The Receptor Tyrosine Kinase TrkA Is Increased and Targetable in HER2-Positive Breast Cancer

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Breast Tissue Samples
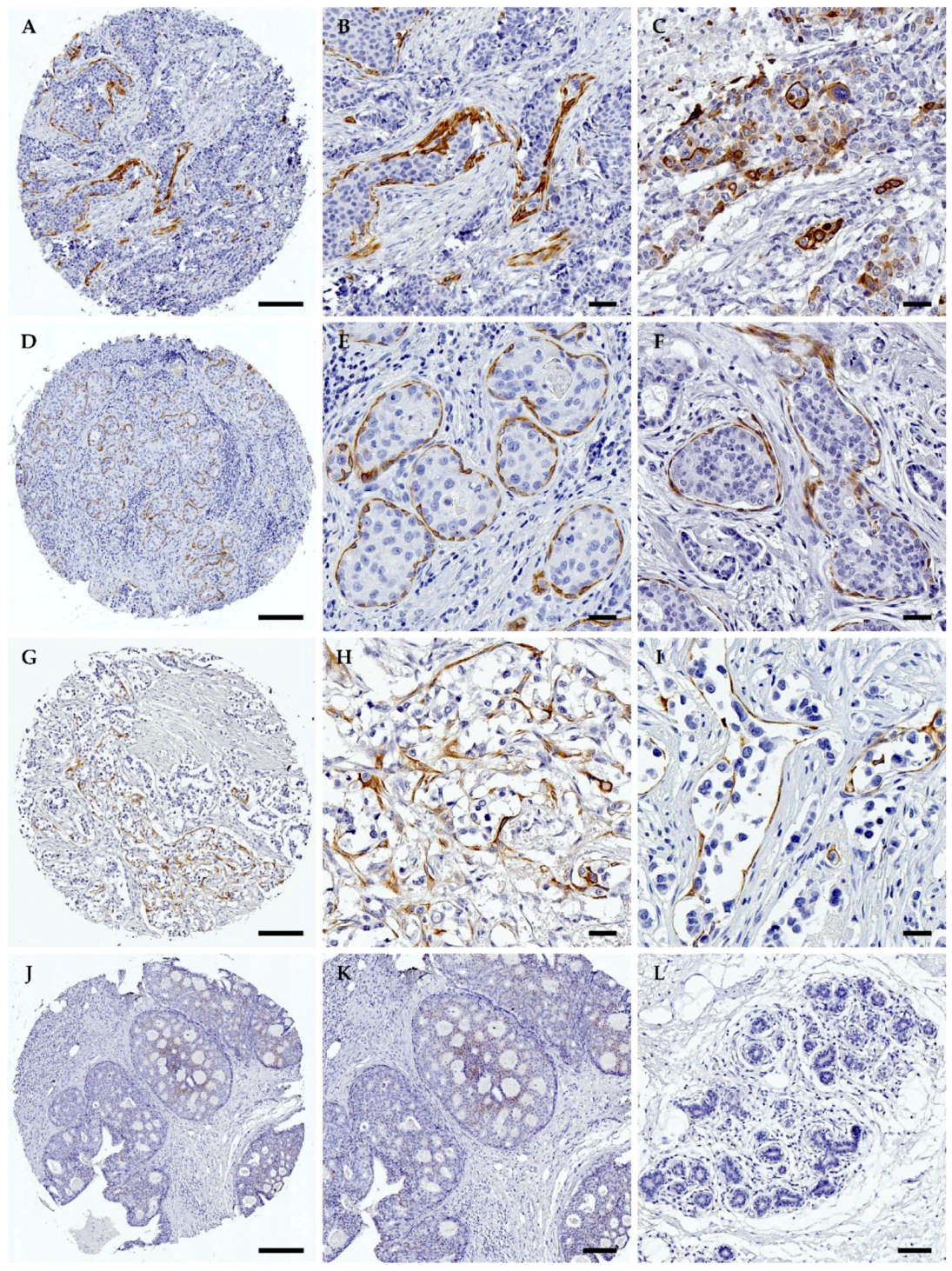
2.2. Immunohistochemistry and Quantification of TrkA Expression
2.3. Cell Culture
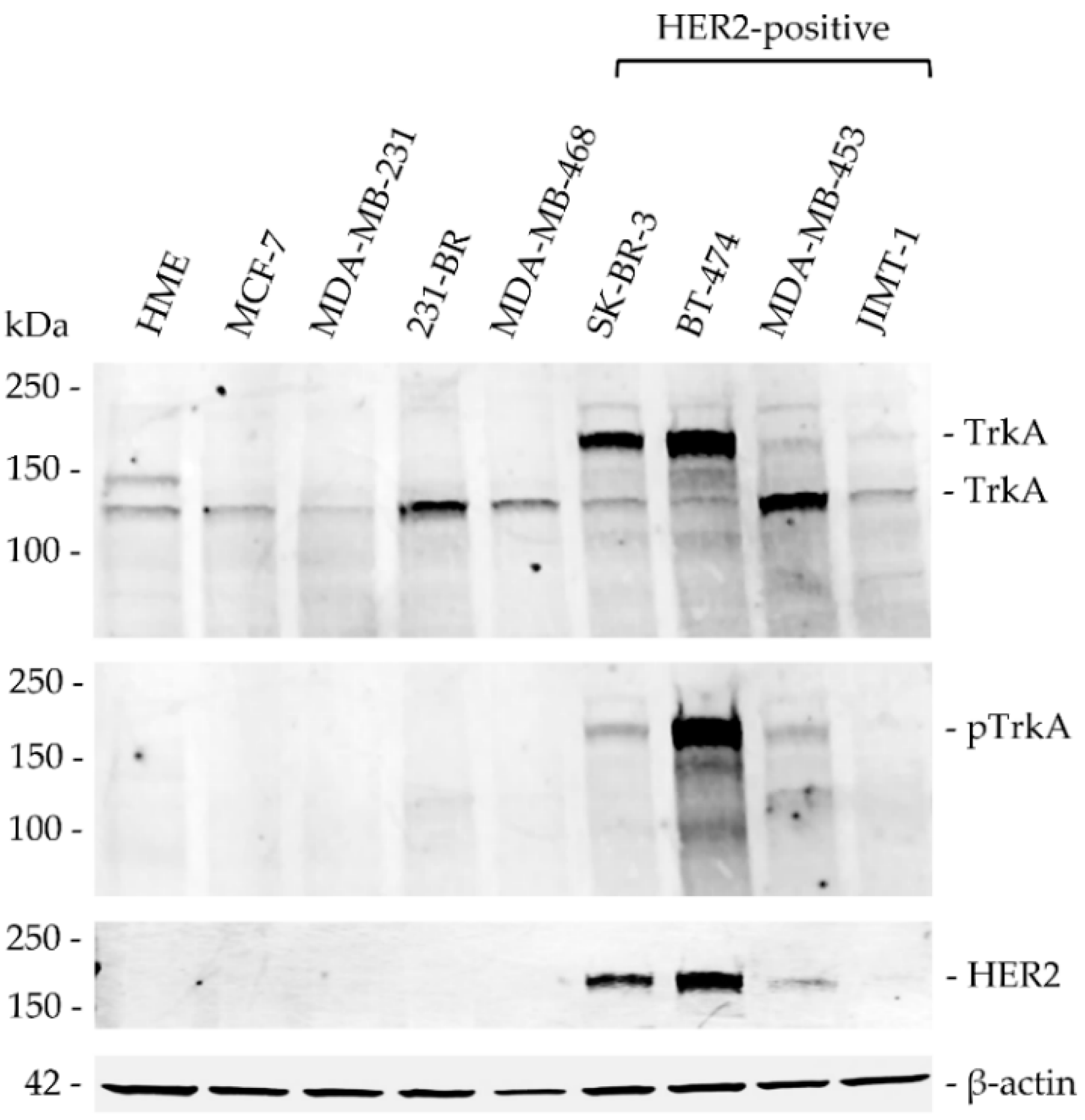
2.4. Protein Extraction and Western Blotting
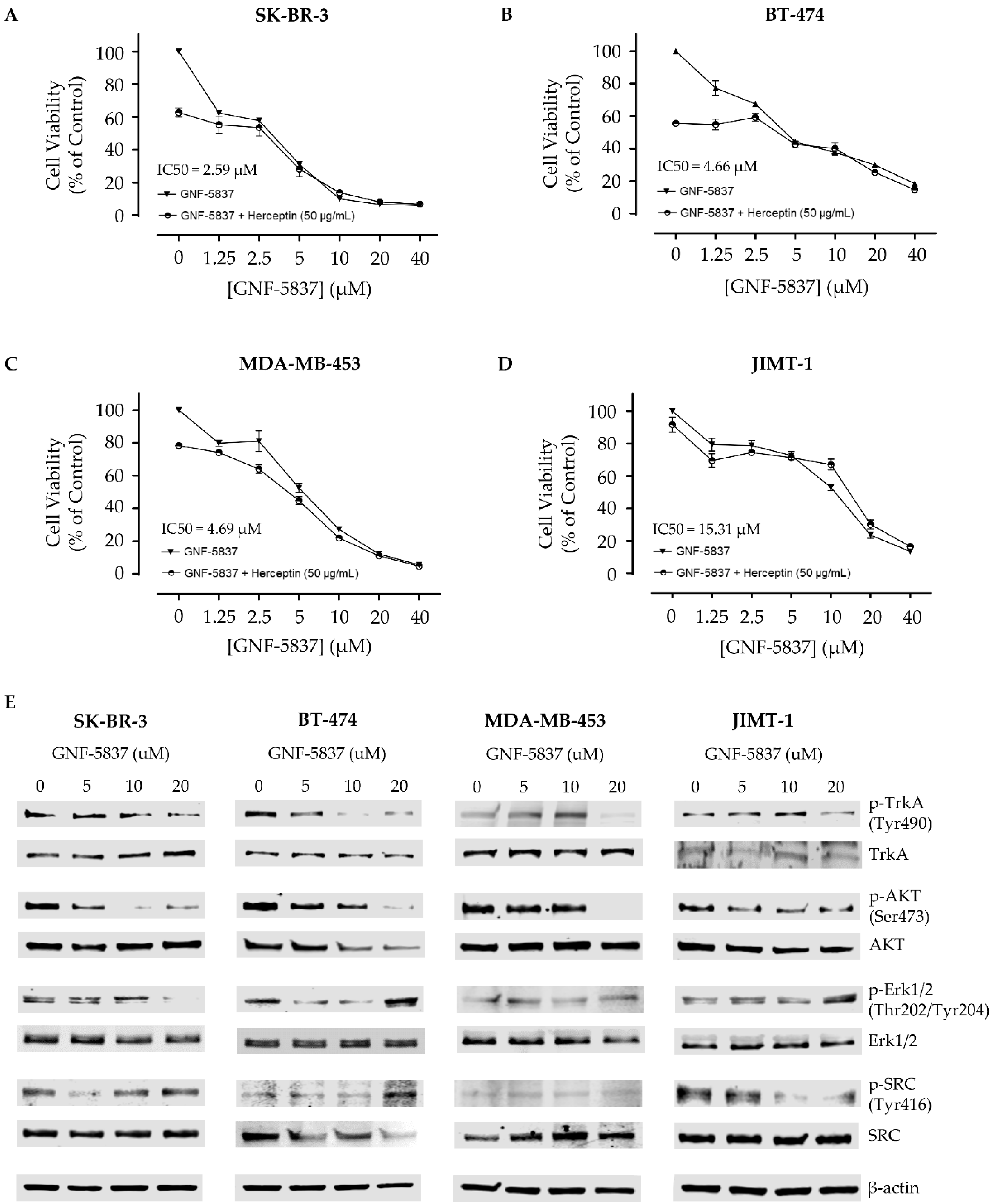
2.5. GNF-5837 Treatment and Cell Viability Assay
3. Results
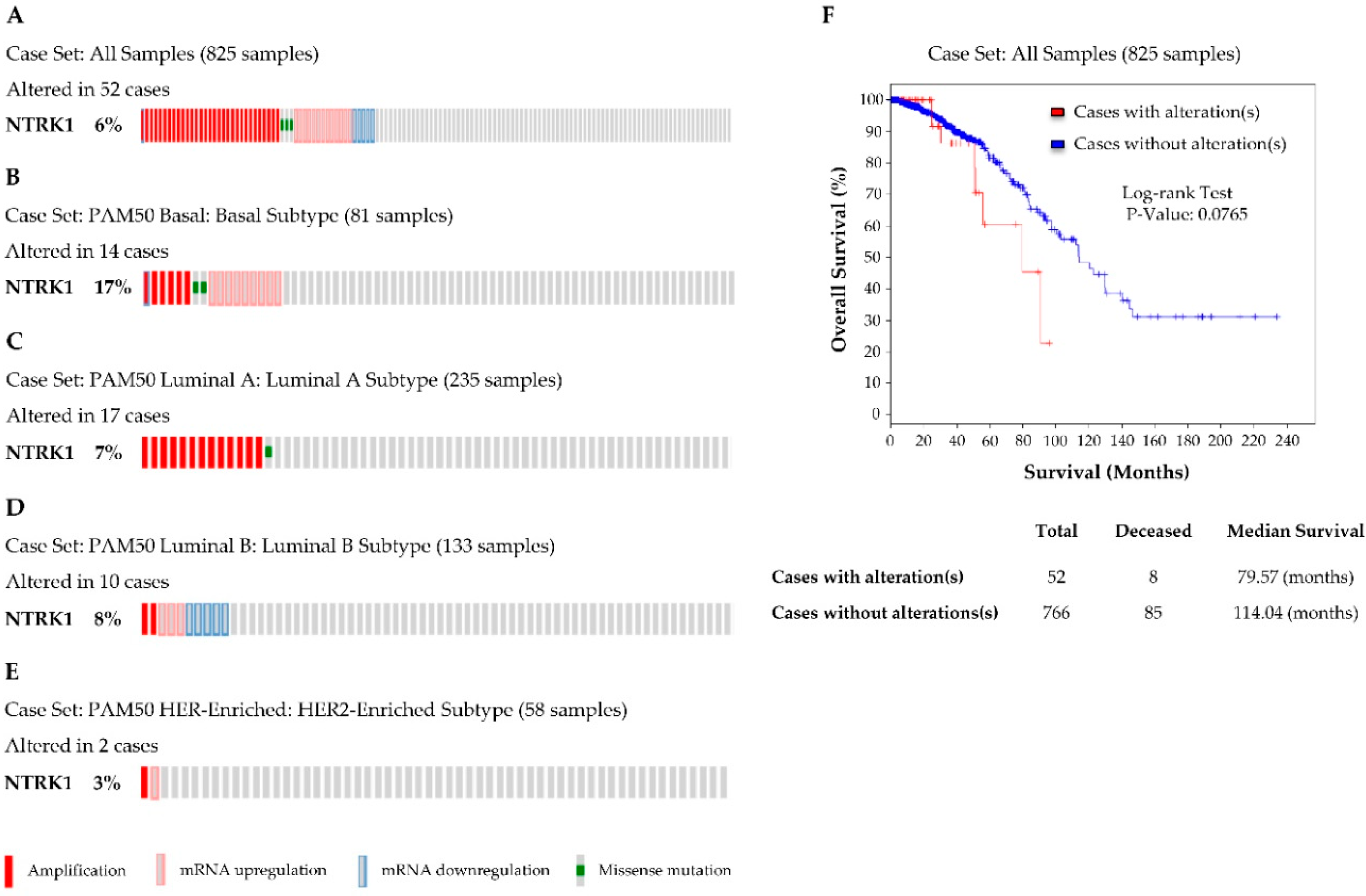
3.1. NTRK1 (TrkA) Gene Expression in Breast Cancer
3.2. TrkA Is Overexpressed in HER2-Positive Breast Tumours
3.3. TrkA Is Expressed in Breast Cancer and Overexpressed in HER2-Positive Cell Lines
3.4. Cytotoxic Effect of the Trk Inhibitor GNF-5837 in HER2-Positive Breast Cancer Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Arteaga, C.L.; Sliwkowski, M.X.; Osborne, C.K.; Perez, E.A.; Puglisi, F.; Gianni, L. Treatment of HER2-positive breast cancer: Current status and future perspectives. Nat. Rev. Clin. Oncol. 2012, 9, 16–32. [Google Scholar] [CrossRef] [PubMed]
- Wolff, A.C.; Hammond, M.E.; Hicks, D.G.; Dowsett, M.; McShane, L.M.; Allison, K.H.; Allred, D.C.; Bartlett, J.M.; Bilous, M.; Fitzgibbons, P.; et al. Recommendations for human epidermal growth factor receptor 2 testing in breast cancer: American Society of Clinical Oncology/College of American Pathologists clinical practice guideline update. J. Clin. Oncol. 2013, 31, 3997–4013. [Google Scholar] [CrossRef] [PubMed]
- Loibl, S.; Gianni, L. HER2-positive breast cancer. Lancet 2017, 389, 2415–2429. [Google Scholar] [CrossRef]
- Niikura, N.; Liu, J.; Hayashi, N.; Mittendorf, E.A.; Gong, Y.; Palla, S.L.; Tokuda, Y.; Gonzalez-Angulo, A.M.; Hortobagyi, G.N.; Ueno, N.T. Loss of human epidermal growth factor receptor 2 (HER2) expression in metastatic sites of HER2-overexpressing primary breast tumors. J. Clin. Oncol. 2012, 30, 593–599. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Niikura, N.; Tomotaki, A.; Miyata, H.; Iwamoto, T.; Kawai, M.; Anan, K.; Hayashi, N.; Aogi, K.; Ishida, T.; Masuoka, H.; et al. Changes in tumor expression of HER2 and hormone receptors status after neoadjuvant chemotherapy in 21,755 patients from the Japanese breast cancer registry. Ann. Oncol. 2016, 27, 480–487. [Google Scholar] [CrossRef]
- Ponde, N.; Brandao, M.; El-Hachem, G.; Werbrouck, E.; Piccart, M. Treatment of advanced HER2-positive breast cancer: 2018 and beyond. Cancer Treat. Rev. 2018, 67, 10–20. [Google Scholar] [CrossRef]
- Skaper, S.D. The neurotrophin family of neurotrophic factors: An overview. Methods Mol. Biol. 2012, 846, 1–12. [Google Scholar]
- Pezet, S.; McMahon, S.B. Neurotrophins: Mediators and modulators of pain. Annu. Rev. Neurosci. 2006, 29, 507–538. [Google Scholar] [CrossRef]
- Miknyoczki, S.J.; Wan, W.; Chang, H.; Dobrzanski, P.; Ruggeri, B.A.; Dionne, C.A.; Buchkovich, K. The neurotrophin-trk receptor axes are critical for the growth and progression of human prostatic carcinoma and pancreatic ductal adenocarcinoma xenografts in nude mice. Clin. Cancer Res. An. Off. J. Am. Assoc. Cancer Res. 2002, 8, 1924–1931. [Google Scholar]
- Lagadec, C.; Meignan, S.; Adriaenssens, E.; Foveau, B.; Vanhecke, E.; Romon, R.; Toillon, R.A.; Oxombre, B.; Hondermarck, H.; Le Bourhis, X. TrkA overexpression enhances growth and metastasis of breast cancer cells. Oncogene 2009, 28, 1960–1970. [Google Scholar] [CrossRef] [Green Version]
- Sasahira, T.; Ueda, N.; Yamamoto, K.; Bhawal, U.K.; Kurihara, M.; Kirita, T.; Kuniyasu, H. Trks are novel oncogenes involved in the induction of neovascularization, tumor progression, and nodal metastasis in oral squamous cell carcinoma. Clin. Exp. Metastasis 2013, 30, 165–176. [Google Scholar] [CrossRef]
- Bradshaw, R.A.; Pundavela, J.; Biarc, J.; Chalkley, R.J.; Burlingame, A.L.; Hondermarck, H. NGF and ProNGF: Regulation of neuronal and neoplastic responses through receptor signaling. Adv. Biol. Regul. 2014. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faulkner, S.; Jobling, P.; Rowe, C.W.; Rodrigues Oliveira, S.M.; Roselli, S.; Thorne, R.F.; Oldmeadow, C.; Attia, J.; Jiang, C.C.; Zhang, X.D.; et al. Neurotrophin Receptors TrkA, p75(NTR), and Sortilin Are Increased and Targetable in Thyroid Cancer. Am. J. Pathol. 2018, 188, 229–241. [Google Scholar] [CrossRef] [Green Version]
- Gao, F.; Griffin, N.; Faulkner, S.; Rowe, C.W.; Williams, L.; Roselli, S.; Thorne, R.F.; Ferdoushi, A.; Jobling, P.; Walker, M.M.; et al. The neurotrophic tyrosine kinase receptor TrkA and its ligand NGF are increased in squamous cell carcinomas of the lung. Sci. Rep. 2018, 8, 8135. [Google Scholar] [CrossRef] [Green Version]
- Vaishnavi, A.; Capelletti, M.; Le, A.T.; Kako, S.; Butaney, M.; Ercan, D.; Mahale, S.; Davies, K.D.; Aisner, D.L.; Pilling, A.B.; et al. Oncogenic and drug-sensitive NTRK1 rearrangements in lung cancer. Nat. Med. 2013, 19, 1469–1472. [Google Scholar] [CrossRef] [Green Version]
- Vaishnavi, A.; Le, A.T.; Doebele, R.C. TRKing down an old oncogene in a new era of targeted therapy. Cancer Discov. 2015, 5, 25–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bhangoo, M.S.; Sigal, D. TRK Inhibitors: Clinical Development of Larotrectinib. Curr. Oncol. Rep. 2019, 21, 14. [Google Scholar] [CrossRef] [PubMed]
- Cocco, E.; Scaltriti, M.; Drilon, A. NTRK fusion-positive cancers and TRK inhibitor therapy. Nat. Rev. Clin. Oncol. 2018, 15, 731–747. [Google Scholar] [CrossRef]
- Drilon, A.; Laetsch, T.W.; Kummar, S.; DuBois, S.G.; Lassen, U.N.; Demetri, G.D.; Nathenson, M.; Doebele, R.C.; Farago, A.F.; Pappo, A.S.; et al. Efficacy of Larotrectinib in TRK Fusion-Positive Cancers in Adults and Children. N. Engl. J. Med. 2018, 378, 731–739. [Google Scholar] [CrossRef]
- Descamps, S.; Pawlowski, V.; Revillion, F.; Hornez, L.; Hebbar, M.; Boilly, B.; Hondermarck, H.; Peyrat, J.P. Expression of nerve growth factor receptors and their prognostic value in human breast cancer. Cancer Res. 2001, 61, 4337–4340. [Google Scholar]
- Com, E.; Lagadec, C.; Page, A.; El Yazidi-Belkoura, I.; Slomianny, C.; Spencer, A.; Hammache, D.; Rudkin, B.B.; Hondermarck, H. Nerve growth factor receptor TrkA signaling in breast cancer cells involves Ku70 to prevent apoptosis. Mol. Cell Proteom. 2007, 6, 1842–1854. [Google Scholar] [CrossRef] [Green Version]
- Demont, Y.; Corbet, C.; Page, A.; Ataman-Onal, Y.; Choquet-Kastylevsky, G.; Fliniaux, I.; Le Bourhis, X.; Toillon, R.A.; Bradshaw, R.A.; Hondermarck, H. Pro-nerve growth factor induces autocrine stimulation of breast cancer cell invasion through tropomyosin-related kinase A (TrkA) and sortilin protein. J. Biol. Chem. 2012, 287, 1923–1931. [Google Scholar] [CrossRef] [Green Version]
- Descamps, S.; Toillon, R.A.; Adriaenssens, E.; Pawlowski, V.; Cool, S.M.; Nurcombe, V.; Le Bourhis, X.; Boilly, B.; Peyrat, J.P.; Hondermarck, H. Nerve growth factor stimulates proliferation and survival of human breast cancer cells through two distinct signaling pathways. J. Biol. Chem. 2001, 276, 17864–17870. [Google Scholar] [CrossRef] [Green Version]
- Davidson, B.; Reich, R.; Lazarovici, P.; Ann Florenes, V.; Nielsen, S.; Nesland, J.M. Altered expression and activation of the nerve growth factor receptors TrkA and p75 provide the first evidence of tumor progression to effusion in breast carcinoma. Breast Cancer Res. Treat. 2004, 83, 119–128. [Google Scholar] [CrossRef]
- Youssef, G.; Gillett, C.; Agbaje, O.; Crompton, T.; Montano, X. Phosphorylation of NTRK1 at Y674/Y675 induced by TP53-dependent repression of PTPN6 expression: A potential novel prognostic marker for breast cancer. Mod. Pathol. An. Off. J. United States Can. Acad. Pathol. Inc. 2014, 27, 361–374. [Google Scholar] [CrossRef] [PubMed]
- Hondermarck, H. Neurotrophins and their receptors in breast cancer. Cytokine Growth Factor Rev. 2012, 23, 357–365. [Google Scholar] [CrossRef]
- Faulkner, S.; Roselli, S.; Demont, Y.; Pundavela, J.; Choquet, G.; Leissner, P.; Oldmeadow, C.; Attia, J.; Walker, M.M.; Hondermarck, H. ProNGF is a potential diagnostic biomarker for thyroid cancer. Oncotarget 2016, 7, 28488–28497. [Google Scholar] [CrossRef]
- Adriaenssens, E.; Vanhecke, E.; Saule, P.; Mougel, A.; Page, A.; Romon, R.; Nurcombe, V.; Le Bourhis, X.; Hondermarck, H. Nerve growth factor is a potential therapeutic target in breast cancer. Cancer Res. 2008, 68, 346–351. [Google Scholar] [CrossRef] [Green Version]
- Roselli, S.; Pundavela, J.; Demont, Y.; Faulkner, S.; Keene, S.; Attia, J.; Jiang, C.C.; Zhang, X.D.; Walker, M.M.; Hondermarck, H. Sortilin is associated with breast cancer aggressiveness and contributes to tumor cell adhesion and invasion. Oncotarget 2015, 6, 10473–10486. [Google Scholar] [CrossRef] [Green Version]
- Cerami, E.; Gao, J.; Dogrusoz, U.; Gross, B.E.; Sumer, S.O.; Aksoy, B.A.; Jacobsen, A.; Byrne, C.J.; Heuer, M.L.; Larsson, E.; et al. The cBio cancer genomics portal: An open platform for exploring multidimensional cancer genomics data. Cancer Discov. 2012, 2, 401–404. [Google Scholar] [CrossRef] [Green Version]
- Cancer Genome Atlas, N. Comprehensive molecular portraits of human breast tumours. Nature 2012, 490, 61–70. [Google Scholar] [CrossRef] [Green Version]
- Hutter, C.; Zenklusen, J.C. The Cancer Genome Atlas: Creating Lasting Value beyond Its Data. Cell 2018, 173, 283–285. [Google Scholar] [CrossRef]
- Watson, F.L.; Porcionatto, M.A.; Bhattacharyya, A.; Stiles, C.D.; Segal, R.A. TrkA glycosylation regulates receptor localization and activity. J. Neurobiol. 1999, 39, 323–336. [Google Scholar] [CrossRef]
- Albaugh, P.; Fan, Y.; Mi, Y.; Sun, F.; Adrian, F.; Li, N.; Jia, Y.; Sarkisova, Y.; Kreusch, A.; Hood, T.; et al. Discovery of GNF-5837, a Selective TRK Inhibitor with Efficacy in Rodent Cancer Tumor Models. ACS Med. Chem. Lett. 2012, 3, 140–145. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.; Wang, H.; Chen, Y.; Wang, M.; Ding, G.; Li, T. Trk inhibitor GNF5837 suppresses the tumor growth, survival and migration of renal cell carcinoma. Oncol. Rep. 2019, 42, 2039–2048. [Google Scholar] [PubMed]
- Tanner, M.; Kapanen, A.I.; Junttila, T.; Raheem, O.; Grenman, S.; Elo, J.; Elenius, K.; Isola, J. Characterization of a novel cell line established from a patient with Herceptin-resistant breast cancer. Mol. Cancer Ther. 2004, 3, 1585–1592. [Google Scholar] [PubMed]
- Smith, S.E.; Mellor, P.; Ward, A.K.; Kendall, S.; McDonald, M.; Vizeacoumar, F.S.; Vizeacoumar, F.J.; Napper, S.; Anderson, D.H. Molecular characterization of breast cancer cell lines through multiple omic approaches. Breast Cancer Res. 2017, 19, 65. [Google Scholar] [CrossRef] [Green Version]
- Kao, J.; Salari, K.; Bocanegra, M.; Choi, Y.L.; Girard, L.; Gandhi, J.; Kwei, K.A.; Hernandez-Boussard, T.; Wang, P.; Gazdar, A.F.; et al. Molecular profiling of breast cancer cell lines defines relevant tumor models and provides a resource for cancer gene discovery. PLoS ONE 2009. [Google Scholar] [CrossRef]
- Gygi, S.P.; Rochon, Y.; Franza, B.R.; Aebersold, R. Correlation between protein and mRNA abundance in yeast. Mol. Cell. Biol. 1999, 19, 1720–1730. [Google Scholar] [CrossRef] [Green Version]
- Vogel, C.; Marcotte, E.M. Insights into the regulation of protein abundance from proteomic and transcriptomic analyses. Nat. Rev. Genet. 2012, 13, 227–232. [Google Scholar] [CrossRef]
- Liu, Y.; Beyer, A.; Aebersold, R. On the Dependency of Cellular Protein Levels on mRNA Abundance. Cell 2016, 165, 535–550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, B.; Wang, J.; Wang, X.; Zhu, J.; Liu, Q.; Shi, Z.; Chambers, M.C.; Zimmerman, L.J.; Shaddox, K.F.; Kim, S.; et al. Proteogenomic characterization of human colon and rectal cancer. Nature 2014, 513, 382–387. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Faulkner, S.; Griffin, N.; Rowe, C.W.; Jobling, P.; Lombard, J.M.; Oliveira, S.M.; Walker, M.M.; Hondermarck, H. Nerve growth factor and its receptor tyrosine kinase TrkA are overexpressed in cervical squamous cell carcinoma. FASEB Bioadv. 2020, 2, 398–408. [Google Scholar] [CrossRef] [PubMed]
- Tagliabue, E.; Castiglioni, F.; Ghirelli, C.; Modugno, M.; Asnaghi, L.; Somenzi, G.; Melani, C.; Menard, S. Nerve growth factor cooperates with p185(HER2) in activating growth of human breast carcinoma cells. J. Biol. Chem. 2000, 275, 5388–5394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Louie, E.; Chen, X.F.; Coomes, A.; Ji, K.; Tsirka, S.; Chen, E.I. Neurotrophin-3 modulates breast cancer cells and the microenvironment to promote the growth of breast cancer brain metastasis. Oncogene 2013, 32, 4064–4077. [Google Scholar] [CrossRef] [PubMed] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TrkA Intensity | |||||
|---|---|---|---|---|---|
| Parameter | 0 | 1 | 2 | 3 | p-Value |
| All Cases (n = 366) | 260 (71%) | 27 (7%) | 48 (13%) | 31 (9%) | |
| Pathological Subtype | <0.0001 a | ||||
| DCIS (n = 50) | 38 (76%) | 12 (24%) | 0 (0%) | 0 (0%) | |
| ILC (n = 158) | 126 (80%) | 7 (4%) | 19 (12%) | 6 (4%) | |
| IDC (n = 158) | 96 (61%) | 8 (5%) | 29 (18%) | 25 (16%) | |
| Clinical Parameters in Invasive Carcinomas | |||||
| Patient Age(Years) | 0.7195 | ||||
| ≤50 (n = 184) | 130 (71%) | 7 (4%) | 30 (16%) | 17 (9%) | |
| >50 (n = 132) | 92 (70%) | 8 (6%) | 18 (14%) | 14 (10%) | |
| Lymph Node Status(N) | 0.0696 | ||||
| Negative (n = 163) | 121 (74%) | 9 (6%) | 21 (13%) | 12 (7%) | |
| Positive (n = 133) | 82 (62%) | 6 (5%) | 27 (20%) | 18 (13%) | |
| HER2 | 0.0027 a | ||||
| HER2 negative (n = 250) | 188 (75%) | 10 (4%) | 31 (12%) | 21 (9%) | |
| HER2 positive (n = 66) | 34 (51%) | 5 (8%) | 17 (26%) | 10 (15%) | |
| Estrogen Receptor | 0.2267 | ||||
| ER negative (n = 181) | 129 (71%) | 5 (3%) | 27 (15%) | 20 (11%) | |
| ER positive (n = 135) | 93 (69%) | 10 (7%) | 21 (16%) | 11 (8%) | |
| Progesterone Receptor | 0.0261 a | ||||
| PR negative (n = 207) | 152 (73%) | 6 (3%) | 26 (13%) | 23 (11%) | |
| PR positive (n = 109) | 70 (64%) | 9 (8%) | 22 (20%) | 8 (8%) | |
| Tumour Size(T) | 0.6360 | ||||
| 1 + 2 (n = 238) | 166 (70%) | 14 (6%) | 36 (15%) | 22 (9%) | |
| 3 + 4 (n = 58) | 38 (65%) | 1 (2%) | 11 (19%) | 8 (14%) | |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Griffin, N.; Marsland, M.; Roselli, S.; Oldmeadow, C.; Attia, J.; Walker, M.M.; Hondermarck, H.; Faulkner, S. The Receptor Tyrosine Kinase TrkA Is Increased and Targetable in HER2-Positive Breast Cancer. Biomolecules 2020, 10, 1329. https://doi.org/10.3390/biom10091329
Griffin N, Marsland M, Roselli S, Oldmeadow C, Attia J, Walker MM, Hondermarck H, Faulkner S. The Receptor Tyrosine Kinase TrkA Is Increased and Targetable in HER2-Positive Breast Cancer. Biomolecules. 2020; 10(9):1329. https://doi.org/10.3390/biom10091329
Chicago/Turabian StyleGriffin, Nathan, Mark Marsland, Severine Roselli, Christopher Oldmeadow, John Attia, Marjorie M. Walker, Hubert Hondermarck, and Sam Faulkner. 2020. "The Receptor Tyrosine Kinase TrkA Is Increased and Targetable in HER2-Positive Breast Cancer" Biomolecules 10, no. 9: 1329. https://doi.org/10.3390/biom10091329