
4-CPA (4-Chlorophenoxyacetic Acid) Induces the Formation and Development of Defective “Fenghou” (Vitis vinifera × V. labrusca) Grape Seeds

 , and
, and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Plant Material
2.2. Pharmacological Treatments
2.3. Organizational Structure Observation and Statistical Analysis
2.4. RNA Extraction and Quantitative Real-Time PCR Analysis
2.5. In Situ Hybridization
2.6. Statistical Analysis
3. Results
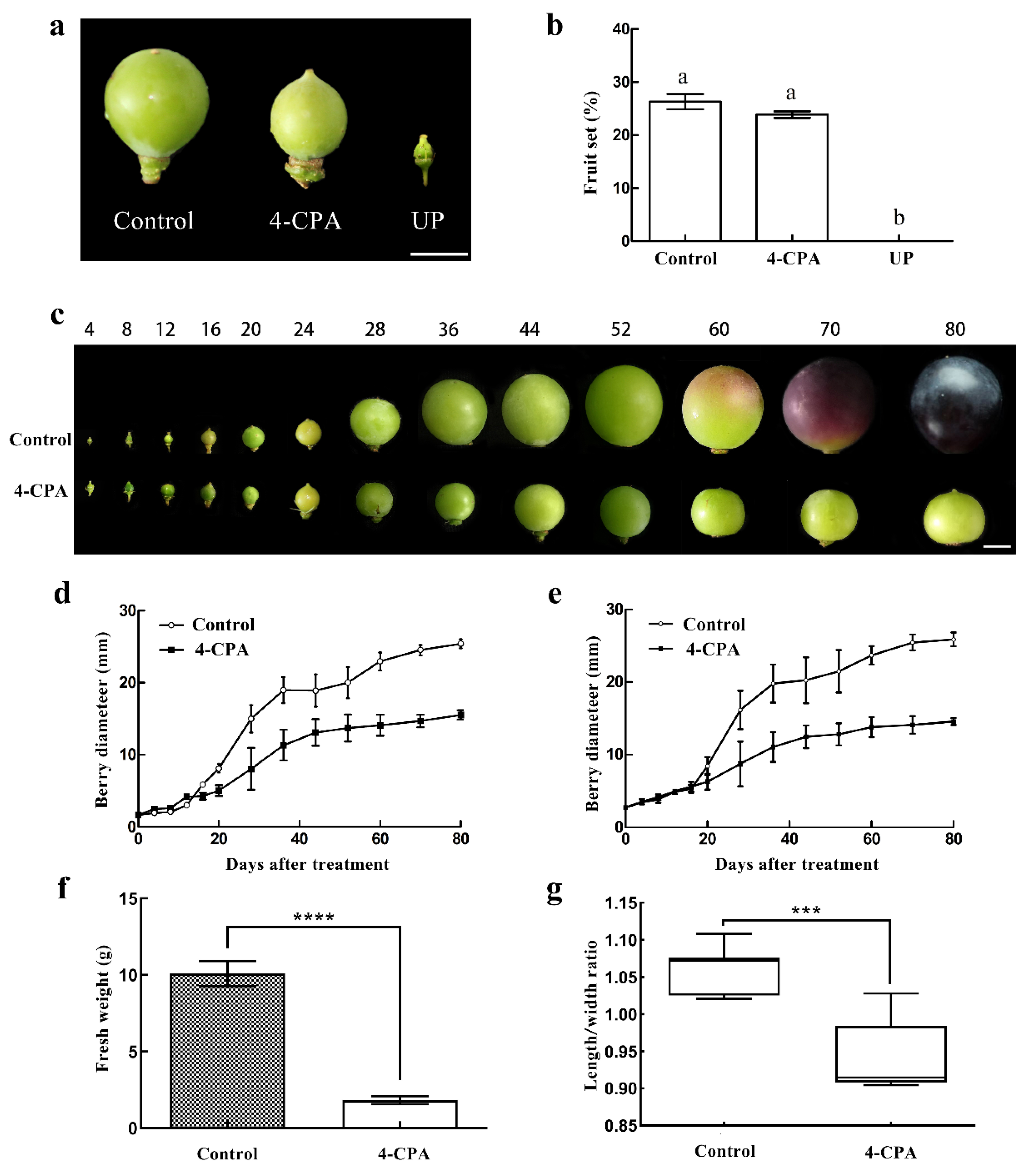
3.1. 4-CPA Inhibits Fruit Growth But Not Fruit Setting
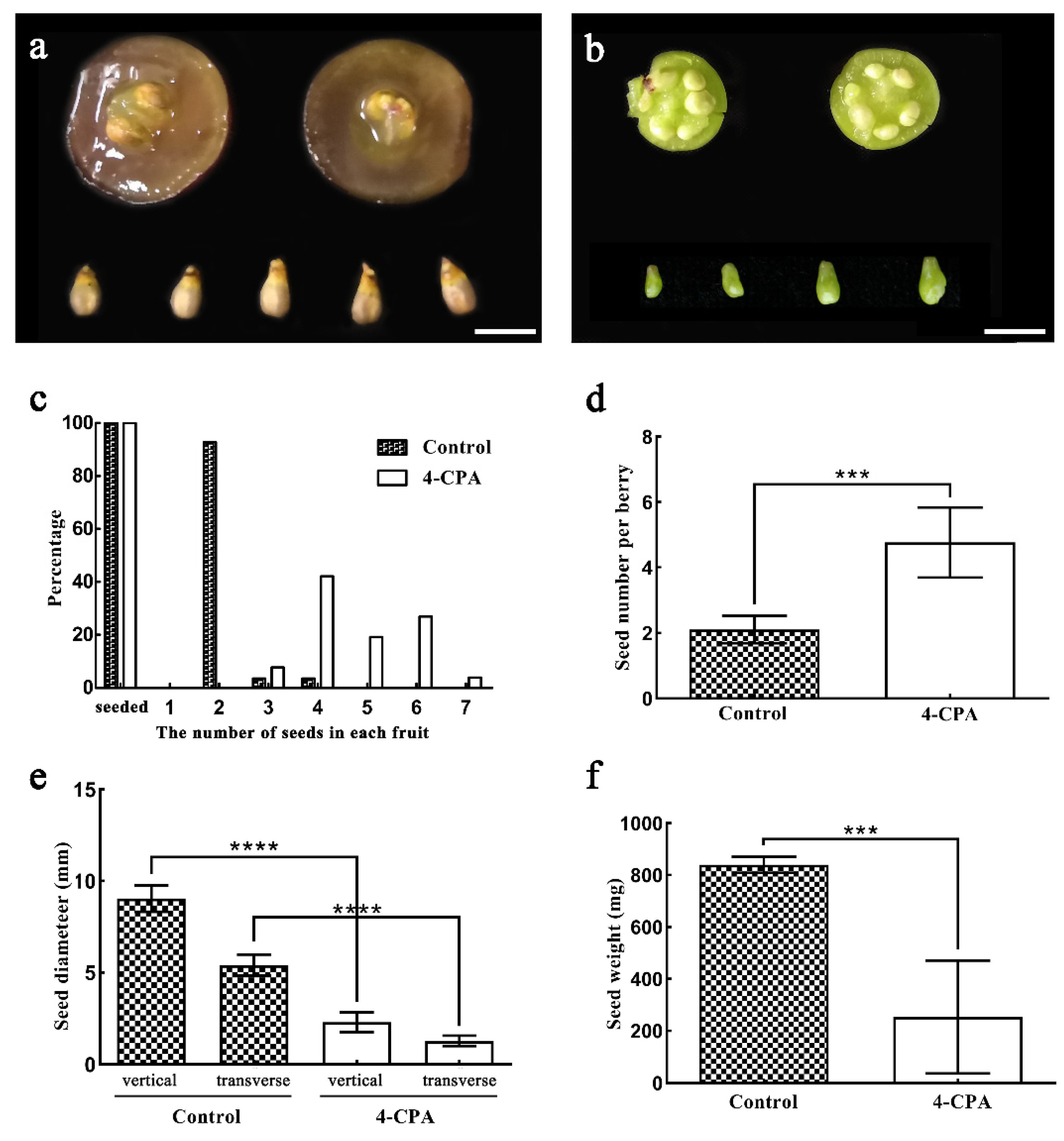
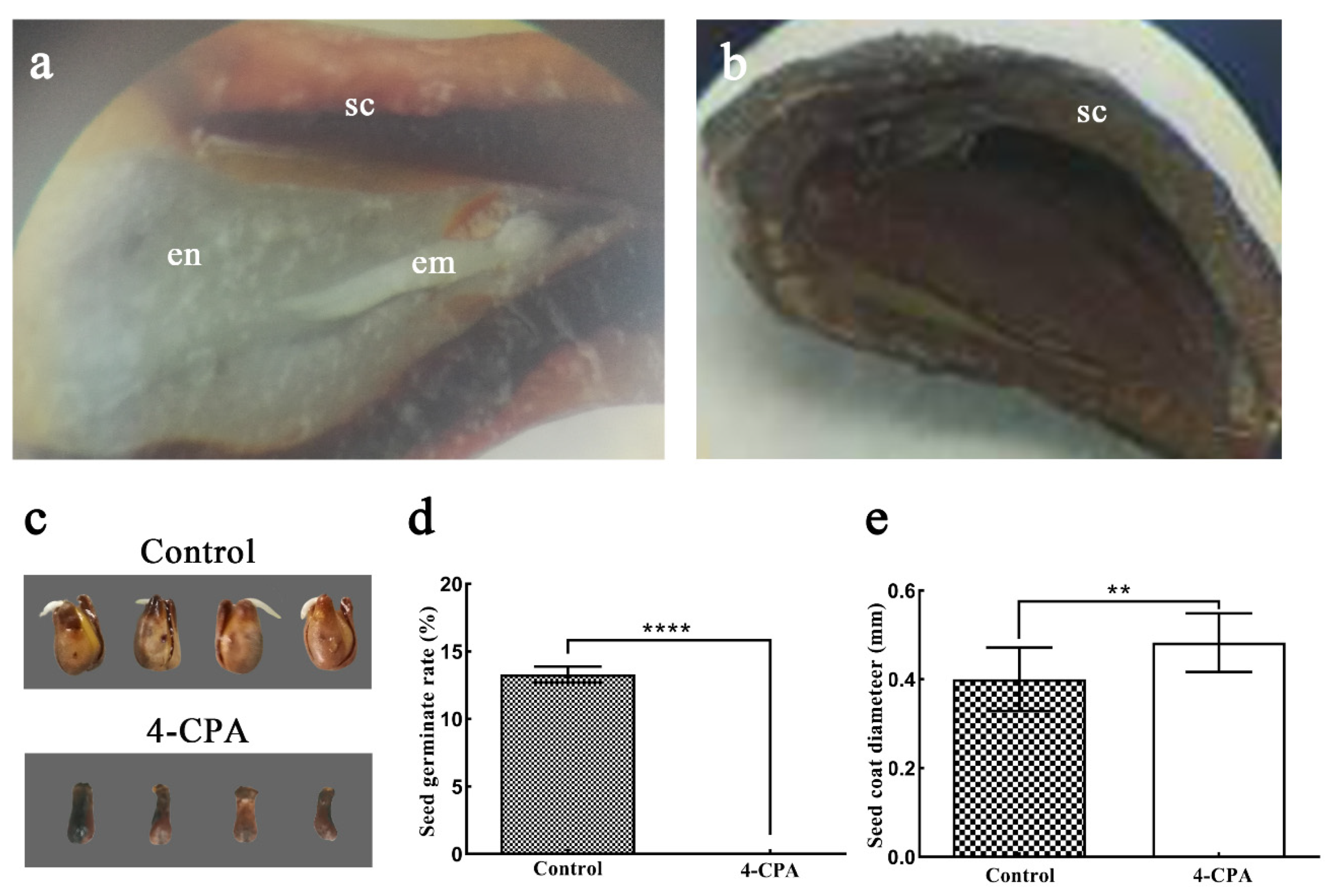
3.2. The Effect of 4-CPA Treatment on Seed Growth and Development
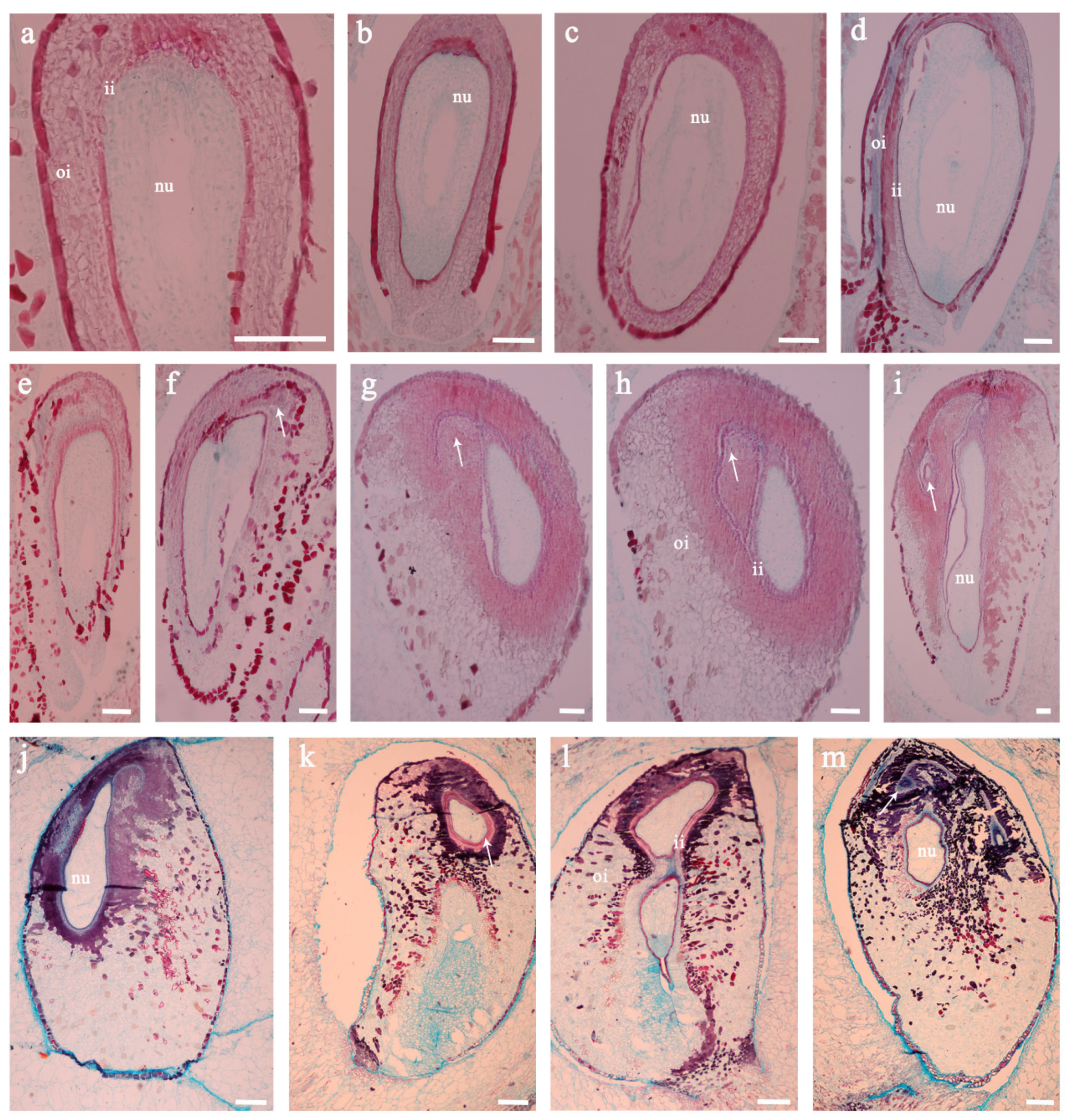
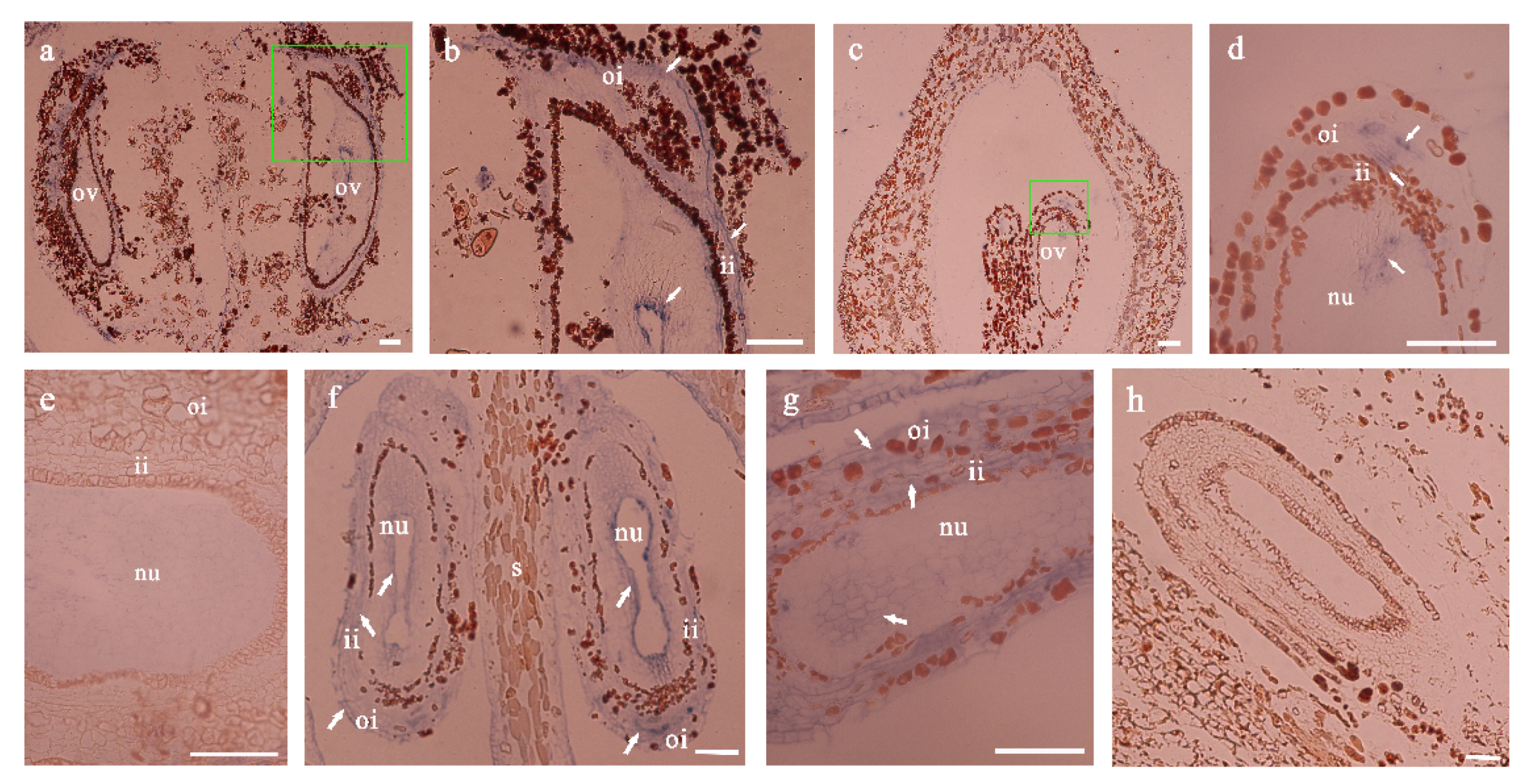
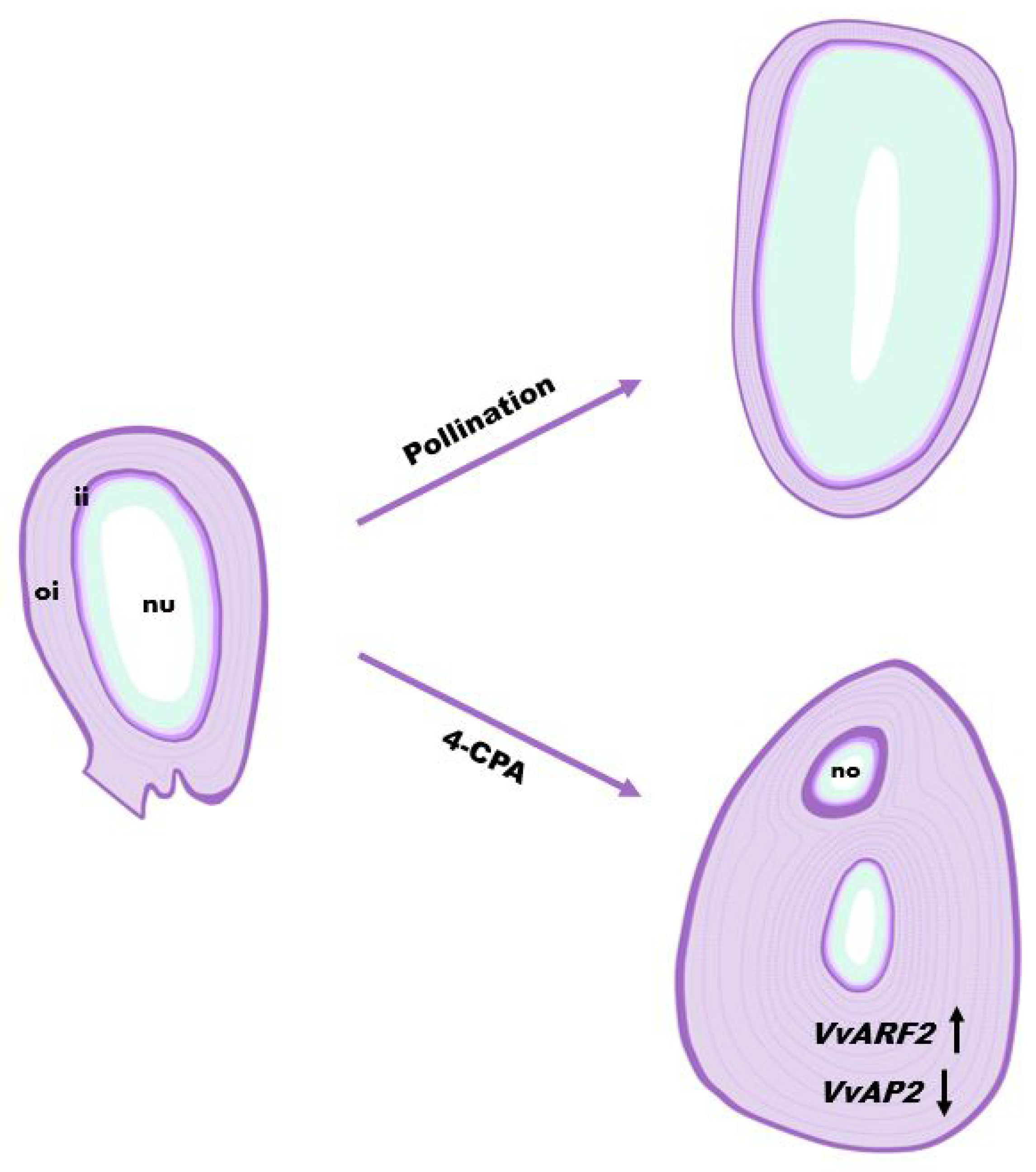
3.3. Observation of Histological Structure of Defective Seed
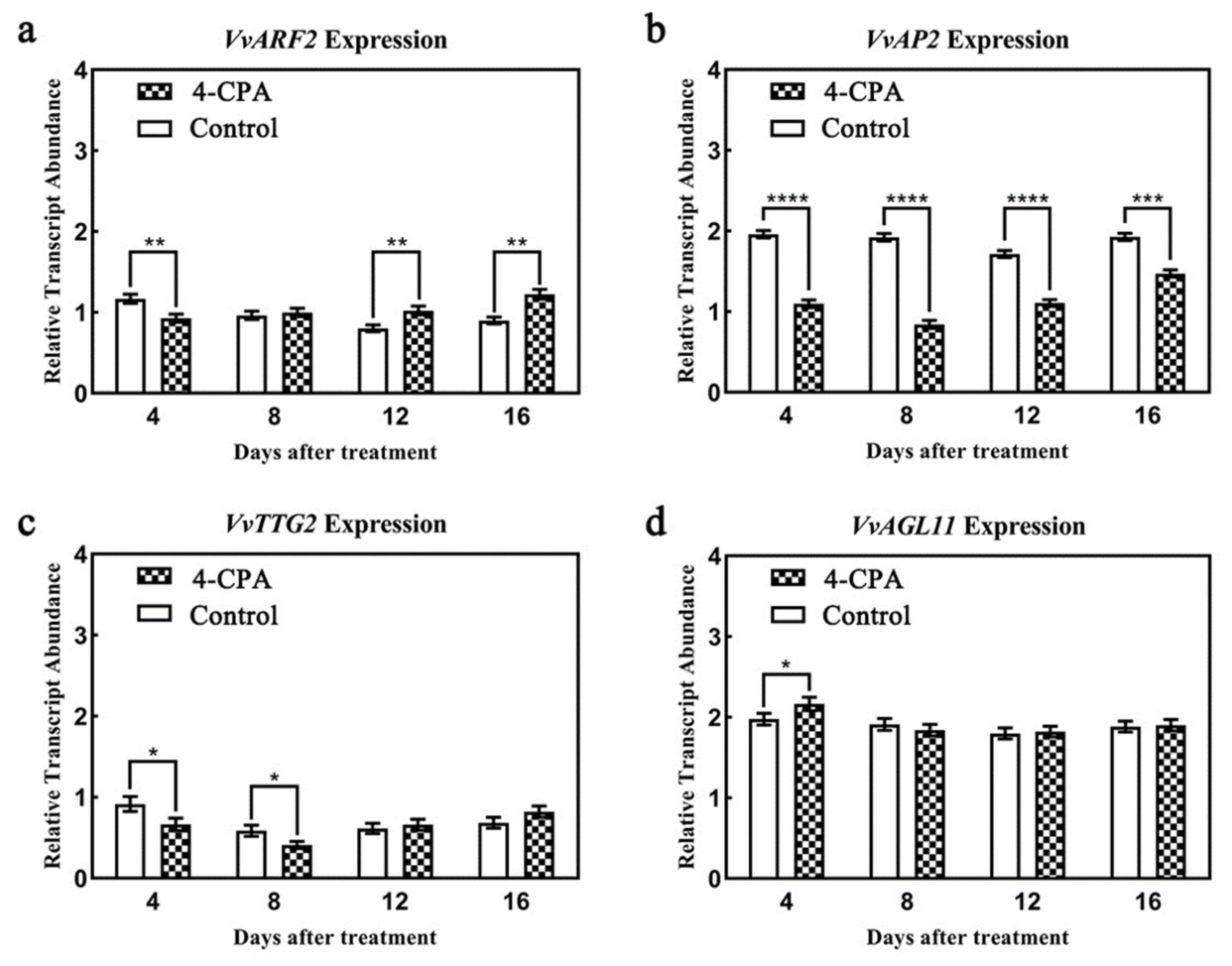
3.4. Auxin Regulates the Expression and Location of Grape-Ovule-Related Genes
4. Discussion
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Acknowledgments
Conflicts of Interest
References
- Qian, C.L.; Ren, N.N.; Wang, J.Y.; Xu, Q.; Chen, X.H.; Qi, X.H. Effects of exogenous application of CPPU, NAA and GA(4+7) on parthenocarpy and fruit quality in cucumber (Cucumis sativus L.). Food Chem. 2018, 243, 410–413. [Google Scholar] [CrossRef]
- Cong, L.; Yue, R.R.; Wang, H.B.; Liu, J.L.; Zhai, R.; Yang, J.; Wu, M.; Si, M.; Zhang, H.Q.; Yang, C.Q.; et al. 2,4-D-induced parthenocarpy in pear is mediated by enhancement of GA(4) biosynthesis. Physiol. Plant 2019, 166, 812–820. [Google Scholar] [CrossRef]
- Beziat, C.; Kleine-Vehn, J. The Road to Auxin-Dependent Growth Repression and Promotion in Apical Hooks. Curr. Biol. 2018, 28, R519–R525. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Radoeva, T.; Lokerse, A.S.; Llavata-Peris, C.I.; Wendrich, J.R.; Xiang, D.Q.; Liao, C.Y.; Vlaar, L.; Boekschoten, M.; Hooiveld, G.; Datla, R.; et al. A Robust Auxin Response Network Controls Embryo and Suspensor Development through a Basic Helix Loop Helix Transcriptional Module. Plant Cell 2019, 31, 52–67. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Du, M.; Spalding, E.P.; Gray, W.M. Rapid Auxin-Mediated Cell Expansion. Annu. Rev. Plant Biol. 2020, 71, 379–402. [Google Scholar] [CrossRef] [Green Version]
- Carrera, E.; Ruiz-Rivero, O.; Peres, L.E.P.; Atares, A.; Garcia-Martinez, J.L. Characterization of the procera Tomato Mutant Shows Novel Functions of the SlDELLA Protein in the Control of Flower Morphology, Cell Division and Expansion, and the Auxin-Signaling Pathway during Fruit-Set and Development. Plant Physiol. 2012, 160, 1581–1596. [Google Scholar] [CrossRef] [Green Version]
- Shinozaki, Y.; Beauvoit, B.P.; Takahara, M.; Hao, S.; Ezura, K.; Andrieu, M.-H.; Nishida, K.; Mori, K.; Suzuki, Y.; Kuhara, S.; et al. Fruit setting rewires central metabolism via gibberellin cascades. Proc. Natl. Acad. Sci. USA 2020, 117, 23970–23981. [Google Scholar] [CrossRef]
- Serrani, J.C.; Ruiz-Rivero, O.; Fos, M.; Garcia-Martinez, J.L. Auxin-induced fruit-set in tomato is mediated in part by gibberellins. Plant J. 2008, 56, 922–934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, J.H.; Israeli, A.; Ori, N.; Sun, T.P. The Interaction between DELLA and ARF/IAA Mediates Crosstalk between Gibberellin and Auxin Signaling to Control Fruit Initiation in Tomato. Plant Cell 2018, 30, 1710–1728. [Google Scholar] [CrossRef] [Green Version]
- Pandolfini, T.; Molesini, B.; Spena, A. Molecular dissection of the role of auxin in fruit initiation. Trends Plant Sci. 2007, 12, 327–329. [Google Scholar] [CrossRef] [PubMed]
- Marti, C.; Orzaez, D.; Ellul, P.; Moreno, V.; Carbonell, J.; Granell, A. Silencing of DELLA induces facultative parthenocarpy in tomato fruits. Plant J. 2007, 52, 865–876. [Google Scholar] [CrossRef] [PubMed]
- Cheng, C.X.; Xu, X.Z.; Singer, S.D.; Li, J.; Zhang, H.J.; Gao, M.; Wang, L.; Song, J.Y.; Wang, X.P. Effect of GA(3) Treatment on Seed Development and Seed-Related Gene Expression in Grape. PLoS ONE 2013, 8, e80044. [Google Scholar]
- Mesejo, C.; Munoz-Fambuena, N.; Reig, C.; Martinez-Fuentes, A.; Agusti, M. Cell division interference in newly fertilized ovules induces stenospermocarpy in cross-pollinated citrus fruit. Plant Sci. 2014, 225, 86–94. [Google Scholar] [CrossRef]
- Lu, L.; Liang, J.J.; Zhu, X.; Xiao, K.; Li, T.Z.; Hu, J.F. Auxin- and cytokinin-induced berries set in grapevine partly rely on enhanced gibberellin biosynthesis. Tree Genet. Genomes 2016, 12, 12. [Google Scholar] [CrossRef]
- Figueiredo, D.D.; Kohler, C. Auxin: A molecular trigger of seed development. Gene Dev. 2018, 32, 479–490. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smit, M.E.; Weijers, D. The role of auxin signaling in early embryo pattern formation. Curr. Opin. Plant Biol. 2015, 28, 99–105. [Google Scholar] [CrossRef] [PubMed]
- Figueiredo, D.D.; Batista, R.A.; Roszak, P.J.; Kohler, C. Auxin production couples endosperm development to fertilization. Nat. Plants 2015, 1, 1–6. [Google Scholar] [CrossRef]
- Matilla, A.J. Auxin: Hormonal Signal Required for Seed Development and Dormancy. Plants 2020, 9, 17. [Google Scholar] [CrossRef]
- Figueiredo, D.D.; Batista, R.A.; Roszakt, P.J.; Hennig, L.; Kohler, C. Auxin production in the endosperm drives seed coat development in Arabidopsis. Elife 2016, 5, e20542. [Google Scholar] [CrossRef]
- Roszak, P.; Kohler, C. Polycomb group proteins are required to couple seed coat initiation to fertilization. Proc. Natl. Acad. Sci. USA 2011, 108, 20826–20831. [Google Scholar] [CrossRef] [Green Version]
- Serrani, J.C.; Fos, M.; Atares, A.; Garcia-Martinez, J.L. Effect of gibberellin and auxin on parthenocarpic fruit growth induction in the cv micro-tom of tomato. J. Plant Growth Regul. 2007, 26, 211–221. [Google Scholar] [CrossRef]
- Krizek, B.A.; Fletcher, J.C. Molecular mechanisms of flower development: An armchair guide. Nat. Rev. Genet. 2005, 6, 688–698. [Google Scholar] [CrossRef]
- Sharma, P.; Kumar, V.; Singh, S.K.; Thakur, S.; Siwach, P.; Sreenivasulu, Y.; Srinivasan, R.; Bhat, S.R. Promoter Trapping and Deletion Analysis Show Arabidopsis thaliana APETALA2 Gene Promoter Is Bidirectional and Functions as a Pollen- and Ovule-Specific Promoter in the Reverse Orientation. Appl. Biochem. Biotechnol. 2017, 182, 1591–1604. [Google Scholar] [CrossRef]
- Feng, K.; Hou, X.L.; Xing, G.M.; Liu, J.X.; Duan, A.Q.; Xu, Z.S.; Li, M.Y.; Zhuang, J.; Xiong, A.S. Advances in AP2/ERF super-family transcription factors in plant. Crit. Rev. Biotechnol. 2020, 40, 750–776. [Google Scholar] [CrossRef]
- Jofuku, K.D.; Omidyar, P.K.; Gee, Z.; Okamuro, J.K. Control of seed mass and seed yield by the floral homeotic gene APETALA2. Proc. Natl. Acad. Sci. USA 2005, 102, 3117–3122. [Google Scholar] [CrossRef] [Green Version]
- Ohto, M.; Fischer, R.L.; Goldberg, R.B.; Nakamura, K.; Harada, J.J. Control of seed mass by APETALA2. Proc. Natl. Acad. Sci. USA 2005, 102, 3123–3128. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, H.; Johnson, P.; Stepanova, A.; Alonso, J.M.; Ecker, J.R. Convergence of signaling of differential cell growth pathways in the control in Arabidopsis. Dev. Cell 2004, 7, 193–204. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schruff, M.C.; Spielman, M.; Tiwari, S.; Adams, S.; Fenby, N.; Scott, R.J. The AUXIN RESPONSE FACTOR 2 gene of Arabidopsis links auxin signalling, cell division, and the size of seeds and other organs. Development 2006, 133, 251–261. [Google Scholar] [CrossRef] [Green Version]
- Malabarba, J.; Buffon, V.; Mariath, J.E.A.; Gaeta, M.L.; Dornelas, M.C.; Margis-Pinheiro, M.; Pasquali, G.; Revers, L.F. The MADS-box gene Agamous-like 11 is essential for seed morphogenesis in grapevine. J. Exp. Bot. 2017, 68, 1493–1506. [Google Scholar] [CrossRef]
- Malabarba, J.; Buffon, V.; Mariath, J.E.A.; Maraschin, F.S.; Margis-Pinheiro, M.; Pasquali, G.; Revers, L.F. Manipulation of VviAGL11 expression changes the seed content in grapevine (Vitis vinifera L.). Plant Sci. 2018, 269, 126–135. [Google Scholar] [CrossRef] [Green Version]
- Royo, C.; Torres-Perez, R.; Mauri, N.; Diestro, N.; Antonio Cabezas, J.; Marchal, C.; Lacombe, T.; Ibanez, J.; Tornel, M.; Carreno, J.; et al. The Major Origin of Seedless Grapes Is Associated with a Missense Mutation in the MADS-Box Gene VviAGL11. Plant Physiol. 2018, 177, 1234–1253. [Google Scholar] [CrossRef] [Green Version]
- Amato, A.; Cavallini, E.; Zenoni, S.; Finezzo, L.; Begheldo, M.; Ruperti, B.; Tornielli, G.B. A Grapevine TTG2-Like WRKY Transcription Factor Is Involved in Regulating Vacuolar Transport and Flavonoid Biosynthesis. Front. Plant Sci. 2017, 7, 20. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(T)(-Delta Delta C) method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef] [PubMed]
- Drews, G.N.; Bowman, J.L.; Meyerowitz, E.M. Negative Regulation of the Arabidopsis Homeotic Gene Agamous by the Apetala2 Product. Cell 1991, 65, 991–1002. [Google Scholar] [CrossRef]
- Kelley, D.R.; Gasser, C.S. Ovule development: Genetic trends and evolutionary considerations. Sex Plant Reprod. 2009, 22, 229–234. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ingram, G.C. Family life at close quarters: Communication and constraint in angiosperm seed development. Protoplasma 2010, 247, 195–214. [Google Scholar] [CrossRef] [PubMed]
- Ingram, G.C. Dying to live: Cell elimination as a developmental strategy in angiosperm seeds. J. Exp. Bot. 2017, 68, 785–796. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gardarin, A.; Durr, C.; Mannino, M.R.; Busset, H.; Colbach, N. Seed mortality in the soil is related to seed coat thickness. Seed Sci. Res. 2010, 20, 243–256. [Google Scholar] [CrossRef]
- Kelly, K.M.; Staden, J.V.; Bell, W.E. Seed coat structure and dormancy. Plant Growth Regul. 1992, 11, 201–209. [Google Scholar] [CrossRef]
- Cao, J.S.; Li, G.J.; Qu, D.J.; Li, X.; Wang, Y.N. Into the Seed: Auxin Controls Seed Development and Grain Yield. Int. J. Mol. Sci. 2020, 21, 17. [Google Scholar] [CrossRef] [Green Version]
- Larsson, E.; Vivian-Smith, A.; Offringa, R.; Sundberg, E. Auxin Homeostasis in Arabidopsis Ovules Is Anther-Dependent at Maturation and Changes Dynamically upon Fertilization. Front. Plant Sci. 2017, 8, 14. [Google Scholar] [CrossRef] [PubMed]
- Wan, S.B.; Li, W.L.; Zhu, Y.Y.; Liu, Z.M.; Huang, W.D.; Zhan, J.C. Genome-wide identification, characterization and expression analysis of the auxin response factor gene family in Vitis vinifera. Plant Cell Rep. 2014, 33, 1365–1375. [Google Scholar] [CrossRef] [PubMed]
- Shangguan, L.F.; Wang, C.; Kayesh, E.; Zhang, Y.P.; Korir, N.K.; Han, J.; Fang, J.G. Review and structural analysis of the evolution of grapevine (Vitis vinifera L.) genes involved in flower and fruit development. J. Horticult. Sci. Biotechnol. 2012, 87, 243–249. [Google Scholar] [CrossRef]
- Ohto, M.A.; Floyd, S.K.; Fischer, R.L.; Goldberg, R.B.; Harada, J.J. Effects of APETALA2 on embryo, endosperm, and seed coat development determine seed size in Arabidopsis. Sex Plant. Reprod. 2009, 22, 277–289. [Google Scholar] [CrossRef] [PubMed] [Green Version]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Inner Integument | Outer Integument | ||||||
|---|---|---|---|---|---|---|---|
| Cell Layers | Cell Diameter (μm) | Cell Area (μm2) | Cell Layers | Cell Diameter (μm) | Cell Area (μm2) | ||
| 4DAT | Control | 2.67 ± 0.47 | 4.42 ± 0.45 | 61.93 ± 13.11 | 5.44 ± 0.50 | 7.18 ± 0.29 | 162.29 ± 13.06 |
| 4-CPA | 2.78 ± 0.42 | 4.75 ± 0.40 | 71.44 ± 12.03 | 7.44 ± 0.50 | 11.74 ± 1.06 | 436.26 ± 80.40 | |
| 8DAT | Control | 2.67 ± 0.47 | 4.51 ± 0.36 | 64.30 ± 10.01 | 5.78 ± 0.63 | 7.44 ± 0.52 | 174.55 ± 24.53 |
| 4-CPA | 3.00 ± 0.00 | 5.44 ± 0.41 | 93.56 ± 13.12 | 8.11 ± 0.31 | 12.29 ± 1.02 | 477.76 ± 75.40 | |
| 12DAT | Control | 2.89 ± 0.31 | 3.39 ± 0.30 | 36.32 ± 6.60 | 6.33 ± 0.47 | 9.32 ± 0.57 | 273.83 ± 32.73 |
| 4-CPA | 3.00 ± 0.00 | 5.10 ± 0.19 | 81.89 ± 6.19 | 12.22 ± 0.92 | 15.04 ± 0.49 | 711.56 ± 45.95 | |
| 16DAT | Control | 3.00 ± 0.00 | 3.44 ± 0.25 | 37.27 ± 5.27 | 6.44 ± 0.68 | 11.16 ± 0.42 | 391.87 ± 29.35 |
| 4-CPA | 3.00 ± 0.00 | 4.88 ± 0.35 | 75.32 ± 10.81 | 13.00 ± 0.94 | 15.07 ± 1.28 | 718.75 ± 119.22 | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Liu, Z.; Wang, Y.; Pu, W.; Zhu, H.; Liang, J.; Wu, J.; Hong, L.; Guan, P.; Hu, J. 4-CPA (4-Chlorophenoxyacetic Acid) Induces the Formation and Development of Defective “Fenghou” (Vitis vinifera × V. labrusca) Grape Seeds. Biomolecules 2021, 11, 515. https://doi.org/10.3390/biom11040515
Liu Z, Wang Y, Pu W, Zhu H, Liang J, Wu J, Hong L, Guan P, Hu J. 4-CPA (4-Chlorophenoxyacetic Acid) Induces the Formation and Development of Defective “Fenghou” (Vitis vinifera × V. labrusca) Grape Seeds. Biomolecules. 2021; 11(4):515. https://doi.org/10.3390/biom11040515
Chicago/Turabian StyleLiu, Zhenhua, Yan Wang, Wenjiang Pu, Haifeng Zhu, Jinjun Liang, Jiang Wu, Liang Hong, Pingyin Guan, and Jianfang Hu. 2021. "4-CPA (4-Chlorophenoxyacetic Acid) Induces the Formation and Development of Defective “Fenghou” (Vitis vinifera × V. labrusca) Grape Seeds" Biomolecules 11, no. 4: 515. https://doi.org/10.3390/biom11040515