Multiple Screening of Pesticides Toxicity in Zebrafish and Daphnia Based on Locomotor Activity Alterations

 ,
,  ,
, 
Abstract
:1. Introduction
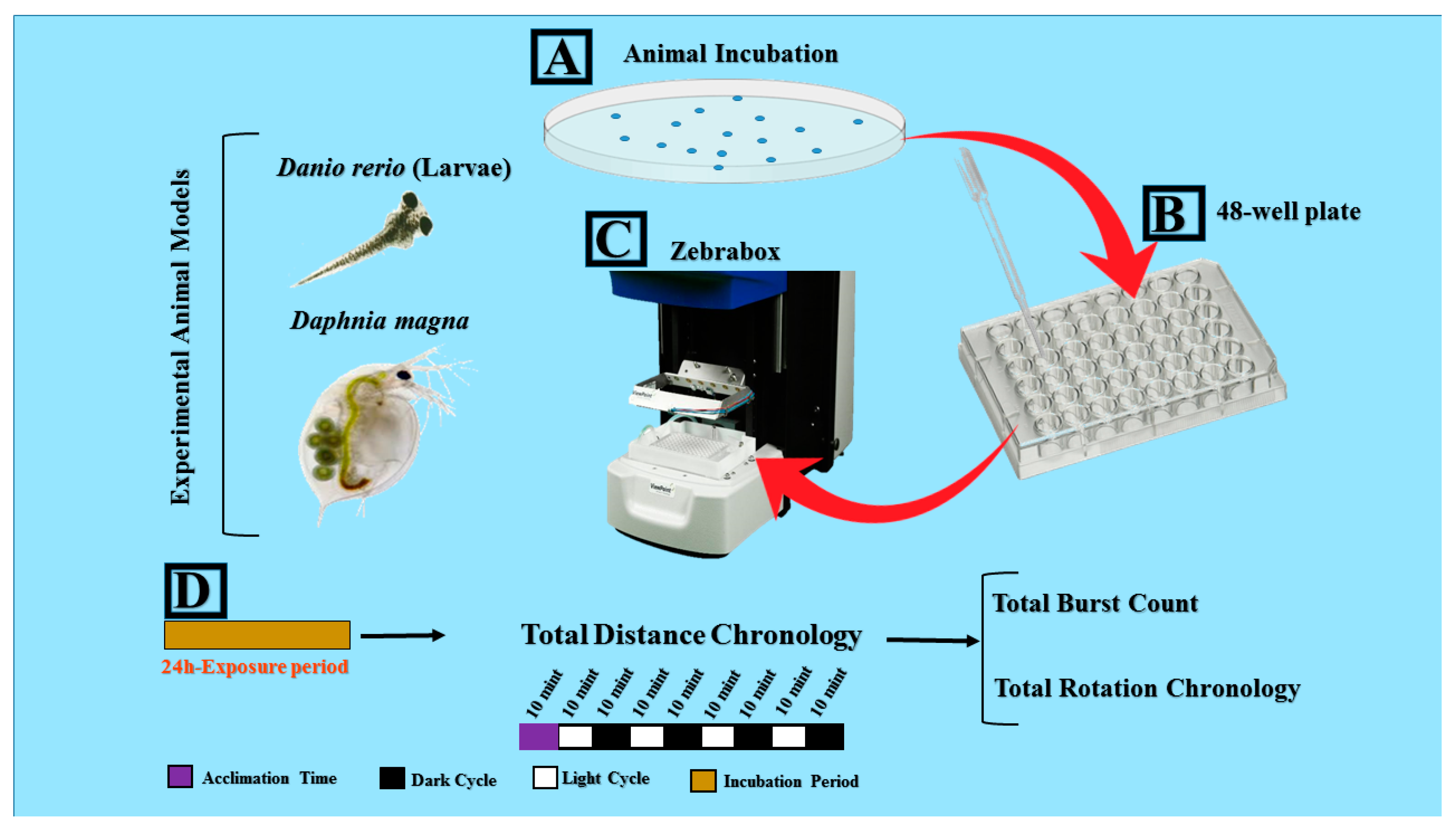
2. Materials and Methods
2.1. Zebrafish Larvae Rearing and Maintenance
2.2. Daphnia Rearing and Maintenance
2.3. Chemical Exposure
2.4. Automated Imaging of Swimming Behaviors Analysis
2.5. Statistical Analysis
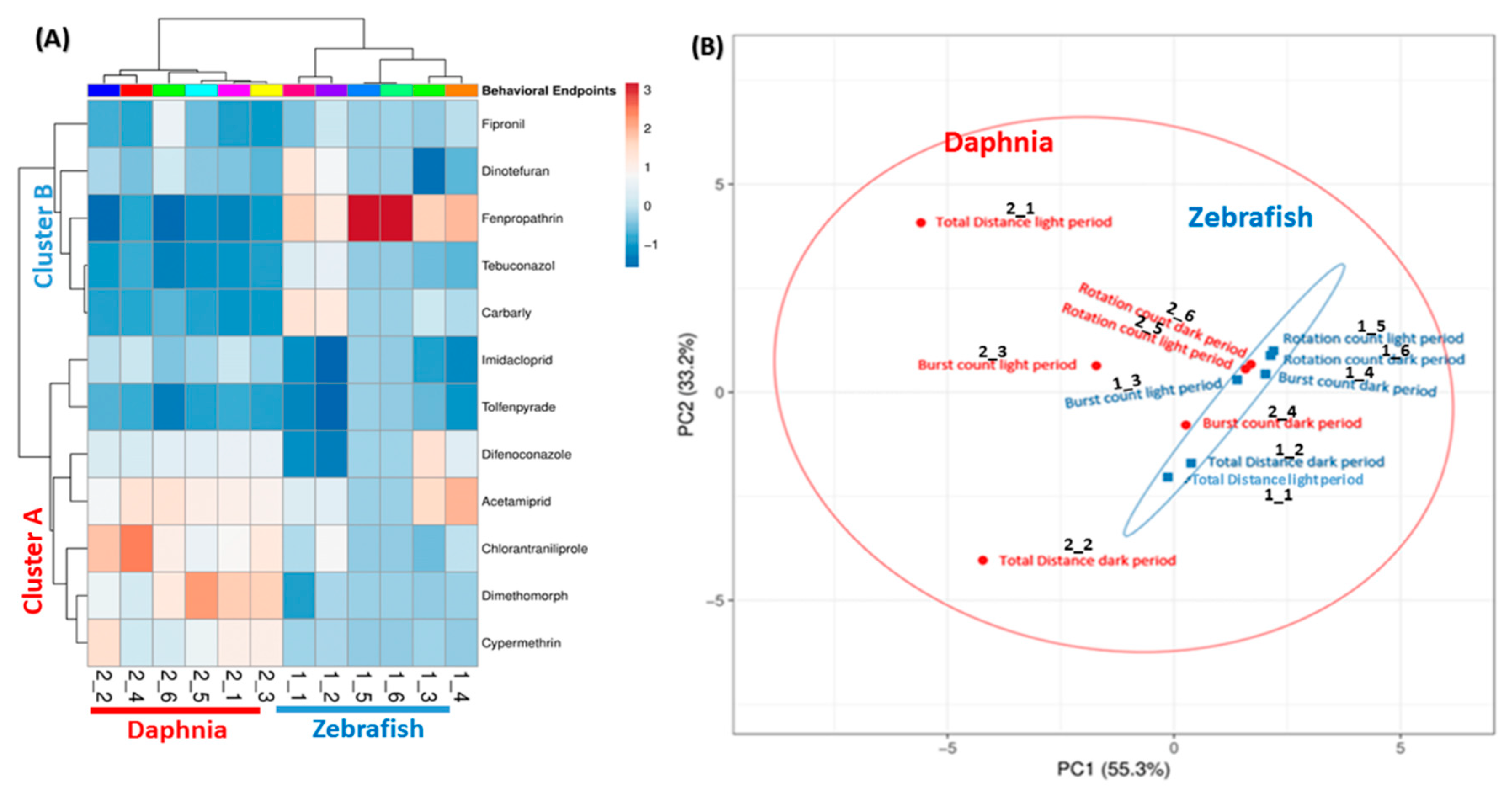
2.6. Principal Component Analysis (PCA), Heatmap, and Hierarchical Clustering Analysis
3. Results
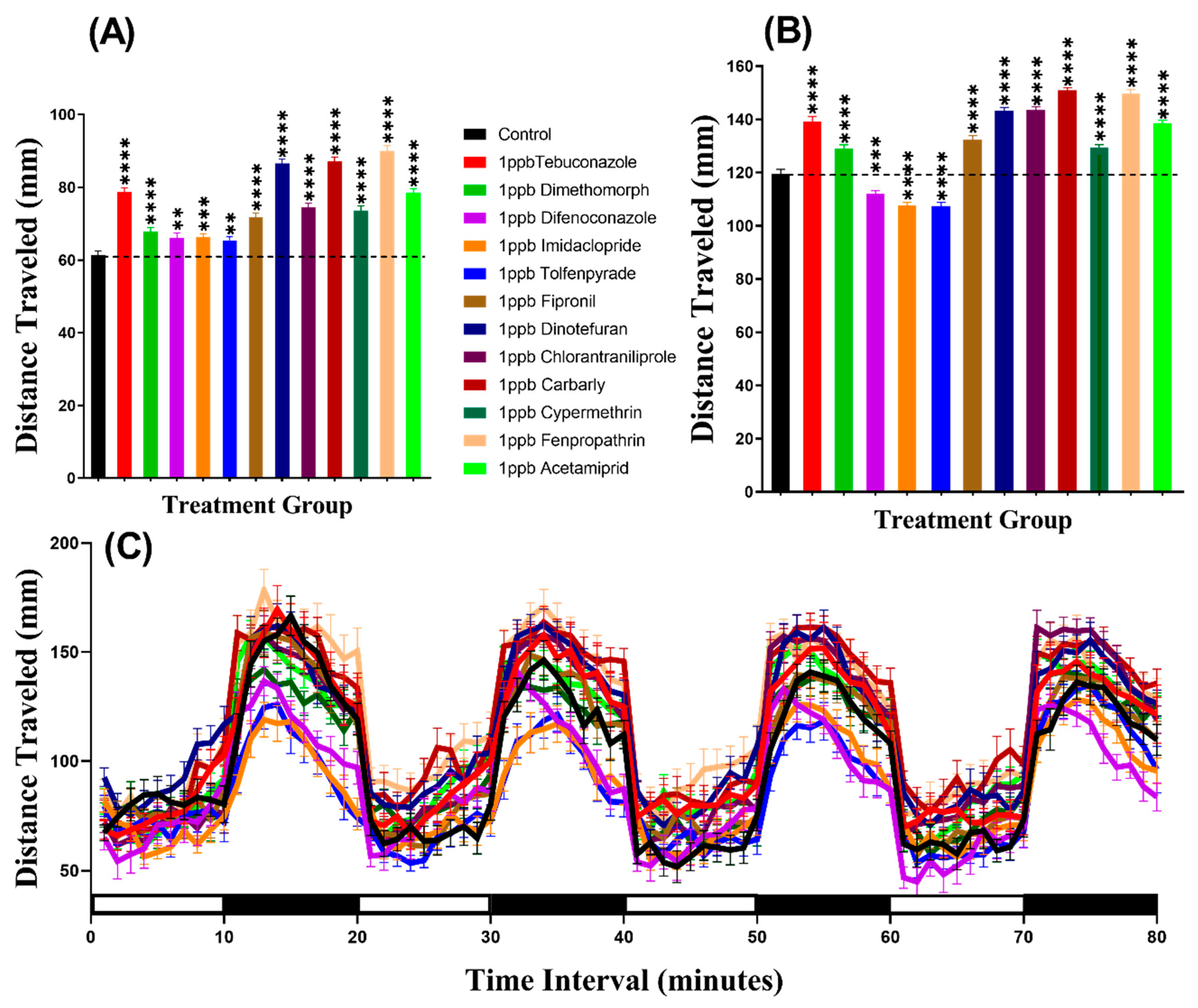
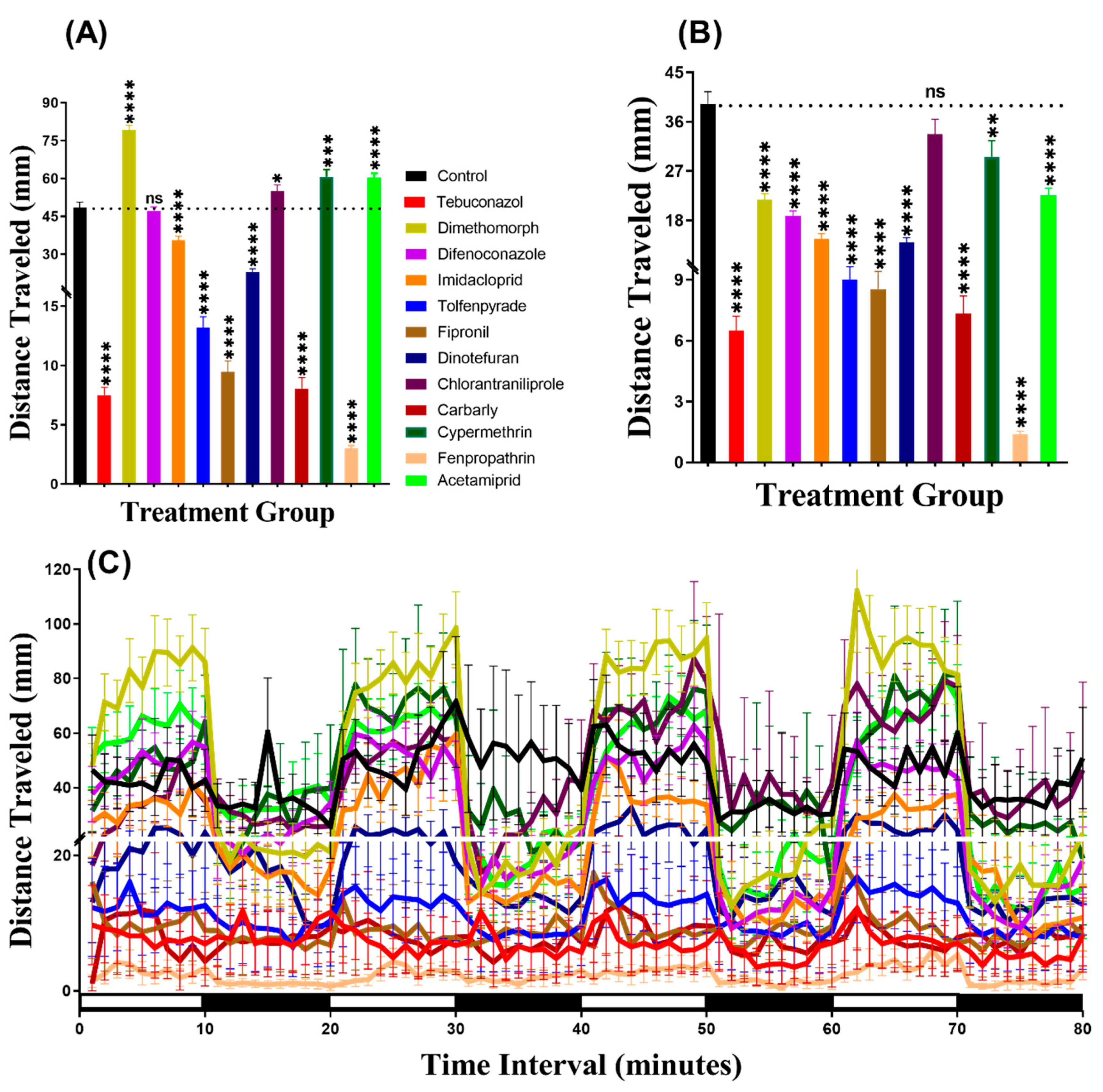
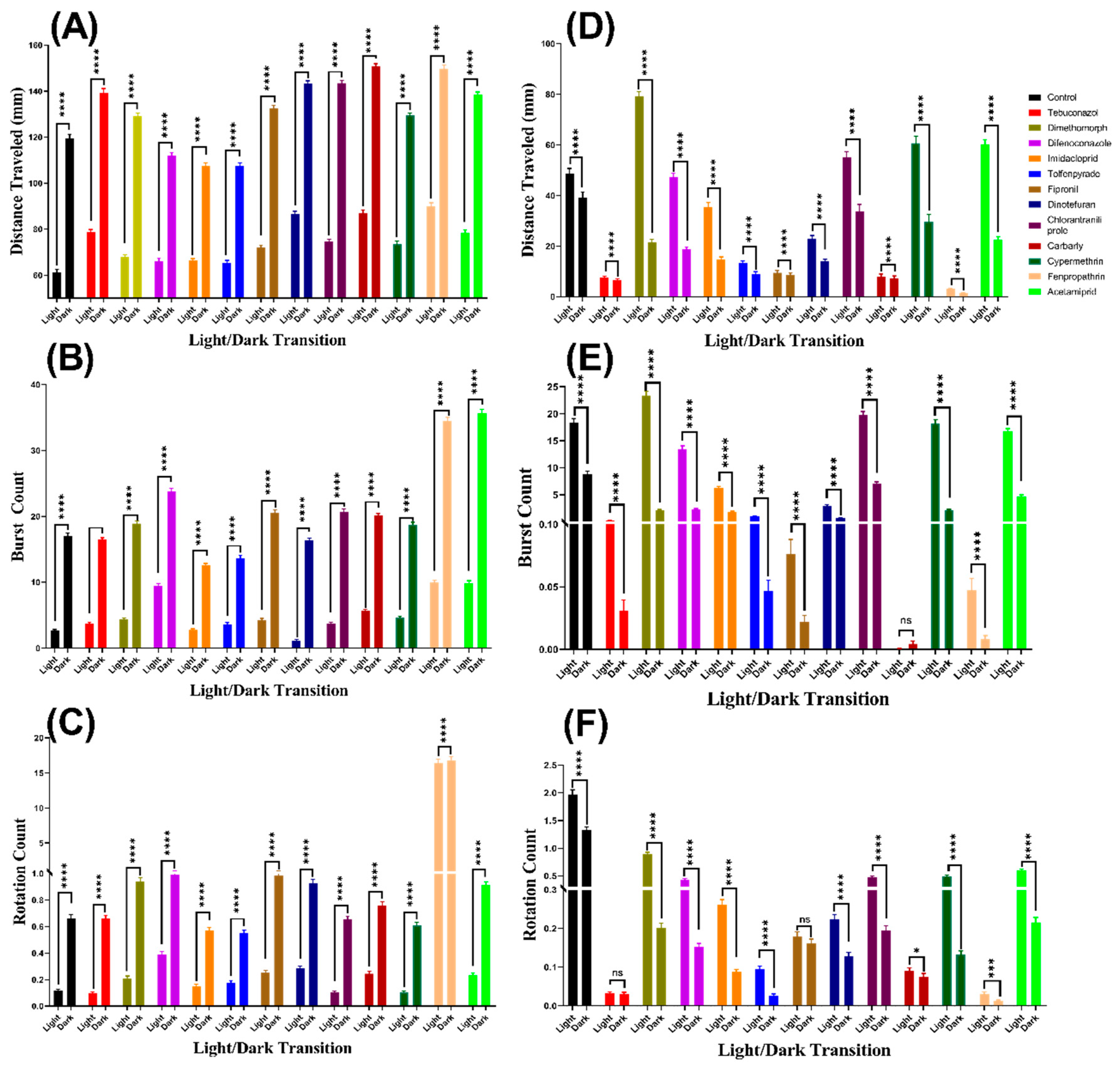
3.1. Total Distance Chronology for Zebrafish after Pesticide Exposure
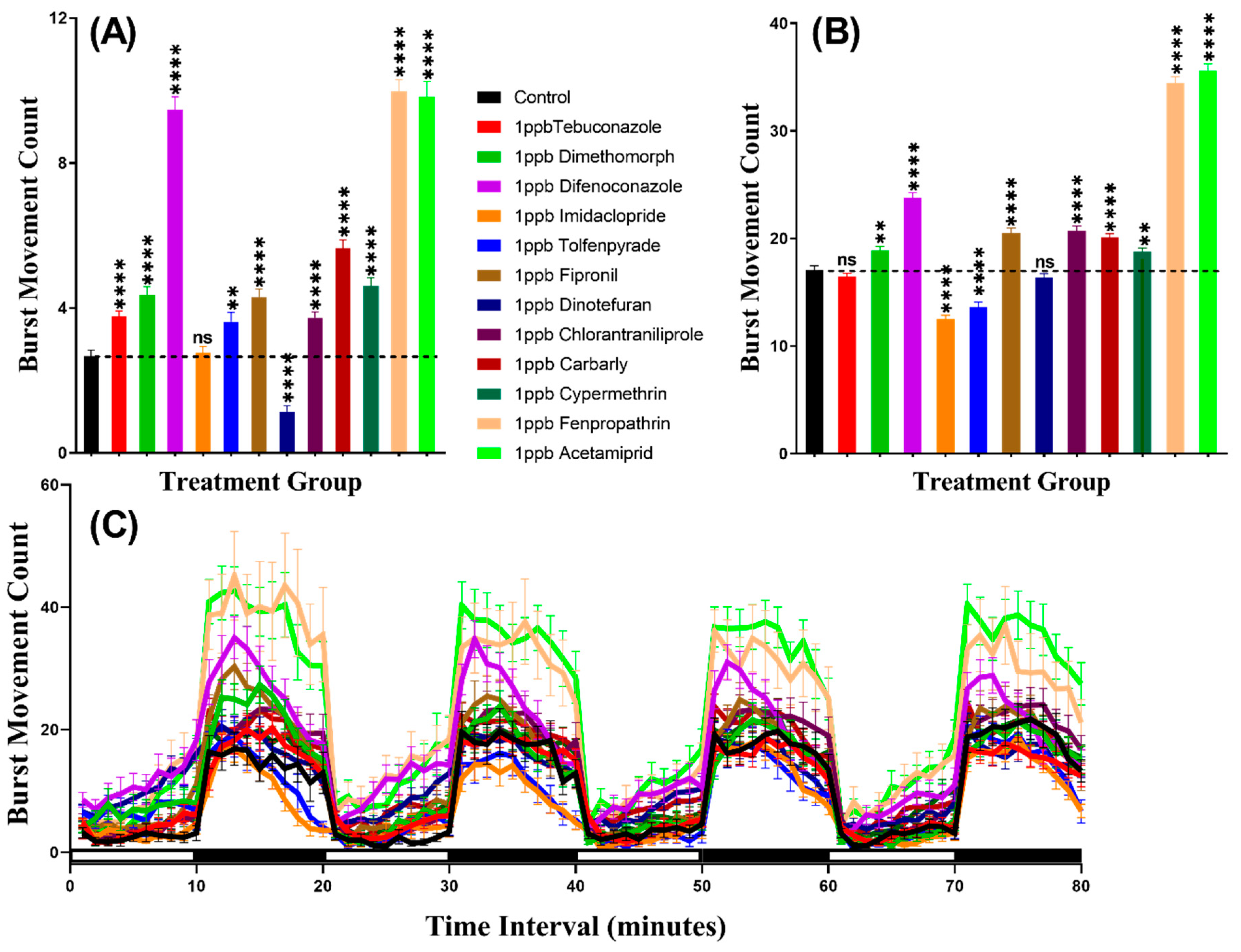
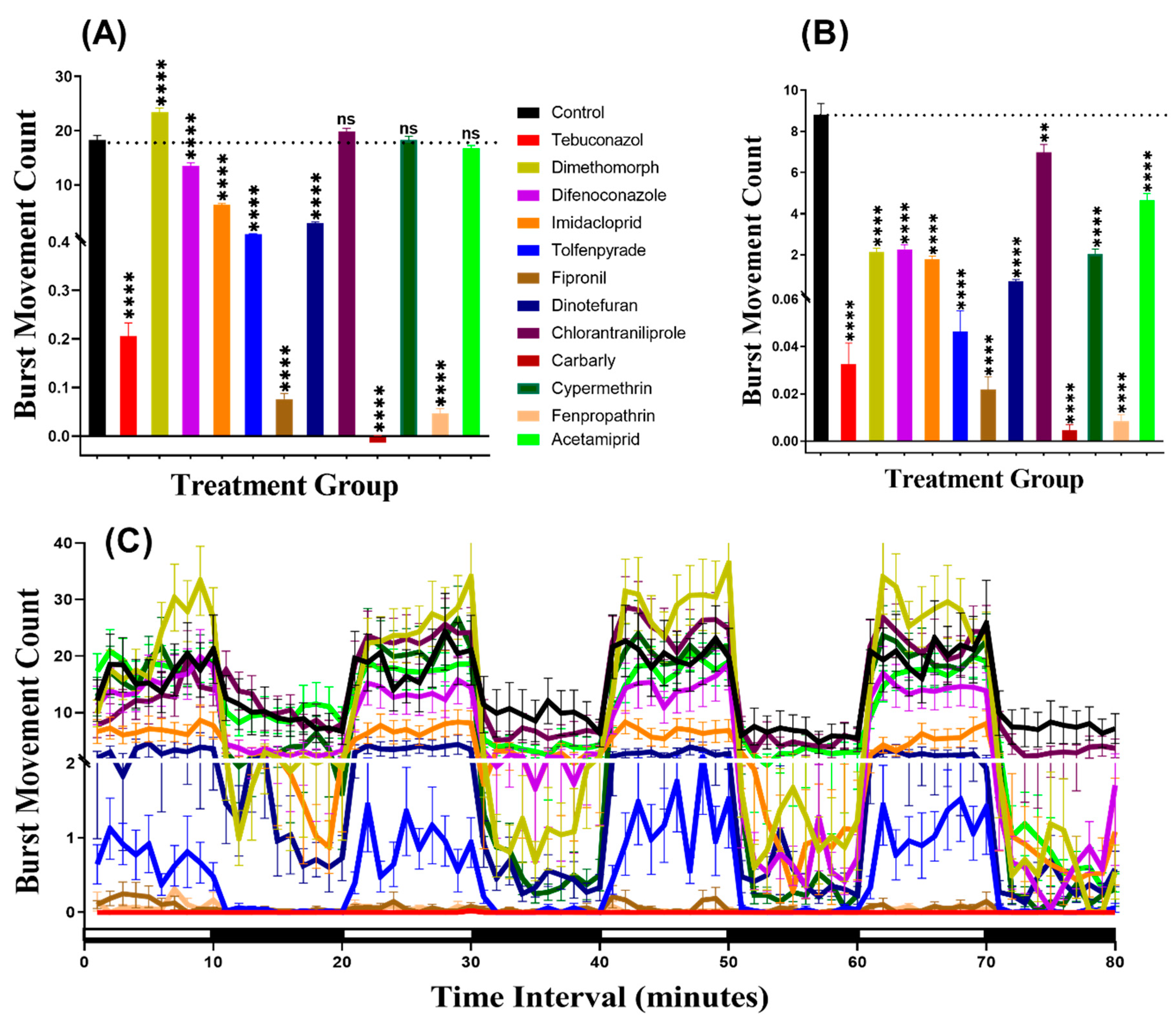
3.2. Burst Count Movement for Zebrafish after Pesticide Exposure
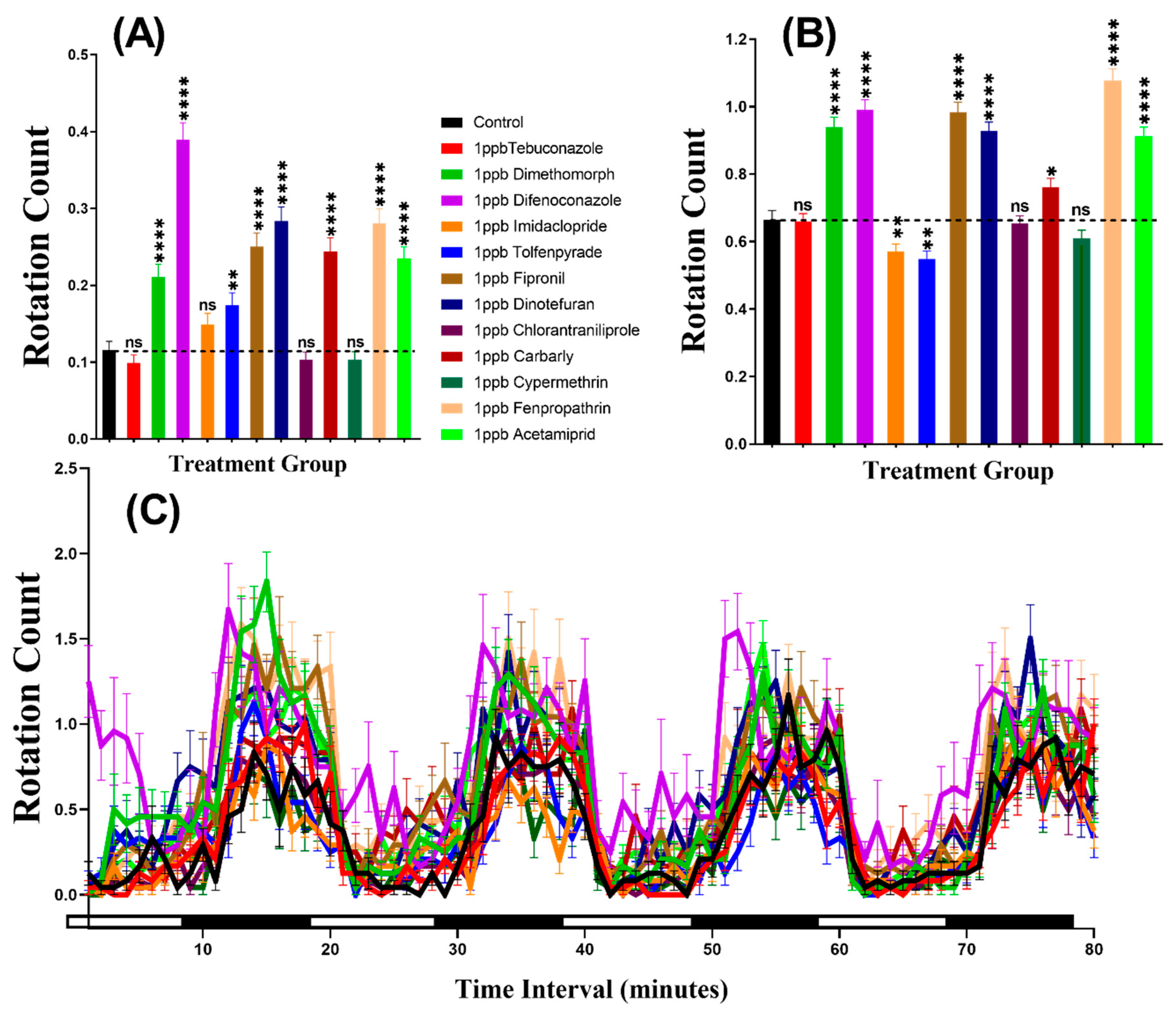
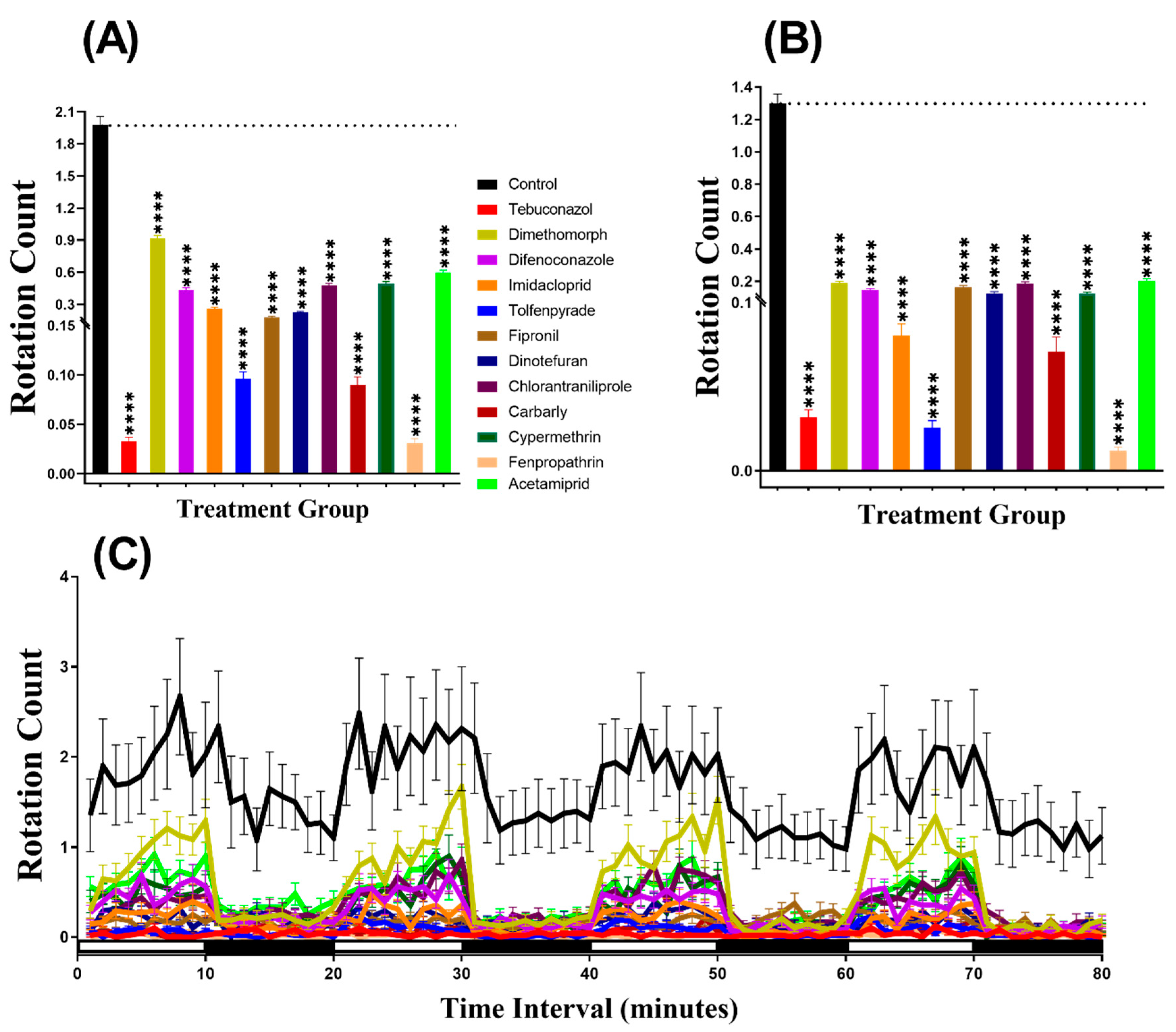
3.3. Swimming Orientation for Zebrafish after Pesticide Exposure
3.4. Total Distance for Daphnia Magna after Pesticide Exposure
3.5. Burst Count Movement for Daphnia magna after Pesticide Exposure
3.6. Swimming Orientation for Daphnia magna after Pesticide Exposure
3.7. PCA Analysis and Clustering of Phenomics Data
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Chemicals Tested | Concentration | Endpoints Tested | Main Highlights | References |
|---|---|---|---|---|
| Ionic and Nanosilver | 10, 20, 30, and 50 nM. | Mortality and growth | Toxicity increased with decreasing particle size. LC50 for nanosilver was 10 to 50 times greater than Ag+ (based on mass concentration). | [51] |
| R-Metalaxyl and rac-Metalaxyl in Acute, Chronic, and Sublethal condition | R-metalaxyl: 0.5, 16, and 169 mg/L for algae, Daphnia magna and zebrafish respectively. rac-metalaxyl: 0.05, 300, and 170 mg/L for algae, Daphnia magna and zebrafish respectively. | LC50 for 24, 48, and 96 h. Na,K-ATPase enzyme activity in adult zebrafish | R-metalaxyl were more toxic to aquatic organisms than rac-metalaxyl. After 24 h Na, K-ATPase enzyme activity increased by ~50% than control for 70 mg/L of R-metalaxyl but no significant values were collected at 10 mg/L. From 48 to 96, 1omg/L R-metalaxyl gradually increased. In contrast, rac-metalaxyl did not change the enzyme activity from 24 to 96 h for both concentration | [52] |
| Cimetidine | Daphnia magna: 0, 0.048, 0.24, 1.2, 6.0, and 30 mg/L Moina macrocopa: 0, 0.3, 3, 30, or 100 mg/L | Mortality and population growth Sex hormones and gene expression level in zebrafish | Cimetidine is not acutely toxic at levels occurring in the aquatic environment. Chronic exposure to cimetidine leads to alteration of the steroidogenesis pathway. Endocrine disruption effects were also observed in early life stage exposure | [53] |
| Cefadroxil and Cefadrine | Daphnia magna: cefadroxil (0, 5.6, 11.7, 22.2, 39.5, and 83.0 mg/L) and cefradine (0, 4.3, 9.7, 19.7, 39.0, and 80.8 mg/L). Oryzias latipes: cefadroxil (0, 1.0, 7.8, 84.8, 718.9, or 8883.1 mg/L) and cefradine (0, 1.0, 7.1, 73.9, 724.6, or 7758.5 mg/L). | Survival and population growth. Endocrine disruption mechanism | Exposure to these drugs caused disruptions to the functioning of the endocrine system altering gene transcription levels and sex hormone levels. Cefadroxil and cefradine impaired growth. | [3] |
| Four CeO2 Nanocrystalline catalyst (CuO–CeO2, CuCe20, CuCe10 and CuCe15) | Danio rerio and Daphnia magna: 1, 10, 50, 100, 250 and 500 mg/L | Hatching success and teratology effects | pure nanocrystalline CeO2 and mixed oxide CuO–CeO2 catalysts are not highly toxic as the other pollutants but still some sublethal effects of CuO–CeO2 were found | [54] |
| ZnO, CuO, Au, and TiO2 Nanoparticles | Daphnia magna: 0.5, 1, 2, 4, and 8 mg/ dm3 for ZnO NPs (30 nm) and CuO NPs; 1, 2, 4, 8, and 10 mg/ dm3 for TiO2 NPs (50 nm); and 0.05, 0.1, 0.2, 0.4, and 0.8 mg/ dm3 for Au NP (20 nm). Danio rerio: 5, 10, 15, 25, and 25 mg/dm3 for ZnO NPs (30 nm, 50 nm) and CuO NP; 100, 500, 800, and 1000 mg/ dm3 for TiO2 NPs (< 20 nm, 30 nm), and 0.25, 0.5, 1, 2, and 4 mg/dm3 for Au NP (20 nm). | Developmental effects and survival rate Biomarker analysis | Almost all four nanoparticles were toxic to both species. The number of offspring surprisingly decreased as the concentration of metal ion increased. The inhibited the CAT and SOD activities caused oxidative stress resulting physiological alteration in the early life stage | [55] |
| Perfluorooctane sulfonic acid (PFOS) and Perfluorooctanoic acid (PFOA) | Acute: PFOS (0, 6.25, 12.5, 25, 50, and 100 mg/L) and PFOA (0, 62.5, 125, 250, 500, and 1000 mg/L). Chronic: PFOA (0, 3.125, 6.25, 12.5, 25, and 50 mg/L) PFOS (0, 0.3125, 0.625, 1.25, 2.5, and 5 mg/L). Adult Medaka: PFOS, 0.01, 0.1, and 1 mg/L; PFOA, 0.1, 1, and 10 mg/L. | Population growth, teratological effects, Histology. | Parental exposure in Japanese medaka transferred adverse effects to offspring. The threshold of PFOS is approximately 10 times higher than that of PFOA in water Daphnia magna but Moina macrocopa was more to both compound than Daphnia magna. | [56] |
| Behavioral Endpoints | Group Comparisons | Danio rerio | Daphnia magna | ||
|---|---|---|---|---|---|
| Significance | p Value | Significance | p Value | ||
| Distance Traveled | Control vs. 1 ppb Tebuconazole | **** | <0.0001 | **** | <0.0001 |
| Control vs. 1 ppb Dimethomorph | ns | >0.9999 | **** | <0.0001 | |
| Control vs. 1 ppb Difenoconazole | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Imidacloprid | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Tolfenpyrade | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Fipronil | ** | 0.0031 | **** | <0.0001 | |
| Control vs. 1 ppb Dinotefuran | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Chlorantraniliprole | **** | <0.0001 | ns | 0.7406 | |
| Control vs. 1 ppb Carbarly | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Cypermethrin | * | 0.0135 | ns | 0.3779 | |
| Control vs. 1 ppb Fenpropathrin | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Acetamiprid | **** | <0.0001 | ns | 0.1502 | |
| Burst Movement Count | Control vs. 1 ppb Tebuconazole | ns | 0.5148 | **** | <0.0001 |
| Control vs. 1 ppb Dimethomorph | **** | <0.0001 | ns | 0.2088 | |
| Control vs. 1 ppb Difenoconazole | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Imidacloprid | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Tolfenpyrade | ** | 0.0014 | **** | <0.0001 | |
| Control vs. 1 ppb Fipronil | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Dinotefuran | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Chlorantraniliprole | **** | <0.0001 | ns | 0.6996 | |
| Control vs. 1 ppb Carbarly | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Cypermethrin | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Fenpropathrin | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Acetamiprid | **** | <0.0001 | **** | <0.0001 | |
| Rotation Count | Control vs. 1 ppb Tebuconazole | ns | 0.6257 | **** | <0.0001 |
| Control vs. 1 ppb Dimethomorph | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Difenoconazole | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Imidacloprid | ns | 0.1705 | **** | <0.0001 | |
| Control vs. 1 ppb Tolfenpyrade | ns | 0.2014 | **** | <0.0001 | |
| Control vs. 1 ppb Fipronil | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Dinotefuran | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Chlorantraniliprole | ns | 0.5932 | **** | <0.0001 | |
| Control vs. 1 ppb Carbarly | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Cypermethrin | ns | 0.1311 | **** | <0.0001 | |
| Control vs. 1 ppb Fenpropathrin | **** | <0.0001 | **** | <0.0001 | |
| Control vs. 1 ppb Acetamiprid | **** | <0.0001 | **** | <0.0001 | |

References
- Carvalho, F.P. Pesticides, environment, and food safety. Food Energy Secur. 2017, 6, 48–60. [Google Scholar] [CrossRef]
- Alexandratos, N.; Bruinsma, J.; Global Perspective Studies Team. World Agriculture towards 2030/2050: The 2012 Revision, (03); Food and Agriculture Organization of the United Nations Agricultural Development Economics Division: Rome, Italy, 2012; Volume 12, pp. 94–126. [Google Scholar]
- Song, H.; Kabir, E.; Jahan, S.A. Exposure to pesticides and the associated human health effects. Sci. Total. Environ. 2017, 575, 525–535. [Google Scholar] [CrossRef]
- Maroni, M.; Fait, A. Health effects in man from long-term exposure to pesticides. A review of the 1975-1991 literature. Toxicology 1993, 78, 1–180. [Google Scholar] [CrossRef]
- Townson, H. Public health impact of pesticides used in agriculture. Trans. R. Soc. Trop. Med. Hyg. 1992, 86, 350. [Google Scholar] [CrossRef]
- He, L.; Xiao, K.; Zhou, C.; Li, G.; Yang, H.; Li, Z.; Cheng, J. Insights into pesticide toxicity against aquatic organism: QSTR models on Daphnia Magna. Ecotoxicol. Environ. Saf. 2019, 173, 285–292. [Google Scholar] [CrossRef]
- Nowell, L.H.; Moran, P.W.; Schmidt, T.S.; Norman, J.E.; Nakagaki, N.; Shoda, M.E.; Mahler, B.J.; Van Metre, P.C.; Stone, W.W.; Sandstrom, M.W.; et al. Complex mixtures of dissolved pesticides show potential aquatic toxicity in a synoptic study of Midwestern U.S. streams. Sci. Total. Environ. 2018, 613, 1469–1488. [Google Scholar] [CrossRef] [PubMed]
- Jang, G.H.; Park, C.-B.; Kang, B.J.; Kim, Y.J.; Lee, K. Sequential assessment via daphnia and zebrafish for systematic toxicity screening of heterogeneous substances. Environ. Pollut. 2016, 216, 292–303. [Google Scholar] [CrossRef] [PubMed]
- Chakri, K.; Touati, L.; Alfarhan, A.H.; Al-Rasheid, K.A.; Samraoui, B. Effect of vertebrate and invertebrate kairomones on the life history of Daphnia magna Straus (Crustacea: Branchiopoda). Comptes Rendus Boil. 2010, 333, 836–840. [Google Scholar] [CrossRef] [PubMed]
- Enserink, L.; De La Haye, M.; Maas, H. Reproductive strategy of Daphnia magna: Implications for chronic toxicity tests. Aquat. Toxicol. 1993, 25, 111–123. [Google Scholar] [CrossRef]
- Gliwicz, Z.M.; Guisande, C. Family planning inDaphnia: Resistance to starvation in offspring born to mothers grown at different food levels. Oecologia 1992, 91, 463–467. [Google Scholar] [CrossRef]
- Jemec, A.; Drobne, D.; Tisler, T.; Trebše, P.; Ros, M.; Sepčić, K. The applicability of acetylcholinesterase and glutathione S-transferase in Daphnia magna toxicity test. Comp. Biochem. Physiol. Part C Toxicol. Pharmacol. 2007, 144, 303–309. [Google Scholar] [CrossRef] [PubMed]
- Kalueff, A.V.; Gebhardt, M.; Stewart, A.M.; Cachat, J.M.; Brimmer, M.; Chawla, J.S.; Craddock, C.; Kyzar, E.J.; Roth, A.; Landsman, S.; et al. Towards a Comprehensive Catalog of Zebrafish Behavior 1.0 and Beyond. Zebrafish 2013, 10, 70–86. [Google Scholar] [CrossRef] [PubMed]
- Kokel, D.; Dunn, T.W.; Ahrens, M.B.; Alshut, R.; Cheung, C.Y.J.; Saint-Amant, L.; Bruni, G.N.; Mateus, R.; Van Ham, T.J.; Shiraki, T.; et al. Identification of nonvisual photomotor response cells in the vertebrate hindbrain. J. Neurosci. 2013, 33, 3834–3843. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Thompson, W.A.; Arnold, V.I.; Vijayan, M.M. Venlafaxine in Embryos Stimulates Neurogenesis and Disrupts Larval Behavior in Zebrafish. Environ. Sci. Technol. 2017, 51, 12889–12897. [Google Scholar] [CrossRef] [PubMed]
- Pretti, C.; Chiappe, C.; Pieraccini, D.; Gregori, M.; Abramo, F.; Monni, G.; Intorre, L. Acute toxicity of ionic liquids to the zebrafish (Danio rerio). Green Chem. 2006, 8, 238–240. [Google Scholar] [CrossRef]
- Asharani, P.V.; Wu, Y.L.; Gong, Z.; Valiyaveettil, S. Toxicity of silver nanoparticles in zebrafish models. Nanotechnology 2008, 19, 255102. [Google Scholar] [CrossRef]
- Hill, A.J.; Teraoka, H.; Heideman, W.; Peterson, R.E. Zebrafish as a Model Vertebrate for Investigating Chemical Toxicity. Toxicol. Sci. 2005, 86, 6–19. [Google Scholar] [CrossRef] [Green Version]
- Rubinstein, A.L. Zebrafish assays for drug toxicity screening. Expert Opin. Drug Metab. Toxicol. 2006, 2, 231–240. [Google Scholar] [CrossRef]
- Restani, G.C.; Fonseca, A.L. Effects of Cylindrospermopsis raciborskii strains (Woloszynska, 1912) Senayya & Subba Raju on the mobility of Daphnia laevis (Cladocera, Daphniidae). Braz. J. Boil. 2014, 74, 23–31. [Google Scholar] [CrossRef] [Green Version]
- Hussain, A.; Audira, G.; Siregar, P.; Lin, Y.-C.; Villalobos, O.; Villaflores, O.B.; Wang, W.-D.; Hsiao, C.-D. Waterborne Exposure of Paclobutrazol at Environmental Relevant Concentration Induce Locomotion Hyperactivity in Larvae and Anxiolytic Exploratory Behavior in Adult Zebrafish. Int. J. Environ. Res. Public Health 2020, 17, 4632. [Google Scholar] [CrossRef]
- Avdesh, A.; Chen, M.; Martin-Iverson, M.; Mondal, A.; Ong, D.; Rainey-Smith, S.; Taddei, K.; Lardelli, M.; Groth, D.M.; Verdile, G.; et al. Regular Care and Maintenance of a Zebrafish (Danio rerio) Laboratory: An Introduction. J. Vis. Exp. 2012, e4196. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kristofco, L.A.; Cruz, L.C.; Haddad, S.P.; Behra, M.L.; Chambliss, C.K.; Brooks, B. Age matters: Developmental stage of Danio rerio larvae influences photomotor response thresholds to diazinion or diphenhydramine. Aquat. Toxicol. 2016, 170, 344–354. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Emran, F.; Rihel, J.; Dowling, J.E. A Behavioral Assay to Measure Responsiveness of Zebrafish to Changes in Light Intensities. J. Vis. Exp. 2008, 20, e923. [Google Scholar] [CrossRef] [PubMed]
- Herrero-Hernández, E.; Andrades, M.; Marín-Benito, J.M.; Sánchez-Martín, M.J.; Rodríguez-Cruz, M.S. Field-scale dissipation of tebuconazole in a vineyard soil amended with spent mushroom substrate and its potential environmental impact. Ecotoxicol. Environ. Saf. 2011, 74, 1480–1488. [Google Scholar] [CrossRef] [PubMed]
- Van Dijk, T.C.; Van Staalduinen, M.A.; Van Der Sluijs, J.P. Macro-Invertebrate Decline in Surface Water Polluted with Imidacloprid. PLoS ONE 2013, 8, e62374. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gunasekara, A.S.; Truong, T.; Goh, K.S.; Spurlock, F.; Tjeerdema, R.S. Environmental fate and toxicology of fipronil. J. Pestic. Sci. 2007, 32, 189–199. [Google Scholar] [CrossRef] [Green Version]
- Walters, J.; Goh, K.S.; Li, L.; Feng, H.; Hernandez, J.; White, J. Environmental monitoring of carbaryl applied in urban areas to control the glassy-winged sharpshooter in California. Environ. Monit. Assess. 2003, 82, 265–280. [Google Scholar] [CrossRef]
- Marino, D.; Ronco, A. Cypermethrin and Chlorpyrifos Concentration Levels in Surface Water Bodies of the Pampa Ondulada, Argentina. Bull. Environ. Contam. Toxicol. 2005, 75, 820–826. [Google Scholar] [CrossRef]
- Kanawi, E.; Budd, R.; Tjeerdema, R.S. Environmental Fate and Ecotoxicology of Fenpropathrin. Rev. Environ. Contam. Toxicol. 2013, 225, 77–93. [Google Scholar] [CrossRef]
- Bownik, A.; Sokołowska, N.; Ślaska, B. Effects of apomorphine, a dopamine agonist, on Daphnia magna: Imaging of swimming track density as a novel tool in the assessment of swimming activity. Sci. Total. Environ. 2018, 635, 249–258. [Google Scholar] [CrossRef]
- Latimer, K.S. Duncan and Prasse’s Veterinary Laboratory Medicine: Clinical Pathology, 5th ed.; Wiley: Danvers, MA, USA, 2011; pp. 365–367. [Google Scholar]
- Irons, T.; MacPhail, R.; Hunter, D.; Padilla, S. Acute neuroactive drug exposures alter locomotor activity in larval zebrafish. Neurotoxicol. Teratol. 2010, 32, 84–90. [Google Scholar] [CrossRef] [PubMed]
- MacPhail, R.; Brooks, J.; Hunter, D.; Padnos, B.; Irons, T.; Padilla, S. Locomotion in larval zebrafish: Influence of time of day, lighting and ethanol. NeuroToxicology 2009, 30, 52–58. [Google Scholar] [CrossRef] [PubMed]
- Baillieul, M.; Blust, R. Analysis of the swimming velocity of cadmium-stressed Daphnia magna. Aquat. Toxicol. 1999, 44, 245–254. [Google Scholar] [CrossRef]
- Gerhardt, A.; De Bisthoven, L.J.; Schmidt, S. Automated Recording of Vertical Negative Phototactic Behaviour in Daphnia magna Straus (Crustacea). Hydrobiologia 2006, 559, 433–441. [Google Scholar] [CrossRef]
- Dhawan, D.B.D.R. Comparison of Lethality, Reproduction, and Behavior as Toxicological Endpoints in the Nematode Caenorhabditis Elegans. J. Toxicol. Environ. Health Part A 1999, 58, 451–462. [Google Scholar] [CrossRef]
- Melvin, S.D.; Wilson, S.P. The utility of behavioral studies for aquatic toxicology testing: A meta-analysis. Chemosphere 2013, 93, 2217–2223. [Google Scholar] [CrossRef]
- Spyker, J.M. Behavioral Teratology and Toxicology. In Behavioral Toxicology; Springer Science and Business Media LLC: New York, NY, USA, 1975; pp. 311–349. [Google Scholar]
- Mitchell, C.L.; Tilson, H.A.; Evans, H.L. Behavioral Toxicology in Risk Assessment: Problems and Research Needs. CRC Crit. Rev. Toxicol. 1982, 10, 265–274. [Google Scholar] [CrossRef]
- Ren, Z.; Zha, J.; Ma, M.; Wang, Z.-J.; Gerhardt, A. The early warning of aquatic organophosphorus pesticide contamination by on-line monitoring behavioral changes of Daphnia magna. Environ. Monit. Assess. 2007, 134, 373–383. [Google Scholar] [CrossRef]
- Freire, P.P.; Fernandez, G.J.; De Moraes, D.; Cury, S.S.; Dal-Pai-Silva, M.; Reis, P.P.; Rogatto, S.R.; Carvalho, R. The expression landscape of cachexia-inducing factors in human cancers. J. Cachex Sarcopenia Muscle 2020. [Google Scholar] [CrossRef] [Green Version]
- Caron, H.; Van Schaik, B.D.; Van Der Mee, M.; Baas, F.; Riggins, G.; Van Sluis, P.; Hermus, M.-C.; Van Asperen, R.; Boon, K.; Voûte, P.A.; et al. The Human Transcriptome Map: Clustering of Highly Expressed Genes in Chromosomal Domains. Science 2001, 291, 1289–1292. [Google Scholar] [CrossRef] [Green Version]
- Kokel, D.; Bryan, J.; Laggner, C.; White, R.; Cheung, C.Y.J.; Mateus, R.; Healey, D.; Kim, S.; Werdich, A.A.; Haggarty, S.J.; et al. Rapid behavior-based identification of neuroactive small molecules in the zebrafish. Nat. Methods 2010, 6, 231–237. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tremolada, P.; Finizio, A.; Villa, S.; Gaggi, C.; Vighi, M. Quantitative inter-specific chemical activity relationships of pesticides in the aquatic environment. Aquat. Toxicol. 2004, 67, 87–103. [Google Scholar] [CrossRef] [PubMed]
- Colwill, R.M.; Creton, R. Imaging escape and avoidance behavior in zebrafish larvae. Rev. Neurosci. 2011, 22, 63–73. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kimmel, C.B.; Patterson, J.; Kimmel, R.O. The development and behavioral characteristics of the startle response in the zebra fish. Dev. Psychobiol. 1974, 7, 47–60. [Google Scholar] [CrossRef]
- Mokry, L.E.; Hoagland, K.D. Acute toxicities of five synthetic pyrethroid insecticides to daphnia magna and ceriodaphnia dubia. Environ. Toxicol. Chem. 1990, 9, 1045. [Google Scholar] [CrossRef]
- Bownik, A. Daphnia swimming behaviour as a biomarker in toxicity assessment: A review. Sci. Total. Environ. 2017, 601, 194–205. [Google Scholar] [CrossRef]
- Martins, J.C.; Oliva-Teles, L.; Vasconcelos, V. Assays with Daphnia magna and Danio rerio as alert systems in aquatic toxicology. Environ. Int. 2007, 33, 414–425. [Google Scholar] [CrossRef]
- Hoheisel, S.M.; Diamond, S.; Mount, D. Comparison of nanosilver and ionic silver toxicity in Daphnia magna and Pimephales promelas. Environ. Toxicol. Chem. 2012, 31, 2557–2563. [Google Scholar] [CrossRef]
- Yao, K.; Zhu, L.; Duan, Z.; Chen, Z.; Li, Y.; Zhu, X. Comparison of R-metalaxyl andrac-metalaxyl in acute, chronic, and sublethal effect on aquatic organisms: Daphnia magna, Scenedesmus quadricanda, and Danio rerio. Environ. Toxicol. 2009, 24, 148–156. [Google Scholar] [CrossRef]
- Rihel, J.; Prober, D.A.; Arvanites, A.; Lam, K.; Zimmerman, S.; Jang, S.; Haggarty, S.J.; Kokel, D.; Rubin, L.L.; Peterson, R.T.; et al. Zebrafish Behavioral Profiling Links Drugs to Biological Targets and Rest/Wake Regulation. Science 2010, 327, 348–351. [Google Scholar] [CrossRef] [Green Version]
- Jemec, A.; Djinović, P.; Tisler, T.; Pintar, A. Effects of four CeO2 nanocrystalline catalysts on early-life stages of zebrafish Danio rerio and crustacean Daphnia magna. J. Hazard. Mater. 2012, 219, 213–220. [Google Scholar] [CrossRef] [PubMed]
- Liu, J.; Fan, D.; Wang, L.; Shi, L.; Ding, J.; Chen, Y.; Shen, S. Effects of ZnO, CuO, Au, and TiO2 nanoparticles on Daphnia magna and early life stages of zebrafish Danio rerio. Environ. Prot. Eng. 2014, 40, 40. [Google Scholar] [CrossRef]
- Ji, K.; Kim, Y.; Oh, S.; Ahn, B.; Jo, H.; Choi, K. Toxicity of perfluorooctane sulfonic acid and perfluorooctanoic acid on freshwater macroinvertebrates (daphnia magna and moina macrocopa) and fish (oryzias latipes). Environ. Toxicol. Chem. 2008, 27, 2159. [Google Scholar] [CrossRef] [PubMed]








| Number | Pesticide | Functional Grouping to | WHO GHS Classification for Aquatic Acute Toxicity * | Aquatic Environmental Concentration µg/L | Reference |
|---|---|---|---|---|---|
| 1 | Tebuconazole | Triazole fungicide | 1 | N.A. | N.A. |
| 2 | Difenoconazole | Triazole fungicide | 1 | 9.1 | [25] |
| 3 | Dimethomorph | Morpholine fungicide | N.A. | 24.40 | EPA, 1998 |
| 4 | Imidacloprid | Neonicotinoid insecticide | 1 | 320 | [26] |
| 5 | Tolfenpyrad | Pyrazole insecticide | 1 | 26.9 | EPA, 2014 |
| 6 | Fipronil | Phenylpyrazole insecticide | 1 | 0.117 | [27] |
| 7 | Dinotefuran | Neonicotinoid insecticide | N.A. | 9.64 | US EPA, 2004 |
| 8 | Chlorantraniliprole | Ryanoid insecticide | 1 | N.A. | N.A. |
| 9 | Carbaryl | Carbamate insecticide | 1 | 0.125 | [28] |
| 10 | Cypermethrin | Synthetic pyrethroid insecticide | 1 | 3.5 | [29] |
| 11 | Fenpropathrin | Pyrethroid insecticide | 1 | undetectable | [30] |
| 12 | Acetamiprid | Neonicotinoid insecticide | N.A. | 0.0544 | [30] |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hussain, A.; Audira, G.; Malhotra, N.; Uapipatanakul, B.; Chen, J.-R.; Lai, Y.-H.; Huang, J.-C.; Chen, K.H.-C.; Lai, H.-T.; Hsiao, C.-D. Multiple Screening of Pesticides Toxicity in Zebrafish and Daphnia Based on Locomotor Activity Alterations. Biomolecules 2020, 10, 1224. https://doi.org/10.3390/biom10091224
Hussain A, Audira G, Malhotra N, Uapipatanakul B, Chen J-R, Lai Y-H, Huang J-C, Chen KH-C, Lai H-T, Hsiao C-D. Multiple Screening of Pesticides Toxicity in Zebrafish and Daphnia Based on Locomotor Activity Alterations. Biomolecules. 2020; 10(9):1224. https://doi.org/10.3390/biom10091224
Chicago/Turabian StyleHussain, Akhlaq, Gilbert Audira, Nemi Malhotra, Boontida Uapipatanakul, Jung-Ren Chen, Yu-Heng Lai, Jong-Chin Huang, Kelvin H.-C. Chen, Hong-Thih Lai, and Chung-Der Hsiao. 2020. "Multiple Screening of Pesticides Toxicity in Zebrafish and Daphnia Based on Locomotor Activity Alterations" Biomolecules 10, no. 9: 1224. https://doi.org/10.3390/biom10091224