Cerrena unicolor Laccases, Genes Expression and Regulation of Activity

 , , ,
, , ,  , , , and
, , , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Growth Conditions
2.2. Laccase Activity and Protein Assay
2.3. Separation of Protein Samples by Native PAGE and SDS-PAGE
2.4. Peptide Sequencing by LC-MS/MS
2.5. RNA Isolation and RT-PCR
2.6. 2-D gel Electrophoresis
2.7. Purification of Endogenous Aspartic-Like Protease from the C. unicolor Culture Fluid
2.8. Measurements of Protease Activity
2.9. Laccase-Protease Experiments
3. Results
3.1. LC-MS/MS Analyses
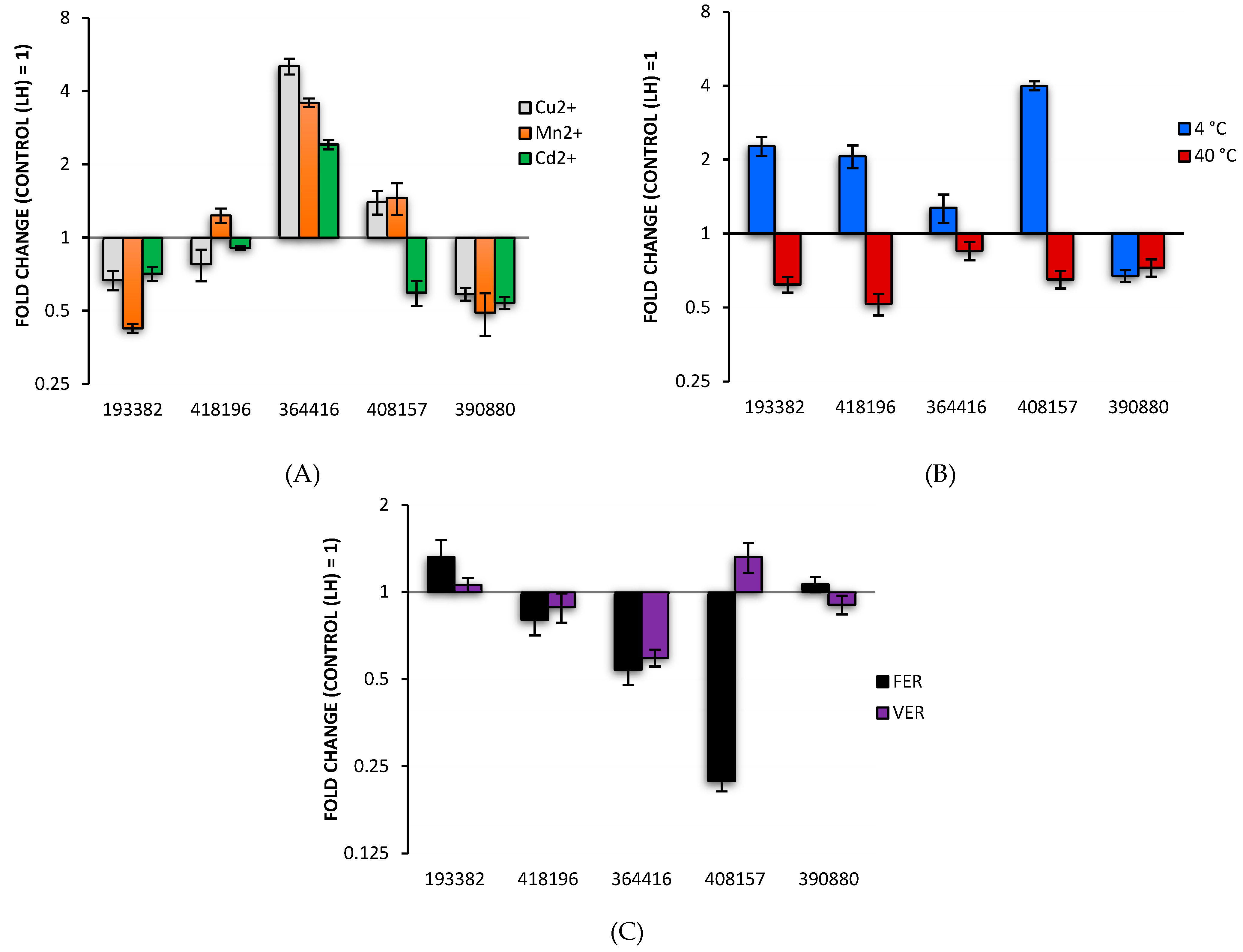
3.2. Analysis of Differentially Expressed Laccase Genes
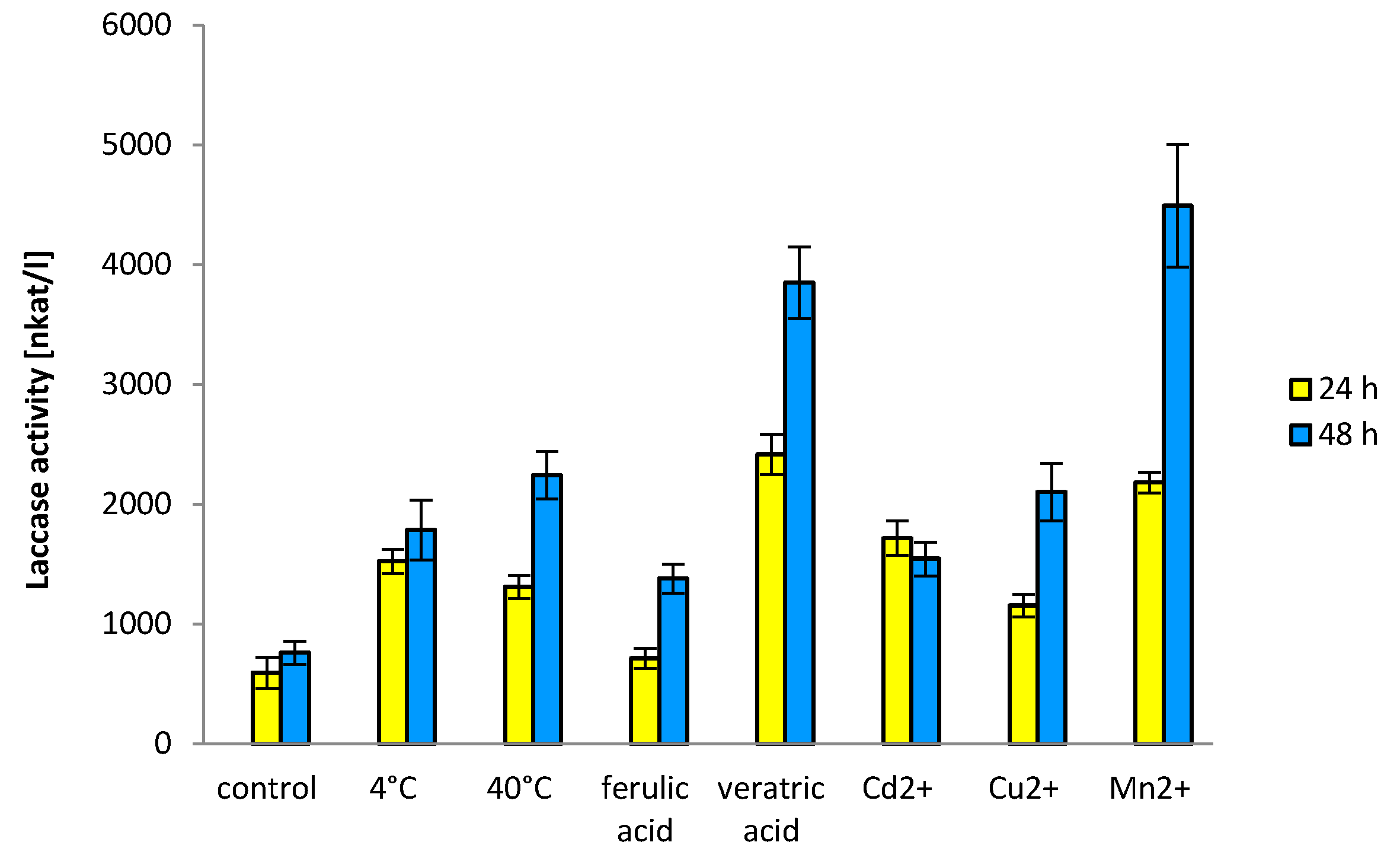
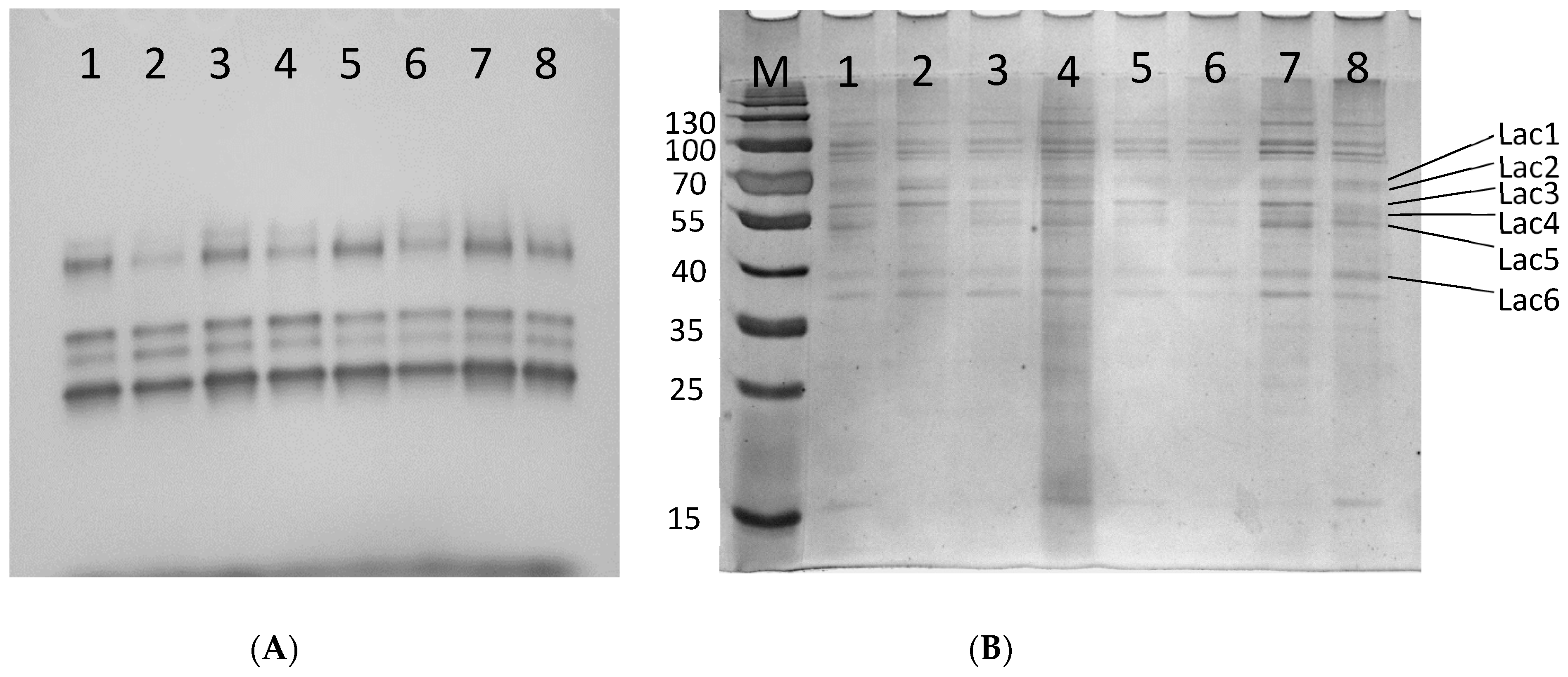
3.3. Laccase Enzymatic Activity and Electrophoretic Profiles
3.4. 2-D Electrophoresis and LC-MS/MS
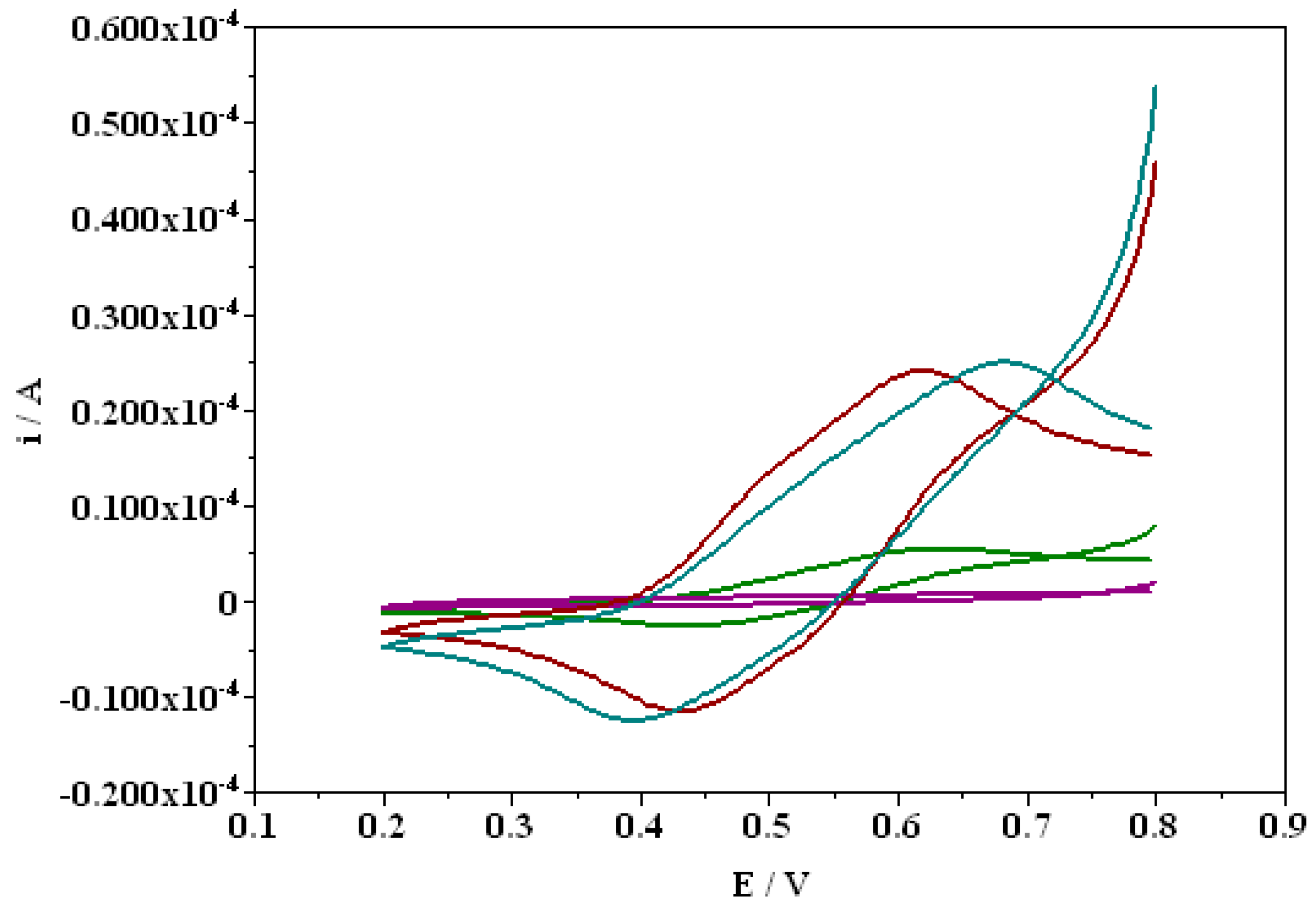
3.5. Proteolytic Modifications of Laccase by C. unicolor Endogenous Protease
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Belova, O.V.; Lisov, A.V.; Vinokurova, N.G.; Kostenevich, A.A.; Sapunova, L.I.; Lobanok, A.G.; Leont’evskii, A.A. Xylanase and cellulase of fungus Cerrena unicolor VKM F-3196: Production, properties, and applications for the saccharification of plant material. Prikl. Biokhim. Mikrobiol. 2014, 50, 171–176. [Google Scholar] [CrossRef] [PubMed]
- Rogalski, J.; Dawidowicz, A.; Jozwik, E.; Leonowicz, A. Immobilization of laccase from Cerrena unicolor on controlled porosity glass. J. Mol. Catal. B Enzym. 1999, 6, 29–39. [Google Scholar] [CrossRef]
- Elisashvili, V.I.; Kvesitadze, G.I. Laccase and Ligninase Activities of Cerrena-Unicolor. Bioconversion Plant Raw Mater. Biotechnol. Adv. 1991, 122, 19–29. [Google Scholar]
- Janusz, G.; Mazur, A.; Wielbo, J.; Koper, P.; Zebracki, K.; Pawlik, A.; Ciolek, B.; Paszczynski, A.; Kubik-Komar, A. Comparative transcriptomic analysis of Cerrena unicolor revealed differential expression of genes engaged in degradation of various kinds of wood. Microbiol. Res. 2018, 207, 256–268. [Google Scholar] [CrossRef] [PubMed]
- Fink-Boots, M.; Wilkolazka, A.; Malarczyk, E.; Leonowicz, A. Changes in sets of laccase isoforms under stress conditions in Cerrena unicolor. In Proceedings of the 7th International Conference on Biotechnology in the Pulp and Paper Industry, Vancouver, BC, Canada, 16–19 June 1998; Volume B, pp. B179–B181. [Google Scholar]
- Yang, J.; Wang, G.; Ng, T.B.; Lin, J.; Ye, X. Laccase Production and Differential Transcription of Laccase Genes in Cerrena sp. in Response to Metal Ions, Aromatic Compounds, and Nutrients. Front. Microbiol. 2015, 6, 1558. [Google Scholar] [CrossRef]
- Janusz, G.; Rogalski, J.; Szczodrak, J. Increased production of laccase by Cerrena unicolor in submerged liquid cultures. World J. Microbiol. Biotechnol. 2007, 23, 1459–1464. [Google Scholar] [CrossRef]
- Jarosz-Wilkolazka, A.; Graz, M.; Braha, B.; Menge, S.; Schlosser, D.; Krauss, G.J. Species-specific Cd-stress response in the white rot basidiomycetes Abortiporus biennis and Cerrena unicolor. Biometals 2006, 19, 39–49. [Google Scholar] [CrossRef] [PubMed]
- Fink-Boots, M.D.; Jaszek, M.B.; Leonowicz, A. Heat shock stimulation of laccase in Abortiporus biennis and Cerrena unicolor. Biol. Sci. Symp. 1997, 395–396. [Google Scholar]
- Sojka-Ledakowicz, J.; Lichawska-Olczyk, J.; Ledakowicz, S.; Michniewicz, A. Bio-scouring of linen fabrics with laccase complex from Cerrena unicolor. Fibres Text. East. Eur. 2007, 15, 86–89. [Google Scholar]
- Zhao, M.; Zhang, B.B.; Lu, L.; Zhao, L.Y.; Liang, S.C. Purification and characterization of laccase from the white rot fungus Cerrena unicolor and its use in dye decolorization. J. Biotechnol. 2008, 136, S327. [Google Scholar] [CrossRef]
- Mann, J.; Markham, J.L.; Peiris, P.; Spooner-Hart, R.N.; Holford, P.; Nair, N.G. Use of olive mill wastewater as a suitable substrate for the production of laccase by Cerrena consors. Int. Biodeterior. Biodegrad. 2015, 99, 138–145. [Google Scholar] [CrossRef]
- Matuszewska, A.; Karp, M.; Jaszek, M.; Janusz, G.; Osinska-Jaroszuk, M.; Sulej, J.; Stefaniuk, D.; Tomczak, W.; Giannopoulos, K. Laccase purified from Cerrena unicolor exerts antitumor activity against leukemic cells. Oncol. Lett. 2016, 11, 2009–2018. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sharma, K.K.; Kuhad, R.C. Laccase: Enzyme revisited and function redefined. Indian J. Microbiol. 2008, 48, 309–316. [Google Scholar] [CrossRef] [Green Version]
- Leatham, G.F.; Stahmann, M.A. Studies on the Laccase of Lentinus-Edodes—Specificity, Localization and Association with the Development of Fruiting Bodies. J. Gen. Microbiol. 1981, 125, 147–157. [Google Scholar] [CrossRef] [Green Version]
- Missall, T.A.; Moran, J.M.; Corbett, J.A.; Lodge, J.K. Distinct stress responses of two functional laccases in Cryptococcus neoformans are revealed in the absence of the thiol-specific antioxidant Tsa1. Eukaryot. Cell 2005, 4, 202–208. [Google Scholar] [CrossRef] [Green Version]
- Janusz, G.; Pawlik, A.; Swiderska-Burek, U.; Polak, J.; Sulej, J.; Jarosz-Wilkolazka, A.; Paszczynski, A. Laccase Properties, Physiological Functions, and Evolution. Int. J. Mol. Sci. 2020, 21, 966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, Y.; Wei, F.; Zhuo, R.; Fan, F.; Liu, H.; Zhang, C.; Ma, L.; Jiang, M.; Zhang, X. Enhancing the laccase production and laccase gene expression in the white-rot fungus Trametes velutina 5930 with great potential for biotechnological applications by different metal ions and aromatic compounds. PLoS ONE 2013, 8, e79307. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fernandez-Fueyo, E.; Castanera, R.; Ruiz-Duenas, F.J.; Lopez-Lucendo, M.F.; Ramirez, L.; Pisabarro, A.G.; Martinez, A.T. Ligninolytic peroxidase gene expression by Pleurotus ostreatus: Differential regulation in lignocellulose medium and effect of temperature and pH. Fungal Genet. Biol. 2014, 72, 150–161. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Soden, D.M.; Dobson, A.D.W. Differential regulation of laccase gene expression in Pleurotus sajor-caju. Microbiology 2001, 147, 1755–1763. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Swatek, A.; Staszczak, M. Effect of Ferulic Acid, a Phenolic Inducer of Fungal Laccase, on 26S Proteasome Activities In Vitro. Int. J. Mol. Sci. 2020, 21, 2463. [Google Scholar] [CrossRef] [Green Version]
- Staszczak, M.; Jarosz-Wilkolazka, A. Inhibition of the proteasome strongly affects cadmium stimulated laccase activity in Trametes versicolor. Biochimie 2005, 87, 755–762. [Google Scholar] [CrossRef] [PubMed]
- Janusz, G.; Jaszek, M.; Matuszewska, A.; Draczkowski, P.; Osinska-Jaroszuk, M. Proteolytic modifications of laccase from Cerrena unicolor. J. Mol. Catal. B Enzym. 2015, 122, 330–338. [Google Scholar] [CrossRef]
- Janusz, G.; Sulej, J.; Jaszek, M.; Osinska-Jaroszuk, M. Effect of different wavelengths of light on laccase, cellobiose dehydrogenase, and proteases produced by Cerrena unicolor, Pycnoporus sanguineus and Phlebia lindtneri. Acta Biochim. Pol. 2016, 63, 223–228. [Google Scholar] [CrossRef] [PubMed]
- Lindeberg, G.; Holm, G. Occurrence of tyrosinase and laccase in fruit bodies and mycelia of some Hymenomycetes. Physiol. Plant. 1952, 5, 100–114. [Google Scholar] [CrossRef]
- Jarosz-Wilkolazka, A.; Fink-Boots, M.; Malarczyk, E.; Leonowicz, A. Formaldehyde as a proof and response to various kind of stress in some Basidiomycetes. Acta Biol. Hung. 1998, 49, 393–403. [Google Scholar]
- Leonowicz, A.; Grzywnowicz, K. Quantitative estimation of laccase forms in some white-rot fungi using syringaldazine as a substrate. Enzym. Microb. Technol. 1981, 3, 55–58. [Google Scholar] [CrossRef]
- Bradford, M.M. A rapid and sensitive method for the quantitation of microgram quantities of protein utilizing the principle of protein-dye binding. Anal. Biochem. 1976, 72, 248–254. [Google Scholar] [CrossRef]
- Laemmli, U.K. Cleavage of structural proteins during the assembly of the head of bacteriophage T4. Nature 1970, 227, 680–685. [Google Scholar] [CrossRef] [PubMed]
- Walker, J.M. The Protein Protocols Handbook, 3rd ed.; Humana Press: New York, NY, USA, 2009; 1991p. [Google Scholar]
- Pfaffl, M.W. A new mathematical model for relative quantification in real-time RT-PCR. Nucleic Acids Res. 2001, 29, e45. [Google Scholar] [CrossRef]
- Wong, C.; Sridhara, S.; Bardwell, J.C.; Jakob, U. Heating greatly speeds Coomassie blue staining and destaining. Biotechniques 2000, 28, 426–432. [Google Scholar] [CrossRef]
- Anson, M.L. The Estimation of Pepsin, Trypsin, Papain, and Cathepsin with Hemoglobin. J. Gen. Physiol. 1938, 22, 79–89. [Google Scholar] [CrossRef]
- Heussen, C.; Dowdle, E.B. Electrophoretic analysis of plasminogen activators in polyacrylamide gels containing sodium dodecyl sulfate and copolymerized substrates. Anal. Biochem. 1980, 102, 196–202. [Google Scholar] [CrossRef]
- Songulashvili, G.; Spindler, D.; Jimenez-Tobon, G.A.; Jaspers, C.; Kerns, G.; Penninckx, M.J. Production of a high level of laccase by submerged fermentation at 120-L scale of Cerrena unicolor C-139 grown on wheat bran. Comptes Rendus Biol. 2015, 338, 121–125. [Google Scholar] [CrossRef] [PubMed]
- Filazzola, M.T.; Sannino, F.; Rao, M.A.; Gianfreda, L. Effect of various pollutants and soil-like constituents on laccase from Cerrena unicolor. J. Environ. Qual. 1999, 28, 1929–1938. [Google Scholar] [CrossRef]
- Savinova, O.S.; Moiseenko, K.V.; Vavilova, E.A.; Chulkin, A.M.; Fedorova, T.V.; Tyazhelova, T.V.; Vasina, D.V. Evolutionary Relationships Between the Laccase Genes of Polyporales: Orthology-Based Classification of Laccase lsozymes and Functional Insight From Trametes hirsuta. Front. Microbiol. 2019, 10, 152. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hua, S.; Zhang, B.; Fu, Y.; Qi, B.; Li, Y.; Tian, F. Enzymatic gene expression by Pleurotus tuoliensis (Bailinggu): Differential regulation under low temperature induction conditions. World J. Microbiol. Biotechnol. 2018, 34, 160. [Google Scholar] [CrossRef]
- Piscitelli, A.; Giardina, P.; Lettera, V.; Pezzella, C.; Sannia, G.; Faraco, V. Induction and transcriptional regulation of laccases in fungi. Curr. Genom. 2011, 12, 104–112. [Google Scholar] [CrossRef] [Green Version]
- Janusz, G.; Kucharzyk, K.H.; Pawlik, A.; Staszczak, M.; Paszczynski, A.J. Fungal laccase, manganese peroxidase and lignin peroxidase: Gene expression and regulation. Enzym. Microb. Technol. 2013, 52, 1–12. [Google Scholar] [CrossRef]
- Janusz, G.; Mazur, A.; Checinsks, A.; Malek, W.; Rogalski, J.; Ohga, S. Cloning and characterization of a laccase gene from biotechnologically important basidiomycete Cerrena unicolor. J. Fac. Agric. Kyushu Univ. 2012, 57, 41–49. [Google Scholar] [CrossRef]
- Staszczak, M.; Zdunek, E.; Leonowicz, A. Studies on the role of proteases in the white-rot fungus Trametes versicolor: Effect of PMSF and chloroquine on ligninolytic enzymes activity. J. Basic Microbiol. 2000, 40, 51–63. [Google Scholar] [CrossRef]
- Osinska-Jaroszuk, M.; Jaszek, M.; Starosielec, M.; Sulej, J.; Matuszewska, A.; Janczarek, M.; Bancerz, R.; Wydrych, J.; Wiater, A.; Jarosz-Wilkolazka, A. Bacterial exopolysaccharides as a modern biotechnological tool for modification of fungal laccase properties and metal ion binding. Bioprocess. Biosyst. Eng. 2018, 41, 973–989. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Protein ID C. unicolor 303 | Transcript ID C. unicolor FCL139 | Score | Query Coverage (%) | MW (Da) | pI |
|---|---|---|---|---|---|
| Laccase | |||||
| 193382 | XLOC_008690 | 8959 | 52 | 55139.50 | 4.64 |
| 390880 | XLOC_011744 | 6576 | 37 | 54248.29 | 5.92 |
| 418196 | XLOC_000669 | 3806 | 25 | 23445.25 | 4.44 |
| 408157 | XLOC_011551 | 720 | 9 | 55484.45 | 5.08 |
| 364416 | XLOC_011286 | 206 | 13 | 54798.69 | 5.20 |
| Aspartic protease | |||||
| 404065 | XLOC_010406 | 218 | 9.5 | 56095.00 | 4.66 |
| Time 0 min | Time 60 min | Time 120 min | |
|---|---|---|---|
| Km (mM) | 0.54 | 0.19 | 0.16 |
| Vmax (µM/min) | 64.43 | 43.46 | 37.85 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pawlik, A.; Ciołek, B.; Sulej, J.; Mazur, A.; Grela, P.; Staszczak, M.; Niścior, M.; Jaszek, M.; Matuszewska, A.; Janusz, G.; et al. Cerrena unicolor Laccases, Genes Expression and Regulation of Activity. Biomolecules 2021, 11, 468. https://doi.org/10.3390/biom11030468
Pawlik A, Ciołek B, Sulej J, Mazur A, Grela P, Staszczak M, Niścior M, Jaszek M, Matuszewska A, Janusz G, et al. Cerrena unicolor Laccases, Genes Expression and Regulation of Activity. Biomolecules. 2021; 11(3):468. https://doi.org/10.3390/biom11030468
Chicago/Turabian StylePawlik, Anna, Beata Ciołek, Justyna Sulej, Andrzej Mazur, Przemysław Grela, Magdalena Staszczak, Mateusz Niścior, Magdalena Jaszek, Anna Matuszewska, Grzegorz Janusz, and et al. 2021. "Cerrena unicolor Laccases, Genes Expression and Regulation of Activity" Biomolecules 11, no. 3: 468. https://doi.org/10.3390/biom11030468