon, M.J.; Behrens, M.M.; Morgado, E.; Portillo, F. Low activity of the yeast cAMP-dependent protein kinase catalytic subunit Tpk3 is due to the poor expression of the TPK3 gene. JBIC J. Biol. Inorg. Chem. 1993, 213, 501–506. [Google Scholar] [CrossRef]
- Chevtzoff, C.; Yoboue, E.D.; Galinier, A.; Casteilla, L.; Daignan-Fornier, B.; Rigoulet, M.; Devin, A. Reactive Oxygen Species-mediated Regulation of Mitochondrial Biogenesis in the Yeast Saccharomyces cerevisiae. J. Biol. Chem. 2010, 285, 1733–1742. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rahman, M.; Hudson, A. Substrates for Yeast Mitochondrial cAMP-Dependent Protein Kinase Activity. Biochem. Biophys. Res. Commun. 1995, 214, 188–194. [Google Scholar] [CrossRef] [PubMed]
- Kadenbach, B.; Hüttemann, M.; Arnold, S.; Lee, I.; Bender, E. Mitochondrial energy metabolism is regulated via nuclear-coded subunits of cytochrome c oxidase. Free Radic. Biol. Med. 2000, 29, 211–221. [Google Scholar] [CrossRef]
- Boy-Marcotte, E.; Perrot, M.; Bussereau, F.; Boucherie, H.; Jacquet, M. Msn2p and Msn4p Control a Large Number of Genes Induced at the Diauxic Transition Which Are Repressed by Cyclic AMP in Saccharomyces cerevisiae. J. Bacteriol. 1998, 180, 1044–1052. [Google Scholar] [CrossRef] [Green Version]
- Erkina, T.Y.; Lavrova, M.V.; Erkine, A.M. Alternative ways of stress regulation in Saccharomyces cerevisiae cells: Transcriptional activators Msn2 and Msn. Cell Tissue Biol. 2009, 3, 121–129. [Google Scholar] [CrossRef]
- Görner, W.; Durchschlag, E.; Martinez-Pastor, M.T.; Estruch, F.; Ammerer, G.; Hamilton, B.; Ruis, H.; Schüller, C. Nuclear localization of the C2H2 zinc finger protein Msn2p is regulated by stress and protein kinase A activity. Genes Dev. 1998, 12, 586–597. [Google Scholar] [CrossRef]
- Hasan, R.; Leroy, C.; Isnard, A.-D.; Labarre, J.; Boy-Marcotte, E.; Toledano, M.B. The control of the yeast H2O2 response by the Msn2/4 transcription factors. Mol. Microbiol. 2002, 45, 233–241. [Google Scholar] [CrossRef]
- Kandror, O.; Bretschneider, N.; Kreydin, E.; Cavalieri, D.; Goldberg, A.L. Yeast Adapt to Near-Freezing Temperatures by STRE/Msn2,4-Dependent Induction of Trehalose Synthesis and Certain Molecular Chaperones. Mol. Cell 2004, 13, 771–781. [Google Scholar] [CrossRef]
- Garreau, H.; Hasan, R.N.; Renault, G.; Estruch, F.; Boy-Marcotte, E.; Jacquet, M. Hyperphosphorylation of Msn2p and Msn4p in response to heat shock and the diauxic shift is inhibited by cAMP in Saccharomyces cerevisiae. Microbiology 2000, 146, 2113–2120. [Google Scholar] [CrossRef] [Green Version]
- Smith, A.; Ward, M.P.; Garrett, S. Yeast PKA represses Msn2p/Msn4p-dependent gene expression to regulate growth, stress response and glycogen accumulation. EMBO J. 1998, 17, 3556–3564. [Google Scholar] [CrossRef] [Green Version]
- Chasman, D.; Ho, Y.; Berry, D.B.; Nemec, C.M.; E MacGilvray, M.; Hose, J.; E Merrill, A.; Lee, M.V.; Will, J.L.; Coon, J.J.; et al. Pathway connectivity and signaling coordination in the yeast stress-activated signaling network. Mol. Syst. Biol. 2014, 10, 759. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Pastor, M.T.; Marchler, G.; Schüller, C.; Marchler-Bauer, A.; Ruis, H.; Estruch, F. The Saccharomyces cerevisiae zinc finger proteins Msn2p and Msn4p are required for transcriptional induction through the stress response element (STRE). EMBO J 1996, 15, 2227–2235. [Google Scholar] [CrossRef] [PubMed]
- Kobayashi, N.; McEntee, K. Evidence for a heat shock transcription factor-independent mechanism for heat shock induction of transcription in Saccharomyces cerevisiae. Proc. Natl. Acad. Sci. USA 1990, 87, 6550–6554. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wieser, R.; Adam, G.; Wagner, A.; Schüller, C.; Marchler, G.; Ruis, H.; Krawiec, Z.; Bilinski, T. Heat shock factor-independent heat control of transcription of the CTT1 gene encoding the cytosolic catalase T of Saccharomyces cerevisiae. J. Biol. Chem. 1991, 266, 12406–12411. [Google Scholar] [CrossRef]
- Causton, H.C.; Ren, B.; Koh, S.S.; Harbison, C.T.; Kanin, E.; Jennings, E.G.; Lee, T.I.; True, H.L.; Lander, E.S.; Young, R.A. Remodeling of Yeast Genome Expression in Response to Environmental Changes. Mol. Biol. Cell 2001, 12, 323–337. [Google Scholar] [CrossRef] [Green Version]
- Sadeh, A.; Movshovich, N.; Volokh, M.; Gheber, L.; Aharoni, A. Fine-tuning of the Msn2/4–mediated yeast stress responses as revealed by systematic deletion of Msn2/4 partners. Mol. Biol. Cell 2011, 22, 3127–3138. [Google Scholar] [CrossRef]
- Durchschlag, E.; Reiter, W.; Ammerer, G.; Schüller, C. Nuclear Localization Destabilizes the Stress-regulated Transcription Factor Msn. J. Biol. Chem. 2004, 279, 55425–55432. [Google Scholar] [CrossRef] [Green Version]
- Jacquet, M.; Renault, G.; Lallet, S.; De Mey, J.; Goldbeter, A. Oscillatory nucleocytoplasmic shuttling of the general stress response transcriptional activators Msn2 and Msn4 in Saccharomyces cerevisiae. J. Cell Biol. 2003, 161, 497–505. [Google Scholar] [CrossRef]
- Görner, W.; Durchschlag, E.; Wolf, J.; Brown, E.L.; Ammerer, G.; Ruis, H.; Schüller, C. Acute glucose starvation activates the nuclear localization signal of a stress-specific yeast transcription factor. EMBO J. 2002, 21, 135–144. [Google Scholar] [CrossRef] [Green Version]
- Garmendia-Torres, C.; Goldbeter, A.; Jacquet, M. Nucleocytoplasmic Oscillations of the Yeast Transcription Factor Msn2: Evidence for Periodic PKA Activation. Curr. Biol. 2007, 17, 1044–1049. [Google Scholar] [CrossRef]
- Pfanzagl, V.; Görner, W.; Radolf, M.; Parich, A.; Schuhmacher, R.; Strauss, J.; Reiter, W.; Schüller, C. A constitutive active allele of the transcription factor Msn2 mimicking low PKA activity dictates metabolic remodeling in yeast. Mol. Biol. Cell 2018, 29, 2848–2862. [Google Scholar] [CrossRef] [PubMed]
- Vabulas, R.M.; Raychaudhuri, S.; Hayer-Hartl, M.; Hartl, F.U. Protein Folding in the Cytoplasm and the Heat Shock Response. Cold Spring Harb. Perspect. Biol. 2010, 2, a004390. [Google Scholar] [CrossRef] [PubMed]
- Verghese, J.; Abrams, J.; Wang, Y.; Morano, K.A. Biology of the Heat Shock Response and Protein Chaperones: Budding Yeast (Saccharomyces cerevisiae) as a Model System. Microbiol. Mol. Biol. Rev. 2012, 76, 115–158. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Trotter, E.W.; Kao, C.M.-F.; Berenfeld, L.; Botstein, D.; Petsko, G.A.; Gray, J.V. Misfolded Proteins Are Competent to Mediate a Subset of the Responses to Heat Shock in Saccharomyces cerevisiae. J. Biol. Chem. 2002, 277, 44817–44825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kmiecik, S.W.; Mayer, M.P. Molecular mechanisms of heat shock factor 1 regulation. Trends Biochem. Sci. 2021, 47, 218–234. [Google Scholar] [CrossRef]
- Sorger, P.K.; Pelham, H.R. Yeast heat shock factor is an essential DNA-binding protein that exhibits temperature-dependent phosphorylation. Cell 1988, 54, 855–864. [Google Scholar] [CrossRef]
- Ferguson, S.B.; Anderson, E.S.; Harshaw, R.B.; Thate, T.; Craig, N.L.; Nelson, H.C.M. Protein Kinase A Regulates Constitutive Expression of Small Heat-Shock Genes in an Msn2/4p-Independent and Hsf1p-Dependent Manner in Saccharomyces cerevisiae. Genetics 2005, 169, 1203–1214. [Google Scholar] [CrossRef] [Green Version]
- Kanshin, E.; Kubiniok, P.; Thattikota, Y.; D’Amours, D.; Thibault, P. Phosphoproteome dynamics of Saccharomyces cerevisiae under heat shock and cold stress. Mol. Syst. Biol. 2015, 11, 813. [Google Scholar] [CrossRef]
- Lee, P.; Cho, B.-R.; Joo, H.-S.; Hahn, J.-S. Yeast Yak1 kinase, a bridge between PKA and stress-responsive transcription factors, Hsf1 and Msn2/Msn. Mol. Microbiol. 2008, 70, 882–895. [Google Scholar] [CrossRef]
- Pérez-Landero, S.; Sandoval-Motta, S.; Martínez-Anaya, C.; Yang, R.; Folch-Mallol, J.L.; Martínez, L.M.; Ventura, L.; Guillén-Navarro, K.; Aldana-González, M.; Nieto-Sotelo, J. Complex Regulation of Hsf1-Skn7 Activities by the Catalytic Subunits of PKA in Saccharomyces Cerevisiae: Experimental and Computational Evidences. BMC Syst. Biol. 2015, 9, 42. [Google Scholar] [CrossRef] [Green Version]
- Pautasso, C.; Rossi, S. Transcriptional regulation of the protein kinase A subunits in Saccharomyces cerevisiae: Autoregulatory role of the kinase A activity. Biochim. Biophys. Acta 2014, 1839, 275–287. [Google Scholar] [CrossRef]
- Reca, S.; Galello, F.; Ojeda, L.; Pautasso, C.; Cañonero, L.; Moreno, S.; Portela, P.; Rossi, S. Chromatin remodeling and transcription of the TPK1 subunit of PKA during stress in Saccharomyces cerevisiae. Biochim. Biophys. Acta 2020, 1863, 194599. [Google Scholar] [CrossRef] [PubMed]
- Cañonero, L.; Pautasso, C.; Galello, F.; Sigaut, L.; Pietrasanta, L.; Arroyo, J.; Bermúdez-Moretti, M.; Portela, P.; Rossi, S. Heat stress regulates the expression of TPK1 gene at transcriptional and post-transcriptional levels in Saccharomyces cerevisiae. Biochim. Biophys. Acta 2022, 1869, 119209. [Google Scholar] [CrossRef] [PubMed]
- Balagopal, V.; Parker, R. Polysomes, P bodies and stress granules: States and fates of eukaryotic mRNAs. Curr. Opin. Cell Biol. 2009, 21, 403–408. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Protter, D.S.; Parker, R. Principles and Properties of Stress Granules. Trends Cell Biol. 2016, 26, 668–679. [Google Scholar] [CrossRef] [Green Version]
- Wallace, E.W.; Kear-Scott, J.L.; Pilipenko, E.V.; Schwartz, M.H.; Laskowski, P.R.; Rojek, A.E.; Katanski, C.D.; Riback, J.A.; Dion, M.F.; Franks, A.M.; et al. Reversible, Specific, Active Aggregates of Endogenous Proteins Assemble upon Heat Stress. Cell 2015, 162, 1286–1298. [Google Scholar] [CrossRef] [Green Version]
- Barraza, C.E.; Solari, C.A.; Marcovich, I.; Kershaw, C.; Galello, F.; Rossi, S.; Ashe, M.P.; Portela, P. The role of PKA in the translational response to heat stress in Saccharomyces cerevisiae. PLoS ONE 2017, 12, e0185416. [Google Scholar] [CrossRef]
- Kritsiligkou, P.; Nowicki-Osuch, K.; Carter, Z.; Kershaw, C.J.; Creamer, D.R.; Weids, A.J.; Grant, C.M. Tolerance to nascent protein misfolding stress requires fine-tuning of the cAMP/PKA pathway. J. Biol. Chem. 2021, 296, 100690. [Google Scholar] [CrossRef]
- Rodrigues-Pousada, C.; Nevitt, T.; Menezes, R. The yeast stress response. FEBS J. 2005, 272, 2639–2647. [Google Scholar] [CrossRef]
- Godon, C.; Lagniel, G.; Lee, J.; Buhler, J.-M.; Kieffer, S.; Perrot, M.; Boucherie, H.; Toledano, M.B.; Labarre, J. The H2O2 Stimulon in Saccharomyces cerevisiae. J. Biol. Chem. 1998, 273, 22480–22489. [Google Scholar] [CrossRef] [Green Version]
- Charizanis, C.; Juhnke, H.; Krems, B.; Entian, K.-D. The oxidative stress response mediated via Pos9/Skn7 is negatively regulated by the Ras/PKA pathway in Saccharomyces cerevisiae. Mol. Gen. Genet. MGG 1999, 261, 740–752. [Google Scholar] [CrossRef] [PubMed]
- Kaeberlein, M. Lessons on longevity from budding yeast. Nature 2010, 464, 513–519. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Molin, M.; Yang, J.; Hanzén, S.; Toledano, M.B.; Labarre, J.; Nyström, T. Life Span Extension and H2O2 Resistance Elicited by Caloric Restriction Require the Peroxiredoxin Tsa1 in Saccharomyces cerevisiae. Mol. Cell 2011, 43, 823–833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Biteau, B.; Labarre, J.; Toledano, M.B. ATP-dependent reduction of cysteine–sulphinic acid by S. cerevisiae sulphiredoxin. Nature 2003, 425, 980–984. [Google Scholar] [CrossRef]
- Bodvard, K.; Peeters, K.; Roger, F.; Romanov, N.; Igbaria, A.; Welkenhuysen, N.; Palais, G.; Reiter, W.; Toledano, M.B.; Käll, M.; et al. Light-sensing via hydrogen peroxide and a peroxiredoxin. Nat. Commun. 2017, 8, 14791. [Google Scholar] [CrossRef] [Green Version]
- Roger, F.; Picazo, C.; Reiter, W.; Libiad, M.; Asami, C.; Hanzén, S.; Gao, C.; Lagniel, G.; Wel-kenhuysen, N.; Labarre, J.; et al. Peroxiredoxin Promotes Longevity and H2O2-Resistance in Yeast through Redox-Modulation of Protein Kinase A. eLife 2020, 9, e60346. [Google Scholar] [CrossRef]
- Solari, C.A.; Tudisca, V.; Pugliessi, M.; Nadra, A.D.; Moreno, S.; Portela, P. Regulation of PKA activity by an autophosphorylation mechanism in Saccharomyces cerevisiae. Biochem. J. 2014, 462, 567–579. [Google Scholar] [CrossRef]
- Byrne, D.P.; Shrestha, S.; Galler, M.; Cao, M.; Daly, L.A.; Campbell, A.E.; Eyers, C.E.; Veal, E.A.; Kannan, N.; Eyers, P.A. Aurora A regulation by reversible cysteine oxidation reveals evolutionarily conserved redox control of Ser/Thr protein kinase activity. Sci. Signal. 2020, 13. [Google Scholar] [CrossRef]
- Gasch, A.P. The environmental stress response: A common yeast response to diverse environmental stresses. In Yeast Stress Responses; Hohmann, S., Mager, W.H., Eds.; Springer: Berlin/Heidelberg, Germany, 2003; pp. 11–70. [Google Scholar] [CrossRef]
- Grisan, F.; Iannucci, L.F.; Surdo, N.C.; Gerbino, A.; Zanin, S.; Di Benedetto, G.; Pozzan, T.; Lefkimmiatis, K. PKA compartmentalization links cAMP signaling and autophagy. Cell Death Differ. 2021, 28, 2436–2449. [Google Scholar] [CrossRef]
- Feliciello, A.; Gottesman, M.E.; Avvedimento, E.V. cAMP-PKA signaling to the mitochondria: Protein scaffolds, mRNA and phosphatases. Cell. Signal. 2005, 17, 279–287. [Google Scholar] [CrossRef]
- Colombo, S.; Longoni, E.; Gnugnoli, M.; Busti, S.; Martegani, E. Fast detection of PKA activity in Saccharomyces cerevisiae cell population using AKAR fluorescence resonance energy transfer probes. Cell. Signal. 2022, 92, 110262. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Overview of the cAMP/PKA pathway in S. cerevisiae. Pathway activation occurs in response to glucose via the GPCR (Gpr1) and Ras-based systems. These systems collectively stimulate the adenylate cyclase Cyr1 which increases production of cAMP from ATP. PKA is the only known cAMP receptor in yeast, and upon binding the PKA regulatory subunit, cAMP promotes allosteric changes, which liberate the catalytic subunits from the PKA tetramer. This dissociation activates the C subunit kinase activity and enables phosphorylation of PKA targets.
Figure 1.
Overview of the cAMP/PKA pathway in S. cerevisiae. Pathway activation occurs in response to glucose via the GPCR (Gpr1) and Ras-based systems. These systems collectively stimulate the adenylate cyclase Cyr1 which increases production of cAMP from ATP. PKA is the only known cAMP receptor in yeast, and upon binding the PKA regulatory subunit, cAMP promotes allosteric changes, which liberate the catalytic subunits from the PKA tetramer. This dissociation activates the C subunit kinase activity and enables phosphorylation of PKA targets.

Figure 2.
Analysis of the three S. cerevisiae PKA catalytic isoform protein sequences. Tpk1-3 sequences (UniProt IDs: P06244, P06245 and P05986, respectively) were aligned and analysed using COBALT [56]. Areas are coloured according to identity between sequences, red showing conservation between the aligned residues and blue showing identity between two Tpk sequences. Low conservation is seen in the N-terminal regions of the three catalytic subunits, with high conservation across the kinase and C-terminal domains.
Figure 2.
Analysis of the three S. cerevisiae PKA catalytic isoform protein sequences. Tpk1-3 sequences (UniProt IDs: P06244, P06245 and P05986, respectively) were aligned and analysed using COBALT [56]. Areas are coloured according to identity between sequences, red showing conservation between the aligned residues and blue showing identity between two Tpk sequences. Low conservation is seen in the N-terminal regions of the three catalytic subunits, with high conservation across the kinase and C-terminal domains.

Figure 3.
PKA control of the environmental stress response (ESR). Environmental stresses including heat shock, oxidative stresses, osmotic stresses and nutrient starvation constitute a constant challenge for yeast cells. The ESR is mediated by the Msn2/4 transcription factors which are regulated by PKA activity. PKA, predominantly Tpk1/3, phosphorylates the NLS of Msn2/4, preventing transport into the nucleus under normal conditions. Reduction in PKA during stress permits Msn2/4 shuttling and abrogates their association with the export factor Msn5 in the nucleus. The TFs are then free to bind stress response elements (STRE) and promote expression of ESR genes required for stress tolerance.
Figure 3.
PKA control of the environmental stress response (ESR). Environmental stresses including heat shock, oxidative stresses, osmotic stresses and nutrient starvation constitute a constant challenge for yeast cells. The ESR is mediated by the Msn2/4 transcription factors which are regulated by PKA activity. PKA, predominantly Tpk1/3, phosphorylates the NLS of Msn2/4, preventing transport into the nucleus under normal conditions. Reduction in PKA during stress permits Msn2/4 shuttling and abrogates their association with the export factor Msn5 in the nucleus. The TFs are then free to bind stress response elements (STRE) and promote expression of ESR genes required for stress tolerance.

Figure 4.
The effects of heat shock and oxidative stress on PKA isoforms. The responses to heat shock and hydrogen peroxide stress both necessitate alterations in PKA activity, with many changes in localisation and activity being stress- and isoform- specific. Under heat stress, Tpk2 and Tpk3 associate with stress-induced compartments, such as P-bodies and stress granules, with Tpk2 localisation dependent on its PrLD. Although Tpk1 localisation is unaltered, TPK1 mRNA coalesces into cytoplasmic granules. H2O2 induces the re-localisation of Tpk1 and Tpk2, but not Tpk3, into cytosolic foci, which is linked to the nuclear accumulation of Msn2/4. As part of the response to hydrogen peroxide, glutathionylation (SG) and sulfenylation (SOH) of conserved cysteine residues in Tpk1 also occurs, specifically downregulating Tpk1 activity.
Figure 4.
The effects of heat shock and oxidative stress on PKA isoforms. The responses to heat shock and hydrogen peroxide stress both necessitate alterations in PKA activity, with many changes in localisation and activity being stress- and isoform- specific. Under heat stress, Tpk2 and Tpk3 associate with stress-induced compartments, such as P-bodies and stress granules, with Tpk2 localisation dependent on its PrLD. Although Tpk1 localisation is unaltered, TPK1 mRNA coalesces into cytoplasmic granules. H2O2 induces the re-localisation of Tpk1 and Tpk2, but not Tpk3, into cytosolic foci, which is linked to the nuclear accumulation of Msn2/4. As part of the response to hydrogen peroxide, glutathionylation (SG) and sulfenylation (SOH) of conserved cysteine residues in Tpk1 also occurs, specifically downregulating Tpk1 activity.

Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Creamer, D.R.; Hubbard, S.J.; Ashe, M.P.; Grant, C.M. Yeast Protein Kinase A Isoforms: A Means of Encoding Specificity in the Response to Diverse Stress Conditions? Biomolecules 2022, 12, 958. https://doi.org/10.3390/biom12070958
AMA Style
Creamer DR, Hubbard SJ, Ashe MP, Grant CM. Yeast Protein Kinase A Isoforms: A Means of Encoding Specificity in the Response to Diverse Stress Conditions? Biomolecules. 2022; 12(7):958. https://doi.org/10.3390/biom12070958
Chicago/Turabian StyleCreamer, Declan R., Simon J. Hubbard, Mark P. Ashe, and Chris M. Grant. 2022. "Yeast Protein Kinase A Isoforms: A Means of Encoding Specificity in the Response to Diverse Stress Conditions?" Biomolecules 12, no. 7: 958. https://doi.org/10.3390/biom12070958
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.