
Activity of Anti-Microbial Peptides (AMPs) against Leishmania and Other Parasites: An Overview

 , , , and
, , , and
Abstract
:1. Introduction
2. Families of AMPs and Their Anti-Infectious Activities
2.1. Melittin
2.1.1. Melittin against Leishmaniasis
2.1.2. Activity on Other Parasites
2.2. Cecropin
2.2.1. Cecropin and Leishmania
2.2.2. Activity on Other Parasites
2.3. Cathelicidin
2.3.1. Cathelicidin and Leishmania
2.3.2. Activity on Other Parasites
2.4. Defensin
2.4.1. Defensin and Leishmania
2.4.2. Activity on Other Parasites
2.5. Magainin
2.5.1. Magainin and Leishmania
2.5.2. Activity on Other Parasites
2.6. Temporin
2.6.1. Temporin and Leishmania
2.6.2. Activity on Other Parasites
2.7. Dermaseptin
2.7.1. Dermaseptin against Leishmaniasis
2.7.2. Activity on Other Parasites
2.8. Eumenitin and Leishmania
2.9. Histatin Effect on Leishmania
3. Conclusions and Future Perspectives
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Bahar, A.A.; Ren, D. Antimicrobial peptides. Pharmacy 2013, 6, 1543–1575. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mwangi, J.; Hao, X.; Lai, R.; Zhang, Z.Y. Antimicrobial peptides: New hope in the war against multidrug resistance. Zool Res. 2019, 40, 488–505. [Google Scholar] [CrossRef] [PubMed]
- Pasupuleti, M.; Schmidtchen, A.; Malmsten, M. Antimicrobial peptides: Key components of the innate immune system. Crit. Rev. Biotechno.l 2012, 32, 143–171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zasloff, M. Antimicrobial peptides of multicellular organisms. Nature 2002, 415, 389–395. [Google Scholar] [CrossRef]
- Hilchie, A.L.; Wuerth, K.; Hancock, R.E.W. Immune modulation by multifaceted cationic host defense (antimicrobial) peptides. Nat. Chem. Biol. 2013, 9, 761–768. [Google Scholar] [CrossRef]
- Kumar, P.; Kizhakkedathu, J.N.; Straus, S.K. Antimicrobial peptides: Diversity, mechanism of action and strategies to improve the activity and biocompatibility in vivo. Biomolecules 2018, 8, 4. [Google Scholar] [CrossRef] [Green Version]
- Teixeira, M.C.; Carbone, C.; Sousa, M.C.; Espina, M.; Garcia, M.L.; Sanchez-Lopez, E.; Souto, E.B. Nanomedicines for the delivery of antimicrobial peptides (Amps). Nanomaterials 2020, 10, 560. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Li, X.; Wang, Z. APD3: The antimicrobial peptide database as a tool for research and education. Nucleic Acids Res. 2016, 44, D1087–D1093. [Google Scholar] [CrossRef] [Green Version]
- Wang, G.; Li, X.; Wang, Z. APD2: The updated antimicrobial peptide database and its application in peptide design. Nucleic Acids Res. 2009, 37, D933–D937. [Google Scholar] [CrossRef] [Green Version]
- Wang, Z.; Wang, G. APD: The Antimicrobial Peptide Database. Nucleic Acids Res. 2004, 32, D590–D592. [Google Scholar] [CrossRef] [Green Version]
- Thomas, S.; Karnik, S.; Barai, R.S.; Jayaraman, V.K.; Idicula-Thomas, S. CAMP: A useful resource for research on antimicrobial peptides. Nucleic Acids Res. 2010, 38, D774–D780. [Google Scholar] [CrossRef] [Green Version]
- Bulet, P.; Hetru, C.; Dimarcq, J.-L.; Hoffmann, D. Antimicrobial peptides in insects; structure and function. Dev. Comp. Immunol. 1999, 23, 329–344. [Google Scholar] [CrossRef]
- Agerberth, B.; Charo, J.; Werr, J.; Olsson, B.; Idali, F.; Lindbom, L.; Kiessling, R.; Joörnvall, H.; Wigzell, H.; Gudmundsson, G.H. The human antimicrobial and chemotactic peptides LL-37 and α-defensins are expressed by specific lymphocyte and monocyte populations. Blood 2000, 96, 3086–3093. [Google Scholar] [CrossRef]
- Sørensen, O.; Cowland, J.B.; Askaa, J.; Borregaard, N. An ELISA for hCAP-18, the cathelicidin present in human neutrophils and plasma. J. Immunol. Methods 1997, 206, 53–59. [Google Scholar] [CrossRef]
- Simmaco, M.; Mignogna, G.; Barra, D. Antimicrobial peptides from amphibian skin: What do they tell us? Pept. Sci. 1998, 47, 435–450. [Google Scholar] [CrossRef]
- Bals, R.; Wang, X.; Zasloff, M.; Wilson, J.M. The peptide antibiotic LL-37/hCAP-18 is expressed in epithelia of the human lung where it has broad antimicrobial activity at the airway surface. Proc. Natl. Acad. Sci. USA 1998, 95, 9541–9546. [Google Scholar] [CrossRef] [Green Version]
- Ma, Y.; Liu, C.; Liu, X.; Wu, J.; Yang, H.; Wang, Y.; Li, J.; Yu, H.; Lai, R. Peptidomics and genomics analysis of novel antimicrobial peptides from the frog, Rana nigrovittata. Genomics 2010, 95, 66–71. [Google Scholar] [CrossRef] [Green Version]
- Oppenheim, J.J.; Biragyn, A.; Kwak, L.W.; Yang, D. Roles of antimicrobial peptides such as defensins in innate and adaptive immunity. Ann. Rheum. Dis. 2003, 62, ii17–ii21. [Google Scholar] [CrossRef] [Green Version]
- Madison, M.N.; Kleshchenko, Y.Y.; Nde, P.N.; Simmons, K.J.; Lima, M.F.; Villalta, F. Human defensin α-1 causes Trypanosoma cruzi membrane pore formation and induces DNA fragmentation, which leads to trypanosome destruction. Infect. Immun. 2007, 75, 4780–4791. [Google Scholar] [CrossRef] [Green Version]
- Yeaman, M.R.; Yount, N.Y. Mechanisms of antimicrobial peptide action and resistance. Pharmacol. Rev. 2003, 55, 27–55. [Google Scholar] [CrossRef] [Green Version]
- Bals, R.; Wilson, J.M. Cathelicidins-A family of multifunctional antimicrobial peptides. Cell. Mol. Life Sci. 2003, 60, 711–720. [Google Scholar] [CrossRef]
- Wade, J.; Lin, F.; Hossain, M.; Dawson, R. Chemical synthesis and biological evaluation of an antimicrobial peptide gonococcal growth inhibitor. Amino Acids 2012, 43, 2279–2283. [Google Scholar] [CrossRef]
- Papo, N.; Oren, Z.; Pag, U.; Sahl, H.-G.; Shai, Y. The Consequence of Sequence Alteration of an Amphipathic α-Helical Antimicrobial Peptide and Its Diastereomers. J. Biol. Chem. 2002, 277, 33913–33921. [Google Scholar] [CrossRef] [Green Version]
- Rashid, R.; Veleba, M.; Kline, K.A. Focal Targeting of the Bacterial Envelope by Antimicrobial Peptides. Front. Cell Dev. Biol. 2016, 4, 55. [Google Scholar] [CrossRef]
- Shagaghi, N.; Palombo, E.A.; Clayton, A.H.A.; Bhave, M. Antimicrobial peptides: Biochemical determinants of activity and biophysical techniques of elucidating their functionality. World J. Microbiol. Biotechnol. 2018, 34, 1–13. [Google Scholar] [CrossRef]
- World Health Organization. Antimicrobial Resistance. Global Report on Surveillance; World Health Organization: Geneva, Switzerland, 2014; pp. 12–28. [Google Scholar]
- Batoni, G.; Maisetta, G.; Brancatisano, F.L.; Esin, S.; Campa, M. Use of antimicrobial peptides against microbial biofilms: Advantages and limits. Curr. Med. Chem. 2011, 18, 256–279. [Google Scholar] [CrossRef]
- Xu, W.; Zhu, X.; Tan, T.; Li, W.; Shan, A. Design of Embedded-Hybrid Antimicrobial Peptides with Enhanced Cell Selectivity and Anti-Biofilm Activity. PLoS ONE 2014, 9, e098935. [Google Scholar] [CrossRef]
- De La Fuente-Núñez, C.; Cardoso, M.H.; De Souza Cândido, E.; Franco, O.L.; Hancock, R.E.W. Synthetic antibiofilm peptides. Biochim. Biophys. Acta 2016, 1858, 1061–1069. [Google Scholar] [CrossRef] [PubMed]
- Oliveira, M.; Gomes-Alves, A.G.; Sousa, C.; Mirta Marani, M.; Plácido, A.; Vale, N.; Delerue-Matos, C.; Gameiro, P.; Kückelhaus, S.A.S.; Tomas, A.M.; et al. Ocellatin-PT antimicrobial peptides: High-resolution microscopy studies in antileishmania models and interactions with mimetic membrane systems. Biopolymers 2016, 105, 873–886. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Karunaweera, N.D.; Ferreira, M.U. Leishmaniasis: Current challenges and prospects for elimination with special focus on the South Asian region. Parasitology 2018, 145, 425–429. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- World Health Organization. Leishmaniasis 2018 (22nd December); World Health Organization: Geneva, Switzerland, 2018. Available online: http://www.who.int/mediacentre/factsheets/fs375/en/ (accessed on September 2020).
- Nimsarkar, P.; Ingale, P.; Singh, S. Systems Studies Uncover miR-146a as a Target in Leishmania major Infection Model. ACS Omega 2020, 5, 12516–12526. [Google Scholar] [CrossRef]
- Sunter, J.; Gull, K. Shape, form, function and Leishmania pathogenicity: From textbook descriptions to biological understanding. Open Biol. 2017, 7, 170165. [Google Scholar] [CrossRef] [Green Version]
- World Health Organization. Control of the leishmaniases. Report of a Meeting of the WHO Expert Committee on the Control of Leishmaniases, Geneva, Switzerland, 22–26 March 2010. [Google Scholar]
- Lewis, D.J. Phlebotomid Sandflies. Bull World Health Organ. 1971, 44, 535–551. [Google Scholar]
- Meinecke, C.K.; Schottelius, J.; Oskam, L.; Fleischer, B. Congenital transmission of visceral leishmaniasis (Kala Azar) from an asymptomatic mother to her child. Pediatrics 1999, 104, e65. [Google Scholar] [CrossRef] [Green Version]
- Cruz, I.; Morales, M.A.; Noguer, I.; Rodríguez, A.; Alvar, J. Leishmania in discarded syringes from intravenous drug users. Lancet 2002, 359, 1124–1125. [Google Scholar] [CrossRef]
- Cohen, C.; Corazza, F.; De Mol, P.; Brasseur, D. Leishmaniasis acquired in Belgium. Lancet (Lond. England) 1991, 338, 128. [Google Scholar] [CrossRef]
- Reithinger, R.; Dujardin, J.-C.; Louzir, H.; Pirmez, C.; Alexander, B.; Brooker, S. Cutaneous leishmaniasis. Lancet. Infect. Dis. 2007, 7, 581–596. [Google Scholar] [CrossRef] [Green Version]
- Blum, J.; Lockwood, D.N.J.; Visser, L.; Harms, G.; Bailey, M.S.; Caumes, E.; Clerinx, J.; van Thiel, P.P.A.M.; Morizot, G.; Hatz, C.; et al. Local or systemic treatment for New World cutaneous leishmaniasis? Re-evaluating the evidence for the risk of mucosal leishmaniasis. Int. Health 2012, 4, 153–163. [Google Scholar] [CrossRef]
- Oliveira, L.F.; Schubach, A.O.; Martins, M.M.; Passos, S.L.; Oliveira, R.V.; Marzochi, M.C.; Andrade, C.A. Systematic review of the adverse effects of cutaneous leishmaniasis treatment in the New World. Acta Trop. 2011, 118, 87–96. [Google Scholar] [CrossRef]
- Wortmann, G.; Zapor, M.; Ressner, R.; Fraser, S.; Hartzell, J.; Pierson, J.; Weintrob, A.; Magill, A. Lipsosomal amphotericin B for treatment of cutaneous leishmaniasis. Am. J. Trop. Med. Hyg. 2010, 83, 1028–1033. [Google Scholar] [CrossRef] [Green Version]
- Vacchina, P.; Morales, M.A. In vitro screening test using Leishmania promastigotes stably expressing mCherry protein. Antimicrob. Agents Chemother. 2014, 58, 1825–1828. [Google Scholar] [CrossRef] [Green Version]
- Pace, D. Leishmaniasis. J. Infect. 2014, 69, S10–S18. [Google Scholar] [CrossRef]
- Fernández-Rubio, C.; Larrea, E.; Guerrero, J.P.; Herrero, E.S.; Gamboa, I.; Berrio, C.; Plano, D.; Amin, S.; Sharma, A.K.; Nguewa, P.A. Leishmanicidal activity of isoselenocyanate derivatives. Antimicrob. Agents Chemother. 2019, 63, e00904-18. [Google Scholar] [CrossRef] [Green Version]
- Ibanez, E.; Agliano, A.; Prior, C.; Nguewa, P.; Redrado, M.; Gonzalez-Zubeldia, I.; Plano, D.; Palop, J.A.; Sanmartin, C.; Calvo, A. The Quinoline Imidoselenocarbamate EI201 Blocks the AKT/mTOR Pathway and Targets Cancer Stem Cells Leading to a Strong Antitumor Activity. Curr. Med. Chem. 2012, 19, 3031–3043. [Google Scholar] [CrossRef]
- Fernández-rubio, C.; Campbell, D.; Vacas, A.; Ibañez, E.; Moreno, E.; Espuelas, S. Leishmanicidal Activities of Novel Methylseleno-Imidocarbamates. Antimicrob. Agents Chemother. 2015, 59, 5705–5713. [Google Scholar] [CrossRef] [Green Version]
- Puig-Rigall, J.; Fernández-Rubio, C.; González-Benito, J.; Houston, J.E.; Radulescu, A.; Nguewa, P.; González-Gaitano, G. Structural characterization by scattering and spectroscopic methods and biological evaluation of polymeric micelles of poloxamines and TPGS as nanocarriers for miltefosine delivery. Int. J. Pharm. 2020, 578, 119057. [Google Scholar] [CrossRef]
- Sepehri, A.; PeBenito, L.; Pino-Angeles, A.; Lazaridis, T. Membrane Pore Formation by Melittin Derivatives. Biophys. J. 2020, 118, 234a. [Google Scholar] [CrossRef]
- Scheenstra, M.R.; van den Belt, M.; Tjeerdsma-van Bokhoven, J.L.M.; Schneider, V.A.F.; Ordonez, S.R.; van Dijk, A.; Veldhuizen, E.J.A.; Haagsman, H.P. Cathelicidins PMAP-36, LL-37 and CATH-2 are similar peptides with different modes of action. Sci. Rep. 2019, 9, 1–12. [Google Scholar] [CrossRef]
- Coorens, M.; Schneider, V.A.F.; de Groot, A.M.; van Dijk, A.; Meijerink, M.; Wells, J.M.; Scheenstra, M.R.; Veldhuizen, E.J.A.; Haagsman, H.P. Cathelicidins Inhibit Escherichia coli–Induced TLR2 and TLR4 Activation in a Viability-Dependent Manner. J. Immunol. 2017, 199, 1418–1428. [Google Scholar] [CrossRef] [Green Version]
- Kvansakul, M.; Lay, F.T.; Adda, C.G.; Veneer, P.K.; Baxter, A.A.; Phan, T.K.; Poon, I.K.H.; Hulett, M.D. Binding of phosphatidic acid by NsD7 mediates the formation of helical defensin-lipid oligomeric assemblies and membrane permeabilization. Proc. Natl. Acad. Sci. USA 2016, 113, 11202–11207. [Google Scholar] [CrossRef] [Green Version]
- Järvå, M.; Lay, F.T.; Phan, T.K.; Humble, C.; Poon, I.K.H.; Bleackley, M.R.; Anderson, M.A.; Hulett, M.D.; Kvansakul, M. X-ray structure of a carpet-like antimicrobial defensin-phospholipid membrane disruption complex. Nat. Commun. 2018, 9, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Järvå, M.; Phan, T.K.; Lay, F.T.; Caria, S.; Kvansakul, M.; Hulett, M.D. Human β-defensin 2 kills Candida albicans through phosphatidylinositol 4,5-bisphosphate–mediated membrane permeabilization. Sci. Adv. 2018, 4, eaat0979. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Farrotti, A.; Conflitti, P.; Srivastava, S.; Ghosh, J.; Palleschi, A.; Stella, L.; Bocchinfuso, G. Molecular Dynamics Simulations of the Host Defense Peptide Temporin L and Its Q3K Derivative: An Atomic Level View from Aggregation in Water to Bilayer Perturbation. Molecules 2017, 22, 1235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eggimann, G.; Sweeney, K.; Bolt, H.; Rozatian, N.; Cobb, S.; Denny, P. The Role of Phosphoglycans in the Susceptibility of Leishmania mexicana to the Temporin Family of Anti-Microbial Peptides. Molecules 2015, 20, 2775–2785. [Google Scholar] [CrossRef] [Green Version]
- Arcisio-Miranda, M.; dos Santos Cabrera, M.P.; Konno, K.; Rangel, M.; Procopio, J. Effects of the cationic antimicrobial peptide eumenitin from the venom of solitary wasp Eumenes rubronotatus in planar lipid bilayers: Surface charge and pore formation activity. Toxicon 2008, 51, 736–745. [Google Scholar] [CrossRef]
- Jang, W.S.; Bajwa, J.S.; Sun, J.N.; Edgerton, M. Salivary histatin 5 internalization by translocation, but not endocytosis, is required for fungicidal activity in Candida albicans. Mol. Microbiol. 2010, 77, 354–370. [Google Scholar] [CrossRef] [Green Version]
- Oñate-Garzón, J.; Manrique-Moreno, M.; Trier, S.; Leidy, C.; Torres, R.; Patiño, E. Antimicrobial activity and interactions of cationic peptides derived from Galleria mellonella cecropin D-like peptide with model membranes. J. Antibiot. 2017, 70, 238–245. [Google Scholar] [CrossRef]
- Peng, J.; Wu, Z.; Liu, W.; Long, H.; Zhu, G.; Guo, G.; Wu, J. Antimicrobial functional divergence of the cecropin antibacterial peptide gene family in Musca domestica. Parasites Vectors 2019, 12, 537. [Google Scholar] [CrossRef]
- Silvestro, L.; Weiser, J.N.; Axelsen, P.H.; Gupta, K.; Weiser, J.H.; Axelsen, P.H. Antibacterial and Antimembrane Activities of Cecropin A in Escherichia coli. Antimicrob. Agents Chemother. 2000, 44, 602–607. [Google Scholar] [CrossRef] [Green Version]
- Amini, A.; Raheem, S.; Steiner, A.; Deeba, F.; Ahmad, Z. Insect venom peptides as potent inhibitors of Escherichia coli ATP synthase. Int. J. Biol. Macromol. 2020, 150, 23–30. [Google Scholar] [CrossRef]
- Dos Santos Cabrera, M.P.; Arcisio-Miranda, M.; da Costa, L.C.; de Souza, B.M.; Broggio Costa, S.T.; Palma, M.S.; Ruggiero Neto, J.; Procopio, J. Interactions of mast cell degranulating peptides with model membranes: A comparative biophysical study. Arch. Biochem. Biophys. 2009, 486, 1–11. [Google Scholar] [CrossRef]
- Konno, K.; Hisada, M.; Naoki, H.; Itagaki, Y.; Fontana, R.; Rangel, M.; Oliveira, J.S.; Dos santos Cabrera, M.P.; Neto, J.R.; Hide, I.; et al. Eumenitin, a novel antimicrobial peptide from the venom of the solitary eumenine wasp Eumenes rubronotatus. Peptides 2006, 27, 2624–2631. [Google Scholar] [CrossRef]
- Vylkova, S.; Sun, J.N.; Edgerton, M. The role of released ATP in killing Candida albicans and other extracellular microbial pathogens by cationic peptides. Purinergic Signal. 2007, 3, 91–97. [Google Scholar] [CrossRef] [Green Version]
- Saha, A.K.; Mukherjee, T.; Bhaduri, A. Mechanism of action of amphotericin B on Leishmania donovani promastigotes. Mol. Biochem. Parasitol. 1986, 19, 195–200. [Google Scholar] [CrossRef]
- Murray, H.W.; Delph-Etienne, S. Roles of Endogenous Gamma Interferon and Macrophage Microbicidal Mechanisms in Host Response to Chemotherapy in Experimental Visceral Leishmaniasis. Infect. Immun. 2000, 68, 288–293. [Google Scholar] [CrossRef] [Green Version]
- Paris, C.; Loiseau, P.M.; Bories, C.; Bréard, J. Miltefosine Induces Apoptosis-Like Death in Leishmania donovani Promastigotes. Antimicrob. Agents Chemother. 2004, 48, 852–859. [Google Scholar] [CrossRef] [Green Version]
- Verma, N.K.; Dey, C.S. Possible Mechanism of Miltefosine-Mediated Death of Leishmania donovani. Antimicrob. Agents Chemother. 2004, 48, 3010–3015. [Google Scholar] [CrossRef] [Green Version]
- Zuo, X.; Djordjevic, J.T.; Bijosono Oei, J.; Desmarini, D.; Schibeci, S.D.; Jolliffe, K.A.; Sorrell, T.C. Miltefosine Induces Apoptosis-Like Cell Death in Yeast via Cox9p in Cytochrome c Oxidase. Mol. Pharmacol. 2011, 80, 476–485. [Google Scholar] [CrossRef] [Green Version]
- Wadhone, P.; Maiti, M.; Agarwal, R.; Kamat, V.; Martin, S.; Saha, B. Miltefosine Promotes IFN-γ-Dominated Anti-Leishmanial Immune Response. J. Immunol. 2009, 182, 7146–7154. [Google Scholar] [CrossRef] [Green Version]
- Murray, H.W.; Montelibano, C.; Peterson, R.; Sypek, J.P. Interleukin-12 Regulates the Response to Chemotherapy in Experimental Visceral Leishmaniasis. J. Infect. Dis. 2000, 182, 1497–1502. [Google Scholar] [CrossRef]
- Mookerjee Basu, J.; Mookerjee, A.; Sen, P.; Bhaumik, S.; Sen, P.; Banerjee, S.; Naskar, K.; Choudhuri, S.K.; Saha, B.; Raha, S.; et al. Sodium Antimony Gluconate Induces Generation of Reactive Oxygen Species and Nitric Oxide via Phosphoinositide 3-Kinase and Mitogen-Activated Protein Kinase Activation in Leishmania donovani-Infected Macrophages. Antimicrob. Agents Chemother. 2006, 50, 1788–1797. [Google Scholar] [CrossRef] [Green Version]
- Pereira, A.V.; de Barros, G.; Pinto, E.G.; Tempone, A.G.; Orsi, R.d.O.; Dos Santos, L.D.; Calvi, S.; Ferreira, R.S.; Pimenta, D.C.; Barraviera, B. Melittin induces in vitro death of Leishmania (Leishmania) infantum by triggering the cellular innate immune response. J. Venom. Anim. Toxins Incl. Trop. Dis. 2016, 22, 1. [Google Scholar] [CrossRef] [Green Version]
- Raghuraman, H.; Chattopadhyay, A. Melittin: A membrane-active peptide with diverse functions. Biosci. Rep. 2007, 27, 189–223. [Google Scholar] [CrossRef]
- Adade, C.M.; Chagas, G.S.F.; Souto-Padrón, T. Apis mellifera venom induces different cell death pathways in Trypanosoma cruzi. Parasitology 2012, 139, 1444–1461. [Google Scholar] [CrossRef]
- Papo, N.; Shai, Y. Can we predict biological activity of antimicrobial peptides from their interactions with model phospholipid membranes? Peptides 2003, 24, 1693–1703. [Google Scholar] [CrossRef] [PubMed]
- Pérez-Cordero, J.J.; Lozano, J.M.; Cortés, J.; Delgado, G. Leishmanicidal activity of synthetic antimicrobial peptides in an infection model with human dendritic cells. Peptides 2011, 32, 683–690. [Google Scholar] [CrossRef] [PubMed]
- Adade, C.M.; Oliveira, I.R.S.; Pais, J.A.R.; Souto-Padrón, T. Melittin peptide kills Trypanosoma cruzi parasites by inducing different cell death pathways. Toxicon 2013, 69, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Díaz-Achirica, P.; Ubach, J.; Guinea, A.; Andreu, D.; Rivas, L. The plasma membrane of Leishmania donovani promastigotes is the main target for CA(1-8)M(1-18), a synthetic cecropin A-melittin hybrid peptide. Biochem. J. 1998, 330, 453–460. [Google Scholar] [CrossRef] [PubMed]
- Mahdavi Abhari, F.; Pirestani, M.; Dalimi, A. Anti-amoebic activity of a cecropin-melittin hybrid peptide (CM11) against trophozoites of Entamoeba histolytica. Wien. Klin. Wochenschr. 2019, 131, 427–434. [Google Scholar] [CrossRef] [PubMed]
- Carter, V.; Underhill, A.; Baber, I.; Sylla, L.; Baby, M.; Larget-Thiery, I.; Zettor, A.; Bourgouin, C.; Langel, Ü.; Faye, I.; et al. Killer Bee Molecules: Antimicrobial Peptides as Effector Molecules to Target Sporogonic Stages of Plasmodium. PLoS Pathog. 2013, 9, e1003790. [Google Scholar] [CrossRef] [Green Version]
- Brady, D.; Grapputo, A.; Romoli, O.; Sandrelli, F. Insect cecropins, antimicrobial peptides with potential therapeutic applications. Int. J. Mol. Sci. 2019, 20, 5862. [Google Scholar] [CrossRef] [Green Version]
- Akuffo, H.; Hultmark, D.; Engstöm, A.; Frohlich, D.; Kimbrell, D. Drosophila antibacterial protein, cecropin A, differentially affects non-bacterial organisms such as Leishmania in a manner different from other amphipathic peptides. Int. J. Mol. Med. 1998, 1, 77–159. [Google Scholar] [CrossRef]
- Patiño-Márquez, I.A.; Patiño-González, E.; Hernández-Villa, L.; Ortíz-Reyes, B.; Manrique-Moreno, M. Identification and evaluation of Galleria mellonella peptides with antileishmanial activity. Anal. Biochem. 2018, 546, 35–42. [Google Scholar] [CrossRef]
- Hong, S.-M.; Kusakabe, T.; Lee, J.-M.; Tatsuke, T.; Kawaguchi, Y.; Kang, M.-W.; Kang, S.-W.; Kim, K.-A.; Nho, S.-K. Structure and Expression Analysis of the Cecropin-E Gene from the Silkworm. Biotechnol. Biochem. 2008, 72, 1992–1998. [Google Scholar] [CrossRef]
- Al-Hyali, N.S.; Khalil, L.Y.; Aljawady, M.A. Sarcotoxin effect on leukocytic finding and phagocytic activity in mice. J. Anim. Vet. Adv. 2009, 8, 2395–2398. [Google Scholar]
- Wu, Q.; Patočka, J.; Kuča, K. Insect antimicrobial peptides, a mini review. Toxins 2018, 10, 461. [Google Scholar] [CrossRef]
- Ponnuvel, K.M.; Subhasri, N.; Sirigineedi, S.; Murthy, G.N.; Vijayaprakash, N.B. Molecular evolution of the cecropin multigene family in silkworm Bombyx mori. Bioinformation 2010, 5, 97–103. [Google Scholar] [CrossRef] [Green Version]
- Yi, H.Y.; Chowdhury, M.; Huang, Y.D.; Yu, X.Q. Insect antimicrobial peptides and their applications. Appl. Microbiol. Biotechnol. 2014, 98, 5807–5822. [Google Scholar] [CrossRef] [Green Version]
- Samakovlis, C.; Kimbrell, D.A.; Kylsten, P.; Engstrom, A.; Hultmark, D. The immune response in Drosophila: Pattern of cecropin expression and biological activity. EMBO J. 1990, 9, 2969–2976. [Google Scholar] [CrossRef]
- Suttmann, H.; Retz, M.; Paulsen, F.; Harder, J.; Zwergel, U.; Kamradt, J.; Wullich, B.; Unteregger, G.; Stöckle, M.; Lehmann, J. Antimicrobial peptides of the Cecropin-family show potent antitumor activity against bladder cancer cells. BMC Urol. 2008. [Google Scholar] [CrossRef] [Green Version]
- Brown, S.E.; Howard, A.; Kasprzak, A.B.; Gordon, K.H.; East, P.D. The discovery and analysis of a diverged family of novel antifungal moricin-like peptides in the wax moth Galleria mellonella. Insect Biochem. Mol. Biol. 2008, 8, 5. [Google Scholar] [CrossRef]
- Gwadz, R.W.; Kaslow, D.; Lee, J.Y.; Maloy, W.L.; Zasloff, M.; Miller, L.H. Effects of magainins and cecropins on the sporogonic development of malaria parasites in mosquitoes. Infect. Immun. 1989, 57, 2628–2633. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, W.; Koo, H.; Richman, A.M.; Seeley, D.; Vizioli, J.; Klocko, A.D.; O’Brochta, D.A. Ectopic expression of a cecropin transgene in the human malaria vector mosquito Anopheles gambiae (Diptera: Culicidae): Effects on susceptibility to Plasmodium. J. Med. Entomol. 2004, 41, 447–455. [Google Scholar] [CrossRef] [Green Version]
- Boulanger, N.; Brun, R.; Ehret-Sabatier, L.; Kunz, C.; Bulet, P. Immunopeptides in the defense reactions of Glossina morsitans to bacterial and Trypanosoma brucei brucei infections. Insect Biochem. Mol. Biol. 2002, 32, 369–375. [Google Scholar] [CrossRef]
- Jaynes, J.M.; Burton, C.A.; Barr, S.B.; Jeffers, G.W.; Julian, G.R.; White, K.L.; Enright, F.M.; Klei, T.R.; Laine, R.A. In vitro cytocidal effect of novel lytic peptides on Plasmodium falciparum and Trypanosoma cruzi. FASEB J. Off. Publ. Fed. Am. Soc. Exp. Biol. 1988, 2, 2878–2883. [Google Scholar] [CrossRef] [Green Version]
- Kościuczuk, E.M.; Lisowski, P.; Jarczak, J.; Strzałkowska, N.; Jóźwik, A.; Horbańczuk, J.; Krzyżewski, J.; Zwierzchowski, L.; Bagnicka, E. Cathelicidins: Family of antimicrobial peptides. A review. Mol. Biol. Rep. 2012, 39, 10957–10970. [Google Scholar] [CrossRef] [Green Version]
- Galloo, R.L.; Kim, K.J.; Bernfield, M.; Kozak, C.A.; Zanetti, M.; Merluzzi, L.; Gennaro, R. Identification of CRAMP, a cathelin-related antimicrobial peptide expressed in the embryonic and adult mouse. J. Biol. Chem. 1997, 272, 13088–13093. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.; Shin, V.Y.; Ho, J.C.W.; Siu, M.T.; Cheuk, I.W.Y.; Kwong, A. Functional implications of cathelicidin antimicrobial protein in breast cancer and tumor-associated macrophage microenvironment. Biomolecules 2020, 10, 688. [Google Scholar] [CrossRef]
- Crauwels, P.; Bank, E.; Walber, B.; Wenzel, U.A.; Agerberth, B.; Chanyalew, M.; Abebe, M.; König, R.; Ritter, U.; Reiling, N.; et al. Cathelicidin Contributes to the Restriction of Leishmania in Human Host Macrophages. Front. Immunol. 2019, 10, 2697. [Google Scholar] [CrossRef]
- Das, S.; Sardar, A.H.; Abhishek, K.; Kumar, A.; Rabidas, V.N.; Das, P. Cathelicidin augments VDR-dependent anti-leishmanial immune response in Indian Post-Kala-Azar Dermal Leishmaniasis. Int. Immunopharmacol. 2017, 50, 130–138. [Google Scholar] [CrossRef]
- Marr, A.K.; Cen, S.; Hancock, R.E.W.; McMaster, W.R. Identification of synthetic and natural host defense peptides with leishmanicidal activity. Antimicrob. Agents Chemother. 2016, 60, 2484–2491. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lynn, M.A.; Kindrachuk, J.; Marr, A.K.; Jenssen, H.; Panté, N.; Elliott, M.R.; Napper, S.; Hancock, R.E.; McMaster, W.R. Effect of BMAP-28 antimicrobial peptides on Leishmania major promastigote and amastigote growth: Role of leishmanolysin in parasite survival. PLoS Negl. Trop. Dis. 2011, 5, e1141. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cauchard, S.; Van Reet, N.; Büscher, P.; Goux, D.; Grötzinger, J.; Leippe, M.; Cattoir, V.; Laugier, C.; Cauchard, J. Killing of trypanozoon parasites by the equine cathelicidin eCATH1. Antimicrob. Agents Chemother. 2016, 60, 2610–2619. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fang, Y.; He, X.; Zhang, P.; Shen, C.; Mwangi, J.; Xu, C.; Mo, G.; Lai, R.; Zhang, Z. In vitro and in vivo antimalarial activity of LZ1, a peptide derived from snake cathelicidin. Toxins 2019, 11, 379. [Google Scholar] [CrossRef] [Green Version]
- Meade, K.G.; O’Farrelly, C. Β-Defensins: Farming the microbiome for homeostasis and health. Front. Immunol. 2019, 10, 1–20. [Google Scholar] [CrossRef]
- Dong, H.; Lv, Y.; Zhao, D.; Barrow, P.; Zhou, X. Defensins: The Case for Their Use against Mycobacterial Infections. J. Immunol. Res. 2016, 2016, 7515687. [Google Scholar] [CrossRef] [Green Version]
- Kerenga, B.K.; McKenna, J.A.; Harvey, P.J.; Quimbar, P.; Garcia-Ceron, D.; Lay, F.T.; Phan, T.K.; Veneer, P.K.; Vasa, S.; Parisi, K.; et al. Salt-Tolerant Antifungal and Antibacterial Activities of the Corn Defensin ZmD32. Front. Microbiol. 2019, 10, 795. [Google Scholar] [CrossRef]
- Daneshvar, H.; Tavakoli Kareshk, A.; Sharifi, I.; Keyhani, A.; Tavakoli Oliaee, R.; Asadi, A. Host-parasite responses outcome regulate the expression of antimicrobial peptide genes in the skin of BALB/c and c57bl/6 murine strains following leishmania major MRHO/IR/75/ER infection. Iran. J. Parasitol. 2018, 13, 515–523. [Google Scholar]
- Souza, G.S.; de Carvalho, L.P.; de Melo, E.J.T.; da Silva, F.C.V.; Machado, O.L.T.; Gomes, V.M.; de Oliveira Carvalho, A. A synthetic peptide derived of the β2–β3 loop of the plant defensin from Vigna unguiculata seeds induces Leishmania amazonensis apoptosis-like cell death. Amino Acids 2019, 51, 1633–1648. [Google Scholar] [CrossRef]
- Pero, R.; Coretti, L.; Nigro, E.; Lembo, F.; Laneri, S.; Lombardo, B.; Daniele, A.; Scudiero, O. β-defensins in the fight against Helicobacter pylori. Molecules 2017, 22, 424. [Google Scholar] [CrossRef]
- Polesello, V.; Segat, L.; Crovella, S.; Zupin, L. Candida Infections and Human Defensins. Protein Pept. Lett. 2017, 24, 747–756. [Google Scholar] [CrossRef]
- Yoo, Y.J.; Kwon, I.; Oh, S.R.; Perinpanayagam, H.; Lim, S.M.; Ahn, K.B.; Lee, Y.; Han, S.H.; Chang, S.W.; Baek, S.H.; et al. Antifungal Effects of Synthetic Human Beta-defensin-3-C15 Peptide on Candida albicans-infected Root Dentin. J. Endod. 2017, 43, 1857–1861. [Google Scholar] [CrossRef]
- Wilson, S.S.; Wiens, M.E.; Smith, J.G. Antiviral mechanisms of human defensins. J. Mol. Biol. 2013, 425, 4965–4980. [Google Scholar] [CrossRef]
- Holly, M.K.; Diaz, K.; Smith, J.G. Defensins in Viral Infection and Pathogenesis. Annu. Rev. Virol. 2017, 4, 369–391. [Google Scholar] [CrossRef]
- Sarkar, P.; Jana, K.; Sikdar, S.R. Overexpression of biologically safe Rorippa indica defensin enhances aphid tolerance in Brassica juncea. Planta 2017, 246, 1029–1044. [Google Scholar] [CrossRef]
- Cabezas-cruz, A.; Tonk, M.; Bouchut, A.; Pierrot, C.; Pierce, R.J.; Kotsyfakis, M.; Rahnamaeian, M.; Vilcinskas, A.; Khalife, J.; Valdés, J.J. Antiplasmodial Activity Is an Ancient and Conserved Feature of Tick Defensins. Front. Microbiol. 2016, 7, 1682. [Google Scholar] [CrossRef]
- Couto, J.; Tonk, M.; Ferrolho, J.; Antunes, S.; Vilcinskas, A.; de la Fuente, J.; Domingos, A.; Cabezas-Cruz, A. Antiplasmodial activity of tick defensins in a mouse model of malaria. Ticks Tick. Borne. Dis. 2018, 9, 844–849. [Google Scholar] [CrossRef]
- Morampudi, V.; Braun, M.Y.; Souza, S.D. Modulation of Early β-Defensin-2 Production as a Mechanism Developed by Type I Toxoplasma gondii To Evade Human Intestinal Immunity. Infect. Immun. 2011, 79, 2043–2050. [Google Scholar] [CrossRef] [Green Version]
- Carryn, S.; Schaefer, D.A.; Imboden, M.; Homan, E.J.; Bremel, R.D.; Riggs, M.W. Phospholipases and Cationic Peptides Inhibit Cryptosporidium parvum Sporozoite Infectivity by Parasiticidal and Non-Parasiticidal Mechanisms. J. Parasitol. 2012, 98, 199–204. [Google Scholar] [CrossRef]
- Pino-Angeles, A.; Leveritt, J.M.; Lazaridis, T. Pore Structure and Synergy in Antimicrobial Peptides of the Magainin Family. PLoS Comput. Biol. 2016, 12, 1004570. [Google Scholar] [CrossRef] [Green Version]
- Pincus, M. Physiological structure and function of proteins. Cell Physiol. Source Book 2012, 19–42. [Google Scholar] [CrossRef]
- Bagheri, M. Aggregation vs. Fusion of Negatively Charged Lipid Bilayers Induced by Bactenecin and Magainin Derivatives. Biophys. J. 2018, 114, 453a. [Google Scholar] [CrossRef]
- Hasan, M.; Karal, M.A.S.; Levadnyy, V.; Yamazaki, M. Mechanism of Initial Stage of Pore Formation Induced by Antimicrobial Peptide Magainin 2. Langmuir 2018, 34, 3349–3362. [Google Scholar] [CrossRef]
- Bacalum, M.; Radu, M. Cationic Antimicrobial Peptides Cytotoxicity on Mammalian Cells: An Analysis Using Therapeutic Index Integrative Concept. Int. J. Pept. Res. Ther. 2015, 21, 47–55. [Google Scholar] [CrossRef]
- Patocka, J.; Nepovimova, E.; Klimova, B.; Wu, Q.; Kuca, K. Antimicrobial Peptides: Amphibian Host Defense Peptides. Curr. Med. Chem. 2018, 26, 5924–5946. [Google Scholar] [CrossRef]
- Nyström, L.; Malmsten, M. Membrane interactions and cell selectivity of amphiphilic anticancer peptides. Curr. Opin. Colloid Interface Sci. 2018, 38, 1–17. [Google Scholar] [CrossRef]
- Guerrero, E.; Saugar, J.M.; Matsuzaki, K.; Rivas, L. Role of positional hydrophobicity in the leishmanicidal activity of magainin 2. Antimicrob. Agents Chemother. 2004, 48, 2980–2986. [Google Scholar] [CrossRef] [Green Version]
- Kulkarni, M.M.; Karafova, A.; Kamysz, W.; McGwire, B.S. Design of protease-resistant pexiganan enhances antileishmanial activity. Parasitol. Res. 2014, 113, 1971–1976. [Google Scholar] [CrossRef] [PubMed]
- Giacometti, A.; Cirioni, O.; Simona, M.; Prete, D.E.L.; Barchiesi, F. Short-Term Exposure to Membrane-Active Antibiotics Inhibits Cryptosporidium parvum Infection in Cell Culture. Antimicrob. Agents Chemother. 2000, 44, 3473–3475. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ladram, A.; Nicolas, P. Antimicrobial peptides from frog skin: Biodiversity and therapeutic promises. Front. Biosci.–Landmark 2016, 21, 1341–1371. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Crépin, A.; Jégou, J.-F.; André, S.; Ecale, F.; Croitoru, A.; Cantereau, A.; Berjeaud, J.-M.; Ladram, A.; Verdon, J. In vitro and intracellular activities of frog skin temporins against Legionella pneumophila and its eukaryotic hosts. Sci. Rep. 2020, 1, 3978. [Google Scholar] [CrossRef] [Green Version]
- Maisetta, G.; Grassi, L.; Di Luca, M.; Bombardelli, S.; Medici, C.; Brancatisano, F.L.; Esin, S.; Batoni, G. Anti-biofilm properties of the antimicrobial peptide temporin 1Tb and its ability, in combination with EDTA, to eradicate Staphylococcus epidermidis biofilms on silicone catheters. Biofouling 2016, 32, 787–800. [Google Scholar] [CrossRef]
- Marimuthu, S.K.; Nagarajan, K.; Perumal, S.K.; Palanisamy, S.; Subbiah, L. Insilico Alpha-Helical Structural Recognition of Temporin Antimicrobial Peptides and Its Interactions with Middle East Respiratory Syndrome-Coronavirus. Int. J. Pept. Res. Ther. 2020, 26, 1473–1483. [Google Scholar] [CrossRef] [Green Version]
- Mishra, B.; Wang, X.; Lushnikova, T.; Zhang, Y.; Golla, R.M.; Narayana, J.L.; Wang, C.; McGuire, T.R.; Wang, G. Antibacterial, antifungal, anticancer activities and structural bioinformatics analysis of six naturally occurring temporins. Peptides 2018, 106, 9–20. [Google Scholar] [CrossRef]
- Holden, W.M.; Reinert, L.K.; Hanlon, S.M.; Parris, M.J.; Rollins-Smith, L.A. Development of antimicrobial peptide defenses of southern leopard frogs, Rana sphenocephala, against the pathogenic chytrid fungus, Batrachochytrium dendrobatidis. Dev. Comp. Immunol. 2015, 48, 65–75. [Google Scholar] [CrossRef]
- Abbassi, F.; Humblot, V.; Lequin, O.; Raja, Z.; Andre, S.; Bouceba, T.; Correia, I.; Casale, S.; Foulon, T.; Sereno, D.; et al. Insight into the mechanism of action of temporin-SHa, a new broad-spectrum antiparasitic and antibacterial agent. PLoS ONE 2017, 12, e0174024. [Google Scholar] [CrossRef] [Green Version]
- Swithenbank, L.; Cox, P.; Harris, L.G.; Dudley, E.; Sinclair, K.; Lewis, P.; Cappiello, F.; Morgan, C. Temporin A and Bombinin H2 Antimicrobial Peptides Exhibit Selective Cytotoxicity to Lung Cancer Cells. Scientifica 2020, 2020, 3526286. [Google Scholar] [CrossRef]
- Mangoni, M.L.; Saugar, J.M.; Dellisanti, M.; Barra, D.; Simmaco, M.; Rivas, L. Temporins, small antimicrobial peptides with leishmanicidal activity. J. Biol. Chem. 2005, 280, 984–990. [Google Scholar] [CrossRef] [Green Version]
- André, S.; Raja, Z.; Humblot, V.; Piesse, C.; Foulon, T.; Sereno, D.; Oury, B.; Ladram, A. Functional characterization of temporin-she, a new broad-spectrum antibacterial and leishmanicidal temporin-sh paralog from the Sahara frog (Pelophylax saharicus). Int. J. Mol. Sci. 2020, 21, 6713. [Google Scholar] [CrossRef]
- Souza, A.L.A.; Faria, R.X.; Calabrese, K.S.; Hardoim, D.J. Temporizin and Temporizin-1 Peptides as Novel Candidates for Eliminating Trypanosoma cruzi. PLoS ONE 2016, 11, e0157673. [Google Scholar] [CrossRef]
- Feder, R.; Dagan, A.; Mor, A. Structure-activity relationship study of antimicrobial dermaseptin S4 showing the consequences of peptide oligomerization on selective cytotoxicity. J. Biol. Chem. 2000, 275, 4230–4238. [Google Scholar] [CrossRef] [Green Version]
- Lequin, O.; Ladram, A.; Chabbert, L.; Bruston, F.; Convert, O.; Vanhoye, D.; Chassaing, G.; Nicolas, P.; Amiche, M. Dermaseptin S9, an R-Helical Antimicrobial Peptide with a Hydrophobic Core and Cationic Termini. Biochemistry 2006, 45, 468–480. [Google Scholar] [CrossRef]
- Chaves, R.X.; Quelemes, P.V.; Leite, L.M.; Aquino, D.S.A.; Amorim, L.V.; Rodrigues, K.A.F.; Campelo, Y.D.M.; Veras, L.M.; Bemquerer, M.P.; Ramos-Jesus, J.; et al. Antileishmanial and Immunomodulatory Effects of Dermaseptin-01, A Promising Peptide Against Leishmania amazonensis. Curr. Bioact. Compd. 2016, 13, 1. [Google Scholar] [CrossRef]
- Salay, L.C.; Nobre, T.M.; Colhone, M.C.; Zaniquelli, M.E.D.; Ciancaglini, P.; Stabeli, R.G.; Leite, J.R.S.A.; Zucolotto, V. Dermaseptin 01 as antimicrobial peptide with rich biotechnological potential: Study of peptide interaction with membranes containing Leishmania amazonensis lipid-rich extract and membrane models. J. Pept. Sci. 2011, 17, 700–707. [Google Scholar] [CrossRef]
- Yang, Z.; Zheng, J.; Chan, C.F.; Wong, I.L.K.; Heater, B.S.; Chow, L.M.C.; Lee, M.M.M.; Chan, M.K. Targeted delivery of antimicrobial peptide by Cry protein crystal to treat intramacrophage infection. Biomaterials 2019, 217, 119286. [Google Scholar] [CrossRef]
- Nascimento, C.; Miura, L.M.C.V.; Nakano, E.; Kawano, T.; Sa, U.D.; Ministro, C.; Velloso, R. Evaluation of the in vitro Activity of Dermaseptin 01, a Cationic Antimicrobial Peptide, against Schistosoma mansoni. Chem. Biodivers. 2011, 8, 548–558. [Google Scholar]
- Cruzi, A.; Brand, G.D.; Leite, J.R.S.A.; Silva, L.P.; Albuquerque, S.; Prates, V.; Azevedo, R.B.; Silva, J.S.; Sá, V.C.L.; Brandão, R.A.; et al. Protein Structure and Folding: Dermaseptins from Phyllomedusa oreades and Phyllomedusa distincta: ACTIVITY without Cytotoxicity Dermaseptins from Phyllomedusa oreades and Phyllomedusa distincta. J. Biol.Chem. 2002, 277, 49332–49340. [Google Scholar] [CrossRef] [Green Version]
- Leite, S.A.; Silva, L.P.; Izabel, M.; Rodrigues, S.; Prates, M.V.; Brand, G.D.; Lacava, B.M.; Azevedo, R.B.; Bocca, L.; Albuquerque, S.; et al. Phylloseptins: A novel class of anti-bacterial and anti-protozoan peptides from the Phyllomedusa genus. Peptides 2005, 26, 565–573. [Google Scholar] [CrossRef]
- Sabiá, E.F.; Menezes, L.F.S.; De Araújo, I.F.S.; Schwartz, E.F. Natural occurrence in venomous arthropods of antimicrobial peptides active against protozoan parasites. Toxins 2019, 11, 563. [Google Scholar] [CrossRef] [Green Version]
- Rangel, M.; Dos Santas Cabrera, M.P.; Kazuma, K.; Ando, K.; Wang, X.; Kato, M.; Nihei, K.; Hirata, I.Y.; Cross, T.J.; Garcia, A.N.; et al. Chemical and biological characterization of four new linear cationic α-helical peptides from the venoms of two solitary eumenine wasps. Toxicon 2011, 57, 1081–1092. [Google Scholar] [CrossRef] [Green Version]
- Dale, B.A.; Fredericks, L.P. Antimicrobial peptides in the oral environment: Expression and function in health and disease. Curr. Issues Mol. Biol. 2005, 7, 119–133. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Khurshid, Z.; Najeeb, S.; Mali, M.; Moin, S.F.; Raza, S.Q.; Zohaib, S.; Sefat, F.; Zafar, M.S. Histatin peptides: Pharmacological functions and their applications in dentistry. Saudi Pharm. J. 2017, 25, 25–31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hajishengallis, G.; Russell, M.W. Innate Humoral Defense Factors. In Mucosal Immunology; Elsevier: Amsterdam, The Netherlands, 2015; pp. 251–270. [Google Scholar]
- Luque-Ortega, J.R.; Hof, W.; Veerman, E.C.I.; Saugar, J.M.; Rivas, L. Human antimicrobial peptide histatin 5 is a cell-penetrating peptide targeting mitochondrial ATP synthesis in Leishmania. FASEB J. 2008, 22, 1817–1828. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shahrour, H.; Ferrer-Espada, R.; Dandache, I.; Bárcena-Varela, S.; Sánchez-Gómez, S.; Chokr, A.; Martinez-de-Tejada, G. AMPs as Anti-biofilm Agents for Human Therapy and Prophylaxis. Adv. Exp. Med. Biol. 2019, 1117, 257–279. [Google Scholar] [CrossRef] [PubMed]


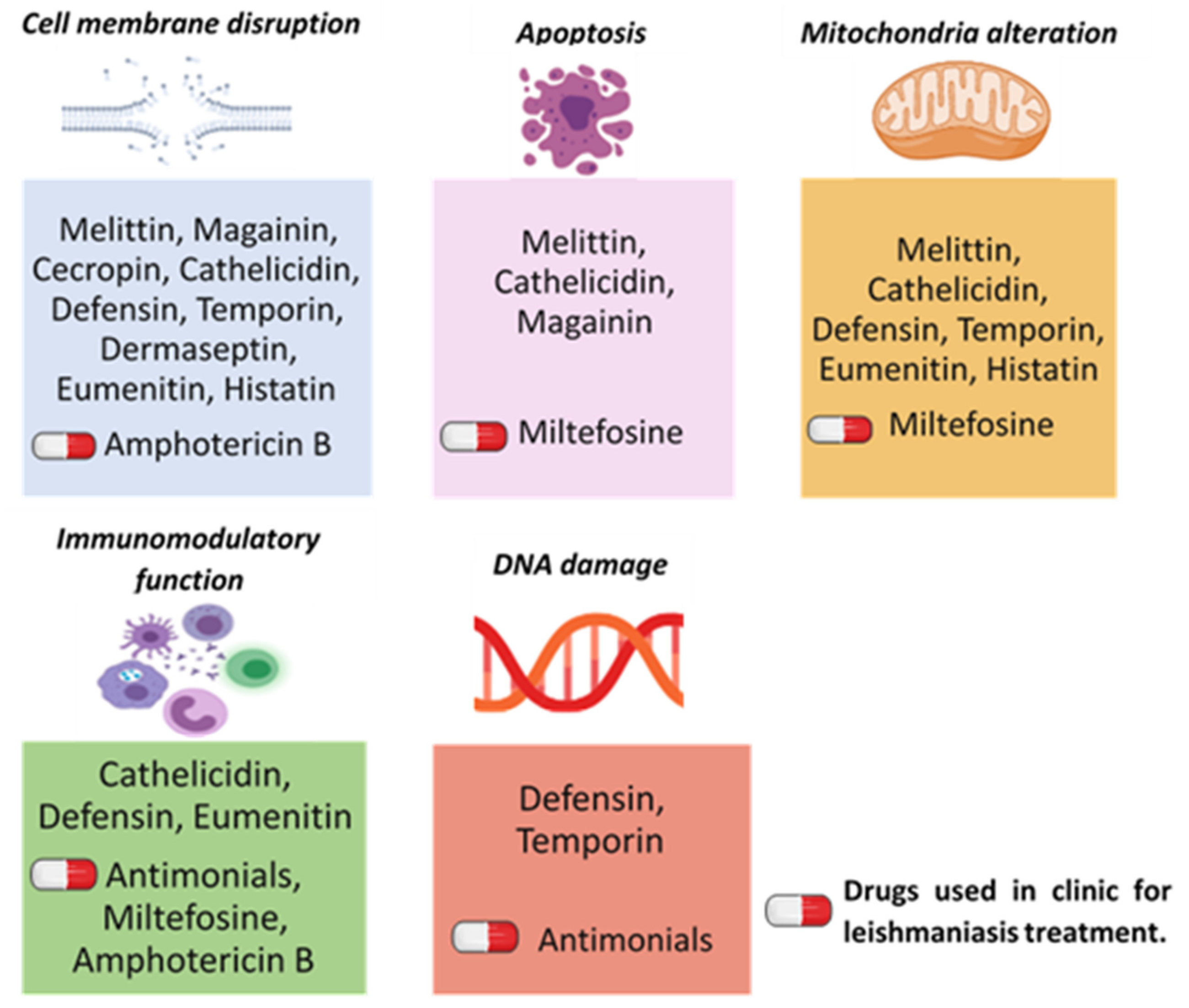{kind=link}
| AMPs Family | Peptide Name | Sequence | Leishmania species | Reference |
|---|---|---|---|---|
| Melittin | Melittin | GIGAVLTTGLPALISWIKRKRQQ | L. major | (Pereira et al., 2016) |
| L. panamensis | ||||
| L. donovani promastigotes | (Pérez-Cordero et al., 2011) | |||
| L. infantum promastigotes and amastigotes | (Díaz-Achirica et al., 1998) | |||
| Cecropin | Cecropin-A | KWKLFKKIEKVGQNIRDGIIKAGPAVAWVGQATQIAK | Leishmania aethiopic | (Pérez-Cordero et al., 2011) |
| L.panamensis amastigotes | (Akuffo et al., 1998) | |||
| Cecropin-D | ENFFKEIERAGQRIRDAIISAAPAVETLAQAQKIIKGGD | Leishmania (V) | (Patiño-Márquez et al., 2018) | |
| L. panamensis promastigotes | ||||
| Cathelicidin | LL-37 | LLGDFFRKSKEKIGKEFKRIVQRIKDFLRNLVPRTES | L. donovani promastigotes and amastigotes | (Marr et al., 2016) |
| L. major amastigotes | ||||
| RI-BMAP-28 | GIRIIPVIIPGYKKWARLIKRGLSRLGG | L. major promastigote | (Marr et al., 2016) | |
| (Lynn et al., 2011) | ||||
| D-BMAP-28 | GGLRSLGRKILRAWKKYGPIIVPIIRIG | L. major promastigote | (Marr et al., 2016) (Lynn et al., 2011) | |
| Defensin | MBD1 | MKTHYFLLVMICFLFSQMEPGVGILTSLGRRTDQYKCLQ HGGFCLRSSCPSNTKLQGTCKPDKPNCCK | L. major | (Daneshvar et al., 2018) |
| MBD2 | MRTLCSLLLICCLLFSYTTPAVGSLKSIGYEAELDHCHTN GGYCVRAICPPSARRPGSCFPEKNPCCKYMK | L. major | (Daneshvar et al., 2018) | |
| MBD3 | MRIHYLLFAFLLVLLSPPAAFSKKI--- NNPVSCLRKGGRCWNR-CIGNTRQIGSCGVPFLKCCKRK | L. major | (Daneshvar et al., 2018) | |
| Vu-Def | MKTCENLADTYRGP | L. amazonensis | (dos Santos et al., 2010) | |
| Magainin | MG-H1 | GIKKFLHIIWKFIKAFVGEIMNS | L. donovani promastigotes | (Guerrero et al., 2004) |
| MG-H2 | IIKKFLHSIWKFGKAFVGEIMNI | L. donovani promastigotes | (Guerrero et al., 2004) | |
| F5W-magainin 2 | GIGKWLHSAKKFGKAFVGEIMNS | L. donovani promastigotes | (Guerrero et al., 2004) | |
| Pexiganan | GIGKFLKKAKKFGKAFVKILKK | L. major | (Zhang et al., 2015) (Kulkarni et al., 2009) | |
| Temporin | Temporin A | FLPLIGRVLSGIL | L. donovani promastigotes | (Mangoni et al., 2005) |
| Temporin B | LLPIVGNLLKSLL | L. pifanoi amastigotes | (Mangoni et al., 2005) | |
| Temporin-She | FLPALAGIAGLLGKIF | L. braziliensis, L. major | (André et al., 2020) | |
| L. infantum | ||||
| SHd | FLPAALAGIGGILGKLF | L. infantum, L. major, L. tropica, L. amazonensis, and L. braziliensis promastigotes | (Mangoni et al., 2005) | |
| L. infantum axenic amastigotes | ||||
| Dermaseptin | DS 01 | GLWSTIKQKGKEAAIAAAKAAGQAALGAL | L. amazonensis promastigotes | (Salay et al., 2011) |
| Dermaseptin S1 | ALWKTMLKKLGTMALHAGKAALGAAADTISQGTQ | L. panamensis | (Pérez-Cordero et al., 2011) (Yang et al., 2019) | |
| L. mexicana promastigotes | ||||
| L. donovani | ||||
| L. amazonensis amastigotes | ||||
| Eumenitin | Eumenitin | LNLKGIFKKVASLLT | L. major Promastigote | (Sabiá et al., 2019) |
| Eumenitin-F | LNLKGLFKKVASLLT | L. major Promastigote | (Sabiá et al., 2019) | |
| Eumenitin R | LNLKGLIKKVASLLN | L. major Promastigote | (Sabiá et al., 2019) | |
| Histatin | Hst5 | DSHAKRHHGYKRKFHEKHHSHRGY | L. donovani promastigotes | (Luque-Ortega et al., 2008) |
| L. pifanoi axenic | ||||
| D- Hst5 | D- DSHAKRHHGYKRKFHEKHHSHRGY | L. donovani promastigotes | (Luque-Ortega et al., 2008) | |
| L. pifanoi axenic | ||||
| Dhvar4 | KRLFKKLLFSLRKY | L. donovani promastigotes | (Luque-Ortega et al., 2008) | |
| L. pifanoi axenic |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
El-Dirany, R.; Shahrour, H.; Dirany, Z.; Abdel-Sater, F.; Gonzalez-Gaitano, G.; Brandenburg, K.; Martinez de Tejada, G.; Nguewa, P.A. Activity of Anti-Microbial Peptides (AMPs) against Leishmania and Other Parasites: An Overview. Biomolecules 2021, 11, 984. https://doi.org/10.3390/biom11070984
El-Dirany R, Shahrour H, Dirany Z, Abdel-Sater F, Gonzalez-Gaitano G, Brandenburg K, Martinez de Tejada G, Nguewa PA. Activity of Anti-Microbial Peptides (AMPs) against Leishmania and Other Parasites: An Overview. Biomolecules. 2021; 11(7):984. https://doi.org/10.3390/biom11070984
Chicago/Turabian StyleEl-Dirany, Rima, Hawraa Shahrour, Zeinab Dirany, Fadi Abdel-Sater, Gustavo Gonzalez-Gaitano, Klaus Brandenburg, Guillermo Martinez de Tejada, and Paul A. Nguewa. 2021. "Activity of Anti-Microbial Peptides (AMPs) against Leishmania and Other Parasites: An Overview" Biomolecules 11, no. 7: 984. https://doi.org/10.3390/biom11070984