
Resveratrol Production in Yeast Hosts: Current Status and Perspectives



1
Key Laboratory of Molecular Biophysics of the Ministry of Education, College of Life Science and Technology, Huazhong University of Science and Technology, Wuhan 430074, China
2
Department of Genetics, Faculty of Agriculture, Zagazig University, Zagazig 44511, Egypt
*
Author to whom correspondence should be addressed.
Biomolecules 2021, 11(6), 830; https://doi.org/10.3390/biom11060830
Submission received: 28 April 2021
/
Revised: 29 May 2021
/
Accepted: 30 May 2021
/
Published: 2 June 2021
(This article belongs to the Topic Compounds with Medicinal Value)
Abstract
:Resveratrol is a plant secondary metabolite known for its therapeutic applications as an antioxidant, anti-cancer, anti-inflammatory, anti-aging, cardio-protective, and neuroprotective agent. Topical formulas of resveratrol are also used for skin disease management and in cosmetic industries. Due to its importance, high resveratrol production is urgently required. Since the last decade, intensive efforts have been devoted to obtaining resveratrol from microorganisms by pathway and metabolic engineering. Yeasts were proven to be excellent host candidates for resveratrol production. In addition to the similar intracellular compartments between yeasts and plants, yeasts exhibit the ability to express genes coding for plant-derived enzymes and to perform post-translational modification. Therefore, this review summarizes the attempts to use yeasts as a platform for resveratrol synthesis as the next promising route in producing high titers of resveratrol from genetically engineered strains.
1. Introduction
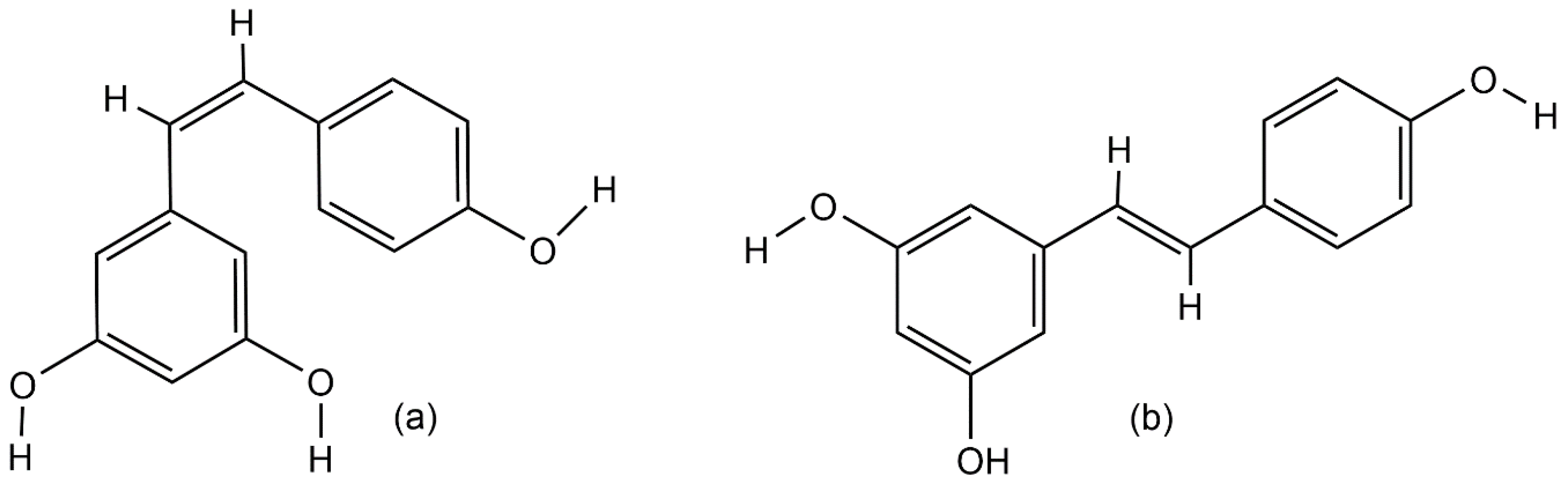
Resveratrol (3,4′,5-trihydroxystilbene) is a natural polyphenolic phytoalexin that belongs to the stilbenoids group of secondary metabolites [1,2,3]. Stilbenes are known for their ability to protect plants from UV light and the effect of chemical fertilizers [4], and for the defense of plants against biotic stresses such as bacterial, fungal, or nematode infections [3,5]. Resveratrol was first identified in 1940 as a constitutive compound of white hellebore roots (Veratrum grandiflorum) [6]. The richest source of resveratrol was found in Polygonum cuspidatum roots, the extract of which is widely used in traditional Chinese and Japanese medicine to treat human fungal diseases, such as gonococcal infection, suppurative dermatitis, tinea favosa and tinea pedis infections, hyperlipidemia, arteriosclerosis, and inflammations [1,7]. Thus far, resveratrol has been found in various plant species, such as grape berries [8], blueberries, cranberries (Vaccinium spp.) [9,10], blackberries, mulberries (Rubus and Morus spp.) [11,12], peanuts (Arachis hypogaea) [13], and jackfruit (Artocarpus heterophyllus) [11]. Naturally, there are two isomeric forms of resveratrol (Figure 1) but the trans isomer is the biologically active form [4]. However, under high pH values or UV-light exposure, resveratrol converts from the trans isoform into the cis isomer [14]. Both isomers exist as glucosides, and 3-O-β-d-resveratrol glucoside (piceid or polydatin) is the richest source of resveratrol in the glucoside form [1,14,15].
Resveratrol was found to be the reason for the well-known French Paradox, assuming that, despite the high consumption of saturated fats, French people show lower coronary heart disease incidence than other communities. This phenomenon can be explained by the consumption of red wine, which is a source of resveratrol [16,17].
Resveratrol has been widely studied in the past three decades [18,19] due to the accumulated evidence of its therapeutic properties. Resveratrol exhibits biological activities as a cardioprotective [20], anti-cancer [21,22,23,24], and possible anti-inflammatory [25] agent. Additional clinical evidence points to resveratrol’s role in cardiovascular disorder treatment [26], its impact on cell immunomodulation [27], and its role in lowering blood pressure [28]. Resveratrol has been suggested to have protective effects on neurodegenerative diseases, such as Alzheimer’s and Parkinson’s diseases [29,30,31,32]. Furthermore, resveratrol exhibits a positive role in treating periodontitis-related tissue defects and increased bone formation in mice morals [33].
In recent years, resveratrol’s topical formulations have been used in cosmetic skincare products [34] and skin disease management. Topical formulations of resveratrol have been reported to be valuable in treating skin diseases, such as acne, eczema, exfoliation, and psoriasis [35,36], and as a potent whitening agent [1]. Resveratrol’s antioxidant, antimicrobial, and antiviral effects provide skin protection from infections and ultraviolet-radiation-mediated oxidative stress (resulting in skin cancer and actinic keratosis) and show anti-aging properties [16,37]. Resveratrol was also found to accelerate skin wound healing [38].
Similar to glucosylated and methylated resveratrol derivatives, resveratrol oligomers present beneficial biological activities. Resveratrol-3-O-d-glycoside (piceid) shows antioxidant, anti-inflammatory, antitumor, hepatoprotective, and neuroprotective effects [39]. Many methylated derivatives, such as pterostilbene, trimethoxystilbene, and DMU212, possess more desirable pharmacokinetic properties than resveratrol and exhibit cardioprotective [40], anti-cancer [41,42,43], and neuroprotective activities [44,45]. Similarly, resveratrol oligomers such as pallidol [46,47], ε-viniferin [46,48,49,50], and labruscol [51] exhibit cytotoxic effects on cancer cell lines. Some isolated polymerized stilbenes, such as hopeaphenol, were found to inhibit the viability of cancer cells [52].
2. Resveratrol Biosynthesis in Nature
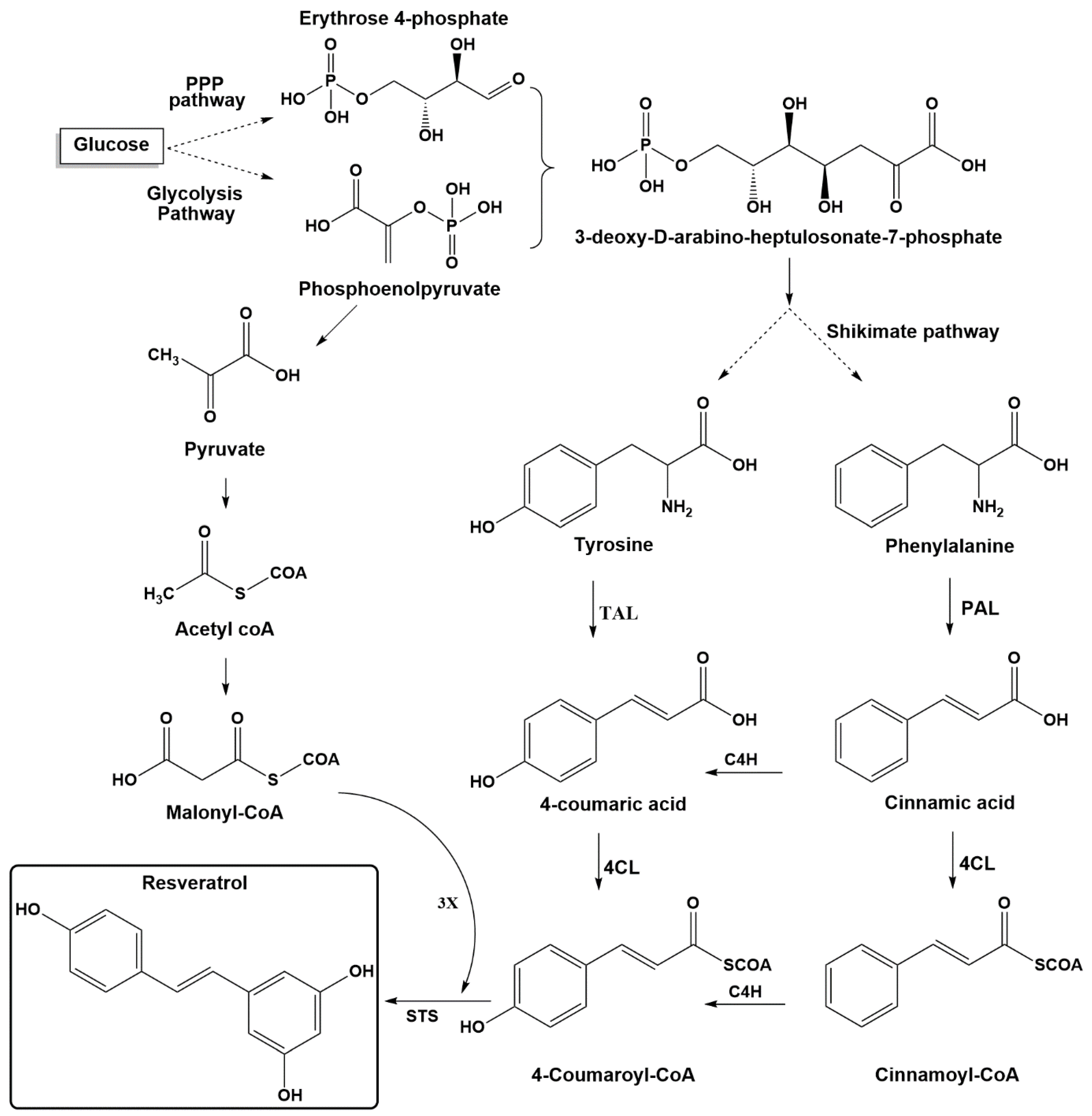
Stilbenes are plant secondary metabolites built from joining two aromatic rings with an ethylene bridge (1,2-diphenylethylene backbone) to form the basic C6-C2-C6 skeleton. Stilbenes share many similarities in their structure, biosynthesis, and biological activities with phenylpropanoids and flavonoids, which originate from the same pathway. The key components for biosynthesis of stilbenes are malonyl-CoA, phenylalanine, and tyrosine (Figure 2). In glycolysis pathway, glucose is converted into pyruvic acid, which leads to acetyl coenzyme A. The latter is the precursor of malonyl-CoA, which plays an essential role in biosynthesis pathways for many secondary plant compounds. The condensation of erythrose-4-phosphate (four carbons), which is generated from the pentose phosphate pathway (parallel reaction pathway to glycolysis) and phosphoenol pyruvate (three carbons, an intermediate in glycolysis), yields the seven-carbon shikimate skeleton. The latter compound is then used in the shikimic acid pathway to generate the aromatic amino acids phenylalanine and tyrosine [1,53].
The biosynthesis pathway for resveratrol starts with phenylalanine or tyrosine, as shown in Figure 2. The first step is to obtain para-coumaric acid from both pathways. The phenylalanine ammonia-lyase (PAL) transforms phenylalanine into cinnamic acid, which is further processed to generate 4-coumaric acid (para-coumaric acid) using cinnamate-4-hydroxylase (C4H). Tyrosine ammonia-lyase (TAL) can directly generate p-coumaric acid from tyrosine. The second step includes transformation of para-coumaric acid into para-coumaroyl-CoA by para-coumaroyl coenzyme A ligase (4CL). The final step involves the condensation of three units of malonyl-CoA with p-coumaroyl-CoA through stilbene synthase (STS) [1,19,54].
The STS genes are usually expressed when induced by biotic or abiotic stimuli [55]. Although STS genes are limited to a few plant families, the Cyperaceae, Dipterocarpaceae, Fabaceae, Gnetaceae, Pinaceae, and Vitaceae families have been reported to have a high degree of STS expression [56,57]. Sequencing of the genome of Vitis vinifera led to a broad diversification of the STS genes; at least 33 full-length coding genes were identified [58], which suggests complex regulation pathways, including the action of various transcription factors such as MYB and WRKY [59,60,61].
Starting from resveratrol, more modifications can be accomplished on its structure by various decorating enzymes. Many stilbene derivatives possess antioxidant and antifungal activities after methylation [62] or glucosylation of the aromatic hydroxyl groups [63]. Polyphenolic secondary metabolites can also be obtained from the oxidation of resveratrol by peroxidases and laccases, allowing for 2−8 resveratrol molecules to condensate [55].
3. Resveratrol Production by Transgenic Yeasts
Due to resveratrol’s pharmacological importance and its possible health and disease applications, resveratrol large-scale production has become a necessity. In nature, plants are the resource for resveratrol production; however, their production is limited by high costs; low quantities (the highest resveratrol concentration in plants was found in the seeds of Paeonia suffruticosa Andr. var. papaveracea (Andr.) Kerner with titers of 0.87 g/kg) [64]; lack of plant sources; and difficulties in the extraction, purification, and concentration processes. Transgenic plants and plant cell suspensions are two methods used for resveratrol production. Although engineered plants are usually restricted to enhancing the expression of STS genes and they have high productivity (up to 650 mg/kg FW of resveratrol), the use of elicitors, long production times, purity, and engineering process difficulties compared with microorganisms, remain as drawbacks for using this method [65]. Similar to transgenic plants, plant cell suspensions need elicitors to induce cells to produce resveratrol with resveratrol titer up to 5 g/L. The major limitation of this method is the requirement of light, which is not possible in large-scale production [66]. Although a high yield of resveratrol can be achieved by chemical synthesis, the production steps are complex, and the production of byproducts and the required toxic organic solvents are major drawbacks for large-scale preparations [67].
Microorganisms have been essential sources for producing pharmaceutically and industrially important compounds for decades due to low-cost cultures, fast production, the ability to construct purification processes, the ease of manipulating their genetic components, as well as the availability of protein and metabolic engineering tools [68]. Yeasts provide a more suitable platform for resveratrol production compared with bacterial hosts. As eukaryotic organisms, yeast species have the ability to express genes coding for plant-derived enzymes, such as cytochrome P450 enzymes; to perform post-translational modification (such as glycosylation); and to functionally fold the eukaryotic recombinant protein [69,70]. Additionally, yeasts and plants share similar intracellular compartments such as the endoplasmic reticulum, which supports eukaryotic and membrane proteins biosynthesis [71]. Another advantage of using yeasts is that Saccharomyces cerevisiae (the most-used yeast species for resveratrol production) is a food-grade organism that can be used safely in human nutrition and pharmaceutical products [72].
S.cerevisiae is a well-studied microorganism model for industrial and pharmaceutical applications. Metabolic engineering of S. cerevisiae was recognized as a robust strategy to produce several plant-derived chemicals [73]. Yarrowia lipolytica is another yeast that has received industrial interest for more than 50 years due to its organic acids production ability. Y. lipolytica is an oleaginous, non-pathogenic yeast that can accumulate lipids up to 40% of its dry cell weight (DCW) and is considered a model organism in diverse research areas [74,75]. Similar to S. cerevisiae, Y. lipolytica is classified as a generally recognized as safe organism (GRAS); therefore, it has the potential to be used in food and pharmaceutical industries. They are also considered powerful hosts for expressing heterologous genes [70,76]; consequently, both are strong candidates for use as cell factories for resveratrol production. Since microorganisms, including yeasts, cannot naturally produce resveratrol, genetic engineering of these strains by heterologous genes is required. Selecting genes to be transferred and the appropriate enzymes were found to markedly affect resveratrol production. Furthermore, pathway engineering, mutagenesis, codon optimization, protein engineering, and using synthetic scaffolds are all different strategies used to enhance resveratrol production capabilities in yeasts. Table 1 lists the genes used to engineer some yeast strains for resveratrol production.
3.1. Pathway Engineering
The introduction of an entire biosynthetic pathway into the microorganism provides the ability to produce resveratrol from its precursors (l-phenylalanine or l-tyrosine) or low-cost materials such as glycerol, glucose, or ethanol [19,89,98]. In this method, PAL or TAL, depending on the used pathway; C4H; 4CL; and STS encoding genes are transferred into the chosen host. One of the first attempts to entirely reconstruct the resveratrol pathway was conducted by Zhang et al. in 2006 in S. cerevisiae. TAL from Rhodobacter sphaeroides, in 4CL from Arabidopsis thaliana, and in STS from V. vinifera (4CL::STS fusion protein) were introduced into the S. cerevisiae strain WAT11. Although the TAL gene was not expressed in the yeast, after 20 h, 5.25 µg/mL of resveratrol was detected. The expression of the coupled genes coding for the fusion protein increased the resveratrol titer up to 15-fold compared with the co-expression of the genes encoding the separate enzymes, which emphasized the importance of the fusion protein and the spatial localization of these two related enzymes to improve resveratrol production [79]. Similarly, the same strain carrying TAL codon-optimized from R. sphaeroides and a similar fusion enzyme 4CL::STS were able to produce a resveratrol titer of 1.06 mg/L without the addition of l-tyrosine, and 1.90 mg/L with tyrosine. Again, the fused protein significantly increased resveratrol biosynthesis [83].
Production of resveratrol by the phenylalanine pathway was reported in S. cerevisiae YPH499. A strain harboring PAL from Populus trichocarpa, C4H and 4CL from Glycine max, and STS from V. vinifera produced resveratrol after being fed with phenylalanine, but the titer was still low (0.29 mg/L) [80]. The combination of both pathways was established in Y. lipolytica, and the final titer obtained was 1.46 mg/L [92]. In a recent study, Y. lipolytica was engineered using TAL from Flavobacterium johnsoniae, PAL and STS from V. vinifera, and C4H and 4CL1 from A. thaliana. The strains showed the ability to produce resveratrol using both pathways separately (using tyrosine or phenylalanine as precursors) or in combination. The importance of gene overexpression by increasing the gene copy number in the resveratrol pathway was clearly demonstrated in this study, in which strains harboring two copies of the PAL, C4H, 4CL1, and STS genes or two copies of TAL, 4CL1, and STS exhibited higher performances than single-copy sets of genes. The best results were obtained from a strain containing two copies of PAL, C4H, TAL, 4CL1, and STS, which produced a high titer of resveratrol, reaching 450 mg/L under fermentation conditions using 100 g/L glycerol as the sole carbon source, which is the highest reported amount of resveratrol produced from the expression of only the resveratrol pathway [94].
Engineering partial pathways or selected genes is an alternative strategy to produce resveratrol. In this case, PAL, TAL, and C4H are usually excluded from the pathway construction, and para-coumaric acid is used as the precursor. S. cerevisiae FY23 was the first used for resveratrol pathway construction in yeast. The 4CL216 from a hybrid poplar and vst1 from a grape vine were constitutively expressed in the strain. After feeding with 5 mM p-coumaric acid, the recombinant strain produced 1.45 µg/L resveratrol [77]. Higher resveratrol titers were then obtained from S. cerevisiae strains CEN-PK113-3B and EC1118, which harbor Nt4CL2 and At4CL2, respectively, and VvSTS. The two strains using p-coumaric acid as a precursor produced 5.8 and 8.2 mg/L resveratrol, respectively, as a final product [78,87].
Using synthetic scaffolds is another strategy used for improving resveratrol production. Nine different constructions containing GTPase binding domain (GBD), Src homology 3 domain (SH3) with 4CL1 from A. thaliana, and PSD95/DlgA/Zo-1 domain (PDZ) with STS from V. vinifera were recruited and optimized in S. cerevisiae WAT11 cells. Resveratrol production in the transformed yeast cells containing the optimal scaffold (GBD1SH32PDZ4) showed a five-fold increase in the production after 36 h (6.7 mg/L) and more than a two-fold rise in the resveratrol titer at 96 h after induction (14.4 mg/L). Using this scaffold strategy increased the resveratrol titer by 2.7-fold compared with the fusion enzyme strategy for the same genes [79,85]. This indicates the effectiveness of protein scaffolds in improving resveratrol synthesis and in increasing pathway enzyme activity compared with the protein fusions strategy.
The importance of selecting pathway genes and codon-optimizing them was explored in a recent report, in which six STS genes from different sources (PhStS, PcPKS5, MaSTS3, RtSTS, VvVST1, and AhSTS) were codon-optimized and then co-expressed with 4CL from Plagiochasma appendiculatum in S. cerevisiae W303. Differences in production time for the final yield were observed among the different strains. Although the lines expressing VvVST1, AhSTS, and RtSTS genes produced resveratrol quickly, their final titer was rather low (27–30 mg/L). Conversely, strains harboring STS from Morusalba and P. cuspidatum accumulated resveratrol up to 39 mg/L using 70 mg/L p-coumaric acid [86]. Codon optimization for resveratrol pathway genes has also been observed in several studies in both S. cerevisiae and Y. lipolytica [88,91,93,95,96]. The highest resveratrol titer, which was produced by the expression of partial pathway genes (4CL and STS) in yeasts, was obtained from industrial Brazilian sugar-cane-fermenting yeast, and the titer was 391 mg/L of resveratrol [81].
3.2. Host Metabolic Engineering (Non-Pathway Genes)
Metabolic engineering of microbial hosts for resveratrol production has achieved significant progress in recent years. The main remaining obstacles for microbial production of resveratrol are the precursor availability and the low activity of stilbene synthase in heterologous hosts. Hence, the primary strategies for increasing productivity using microbial cell factories are increasing the precursor supply (aromatic amino acids and malonyl-CoA) via genetic manipulation of the strain and improving the activity of key enzymes via protein engineering.
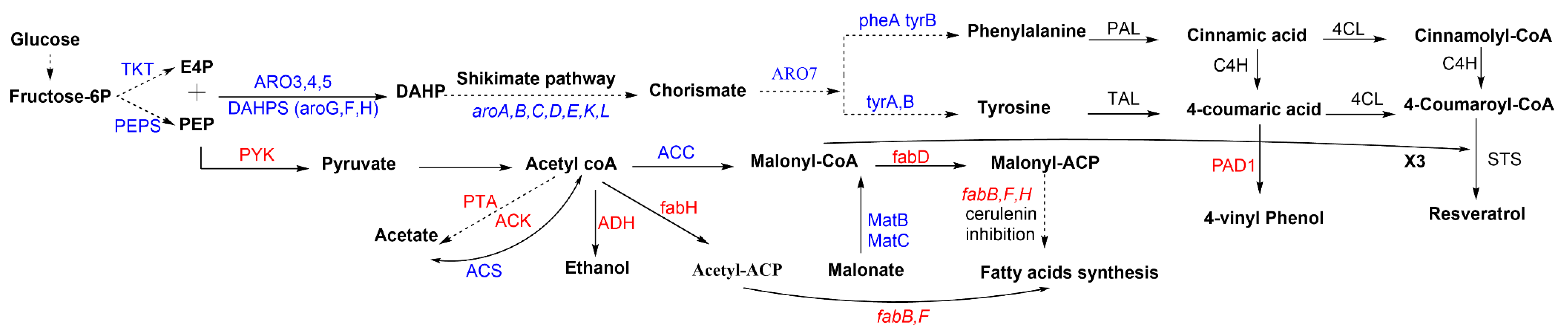
To increase the precursor levels, the introduction of non-pathway exogenous genes and pathway redirection are crucial. Notable efforts have been dedicated to optimizing the aromatic amino acids production and their derived phenylpropenoic acids in yeast [99,100]. Overproduction of aromatic amino acids can be easily achieved by engineering the shikimate pathway. Metabolic engineering of this pathway focused on improving carbon flux toward chorismate, and then Phe and Tyr (Figure 3). Extending the E4P and PEP supply and availability are considered fundamental approaches to enhancing chorismate production [73]. Intracellular malonyl-CoA is the other prime precursor involved in resveratrol biosynthesis. Naturally, the majority of malonyl-CoA is used in fatty acid biosynthesis, leaving a minimal level to be used in resveratrol biosynthesis. Thus, increasing this precursor in microbial hosts provides the opportunity for extra enhancement in resveratrol biosynthesis in the host platform.
Two main strategies are usually applied to enhance the malonyl-CoA pool. The first is to improve acetyl-CoA carboxylation into malonyl-CoA via the acetyl-CoA carboxylase (ACC) enzyme. The other strategy is to inhibit malonyl-CoA consumption by repressing the biosynthesis of fatty acids (Figure 3) [101]. Since the blocking of malonyl-CoA consumption and direct knockouts of the fab genes were found to be lethal to microorganisms [102], the inhibition has been accomplished in bacterial hosts via three procedures: the addition of cerulenin antibiotic to inhibit FabB and FabF [103,104]; using antisense RNA to repress the fab operon, especially the fabD genes [102,105]; and the CRISPRi tool, considered the third promising approach for repressing fab genes and directing carbon flux to malonyl-CoA [106,107]. Although these approaches have only been implemented in E. coli, they might provide valuable tools for future studies on increasing the precursors for resveratrol in yeast strains. S. cerevisiae is known to metabolize p-coumaric acid into 4-vinylphenol by phenyl acrylic acid decarboxylase (Pad1p). Although a PAD1 knockout mutation in S. cerevisiae W303-1A repressed the consumption of p-coumaric acid compared with the wild-type strain that uses about 60% of the p-coumaric acid, no enhancement was detected in resveratrol production (3.1 mg/L) when the PAD1 deletion mutant strain was transformed with a plasmid harboring At4CL1 and AhSTS [82]. In a separate study, the same strain (without PAD1 knockout mutation) was further transferred with RtPAL and AtC4H genes to produce 2.6 mg/L p-coumaric acid and 3.3 mg/L resveratrol, which indicates the minor effect of this strategy in increasing resveratrol production. The ACC1 gene was then overexpressed to increase the malonyl-CoA pool, which was reflected in the increase in resveratrol production to 4.3 mg/L without amino acids addition, and up to 5.8 mg/L with tyrosine added [84]. The overexpression of ACC1 was similarly reported in an E. coli-S. cerevisiae co-culture and Y. lipolytica for resveratrol production [91,93]. An increasing malonyl-CoA pool was also achieved by the overexpression of a plant malonyl-CoA synthetase (AAE13), resulting in a 2.4-fold increase and accumulation in resveratrol in S. cerevisiae [86].
Increasing the phenylalanine precursor in the prephenic acid pathway was achieved by overexpression of feedback-insensitive alleles encoding DAHP synthase (ScARO4K229L) and chorismate mutase (ScARO7G141S). Applying this strategy increased resveratrol production from 2.73 to 4.85 mg/L in S. cerevisiae with HaTAL, At4CL1, At4CL2, and VvVST1. The overexpression of the ScACC1S659A, S1157A gene raised the resveratrol titer to 6.39 mg/L, and another improvement was produced by the multiple-copies integration of pathways genes, which produced the highest titer of 235.57 mg/L, being 36-fold higher than in the last strain. Eventually, 415.65 and 531.41 mg/L of resveratrol were produced from the final strain under fed-batch fermentation with glucose or ethanol as the carbon source, respectively [88]. The same previous strategies were used with the phenylalanine pathway (AtPAL2, AtC4H, At4CL2, and VvVST1) with further metabolic engineering. Overexpression of cytochrome P450 reductase (AtATR2), S. cerevisiae cytochrome B5 (CYB5), and acetyl-CoA synthase (SeACSL641P) was used to increase the precursor supply, as well the deletion of phenylpyruvate decarboxylase (ARO10) to eliminate phenylalanine competing pathways. After this extensive engineering and cultivation in a fed-batch reactor, the strain was able to produce 812 and 755 mg/L of resveratrol from glucose and ethanol, respectively [89].
Y. lipolytica was proven to be a highly promising yeast for producing resveratrol. In Y. lipolytica ST6512, several metabolic strategies were used to finally obtain a strain that produced 12.4 g/L resveratrol, the highest reported resveratrol titer to date from de novo production. In addition to multiple integrations of resveratrol pathway genes (FjTAL, At4CL1, and VvVST1), feedback-insensitive YlARO4K221L and YlARO7G139S were employed to enhance resveratrol production. Further improvement was achieved by optimizing the growth medium and fed-batch fermentation [96].
In bacterial hosts, increased precursor amounts were additionally achieved by overexpression of different genes, such as malonyl-CoA synthetase (matB), malonate carrier protein (matC), tktA, PEP synthetase (ppsA), and acetate assimilation enzyme (acs), or by deleting or inhibiting competing pathway genes such as the pta, ackA, adhE, tyrR, trpR, and fab genes. These strategies might enhance resveratrol production, which requires further experiments on yeast platforms [73,103,104,108,109,110,111].
4. Conclusions
The increased demand for resveratrol for pharmaceutical and cosmetic uses necessitates its production from sustainable sources. Yeast cells are advantageous platforms for resveratrol production because of their fast production, the ease of their cultivation and purification processes, and their remarkable potential for large-scale resveratrol production. Due to their nature as eukaryotic cells and as GRAS organisms, the S. cerevisiae and Y. lipolytica platforms are preferable for resveratrol production. The successful design of a resveratrol production pathway depends on several factors, including determining the appropriate host, examining alternative specific enzymes from various sources, optimizing the codons to maximize the expression of heterologous genes, selecting a convenient transfer gene system (plasmids and their associated genetic elements), and determining favorable culturing parameters. Metabolic engineering has also achieved significant progress in increasing the resveratrol precursors, an important limitation in resveratrol production. Despite these efforts and extensive strain engineering, heterologous stilbene synthase activity and the overall resveratrol production are relatively still low. Except for one study that produced 12 g/L of resveratrol from Y. lipolytica [96], almost all engineered yeast strains do not produce more than 1 g/L, which does not meet the industrial needs and is the main drawback for their use on an industrial scale. Applying all of these elements together in a balanced way as well as testing other strategies that have been applied in E. coli may allow scientists to obtain the desired well-designed resveratrol-producing strain.
Author Contributions
G.G.I. drafted the original manuscript, Y.Y. and J.Y. revised and L.X. and M.Y. corrected the manuscript. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by funding by the National Natural Science Foundation of China (grant number: NSFC31570793), by the Fundamental Research Funds for the Central Universities (grant number: 2016YXMS255), and by the Startup Fund for Talent Scholars of Huazhong University of Science and Technology.
Acknowledgments
It was funded by the National Natural Science Foundation of China (Nos. 31971206 and 31170078).
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
| 4CL | 4-coumaroyl-coA ligase |
| AAE13 | malonyl-CoA synthetase |
| Acetyl-CoA | acetyl-coenzyme A |
| Acetyl-ACP | acetyl-acyl carrier protein |
| ACC | acetyl-CoA carboxylase |
| ACS | acetyl-CoA synthase |
| ACK | acetate kinase |
| ADH | alcohol dehydrogenases |
| araE | arabinose transporter |
| ARO1 | multifunctional AROM complex |
| ARO2 | chorismate synthase |
| ARO3/ARO4/ARO5 | 3-deoxy- d-arabinoheptulosonate-7-phosphate (DAHP) synthase |
| ARO7 | chorismate mutase |
| ARO8 | aromatic amino acid aminotransferase I |
| ARO9 | aromatic amino acid aminotransferase II |
| ARO10 | transaminated amino acid decarboxylase |
| aroA | gene that encodes the 3-phospho-shikimate-1-carboxyvinyltransferase protein |
| aroB | gene that encodes the dehydroquinate synthase protein |
| aroC | gene that encodes the chorismate synthase protein |
| aroD | gene that encodes the dehydroquinate dehydratase protein |
| aroE | gene that encodes the shikimate dehydrogenase protein |
| aroG/aroF/aroH | genes that encode the DAHP synthase |
| aroK/aroL | genes that encode the shikimate kinase isoenzymes I/II |
| ATR2 | NADPH-cytochrome P450 reductase 2 |
| C4H | cinnamate 4-hydroxylase |
| CPR | cytochrome P450 reductase |
| CYB5 | cytochrome b5 |
| DAHP | 3-deoxy-d arabinoheptulosonate 7-phosphate |
| DAHPS | 3-deoxy-d arabinoheptulosonate 7-phosphate (DAHP) synthase |
| E4P | erythrose 4-phosphate |
| fabB/fabF | genes that encode the beta-ketoacyl-acp synthase I/II protein |
| fabD | gene that encodes the malonyl-CoA-acyl carrier protein transacylase |
| fabH | gene that encodes 3-oxoacyl carrier protein synthase III |
| fbr | feedback resistant |
| GRAS | generally recognized as safe |
| Malonyl-CoA | malonyl-coenzyme A |
| Malonyl-ACP | malonyl-acyl carrier protein |
| MatB | malonyl-CoA synthetase |
| MatC | malonate carrier protein |
| PAD | phenyl acrylic acid decarboxylase |
| PAL | phenylalanine ammonia lyase |
| PEP | phosphoenolpyruvate |
| PEPS | phosphoenolpyruvate synthase |
| PEX10 | peroxisomal biogenesis factor 10 |
| PHA2 | prephenate dehydratase |
| PPP | pentose phosphate pathway |
| PTA | phosphate acetyltransferase |
| PYK | pyruvate kinase |
| STS | stilbene synthase |
| TAL | tyrosine ammonia-lyase |
| TKT | transketolase |
| TRP2 | anthranilate synthase |
| TRP3 | indole-3-glycerol-phosphate synthase |
| tyrA/pheA | genes that encode the chorismate mutase protein |
| TyrA | chorismate mutase/prephenate dehydrogenase |
| tyrB | gene that encodes the tyrosine aminotransferase |
| TyrR | transcriptional regulatory protein |
| VST/RS | resveratrol synthase |
| xfpK/xpkA | phosphoketolase. |
References
- Jeandet, P.; Sobarzo-Sánchez, E.; Silva, A.S.; Clément, C.; Nabavi, S.F.; Battino, M.; Rasekhian, M.; Belwal, T.; Habtemariam, S.; Koffas, M. Whole-cell biocatalytic, enzymatic and green chemistry methods for the production of resveratrol and its derivatives. Biotechnol. Adv. 2020, 39, 107461. [Google Scholar] [CrossRef] [PubMed]
- Gugleva, V.; Zasheva, S.; Hristova, M.; Andonova, V. Topical use of resveratrol: Technological aspects. Pharmacia 2020, 67, 89. [Google Scholar] [CrossRef]
- Jeandet, P.; Delaunois, B.; Conreux, A.; Donnez, D.; Nuzzo, V.; Cordelier, S.; Clément, C.; Courot, E. Biosynthesis, metabolism, molecular engineering, and biological functions of stilbene phytoalexins in plants. Biofactors 2010, 36, 331–341. [Google Scholar] [CrossRef] [PubMed]
- Rabesiaka, M.; Rakotondramasy-Rabesiaka, L.; Mabille, I.; Porte, C.; Havet, J.-L. Extraction of trans-resveratrol from red wine and optimization by response surface methodology. Sep. Purif. Technol. 2011, 81, 56–61. [Google Scholar] [CrossRef]
- Jeandet, P.; Douillet-Breuil, A.-C.; Bessis, R.; Debord, S.; Sbaghi, M.; Adrian, M. Phytoalexins from the Vitaceae: Biosynthesis, phytoalexin gene expression in transgenic plants, antifungal activity, and metabolism. J. Agric. Food Chem. 2002, 50, 2731–2741. [Google Scholar] [CrossRef]
- Takaoka, M. Of the phenolic substrate of hellebore (Veratrum grandiflorum Loes. fil.). J. Fac. Sci. Hokkaido Imper. Univ. 1940, 3, 1–16. [Google Scholar]
- Nonomura, S.; Kanagawa, H.; Makimoto, A. Chemical Constituents of Polygonaceous Plants. I. Studies on the Components of Ko-jo-kon. (Polygonum cuspidatum SIEB. et ZUCC.). Yakugaku Zasshi 1963, 83, 988–990. [Google Scholar] [CrossRef] [Green Version]
- Jeandet, P.; Bessis, R.; Gautheron, B. The production of resveratrol (3, 5, 4’-trihydroxystilbene) by grape berries in different developmental stages. Am. J. Enol. Vitic. 1991, 42, 41–46. [Google Scholar]
- Lyons, M.M.; Yu, C.; Toma, R.; Cho, S.Y.; Reiboldt, W.; Lee, J.; van Breemen, R. Resveratrol in raw and baked blueberries and bilberries. J. Agric. Food Chem. 2003, 51, 5867–5870. [Google Scholar] [CrossRef]
- Wang, Y.; Catana, F.; Yang, Y.; Roderick, R.; van Breemen, R.B. An LC-MS method for analyzing total resveratrol in grape juice, cranberry juice, and in wine. J. Agric. Food Chem. 2002, 50, 431–435. [Google Scholar] [CrossRef]
- Shrikanta, A.; Kumar, A.; Govindaswamy, V. Resveratrol content and antioxidant properties of underutilized fruits. J. Food Sci. Technol. 2015, 52, 383–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mudnic, I.; Budimir, D.; Modun, D.; Gunjaca, G.; Generalic, I.; Skroza, D.; Katalinic, V.; Ljubenkov, I.; Boban, M. Antioxidant and vasodilatory effects of blackberry and grape wines. J. Med. Food 2012, 15, 315–321. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sales, J.M.; Resurreccion, A.V. Resveratrol in peanuts. Crit. Rev. Food Sci. Nutr. 2014, 54, 734–770. [Google Scholar] [CrossRef] [PubMed]
- Pezzuto, J.M.; Moon, R.C.; Jang, M.-S.; Ouali, A.; Lin, S.; Barillas, K.S. Pharmaceutical Formulations of Resveratrol and Methods of Use Thereof. U.S. Patent US6414037B1, 2 July 2002. [Google Scholar]
- Ratz-Łyko, A.; Arct, J. Resveratrol as an active ingredient for cosmetic and dermatological applications: A review. J. Cosmet. Laser Ther. 2019, 21, 84–90. [Google Scholar] [CrossRef] [PubMed]
- Singh, G.; Pai, R.S. Recent advances of resveratrol in nanostructured based delivery systems and in the management of HIV/AIDS. J. Control. Release 2014, 194, 178–188. [Google Scholar] [CrossRef]
- Ndiaye, M.; Philippe, C.; Mukhtar, H.; Ahmad, N. The grape antioxidant resveratrol for skin disorders: Promise, prospects, and challenges. Arch. Biochem. Biophys. 2011, 508, 164–170. [Google Scholar] [CrossRef] [Green Version]
- Pangeni, R.; Sahni, J.K.; Ali, J.; Sharma, S.; Baboota, S. Resveratrol: Review on therapeutic potential and recent advances in drug delivery. Expert Opin. Drug Deliv. 2014, 11, 1285–1298. [Google Scholar] [CrossRef]
- Jeandet, P.; Delaunois, B.; Aziz, A.; Donnez, D.; Vasserot, Y.; Cordelier, S.; Courot, E. Metabolic engineering of yeast and plants for the production of the biologically active hydroxystilbene, resveratrol. J. Biomed. Biotechnol. 2012, 2012, 1–14. [Google Scholar] [CrossRef]
- Voloshyna, I.; Hussaini, S.M.; Reiss, A.B. Resveratrol in cholesterol metabolism and atherosclerosis. J. Med. Food 2012, 15, 763–773. [Google Scholar] [CrossRef]
- Rauf, A.; Imran, M.; Butt, M.S.; Nadeem, M.; Peters, D.G.; Mubarak, M.S. Resveratrol as an anti-cancer agent: A review. Crit. Rev. Food Sci. Nutr. 2018, 58, 1428–1447. [Google Scholar] [CrossRef]
- Varoni, E.M.; Lo Faro, A.F.; Sharifi-Rad, J.; Iriti, M. Anticancer molecular mechanisms of resveratrol. Front. Nutr. 2016, 3, 8. [Google Scholar] [CrossRef] [Green Version]
- Cai, H.; Scott, E.; Kholghi, A.; Andreadi, C.; Rufini, A.; Karmokar, A.; Britton, R.G.; Horner-Glister, E.; Greaves, P.; Jawad, D.; et al. Cancer chemoprevention: Evidence of a nonlinear dose response for the protective effects of resveratrol in humans and mice. Sci. Transl. Med. 2015, 7, ra117–ra298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carter, L.G.; D’Orazio, J.A.; Pearson, K.J. Resveratrol and cancer: Focus on in vivo evidence. Endocr. Relat. Cancer 2014, 21, R209–R225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- de Sá Coutinho, D.; Pacheco, M.T.; Frozza, R.L.; Bernardi, A. Anti-inflammatory effects of resveratrol: Mechanistic insights. Int. J. Mol. Sci. 2018, 19, 1812. [Google Scholar] [CrossRef] [Green Version]
- Zordoky, B.N.M.; Robertson, I.M.; Dyck, J.R.B. Preclinical and clinical evidence for the role of resveratrol in the treatment of cardiovascular diseases. Biochim. Biophys. Acta Mol. Basis Dis. 2015, 1852, 1155–1177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Malaguarnera, L. Influence of resveratrol on the immune response. Nutrients 2019, 11, 946. [Google Scholar] [CrossRef] [Green Version]
- Prysyazhna, O.; Wolhuter, K.; Switzer, C.; Santos, C.; Yang, X.; Lynham, S.; Shah, A.M.; Eaton, P.; Burgoyne, J.R. Blood pressure lowering by the antioxidant resveratrol is counterintuitively mediated by oxidation of cGMP-dependent protein kinase. Circulation 2019, 140, 126–137. [Google Scholar] [CrossRef]
- Rao, Y.L.; Ganaraja, B.; Joy, T.; Pai, M.M.; Ullal, S.D.; Murlimanju, B.V. Neuroprotective effects of resveratrol in Alzheimer’s disease. Front. Biosci. 2020, 12, 139–149. [Google Scholar]
- Bastianetto, S.; Ménard, C.; Quirion, R. Neuroprotective action of resveratrol. Biochim. Biophys. Acta (BBA) Mol. Basis Dis. 2015, 1852, 1195–1201. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rege, S.D.; Geetha, T.; Griffin, G.D.; Broderick, T.L.; Babu, J.R. Neuroprotective effects of resveratrol in Alzheimer disease pathology. Front. Aging Neurosci. 2014, 6, 218. [Google Scholar] [CrossRef] [Green Version]
- Singh, N.; Agrawal, M.; Doré, S. Neuroprotective Properties and Mechanisms of Resveratrol in in Vitro and in Vivo Experimental Cerebral Stroke Models. ACS Chem. Neurosci. 2013, 4, 1151–1162. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Adhikari, N.; Prasad Aryal, Y.; Jung, J.-K.; Ha, J.-H.; Choi, S.-Y.; Kim, J.-Y.; Lee, T.-H.; Kim, S.-H.; Yamamoto, H.; Suh, J.-Y.; et al. Resveratrol enhances bone formation by modulating inflammation in the mouse periodontitis model. J. Periodontal Res. 2021, in press. [Google Scholar] [CrossRef]
- Jeandet, P.; Clément, C.; Tisserant, L.-P.; Crouzet, J.; Courot, É. Use of grapevine cell cultures for the production of phytostilbenes of cosmetic interest. Comptes Rendus Chim. 2016, 19, 1062–1070. [Google Scholar] [CrossRef] [Green Version]
- Tsai, M.-J.; Lu, I.J.; Fu, Y.-S.; Fang, Y.-P.; Huang, Y.-B.; Wu, P.-C. Nanocarriers enhance the transdermal bioavailability of resveratrol: In-vitro and in-vivo study. Colloids Surf. B Biointerfaces 2016, 148, 650–656. [Google Scholar] [CrossRef]
- Pelliccia, M.; Giannella, A.; Giannella, J. Use of Resveratrol for the Treatment of Exfoliative Eczema, Acne and Psoriasis. U.S. Patent US20010056071A1, 27 December 2001. [Google Scholar]
- Baxter, R.A. Anti-aging properties of resveratrol: Review and report of a potent new antioxidant skin care formulation. J. Cosmet. Dermatol. 2008, 7, 2–7. [Google Scholar] [CrossRef]
- Zhou, X.; Ruan, Q.; Ye, Z.; Chu, Z.; Xi, M.; Li, M.; Hu, W.; Guo, X.; Yao, P.; Xie, W. Resveratrol accelerates wound healing by attenuating oxidative stress-induced impairment of cell proliferation and migration. Burns 2021, 47, 133–139. [Google Scholar] [CrossRef] [PubMed]
- Du, Q.-H.; Peng, C.; Zhang, H. Polydatin: A review of pharmacology and pharmacokinetics. Pharm. Biol. 2013, 51, 1347–1354. [Google Scholar] [CrossRef]
- Lv, M.; Liu, K.; Fu, S.; Li, Z.; Yu, X. Pterostilbene attenuates the inflammatory reaction induced by ischemia/reperfusion in rat heart. Mol. Med. Rep. 2015, 11, 724–728. [Google Scholar] [CrossRef] [Green Version]
- Aldawsari, F.S.; Velázquez-Martínez, C.A. 3,4′,5-trans-Trimethoxystilbene; a natural analogue of resveratrol with enhanced anticancer potency. Investig. New Drugs 2015, 33, 775–786. [Google Scholar] [CrossRef]
- McCormack, D.E.; Mannal, P.; McDonald, D.; Tighe, S.; Hanson, J.; McFadden, D. Genomic analysis of pterostilbene predicts its antiproliferative effects against pancreatic cancer in vitro and in vivo. J. Gastrointest. Surg. 2012, 16, 1136–1143. [Google Scholar] [CrossRef] [Green Version]
- Sale, S.; Verschoyle, R.D.; Boocock, D.; Jones, D.; Wilsher, N.; Ruparelia, K.C.; Potter, G.A.; Farmer, P.B.; Steward, W.P.; Gescher, A.J. Pharmacokinetics in mice and growth-inhibitory properties of the putative cancer chemopreventive agent resveratrol and the synthetic analogue trans 3, 4, 5, 4′-tetramethoxystilbene. Br. J. Cancer 2004, 90, 736–744. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Chen, L.; Liu, X.; Li, X.; Cao, Y.; Bai, Y.; Qi, F. Pterostilbene inhibits amyloid-β-induced neuroinflammation in a microglia cell line by inactivating the NLRP3/caspase-1 inflammasome pathway. J. Cell. Biochem. 2018, 119, 7053–7062. [Google Scholar] [CrossRef]
- Li, Y.-R.; Li, S.; Lin, C.-C. Effect of resveratrol and pterostilbene on aging and longevity. BioFactors 2018, 44, 69–82. [Google Scholar] [CrossRef] [PubMed]
- Nivelle, L.; Hubert, J.; Courot, E.; Jeandet, P.; Aziz, A.; Nuzillard, J.-M.; Renault, J.-H.; Clément, C.; Martiny, L.; Delmas, D.; et al. Anti-cancer activity of resveratrol and derivatives produced by grapevine cell suspensions in a 14 L stirred bioreactor. Molecules 2017, 22, 474. [Google Scholar] [CrossRef] [PubMed]
- González-Sarrías, A.; Gromek, S.; Niesen, D.; Seeram, N.P.; Henry, G.E. Resveratrol Oligomers Isolated from Carex Species Inhibit Growth of Human Colon Tumorigenic Cells Mediated by Cell Cycle Arrest. J. Agric. Food Chem. 2011, 59, 8632–8638. [Google Scholar] [CrossRef] [PubMed]
- Barjot, C.; Tournaire, M.; Castagnino, C.; Vigor, C.; Vercauteren, J.; Rossi, J.-F. Evaluation of antitumor effects of two vine stalk oligomers of resveratrol on a panel of lymphoid and myeloid cell lines: Comparison with resveratrol. Life Sci. 2007, 81, 1565–1574. [Google Scholar] [CrossRef]
- Muhtadi; Hakim, E.H.; Juliawaty, L.D.; Syah, Y.M.; Achmad, S.A.; Latip, J.; Ghisalberti, E.L. Cytotoxic resveratrol oligomers from the tree bark of Dipterocarpus hasseltii. Fitoterapia 2006, 77, 550–555. [Google Scholar] [CrossRef]
- Chowdhury, S.A.; Kishino, K.; Satoh, R.; Hashimoto, K.; Kikuchi, H.; Nishikawa, H.; Shirataki, Y.; Sakagami, H. Tumor-specificity and apoptosis-inducing activity of stilbenes and flavonoids. Anticancer. Res. 2005, 25, 2055–2063. [Google Scholar]
- Nivelle, L.; Hubert, J.; Courot, E.; Borie, N.; Renault, J.-H.; Nuzillard, J.-M.; Harakat, D.; Clément, C.; Martiny, L.; Delmas, D.; et al. Cytotoxicity of labruscol, a new resveratrol dimer produced by grapevine cell suspensions, on human skin melanoma cancer cell line HT-144. Molecules 2017, 22, 1940. [Google Scholar] [CrossRef] [Green Version]
- Rohaiza, S.; Yaacob, W.; Din, L.; Nazlina, I. Cytotoxic oligostilbenes from Shorea hopeifolia. Afr. J. Pharm. Pharmacol. 2011, 5, 1272–1277. [Google Scholar] [CrossRef] [Green Version]
- Nabavi, S.M.; Šamec, D.; Tomczyk, M.; Milella, L.; Russo, D.; Habtemariam, S.; Suntar, I.; Rastrelli, L.; Daglia, M.; Xiao, J.; et al. Flavonoid biosynthetic pathways in plants: Versatile targets for metabolic engineering. Biotechnol. Adv. 2020, 38, 107316. [Google Scholar] [CrossRef] [PubMed]
- Langcake, P.; Pryce, R.J. The production of resveratrol and the viniferins by grapevines in response to ultraviolet irradiation. Phytochemistry 1977, 16, 1193–1196. [Google Scholar] [CrossRef]
- Keylor, M.H.; Matsuura, B.S.; Stephenson, C.R.J. Chemistry and biology of resveratrol-derived natural products. Chem. Rev. 2015, 115, 8976–9027. [Google Scholar] [CrossRef]
- Shen, T.; Wang, X.-N.; Lou, H.-X. Natural stilbenes: An overview. Nat. Prod. Rep. 2009, 26, 916–935. [Google Scholar] [CrossRef] [PubMed]
- Xiao, K.; Zhang, H.-J.; Xuan, L.-J.; Zhang, J.; Xu, Y.-M.; Bai, D.-L. Stilbenoids: Chemistry and bioactivities. In Studies in Natural Products Chemistry; Atta-ur-Rahman, Ed.; Elsevier: Amsterdam, The Netherlands, 2008; pp. 453–646. [Google Scholar]
- Vannozzi, A.; Dry, I.B.; Fasoli, M.; Zenoni, S.; Lucchin, M. Genome-wide analysis of the grapevine stilbene synthase multigenic family: Genomic organization and expression profiles upon biotic and abiotic stresses. BMC Plant Biol. 2012, 12, 130. [Google Scholar] [CrossRef]
- Jeandet, P.; Clément, C.; Cordelier, S. Regulation of resveratrol biosynthesis in grapevine: New approaches for disease resistance? J. Exp. Bot. 2019, 70, 375–378. [Google Scholar] [CrossRef]
- Vannozzi, A.; Wong, D.C.J.; Höll, J.; Hmmam, I.; Matus, J.T.; Bogs, J.; Ziegler, T.; Dry, I.; Barcaccia, G.; Lucchin, M. Combinatorial regulation of stilbene synthase genes by wrky and myb transcription factors in grapevine (Vitis vinifera L.). Plant Cell Physiol. 2018, 59, 1043–1059. [Google Scholar] [CrossRef]
- Höll, J.; Vannozzi, A.; Czemmel, S.; D’Onofrio, C.; Walker, A.R.; Rausch, T.; Lucchin, M.; Boss, P.K.; Dry, I.B.; Bogs, J. The R2R3-MYB transcription factors MYB14 and MYB15 regulate stilbene biosynthesis in Vitis vinifera. Plant Cell 2013, 25, 4135–4149. [Google Scholar] [CrossRef] [Green Version]
- Rimando, A.M.; Pan, Z.; Polashock, J.J.; Dayan, F.E.; Mizuno, C.S.; Snook, M.E.; Liu, C.-J.; Baerson, S.R. In planta production of the highly potent resveratrol analogue pterostilbene via stilbene synthase and O-methyltransferase co-expression. Plant Biotechnol. J. 2012, 10, 269–283. [Google Scholar] [CrossRef]
- Hall, D.; De Luca, V. Mesocarp localization of a bi-functional resveratrol/hydroxycinnamic acid glucosyltransferase of Concord grape (Vitis labrusca). Plant J. 2007, 49, 579–591. [Google Scholar] [CrossRef] [PubMed]
- Tian, B.; Liu, J. Resveratrol: A review of plant sources, synthesis, stability, modification and food application. J. Sci. Food Agric. 2020, 100, 1392–1404. [Google Scholar] [CrossRef]
- Delaunois, B.; Cordelier, S.; Conreux, A.; Clément, C.; Jeandet, P. Molecular engineering of resveratrol in plants. Plant Biotechnol. J. 2009, 7, 2–12. [Google Scholar] [CrossRef]
- Donnez, D.; Jeandet, P.; Clément, C.; Courot, E. Bioproduction of resveratrol and stilbene derivatives by plant cells and microorganisms. Trends Biotechnol. 2009, 27, 706–713. [Google Scholar] [CrossRef] [PubMed]
- Fan, E.; Zhang, K.; Zhu, M.; Wang, Q. Obtaining resveratrol: From chemical synthesis to biotechnological production. Mini-Rev. Org. Chem. 2010, 7, 272–281. [Google Scholar] [CrossRef]
- Sun, X.; Shen, X.; Jain, R.; Lin, Y.; Wang, J.; Sun, J.; Yan, Y.; Yuan, Q. Synthesis of chemicals by metabolic engineering of microbes. Chem. Soc. Rev. 2015, 44, 3760–3785. [Google Scholar] [CrossRef] [PubMed]
- Braga, A.; Ferreira, P.; Oliveira, J.; Rocha, I.; Faria, N. Heterologous production of resveratrol in bacterial hosts: Current status and perspectives. World J. Microbiol. Biotechnol. 2018, 34, 122. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Madzak, C. Yarrowia lipolytica: Recent achievements in heterologous protein expression and pathway engineering. Appl. Microbiol. Biotechnol. 2015, 99, 4559–4577. [Google Scholar] [CrossRef] [PubMed]
- Rainha, J.; Gomes, D.; Rodrigues, L.R.; Rodrigues, J.L. Synthetic biology approaches to engineer Saccharomyces cerevisiae towards the industrial production of valuable polyphenolic compounds. Life 2020, 10, 56. [Google Scholar] [CrossRef] [PubMed]
- Thapa, S.B.; Pandey, R.P.; Park, Y.I.; Sohng, J.K. Biotechnological advances in resveratrol production and its chemical diversity. Molecules 2019, 24, 2571. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wang, J.; Yang, Y.; Yan, Y. Bioproduction of Resveratrol. In Biotechnology of Natural Products; Schwab, W., Lange, B.M., Wüst, M., Eds.; Springer International Publishing: Cham, Switzerland, 2018; pp. 61–79. [Google Scholar]
- Groenewald, M.; Boekhout, T.; Neuvéglise, C.; Gaillardin, C.; Van Dijck, P.W.; Wyss, M. Yarrowia lipolytica: Safety assessment of an oleaginous yeast with a great industrial potential. Crit. Rev. Microbiol. 2014, 40, 187–206. [Google Scholar] [CrossRef]
- Beopoulos, A.; Chardot, T.; Nicaud, J.-M. Yarrowia lipolytica: A model and a tool to understand the mechanisms implicated in lipid accumulation. Biochimie 2009, 91, 692–696. [Google Scholar] [CrossRef] [PubMed]
- Madzak, C. Engineering Yarrowia lipolytica for use in biotechnological applications: A review of major achievements and recent innovations. Mol. Biotechnol. 2018, 60, 621–635. [Google Scholar] [CrossRef]
- Becker, J.V.; Armstrong, G.O.; Van Der Merwe, M.J.; Lambrechts, M.G.; Vivier, M.A.; Pretorius, I.S. Metabolic engineering of Saccharomyces cerevisiae for the synthesis of the wine-related antioxidant resveratrol. FEMS Yeast Res. 2003, 4, 79–85. [Google Scholar] [CrossRef] [Green Version]
- Beekwilder, J.; Wolswinkel, R.; Jonker, H.; Hall, R.; de Vos, C.H.R.; Bovy, A. Production of resveratrol in recombinant microorganisms. Appl. Environ. Microbiol. 2006, 72, 5670–5672. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Y.; Li, S.-Z.; Li, J.; Pan, X.; Cahoon, R.E.; Jaworski, J.G.; Wang, X.; Jez, J.M.; Chen, F.; Yu, O. Using unnatural protein fusions to engineer resveratrol biosynthesis in yeast and mammalian cells. J. Am. Chem. Soc. 2006, 128, 13030–13031. [Google Scholar] [CrossRef]
- Trantas, E.; Panopoulos, N.; Ververidis, F. Metabolic engineering of the complete pathway leading to heterologous biosynthesis of various flavonoids and stilbenoids in Saccharomyces cerevisiae. Metab. Eng. 2009, 11, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Sydor, T.; Schaffer, S.; Boles, E. Considerable increase in resveratrol production by recombinant industrial yeast strains with use of rich medium. Appl. Environ. Microbiol. 2010, 76, 3361–3363. [Google Scholar] [CrossRef] [Green Version]
- Shin, S.-Y.; Han, N.S.; Park, Y.-C.; Kim, M.-D.; Seo, J.-H. Production of resveratrol from p-coumaric acid in recombinant Saccharomyces cerevisiae expressing 4-coumarate: Coenzyme A ligase and stilbene synthase genes. Enzym. Microb. Technol. 2011, 48, 48–53. [Google Scholar] [CrossRef]
- Wang, Y.; Halls, C.; Zhang, J.; Matsuno, M.; Zhang, Y.; Yu, O. Stepwise increase of resveratrol biosynthesis in yeast Saccharomyces cerevisiae by metabolic engineering. Metab. Eng. 2011, 13, 455–463. [Google Scholar] [CrossRef] [PubMed]
- Shin, S.-Y.; Jung, S.-M.; Kim, M.-D.; Han, N.S.; Seo, J.-H. Production of resveratrol from tyrosine in metabolically engineered Saccharomyces cerevisiae. Enzym. Microb. Technol. 2012, 51, 211–216. [Google Scholar] [CrossRef]
- Wang, Y.; Yu, O. Synthetic scaffolds increased resveratrol biosynthesis in engineered yeast cells. J. Biotechnol. 2012, 157, 258–260. [Google Scholar] [CrossRef]
- Wang, Y.; Chen, H.; Yu, O. A plant malonyl-CoA synthetase enhances lipid content and polyketide yield in yeast cells. Appl. Microbiol. Biotechnol. 2014, 98, 5435–5447. [Google Scholar] [CrossRef]
- Sun, P.; Liang, J.-L.; Kang, L.-Z.; Huang, X.-Y.; Huang, J.-J.; Ye, Z.-W.; Guo, L.-Q.; Lin, J.-F. Increased resveratrol production in wines using engineered wine strains Saccharomyces cerevisiae EC1118 and relaxed antibiotic or auxotrophic selection. Biotechnol. Prog. 2015, 31, 650–655. [Google Scholar] [CrossRef]
- Li, M.; Kildegaard, K.R.; Chen, Y.; Rodriguez, A.; Borodina, I.; Nielsen, J. De novo production of resveratrol from glucose or ethanol by engineered Saccharomyces cerevisiae. Metab. Eng. 2015, 32, 1–11. [Google Scholar] [CrossRef]
- Li, M.; Schneider, K.; Kristensen, M.; Borodina, I.; Nielsen, J. Engineering yeast for high-level production of stilbenoid antioxidants. Sci. Rep. 2016, 6, 36827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Villa-Ruano, N.; Rivera, A.; Rubio-Rosas, E.; Landeta-Cortés, G.; Varela-Caselis, J.L.; Romero-Arenas, O. Comparative activity of six recombinant stilbene synthases in yeast for resveratrol production. Appl. Sci. 2020, 10, 4847. [Google Scholar] [CrossRef]
- Yuan, S.-F.; Yi, X.; Johnston, T.G.; Alper, H.S. De novo resveratrol production through modular engineering of an Escherichia coli–Saccharomyces cerevisiae co-culture. Microb. Cell Factories 2020, 19, 143. [Google Scholar] [CrossRef] [PubMed]
- Huang, L.L.; Xue, Z.; Zhu, Q.Q. Method for the Production of Resveratrol in a Recombinant Oleaginous Microorganism. U.S. Patent USOO7772444B2, 10 August 2010. [Google Scholar]
- Palmer, C.M.; Miller, K.K.; Nguyen, A.; Alper, H.S. Engineering 4-coumaroyl-CoA derived polyketide production in Yarrowia lipolytica through a β-oxidation mediated strategy. Metab. Eng. 2020, 57, 174–181. [Google Scholar] [CrossRef]
- He, Q.; Szczepańska, P.; Yuzbashev, T.; Lazar, Z.; Ledesma-Amaro, R. De novo production of resveratrol from glycerol by engineering different metabolic pathways in Yarrowia lipolytica. Metab. Eng. Commun. 2020, 11, e00146. [Google Scholar] [CrossRef] [PubMed]
- Gu, Y.; Ma, J.; Zhu, Y.; Ding, X.; Xu, P. Engineering Yarrowia lipolytica as a chassis for de novo synthesis of five aromatic-derived natural products and chemicals. ACS Synth. Biol. 2020, 9, 2096–2106. [Google Scholar] [CrossRef] [PubMed]
- Sáez-Sáez, J.; Wang, G.; Marella, E.R.; Sudarsan, S.; Cernuda Pastor, M.; Borodina, I. Engineering the oleaginous yeast Yarrowia lipolytica for high-level resveratrol production. Metab. Eng. 2020, 62, 51–61. [Google Scholar] [CrossRef]
- Wang, L.; Deng, A.; Zhang, Y.; Liu, S.; Liang, Y.; Bai, H.; Cui, D.; Qiu, Q.; Shang, X.; Yang, Z.; et al. Efficient CRISPR–Cas9 mediated multiplex genome editing in yeasts. Biotechnol. Biofuels 2018, 11, 277. [Google Scholar] [CrossRef]
- Chen, R.; Yang, S.; Zhang, L.; Zhou, Y.J. Advanced strategies for production of natural products in yeast. iScience 2020, 23, 100879. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jiang, H.; Wood, K.V.; Morgan, J.A. Metabolic engineering of the phenylpropanoid pathway in Saccharomyces cerevisiae. Appl. Environ. Microbiol. 2005, 71, 2962–2969. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodriguez, A.; Kildegaard, K.R.; Li, M.; Borodina, I.; Nielsen, J. Establishment of a yeast platform strain for production of p-coumaric acid through metabolic engineering of aromatic amino acid biosynthesis. Metab. Eng. 2015, 31, 181–188. [Google Scholar] [CrossRef] [Green Version]
- Shrestha, A.; Pandey, R.P.; Sohng, J.K. Biosynthesis of resveratrol and piceatannol in engineered microbial strains: Achievements and perspectives. Appl. Microbiol. Biotechnol. 2019, 103, 2959–2972. [Google Scholar] [CrossRef] [PubMed]
- Yang, Y.; Lin, Y.; Li, L.; Linhardt, R.J.; Yan, Y. Regulating malonyl-CoA metabolism via synthetic antisense RNAs for enhanced biosynthesis of natural products. Metab. Eng. 2015, 29, 217–226. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Shao, D.; Shi, J.; Huang, Q.; Yang, H.; Jin, M. Strategies for enhancing resveratrol production and the expression of pathway enzymes. Appl. Microbiol. Biotechnol. 2016, 100, 7407–7421. [Google Scholar] [CrossRef]
- Lim, C.G.; Fowler, Z.L.; Hueller, T.; Schaffer, S.; Koffas, M.A. High-yield resveratrol production in engineered Escherichia coli. Appl. Environ. Microbiol. 2011, 77, 3451–3460. [Google Scholar] [CrossRef] [Green Version]
- Wu, J.; Yu, O.; Du, G.; Zhou, J.; Chen, J. Fine-tuning of the fatty acid pathway by synthetic antisense RNA for enhanced (2S)-naringenin production from L-tyrosine in Escherichia coli. Appl. Environ. Microbiol. 2014, 80, 7283–7292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liang, J.-l.; Guo, L.-q.; Lin, J.-f.; He, Z.-q.; Cai, F.-j.; Chen, J.-f. A novel process for obtaining pinosylvin using combinatorial bioengineering in Escherichia coli. World J. Microbiol. Biotechnol. 2016, 32, 102. [Google Scholar] [CrossRef] [PubMed]
- Wu, J.; Du, G.; Chen, J.; Zhou, J. Enhancing flavonoid production by systematically tuning the central metabolic pathways based on a CRISPR interference system in Escherichia coli. Sci. Rep. 2015, 5, 1–14. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Na, D.; Yoo, S.M.; Chung, H.; Park, H.; Park, J.H.; Lee, S.Y. Metabolic engineering of Escherichia coli using synthetic small regulatory RNAs. Nat. Biotechnol. 2013, 31, 170–174. [Google Scholar] [CrossRef]
- Zha, W.; Rubin-Pitel, S.B.; Shao, Z.; Zhao, H. Improving cellular malonyl-CoA level in Escherichia coli via metabolic engineering. Metab. Eng. 2009, 11, 192–198. [Google Scholar] [CrossRef]
- Lütke-Eversloh, T.; Stephanopoulos, G. L-tyrosine production by deregulated strains of Escherichia coli. Appl. Microbiol. Biotechnol. 2007, 75, 103–110. [Google Scholar] [CrossRef] [PubMed]
- Bulter, T.; Bernstein, J.R.; Liao, J.C. A perspective of metabolic engineering strategies: Moving up the systems hierarchy. Biotechnol. Bioeng. 2003, 84, 815–821. [Google Scholar] [CrossRef] [PubMed]
Figure 1.
Chemical structures of resveratrol: (a) cis-resveratrol isomer and (b) trans-resveratrol isomer.
Figure 1.
Chemical structures of resveratrol: (a) cis-resveratrol isomer and (b) trans-resveratrol isomer.

Figure 2.
Resveratrol biosynthesis via the phenylalanine/tyrosine pathway. PPP: pentose phosphate pathway, TAL: tyrosine ammonia-lyase, PAL: phenylalanine ammonia lyase, C4H: cinnamate 4-hydroxylase, 4CL: 4-coumaroyl-coA ligase, STS: stilbene synthase.
Figure 2.
Resveratrol biosynthesis via the phenylalanine/tyrosine pathway. PPP: pentose phosphate pathway, TAL: tyrosine ammonia-lyase, PAL: phenylalanine ammonia lyase, C4H: cinnamate 4-hydroxylase, 4CL: 4-coumaroyl-coA ligase, STS: stilbene synthase.

Figure 3.
Genes and enzymes involved in the resveratrol synthesis pathway. Dotted arrows refer to multiple steps. Genes and enzymes in blue are targets for overexpression. Genes and enzymes in red are targets for knockout or inhibition. 4CL: 4-coumaroyl-coA ligase, ACC: acetyl-CoA carboxylase, Acetyl-CoA: acetyl-coenzyme A, Acetyl-ACP: acetyl-acyl carrier protein, ACS: acetyl-CoA synthase, ACK: acetate kinase, ADH: alcohol dehydrogenases, ARO3/ARO4/ARO5: 3-deoxy- d-arabinoheptulosonate-7-phosphate (DAHP) synthase, ARO7: chorismate mutase, aroA: gene that encodes the 3-phospho-shikimate-1-carboxyvinyltransferase protein, aroB: gene that encodes the dehydroquinate synthase protein, aroC: gene that encodes the chorismate synthase protein, aroD: gene that encodes the dehydroquinate dehydratase protein, aroE: gene that encodes the shikimate dehydrogenase protein, aroG/aroF/aroH: genes that encode the DAHP synthase, aroK/aroL: genes that encode the shikimate kinase isoenzymes I/II, C4H: cinnamate 4-hydroxylase, DAHP: 3-deoxy-d arabinoheptulosonate 7-phosphate, DAHPS: 3-deoxy-d arabinoheptulosonate 7-phosphate (DAHP) synthase, E4P: erythrose 4-phosphate, fabB/fabF: genes that encode the beta-ketoacyl-acp synthase I/II protein, fabD: gene that encodes the malonyl-CoA-acyl carrier protein transacylase, fabH: gene that encodes 3-oxoacyl carrier protein synthase III, fbr: feedback resistant, Malonyl-CoA: malonyl-coenzyme A, Malonyl-ACP: malonyl-acyl carrier protein, MatB: malonyl-CoA synthetase, MatC: malonate carrier protein, PAD: phenyl acrylic acid decarboxylase, PAL: phenylalanine ammonia lyase, PEP: phosphoenolpyruvate, PEPS: phosphoenolpyruvate synthase, PTA: phosphate acetyltransferase, PYK, pyruvate kinase, STS: stilbene synthase, TAL: tyrosine ammonia-lyase, TKT: transketolase, tyrA/pheA: genes that encode the chorismate mutase protein, tyrB: gene that encodes the tyrosine aminotransferase.
Figure 3.
Genes and enzymes involved in the resveratrol synthesis pathway. Dotted arrows refer to multiple steps. Genes and enzymes in blue are targets for overexpression. Genes and enzymes in red are targets for knockout or inhibition. 4CL: 4-coumaroyl-coA ligase, ACC: acetyl-CoA carboxylase, Acetyl-CoA: acetyl-coenzyme A, Acetyl-ACP: acetyl-acyl carrier protein, ACS: acetyl-CoA synthase, ACK: acetate kinase, ADH: alcohol dehydrogenases, ARO3/ARO4/ARO5: 3-deoxy- d-arabinoheptulosonate-7-phosphate (DAHP) synthase, ARO7: chorismate mutase, aroA: gene that encodes the 3-phospho-shikimate-1-carboxyvinyltransferase protein, aroB: gene that encodes the dehydroquinate synthase protein, aroC: gene that encodes the chorismate synthase protein, aroD: gene that encodes the dehydroquinate dehydratase protein, aroE: gene that encodes the shikimate dehydrogenase protein, aroG/aroF/aroH: genes that encode the DAHP synthase, aroK/aroL: genes that encode the shikimate kinase isoenzymes I/II, C4H: cinnamate 4-hydroxylase, DAHP: 3-deoxy-d arabinoheptulosonate 7-phosphate, DAHPS: 3-deoxy-d arabinoheptulosonate 7-phosphate (DAHP) synthase, E4P: erythrose 4-phosphate, fabB/fabF: genes that encode the beta-ketoacyl-acp synthase I/II protein, fabD: gene that encodes the malonyl-CoA-acyl carrier protein transacylase, fabH: gene that encodes 3-oxoacyl carrier protein synthase III, fbr: feedback resistant, Malonyl-CoA: malonyl-coenzyme A, Malonyl-ACP: malonyl-acyl carrier protein, MatB: malonyl-CoA synthetase, MatC: malonate carrier protein, PAD: phenyl acrylic acid decarboxylase, PAL: phenylalanine ammonia lyase, PEP: phosphoenolpyruvate, PEPS: phosphoenolpyruvate synthase, PTA: phosphate acetyltransferase, PYK, pyruvate kinase, STS: stilbene synthase, TAL: tyrosine ammonia-lyase, TKT: transketolase, tyrA/pheA: genes that encode the chorismate mutase protein, tyrB: gene that encodes the tyrosine aminotransferase.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Production of resveratrol in different yeast hosts in different engineered yeast strains, used genes and their sources, used genetic systems for engineering, precursors, the titers obtained, and the applied scale.
Table 1.
Production of resveratrol in different yeast hosts in different engineered yeast strains, used genes and their sources, used genetic systems for engineering, precursors, the titers obtained, and the applied scale.
| Yeast/Parent Strain | Pathway Genes (Source) | Pathway/Host Engineering | Genetic System | Precursor | Titer (mg/L) | Scale | Year of Publication | Reference |
|---|---|---|---|---|---|---|---|---|
| S. cerevisiae FY23 | 4CL216 (P. trichocarpa × P. deltoides) VTS1 (V. vinifera) | - | Episomal plasmid | p-Coumaric acid | 0.00145 | Flask | 2003 | [77] |
| S. cerevisiae CEN-PK113-3B | 4CL2 (N. tabacum) STS (V. vinifera) | - | One copy genome integration | p-Coumaric acid | 5.8 | Flask | 2006 | [78] |
| S. cerevisiae WAT11 | TAL (R. sphaeroides) 4CL (A. thaliana)::STS (V. vinifera) | - | Episomal plasmid | p-Coumaric acid | 5.25 | Flask | 2006 | [79] |
| S. cerevisiae YPH499 | PAL, CPRa (P. trichocarpa × P. deltoides) C4H, 4CL (G. max) STS (V. vinifera) | - | Episomal plasmid | Phenylalanine p-Coumaric acid | 0.29 0.31 | Flask | 2009 | [80] |
| Industrial Brazilian yeast (S. cerevisiae) | 4CL1 (A. thaliana) STS (V. vinifera) | - | Episomal plasmid | p-Coumaric acid | 262–391 | Flask | 2010 | [81] |
| S. cerevisiae W303-1A | 4CL1 (A. thaliana) STS (A. hypogaea) | PAD1 knockout | Episomal plasmid | p-Coumaric acid | 3.1 | Flask | 2011 | [82] |
| S. cerevisiae WAT11 | TAL (R. sphaeroides) 4CL::STS, 4CL1 (A. thaliana)-STS (V. vinifera) fusion enzyme | Expression of araE transporter (E. coli) | One copy genome integration | Tyrosine, p-Coumaric acid Grape Juice | 3.1 2.3 3.44 | Shake flask | 2011 | [83] |
| S. cerevisiae W303-1A | PAL (R. toruloides) C4H, 4CL1 (A. thaliana) STS (A. hypogaea) | Overexpression of ACC1 | Episomal plasmid | Tyrosine | 5.8 | Batch bioreactor | 2012 | [84] |
| S. cerevisiae WAT11 | 4CL1 (A. thaliana) STS (V. vinifera) | Synthetic scaffold | Episomal plasmid | p-Coumaric acid | 14.4 | Flask | 2012 | [85] |
| S. cerevisiae WAT11 | 4CL::STS, 4CL1 (A. thaliana)-STS (V. vinifera) fusion enzyme | Overexpression of: AAE13 | One copy genome integration | p-Coumaric acid | Up to 3.7 | Flask | 2014 | [86] |
| S. cerevisiae EC1118 | 4CL (A. thaliana) STS (V. vinifera) | - | Episomal plasmids | p-coumaric acid | 8.249 | Flask | 2015 | [87] |
| S. cerevisiae CEN. PK102-5B | TAL (H. aurantiacus) TAL (F. johnsoniae) 4CL1 and 4CL2 (A. thaliana) RS (V. vinifera) | Overexpression of ARO4fbr, ARO7fbr, and ACC1 | Multiple copy genome integration | Glucose Ethanol | 415.65 531.41 | Fed-batch bioreactor Fed-batch bioreactor | 2015 | [88] |
| S. cerevisiae CEN. PK102-5B | PAL2, C4H, 4CL2 (A. thaliana) VST1 (V. vinifera) | Overexpression of ARO4fbr, ARO7fbr, ACC1, CYB5 (S. cerevisiae), ATR2 (A. thaliana), ACS (S. enterica), and deletion of aro10 | Multiple-copy genome integration | Glucose Ethanol | 812 755 | Fed-batch bioreactor Fed-batch bioreactor | 2016 | [89] |
| S. cerevisiae W303 | 4CL1 (P. appendiculatum) STS (P. henryana) STS (P. cuspidatum) STS (M. alba var. atropurpurea) STS (R. tataricum) STS (V. vinifera) STS (A. hypogaea) One STS gene for each yeast line | - | Episomal plasmids | p-Coumaric acid | 23.7–39.9 | Batch bioreactor | 2020 | [90] |
| Co-culture of E. coli NEB10β and S. cerevisiae BY4741 | TAL (T. cutaneum) | Overexpression of aroG and tyrA in a tyrR knockout strain | Bacterial Expression Vectors | Glucose | 36 | Co-culture fermentation | 2020 | [91] |
| 4CL (A. thaliana) STS (V. vinifera) | Overexpression of: ACC1 | One copy genome integration | p-Coumaric acid (secreted from E. coli) | |||||
| Y. lipolytica ATCC 20362 | PAL/TAL (R.glutinis) 4CL (S. coelicolor) STS (V. vinifera) | - | l-tyrosine | 1.46 | 2010 | [92] | ||
| Y. lipolytica | 4CL (N. tabacum) STS (A. hypogaea) | Overexpression of: ACC1, PEX10 | Randomly genome integration | p-Coumaric acid | 48.7 | Flask | 2020 | [93] |
| Y. lipolytica Po1d (wt), derived from W29 | TAL (F. johnsoniae) PAL (V. vinifera) C4H, 4CL1 (A. thaliana) VST (V. vinifera) | - | Multiple copy genome integration | Glycerol | 430 | Bioreactor | 2020 | [94] |
| Y. lipolytica Po1fk derived from W29 | TAL (R. toruloides) 4CL (P. crispum) STS (V. vinifera) | ARO4fbr (S. cerevisiae) aroGfbr (E. coli) xfpK (B. breve) xpkA (A. capsulatum) Overexpression: of ARO1, ARO2, ARO3, ARO4, ARO5, TKT Deletion of: TRP2, TRP3, ARO8, ARO9, PYK, PHA2 | One copy genome integration | Glucose | 12.67 | Flask | 2020 | [95] |
| Y. lipolytica ST6512 (W29) | TAL (F. johnsoniae) 4CL1(A. thaliana) VST1 (V. vinifera) | Overexpression of: ARO4fbr and ARO7fbr | Multiple copy genome integration | Glucose Glucose | 409 12355 | Flask Fed-batch bioreactor | 2020 | [96] |
| Ogataea polymorpha | TAL (H. aurantiacus) 4CL (A. thaliana) STS (V. vinifera) | - | CRISPR–Cas9-assisted multiplex genome editing, multi-copy integration | Tyrosine | 97.23 | Flask | 2018 | [97] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ibrahim, G.G.; Yan, J.; Xu, L.; Yang, M.; Yan, Y. Resveratrol Production in Yeast Hosts: Current Status and Perspectives. Biomolecules 2021, 11, 830. https://doi.org/10.3390/biom11060830
AMA Style
Ibrahim GG, Yan J, Xu L, Yang M, Yan Y. Resveratrol Production in Yeast Hosts: Current Status and Perspectives. Biomolecules. 2021; 11(6):830. https://doi.org/10.3390/biom11060830
Chicago/Turabian StyleIbrahim, Gehad G., Jinyong Yan, Li Xu, Min Yang, and Yunjun Yan. 2021. "Resveratrol Production in Yeast Hosts: Current Status and Perspectives" Biomolecules 11, no. 6: 830. https://doi.org/10.3390/biom11060830
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.