Identification and Characterization of a Novel Thermostable and Salt-Tolerant β-1,3 Xylanase from Flammeovirga pacifica Strain WPAGA1



Abstract
:1. Introduction
2. Materials and Methods
2.1. Preparation of β-1,3 Xylan and β-1,3 Oligosaccharides
2.2. Plasmid Construction
2.3. SDS-Polyacrylamide Gel Electrophoresis (SDS-PAGE) with Precast Gels
2.4. Expression and Purification of Xyl88 Recombinant Protein
2.5. Enzymatic Activity Measurement
2.6. Enzymatic Property Measurement
2.7. TLC analysis of rXyl88 Hydrolysate
2.8. Protein Sequence-Structure Analysis
3. Results
3.1. Expression and Purification of Recombinant Protein rXyl88
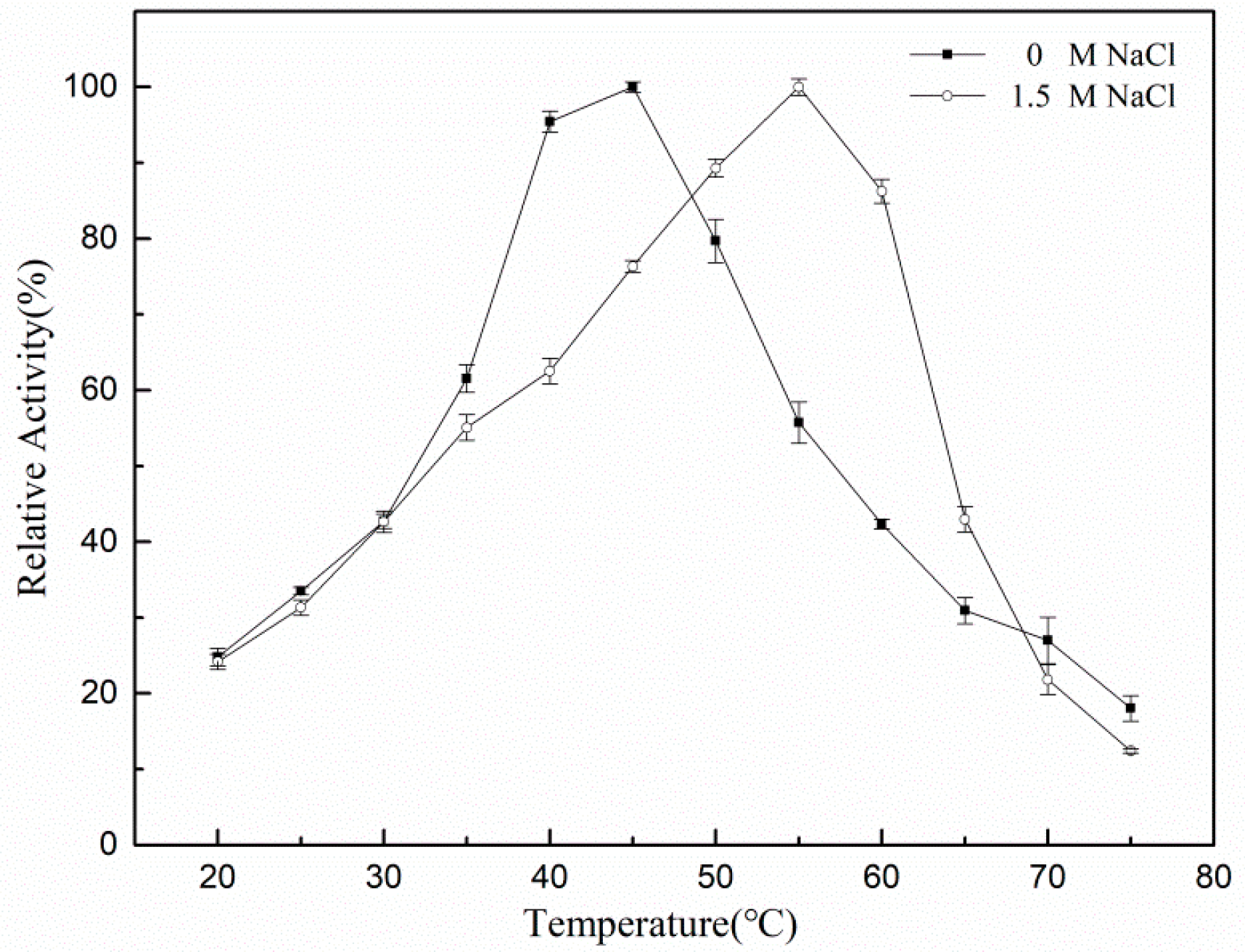
3.2. Measurement of rXyl88 Enzymatic Properties
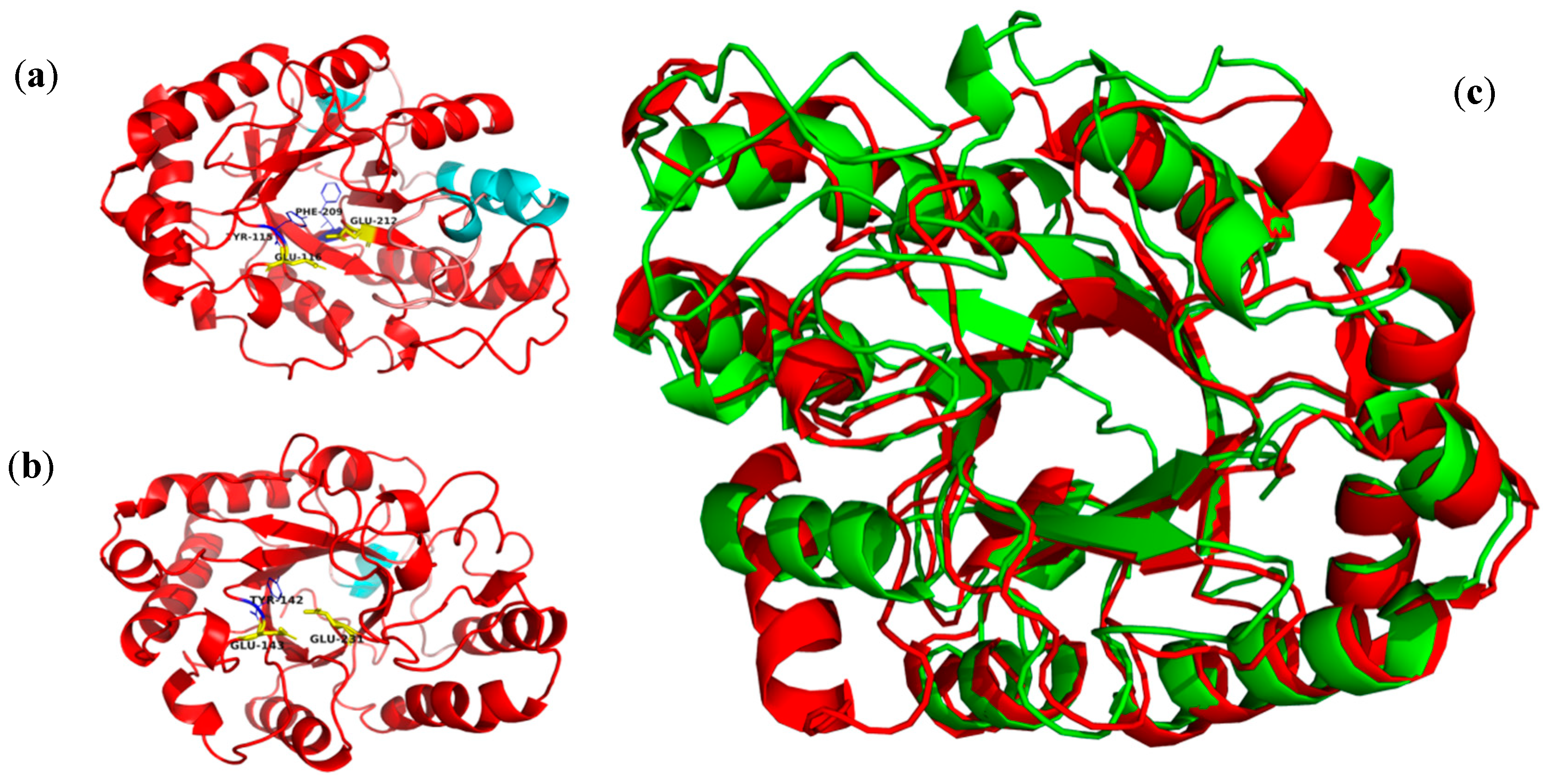
3.3. Thermophilic Salt-Tolerant Mechanism of β-1,3 Xylanase
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
References
- Hehemann, J.H.; Boraston, A.B.; Czjzek, M. A sweet new wave: Structures and mechanisms of enzymes that digest polysaccharides from marine algae. Curr. Opin. Struct. Biol. 2014, 28, 77–86. [Google Scholar] [CrossRef] [PubMed]
- Park, D.; Jagtap, S.; Nair, S.K. Structure of a PL17 family alginate lyase demonstrates functional similarities among exotype depolymerases. J. Biol. Chem. 2014, 289, 8645–8655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Araki, C. Acetylation of agar like substance of Gelidium amansii. J. Chem. Soc. 1937, 58, 1338–1350. [Google Scholar]
- Tobacman, J.K. Review of harmful gastrointestinal effects of carrageenan in animal experiments. Environ. Health Perspect. 2001, 109, 983–994. [Google Scholar] [CrossRef] [PubMed]
- Laureano-Perez, L.; Teymouri, F.; Alizadeh, H.; Dale, B.E. Understanding factors that limit enzymatic hydrolysis of biomass. Appl. Biochem. Biotechnol. 2005, 124, 1081–1099. [Google Scholar] [CrossRef]
- Rahman, M.A.; Halfar, J. First evidence of chitin in calcified coralline algae: New insights into the calcification process of Clathromorphum compactum. Sci. Rep. 2014, 4, 6162. [Google Scholar] [CrossRef]
- Muzzarelli, R.A. Chitin; Elsevier: Amsterdam, The Netherlands, 2013. [Google Scholar]
- Scheller, H.V.; Ulvskov, P. Hemicelluloses. Annu. Rev. Plant Biol. 2010, 61, 263–289. [Google Scholar] [CrossRef] [PubMed]
- Goddard-Borger, E.D.; Sakaguchi, K.; Reitinger, S.; Watanabe, N.; Ito, M.; Withers, S.G.J. Mechanistic insights into the 1,3-xylanases: Useful enzymes for manipulation of algal biomass. J. Am. Chem. Soc. 2012, 134, 3895–3902. [Google Scholar] [CrossRef]
- Macartain, P.; Gill, C.I.R.; Brooks, M.; Campbell, R.; Rowland, I.R. Nutritional value of edible seaweeds. Nutr. Rev. 2007, 65, 535. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, V.T.; Ueng, J.P.; Tsai, G.J. Proximate composition, total phenolic content, and antioxidant activity of seagrape (Caulerpa lentillifera). J. Food Sci. 2011, 76, C950–C958. [Google Scholar] [CrossRef]
- Coughlan, M.; Hazlewood, G.P. Β-1, 4-smallcap D-Xylan-degrading enzyme systems: Biochemistry, molecular biology and applications. Biotechnol. Appl. Biochem. 1993, 17, 259–289. [Google Scholar] [PubMed]
- Polizeli, M.; Rizzatti, A.; Monti, R.; Terenzi, H.; Jorge, J.A.; Amorim, D. Xylanases from fungi: Properties and industrial applications. Appl. Microbiol. Biotechnol. 2005, 67, 577–591. [Google Scholar] [CrossRef] [PubMed]
- Maeda, R.; Ida, T.; Ihara, H.; Sakamoto, T. Induction of apoptosis in MCF-7 cells by beta-1,3-xylooligosaccharides prepared from Caulerpa lentillifera. Biosci. Biotechnol. Biochem. 2012, 76, 1032–1034. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Umemoto, Y.; Araki, S.T. d-Xylose Isomerase from a Marine Bacterium, Vibrio sp. Strain XY-214, and d-Xylulose Production from β-1,3-Xylan. Mar. Biotechnol. 2012, 14, 10–20. [Google Scholar] [CrossRef]
- Gírio, F.M.; Fonseca, C.; Carvalheiro, F.; Duarte, L.C.; Marques, S.; Bogel-Łukasik, R. Hemicelluloses for fuel ethanol: A review. Bioresour. Technol. 2010, 101, 4775–4800. [Google Scholar] [CrossRef]
- Granström, T.B.; Izumori, K.; Leisola, M. A rare sugar xylitol. Part II: Biotechnological production and future applications of xylitol. Appl. Microbiol. Biotechnol. 2007, 74, 273. [Google Scholar]
- Tanzer, J. Xylitol chewing gum and dental caries. Int. Dent. J. 1995, 45, 65–76. [Google Scholar]
- Iriki, Y.; Suzuki, T.; Nisizawa, K.; Miwa, T. Xylan of siphonaceous green algae. Nature 1960, 187, 82–83. [Google Scholar] [CrossRef]
- Aoki, Y.; Kamei, Y. Preparation of recombinant polysaccharide-degrading enzymes from the marine bacterium, Pseudomonas sp. ND137 for the production of protoplasts of Porphyra yezoensis. Eur. J. Phycol. 2006, 41, 321–328. [Google Scholar] [CrossRef] [Green Version]
- Hashimoto, H.; Tamai, Y.; Okazaki, F.; Tamaru, Y.; Shimizu, T.; Araki, T.; Sato, M. The first crystal structure of a family 31 carbohydrate-binding module with affinity to β-1,3-xylan. FEBS Lett. 2005, 579, 4324–4328. [Google Scholar] [CrossRef] [Green Version]
- Kiyohara, M.; Sakaguchi, K.; Yamaguchi, K.; Araki, T.; Ito, M. Characterization and application of carbohydrate-binding modules of β-1,3-xylanase XYL4. J. Biochem. 2009, 146, 633–641. [Google Scholar] [CrossRef] [PubMed]
- Yamaura, I.; Matsumoto, T.; Funatsu, M.; Mukai, E. Purification and some properties of endo-1,3-beta-D-xylanase from Pseudomonas sp. PT-5. Agric. Biol. Chem. 1990, 54, 921–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dubois, M.; Gilles, K.A.; Hamilton, J.K.; Rebers, P.T.; Smith, F. Colorimetric method for determination of sugars and related substances. Anal. Chem. 1956, 28, 350–356. [Google Scholar] [CrossRef]
- Kiyohara, M.; Hama, Y.; Yamaguchi, K.; Ito, M.J. Structure of β-1,3-xylooligosaccharides generated from Caulerpa racemosa var. laete-virens β-1,3-xylan by the action of β-1,3-xylanase. J. Biochem. 2006, 140, 369–373. [Google Scholar] [CrossRef] [PubMed]
- Kruger, N.J. The Bradford method for protein quantitation. In The Protein Protocols Handbook; Springer: Berlin/Heidelberg, Germany, 2009; pp. 17–24. [Google Scholar]
- Miller, G.L. Use of dinitrosalicylic acid reagent for determination of reducing sugar. Anal. Chem. 1959, 31, 426–428. [Google Scholar] [CrossRef]
- Bailey, R.; Bourne, E.J. Colour reactions given by sugars and diphenylamine-aniline spray reagents on paper chromatograms. J. Chromatogr. 1960, 4, 206–213. [Google Scholar] [CrossRef]
- Geer, L.Y.; Domrachev, M.; Lipman, D.J.; Bryant, S.H. CDART: Protein homology by domain architecture. Genome Res. 2002, 12, 1619–1623. [Google Scholar] [CrossRef] [Green Version]
- Arnold, K.; Bordoli, L.; Kopp, J.; Schwede, T.J.B. The SWISS-MODEL workspace: A web-based environment for protein structure homology modelling. Bioinformatics 2006, 22, 195–201. [Google Scholar] [CrossRef] [Green Version]
- Roy, A.; Kucukural, A.; Zhang, Y. I-TASSER: A unified platform for automated protein structure and function prediction. Nat. Protoc. 2010, 5, 725–738. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, D.E.; Chivian, D.; Baker, D. Protein structure prediction and analysis using the Robetta server. Nucleic Acids Res. 2004, 32, 526–531. [Google Scholar] [CrossRef] [Green Version]
- Chen, V.B.; Arendall, W.B.; Headd, J.J.; Keedy, D.A.; Immormino, R.M.; Kapral, G.J.; Murray, L.W.; Richardson, J.S.; Richardson, D.C. MolProbity: All-atom structure validation for macromolecular crystallography. Acta Crystallogr. 2010, 66, 12–21. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Delano, W. Pymol Molecular Graphics System: An open-source molecular graphics tool. Ccp4 Newsl. Protein 2002, 40, 82–92. [Google Scholar]
- Pettersen, E.F.; Goddard, T.D.; Huang, C.C.; Couch, G.S.; Greenblatt, D.M.; Meng, E.C.; Ferrin, T.E. UCSF Chimera-a visualization system for exploratory research and analysis. J. Comput. Chem. 2004, 25, 1605–1612. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Damiano, P.; Giovanni, M.; Tosatto, S.C.E. The RING 2.0 web server for high quality residue interaction networks. Nucleic Acids Res. 2016, 44, W367. [Google Scholar]
- Sakaguchi, K.; Kiyohara, M.; Watanabe, N.; Yamaguchi, K.; Ito, M.; Kawamura, T.; Tanaka, I. Preparation and preliminary X-ray analysis of the catalytic module of beta-1,3-xylanase from the marine bacterium Vibrio sp. AX-4. Acta Crystallogr. 2004, 60, 1470–1472. [Google Scholar]
- Phillips, J.C.; Braun, R.; Wang, W.; Gumbart, J.; Tajkhorshid, E.; Villa, E.; Chipot, C.; Skeel, R.D.; Kale, L.; Schulten, K. Scalable molecular dynamics with NAMD. J. Comput. Chem. 2005, 26, 1781–1802. [Google Scholar] [CrossRef] [Green Version]
- Humphrey, W.; Dalke, A.; Schulten, K. VMD: Visual molecular dynamics. J. Mol. Graph 1996, 14, 33–38. [Google Scholar] [CrossRef]
- Lebowitz, J.; Percus, J.; Verlet, L. Ensemble dependence of fluctuations with application to machine computations. Phys. Rev. 1967, 153, 250. [Google Scholar] [CrossRef]
- Araki, T.; Tani, S.; Maeda, K.; Hashikawa, S.; Nakagawa, H.; Morishita, T.J.B. Purification and characterization of β-1,3-xylanase from a marine bacterium, Vibrio sp. XY-214. Biosci. Biotechnol. Biochem. 1999, 63, 2017–2019. [Google Scholar] [CrossRef]
- Araki, T.; Inoue, N.; Morishita, T. Purification and characterization of beta-1,3-xylanase from a marine bacterium, Alcaligenes sp. XY-234. J. Gen. Appl. Microbiol. 1998, 44, 269–274. [Google Scholar] [CrossRef] [Green Version]
- Kiyohara, M.; Sakaguchi, K.; Yamaguchi, K.; Araki, T.; Nakamura, T.; Ito, M. Molecular cloning and characterization of a novel beta-1,3-xylanase possessing two putative carbohydrate-binding modules from a marine bacterium Vibrio sp. strain AX-4. Biochem. J. 2005, 388, 949–957. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Okazaki, F.; Nakashima, N.; Ogino, C.; Tamaru, Y.; Kondo, A. Biochemical characterization of a thermostable β-1,3-xylanase from the hyperthermophilic eubacterium, Thermotoga neapolitana strain DSM 4359. Appl. Microbiol. Biotechnol. 2013, 97, 6749–6757. [Google Scholar] [CrossRef] [PubMed]
- Aloy, P.; Ceulemans, H.; Stark, A.; Russell, R.B. The Relationship Between Sequence and Interaction Divergence in Proteins. J. Mol. Biol. 2003, 332, 989–998. [Google Scholar] [CrossRef] [PubMed]
- Hamre, A.; Frøberg, E.; Eijsink, V.; Sørlie, M. Thermodynamics of tunnel formation upon substrate binding in a processive glycoside hydrolase. Arch. Biochem. Biophys. 2017, 620, 35–42. [Google Scholar] [CrossRef]
- Unni, S.; Huang, Y.; Hanson, R.M.; Tobias, M.; Krishnan, S.; Li, W.W.; Nielsen, J.E.; Baker, N.A. Web servers and services for electrostatics calculations with APBS and PDB2PQR. J. Comput. Chem. 2011, 32, 1488–1491. [Google Scholar] [CrossRef] [Green Version]
- Pucci, F.; Rooman, M. Physical and molecular bases of protein thermal stability and cold adaptation. Curr. Opin. Biotechnol. 2017, 42, 117–128. [Google Scholar] [CrossRef]
- Müllersantos, M.; de Souza, E.M.; Pedrosa, F.O.; Mitchell, D.A.; Longhi, S.; Carrière, F.; Canaan, S.; Krieger, N. First evidence for the salt-dependent folding and activity of an esterase from the halophilic archaea Haloarcula marismortui. Biochim. Et Biophys. Acta (Bba) Mol. Cell Biol. Lipids 2009, 1791, 719–729. [Google Scholar]
- Elcock, A.H.; Mccammon, J.A. Electrostatic contributions to the stability of halophilic proteins. J. Mol. Biol. 1998, 280, 731–748. [Google Scholar] [CrossRef]
- Zhang, T.; Datta, S.; Eichler, J.; Ivanova, N.; Axen, S.D.; Kerfeld, C.A.; Chen, F.; Kyrpides, N.; Hugenholtz, P.; Cheng, J.F. Identification of a haloalkaliphilic and thermostable cellulase with improved ionic liquid tolerance. Green Chem. 2011, 13, 2083–2090. [Google Scholar] [CrossRef] [Green Version]
- Klee, E.C.C.B. Calcium as a Cellular Regulator; Oxford University Press: New York, NY, USA, 1999. [Google Scholar]
- Sissi, C.; Palumbo, M. Effects of magnesium and related divalent metal ions in topoisomerase structure and function. Nucleic Acids Res. 2009, 37, 702–711. [Google Scholar] [CrossRef] [Green Version]
- Keowmaneechai, E.; McClements, D.J.; Chemistry, F. Influence of EDTA and citrate on physicochemical properties of whey protein-stabilized oil-in-water emulsions containing CaCl2. J. Agric. Food Chem. 2002, 50, 7145–7153. [Google Scholar] [CrossRef] [PubMed]
- Siglioccolo, A.; Paiardini, A.; Piscitelli, M.; Pascarella, S. Structural adaptation of extreme halophilic proteins through decrease of conserved hydrophobic contact surface. BMC Struct. Biol. 2011, 11, 50. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tompa, D.R.; Gromiha, M.M.; Saraboji, K. Contribution of main chain and side chain atoms and their locations to the stability of thermophilic proteins. J. Mol. Graph. Model. 2016, 64, 85–93. [Google Scholar] [CrossRef] [PubMed]













{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Metal Ion (10 mmol/L) | Rel Act. (%) | Metal Ion (10 mmol/L) | Rel act. (%) |
|---|---|---|---|
| Control | 100.00 | Cu2+ | 18.73 ± 0.73 |
| Na+ | 116.35 ± 2.13 | Ca2+ | 72.13 ± 0.62 |
| K+ | 99.12 ± 1.29 | Co2+ | 37.07 ± 0.59 |
| Zn2+ | 35.96 ± 1.74 | Ni2+ | 35.33 ± 0.84 |
| Mn2+ | 48.54 ± 2.41 | Fe3+ | 13.72 ± 0.25 |
| Ba2+ | 106.44 ± 3.37 | EDTA | 73.05 ± 2.59 |
| Mg2+ | 117.63 ± 4.16 |
| NaCl (mol·L−1) | Substrate | |||
|---|---|---|---|---|
| 0 | β-1,3-Xylan | 5.75 ± 0.96 | 517.54 ± 41.54 | 90.01 ± 7.22 |
| Glycol β-1,3-Xylan | 4.83 ± 0.64 | 1501.61 ± 88.41 | 310.89 ± 18.31 | |
| 1.5 | β-1,3-Xylan | 5.58 ± 0.88 | 436.68 ± 54.81 | 78.26 ± 9.82 |
| Glycol β-1,3-Xylan | 4.59 ± 0.91 | 1441.95 ± 130.99 | 314.15 ± 28.51 |
| 3D Structure | Nodes | Edges | Protein–Protein Hydrogen Bond | Salt Bridge | Π–π Stacking | Disulphide Bond | Van der Waals | π-Cation | Protein–Water Hydrogen Bond | |
|---|---|---|---|---|---|---|---|---|---|---|
| 310 K | 328 K | |||||||||
| rXYL4 | 303 | 613 | 326 | 8 | 29 | 1 | 249 | 0 | 259 | 274 |
| Xyl88 | 384 | 793 | 345 | 7 | 25 | 0 | 416 | 0 | 255 | 258 |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yi, Z.; Cai, Z.; Zeng, B.; Zeng, R.; Zhang, G. Identification and Characterization of a Novel Thermostable and Salt-Tolerant β-1,3 Xylanase from Flammeovirga pacifica Strain WPAGA1. Biomolecules 2020, 10, 1287. https://doi.org/10.3390/biom10091287
Yi Z, Cai Z, Zeng B, Zeng R, Zhang G. Identification and Characterization of a Novel Thermostable and Salt-Tolerant β-1,3 Xylanase from Flammeovirga pacifica Strain WPAGA1. Biomolecules. 2020; 10(9):1287. https://doi.org/10.3390/biom10091287
Chicago/Turabian StyleYi, Zhiwei, Zhengwen Cai, Bo Zeng, Runying Zeng, and Guangya Zhang. 2020. "Identification and Characterization of a Novel Thermostable and Salt-Tolerant β-1,3 Xylanase from Flammeovirga pacifica Strain WPAGA1" Biomolecules 10, no. 9: 1287. https://doi.org/10.3390/biom10091287