Brain Cytoplasmic RNAs in Neurons: From Biosynthesis to Function

1
Department of Chemistry, Korea Advanced Institute of Science and Technology (KAIST), Daejeon 34141, Korea
2
Center for Multiscale Chiral Architectures (CMCA), Daejeon 34141, Korea
3
Innovative Target Research Center, Korea Research Institute of Chemical Technology, Daejeon 34114, Korea
*
Author to whom correspondence should be addressed.
Biomolecules 2020, 10(2), 313; https://doi.org/10.3390/biom10020313
Submission received: 16 December 2019
/
Revised: 13 February 2020
/
Accepted: 13 February 2020
/
Published: 17 February 2020
(This article belongs to the Special Issue RNA Trafficking and Local Translation in Neuronal Health and Disease)
Abstract
:Flexibility in signal transmission is essential for high-level brain function. This flexibility is achieved through strict spatial and temporal control of gene expression in neurons. Given the key regulatory roles of a variety of noncoding RNAs (ncRNAs) in neurons, studying neuron-specific ncRNAs provides an important basis for understanding molecular principles of brain function. This approach will have wide use in understanding the pathogenesis of brain diseases and in the development of therapeutic agents in the future. Brain cytoplasmic RNAs (BC RNAs) are a leading paradigm for research on neuronal ncRNAs. Since the first confirmation of brain-specific expression of BC RNAs in 1982, their investigation has been an area of active research. In this review, we summarize key studies on the characteristics and functions of BC RNAs in neurons.
1. Introduction
Prior to completion of sequencing of the human genome, the complexity of living organisms had been assumed to derive from the diversity of protein-coding genes, which were estimated to number ≈120,000 in the human genome [1,2]. The Human Genome Project, however, revealed the shocking finding that only ≈2% of the human genome encodes proteins [3]. The amount of noncoding DNA was much greater than had previously been predicted, and could no longer be treated as “junk DNA”. This realization prompted follow-up studies that sought to determine the biological significance of noncoding DNA. Subsequent extensive transcriptome analyses have established that the human genome is pervasively transcribed, with more than 60% of the genome producing RNAs [4,5,6,7].
Noncoding RNAs (ncRNAs), commonly defined as transcripts that lack the ability to produce a protein, are classified according to size as long noncoding RNAs (lncRNAs; >200 nucleotides (nt)) and small noncoding RNAs (sncRNAs; <200 nt). About 60,000 lncRNAs have been identified in human cells; as an indication that ncRNA research is still in its infancy, ≈70% of these lncRNAs remain unnamed [2,8]. Although these RNAs were initially thought to be transcription byproducts without specific functions [9,10], recent studies have shown that RNAs themselves or their transcription actively contribute to biological phenomena through a variety of mechanisms [8,11,12,13,14,15].
The brain, the most complex organ in the body, interprets a variety of stimuli from the outside world, generating the appropriate signal and passing it on to other organs. This process, carried out by 80 billion neurons spread throughout the central nervous system [16], owes much to the operation of networks of ncRNAs. Brain activity at the cellular level can be interpreted as a specific combination of neuronal activation and inactivation [17,18]. Again, at the molecular level, this can be expressed as a change in the balance between activation and inactivation of the expression of specific genes in neurons. For normal brain activity, this process must be signal-dependent and requires the highest-level expression control system [19,20,21,22,23]. Neurons utilize a variety of ncRNAs to achieve this [20,23,24,25,26]. In 2008, Mercer confirmed the expression of 849 ncRNAs in the adult mouse brain, many of which exhibited characteristic location patterns [27]. The hypothesis that all these ncRNAs have a definite function may be an exaggerated idea [28], but the research that reveals the meaning of their existence is essential for a deeper understanding of the brain, a complex organ.
In 1982, Sutcliffe and colleagues first discovered a polymerase III (pol III)-dependent ncRNA that was expressed only in the rat brain [29]. While analyzing poly A+ transcripts in the brain, liver, and kidney tissues, they found a small (152 nt) transcript that was specifically expressed in the brain and named it brain cytoplasmic 1 RNA (BC1 RNA) [29,30]. A subsequent study in 1987 identified a 200 nt-long RNA expressed in the primate brain that had common expression characteristics with BC1 RNA, naming it brain cytoplasmic 200 RNA (BC200 RNA) [31].
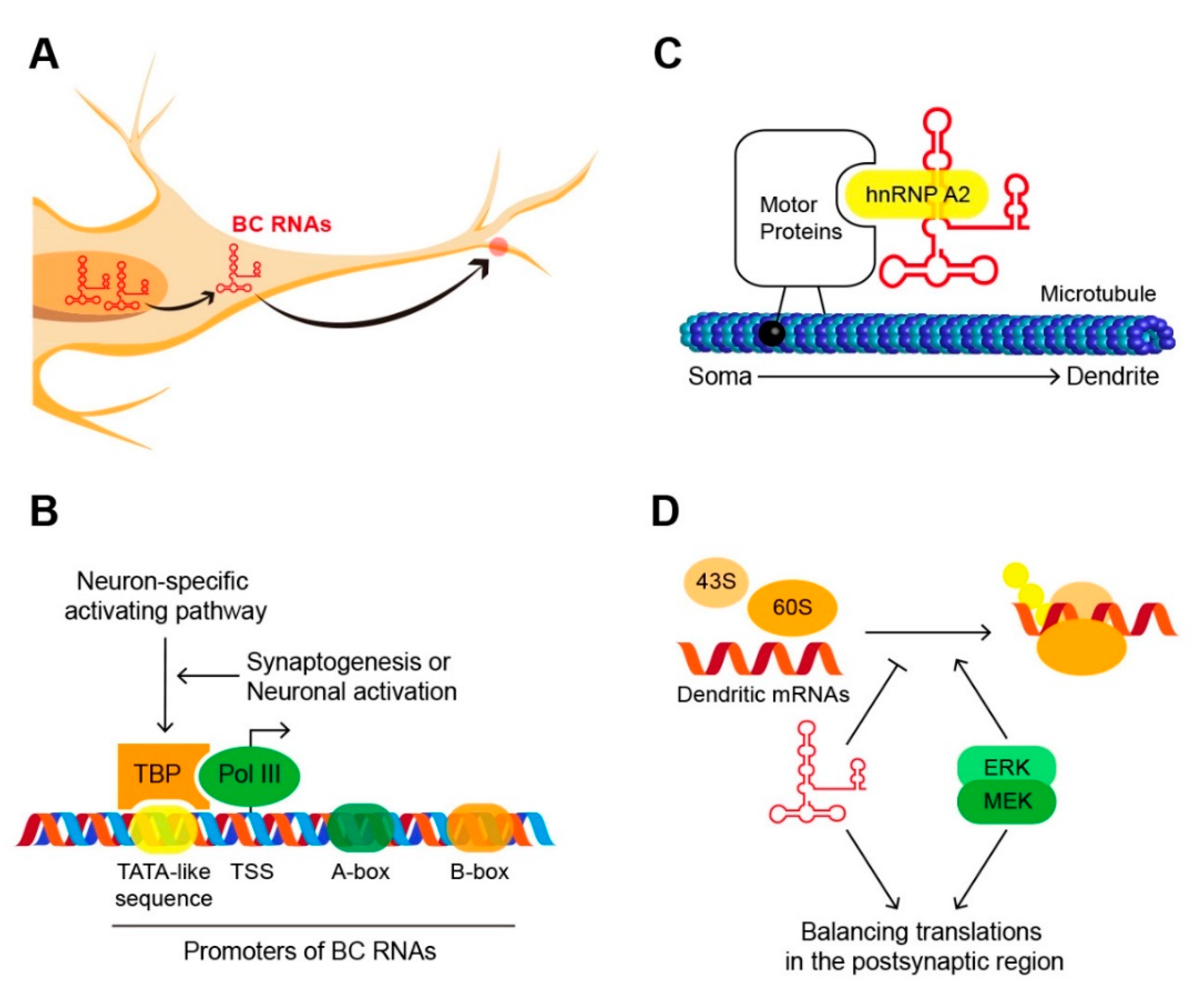
BC1 RNA and BC200 RNA, collectively called brain cytoplasmic RNAs (BC RNAs), are specifically located in neuronal dendrites and control protein translation by inhibiting translation–initiation factors (Figure 1) [32,33]. This regulation plays an important role in maintaining the appropriate protein translation balance in post-synaptic regions. The association of some neurological diseases with BC RNAs illustrates how important regulation by BC RNAs is for normal brain activity [34]. This review summarizes key studies of BC RNAs in neurons and may provide important insights into future research on ncRNAs in the brain—the subject of considerable recent attention.
2. BC RNAs: BC1 and BC200 RNA
In 1984, Sutcliffe and colleagues extended their initial work, examining BC1 RNA expression in additional tissues of the rat, including the adrenal gland, spleen, testis, lung, heart, muscle, and gut. At this point, BC1 RNA had still been identified only in the brain [30], and was thought to be a transcriptional byproduct of mRNA splicing because it was too short to function as mRNA. However, it was identified as an ncRNA because the corresponding DNA in the genome contained its own promoter and the ncRNA itself was located in the cytoplasm. In 1987, Dechiara and Brosius first sequenced BC1 RNA by generating complete cDNA [35]. They showed that BC1 RNA is transcribed from a single gene derived from a retroposed transfer RNAAla (tRNAAla) harboring 3′ unique sequence. Their subsequent study confirmed that BC1 RNA is conserved among rodents, including the guinea pig, Syrian golden hamster, and mouse [36].
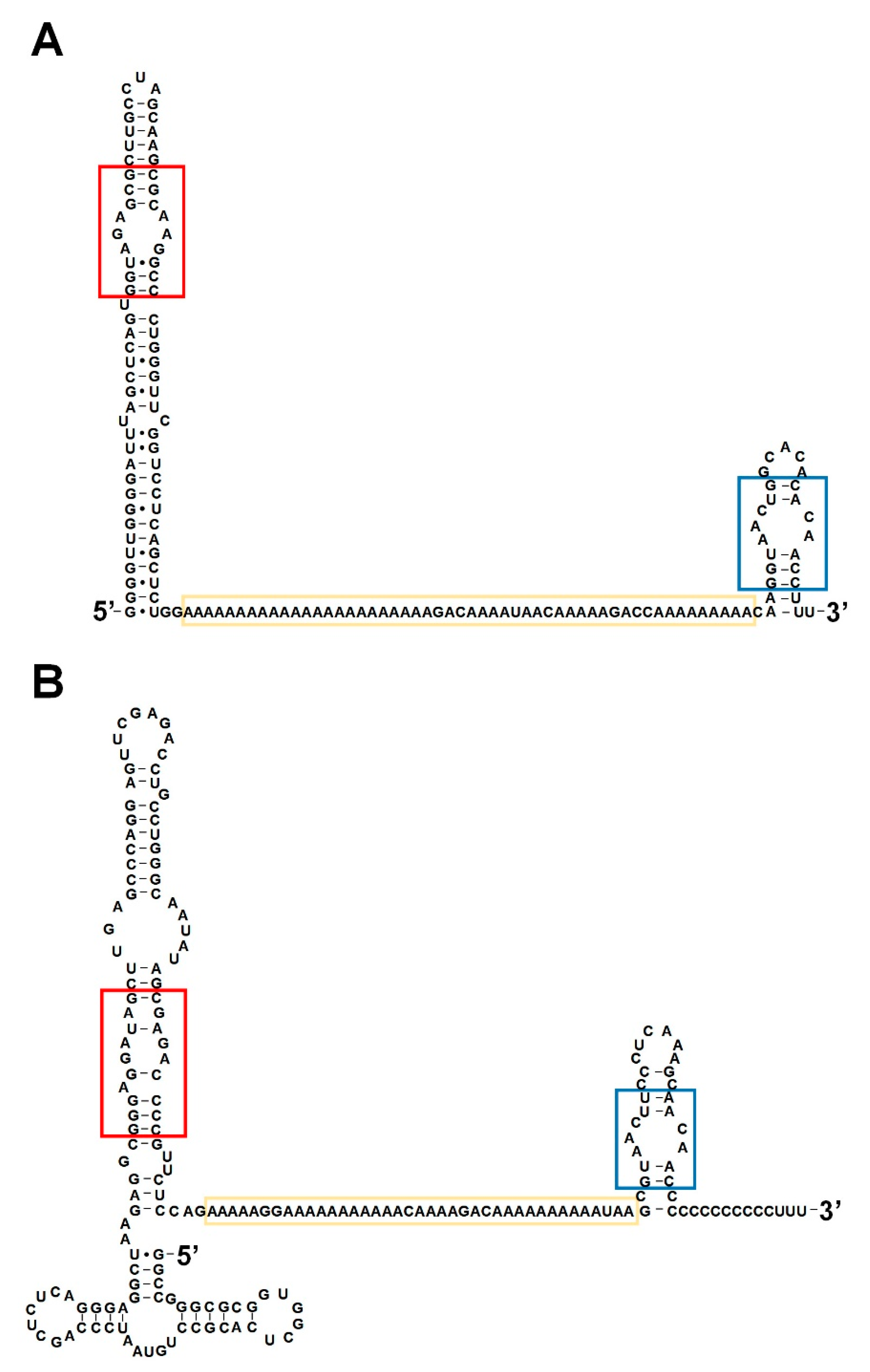
In 1987, Watson and Sutcliffe identified a 200 nt-long RNA that exhibited brain-specific expression in primates and named it BC200 RNA. This gene is derived from a monomeric Alu element and the transcript has a poly A region similar to BC1 RNA, but the overall sequences are very different [31,36]. However, research over the past three decades has confirmed that BC1 and BC200 RNA are functional analogues that perform the same function in each neuron through key common motifs including the GA kink motif, the poly A region, and the C-loop motif (Figure 2) [32,37,38,39].
3. Biosynthesis and RNA Stability
In 2007, Khanam et al. showed in transgenic mice that the BC200 RNA gene harboring only 250 bp upstream sequences expressed a stable amount of RNA comparable to a construct harboring about 2.3 kb upstream [40]. In 2017, Kim et al. conducted a serial upstream deletion analysis to identify the promoter. They found that TATA-binding protein (TBP) binds to the position 28 to 22 upstream sequences, recruiting RNA polymerase III complex to initiate transcription. Additionally, they further confirmed the presence of an internal promoter in the 5′ terminal region of the transcript [41]. The BC1 RNA gene similarly has two types of promoters in the upstream and internal regions [42].
Normally, BC RNAs are expressed only in neurons, but this regulated expression disappears completely during tumorigenesis. In 1997, Chen et al. confirmed that BC200 RNA was abnormally expressed in esophageal, lung, breast, colon, and cervical cancer tissues [43]. Since then, studies have been conducted to analyze the causes of such abnormal expression. A study by Singh et al. showed that BC200 RNA expression is promoted in an estrogen receptor-dependent manner in breast cancer cell lines [44]. Using chromatin immunoprecipitation (ChIP) assays, these authors confirmed that estrogen receptor-α binds directly to the upstream region of BC200 RNA to promote transcription. Another study by Hu and Lu confirmed that BC200 RNA is induced by an oncogenic transcriptional factor, c-MYC in non-small cell lung cancer cells [45]. However, it is not yet known whether these factors and mechanisms are involved in the expression of BC RNAs in neurons.
Recently, Sonawane and his colleagues published important results on tissue-specific gene expression [46]. They collected expression information for 30,243 proteins from 38 tissues and analyzed the regulatory network pattern in each tissue by incorporating the initial regulatory network information for 644 transcription factors and transcription factor protein–protein interactions [47,48]. They found that tissue-specific expression is determined by tissue-specific regulatory network pathways rather than by the expression of specific transcription factors. This finding shows that identifying the neuron-specific regulatory network that controls ncRNA expression is necessary to further understand the biosynthesis of BC RNAs.
Another characteristic of the biosynthesis of BC RNAs is that the level of expression is strongly affected by the development and activity of neurons. In 1998, Muslimov et al. observed that BC1 RNA remained low until synapses were fully formed and that its expression was suppressed by neuronal activity inhibitor, tetrodotoxin [49]. Activity-dependent expression provided an important clue for the functional significance of BC RNAs in neurons.
RNA stability is another factor that determines the number of intracellular transcripts [50]. Kim et al. compared the stability of BC200 RNA in various cancer cell lines and showed that the half-life of BC200 RNA changes significantly depending on the cell line [41]. This indicates that the environment in which BC RNA exists varies greatly from cell to cell.
4. Functions of BC RNAs
BC RNAs are strong translational regulators. In 2002, using an in vitro system, Wang and colleagues first discovered that BC1 RNA exerts a translation-inhibitory effect [51]. Subsequence studies confirmed that BC1 RNA binds to eukaryotic initiation factor 4A (eIF4A) and poly A binding protein (PABP), and inhibits cap-dependent and internal ribosome entry site (IRES)-dependent translation [52,53]. eIF4A is an ATP-dependent RNA helicase that initiates translation by unwinding the structure of the 5′ mRNA [54]. In 2008, Lin et al. found that BC1 RNA extinguishes the molecular force of eIF4A obtained through ATP hydrolysis. This was the first confirmation that BC1 RNA directly inhibits the activity of a single enzyme [55]. BC RNAs were also found to regulate translation in a manner that inhibits the function of eIF4B [56,57].
The generation of BC1-knockout (BC1-KO) mice by Skryabin et al. was an important starting point for the physiological study of BC RNAs [58]. In 2004, Lewejohann and colleagues found that BC1-KO mice have behavioral changes, such as reduced exploration, increased anxiety, and reduced survival in outdoor environments [59]. Independent studies by Zhong and Maccarrone confirmed abnormal increases in transmission that depended on metabotropic glutamate receptors (mGluRs) in BC1-KO mice [60,61]. In 2017, Briz et al. confirmed that a BC1 deficiency causes expression imbalances in the postsynaptic region, promoting an abnormal expression of glutamate receptor subunits that results in excessive signal transmission. The resulting hyperexcitation of neurons extinguishes synaptic plasticity, leading to severe defects in cognition and learning [62].
Taken together, these observations indicate that BC RNAs regulate signal transmission by maintaining a balance between translational activation and inhibition in postsynaptic regions. This regulation allows neurons to exhibit experience- and context-dependent plasticity and ultimately perform higher-level brain functions, such as cognition and learning.
5. Regulatory Factors
Although BC RNAs can affect global expression in vitro, they may have a limited effect in vivo [53,63]. This suggests the presence of higher regulators that control the functions of BC RNAs in cells. The specific localizing of BC RNA in neurons is one of the well-studied mechanisms of BC RNA activity control [64]. Neurons are structurally and functionally asymmetrical cells, and thus the location of the components is finely adjusted according to their function [65,66]. BC RNAs are preferentially localized in the dendrite through the GA kink-turn motif in the 5′ stem-loop [51,64,67,68,69]. This is a common structural characteristic of dendritic mRNAs and is recognized by heterogeneous ribonucleoprotein A2 (hnRNP A2), leading to microtubule-dependent dendritic localization of RNAs [68,69,70]. This mechanism limits the activity of BC RNAs to the microenvironment within the neuronal dendrites.
In 2019, a study by Muslimov et al. confirmed that the dendritic localization of BC200 RNA was severely impaired in the autoimmune disease systemic lupus erythematosus (SLE) [38]. They further detected specific antibodies against BC200 RNA in the serum of SLE patients and found that the antibody competitively inhibited binding to hnRNP A2. These findings show how important the dendritic localization of BC200 RNA is for normal brain activity.
In 2003, Zalfa and colleagues reported that BC1 RNA binds to fragile X mental retardation protein (FMRP), suggesting that the BC1 RNA-FMRP complex might regulate the translation of specific mRNAs [71]. However, in 2008, Iacoangeli et al. revealed that BC1-FMRP binding affinity is as low as the tRNA-FMRP level and there is no binding in vivo either [72]. These verification experiments led to the conclusion that the interaction between BC1 and FMRP is merely nonspecific binding. The recent results of Booy et al. also support this conclusion [73]. Their analysis of proteins interacting with BC RNAs at the genome level showed binding to 14 proteins including 8 new proteins, but no direct binding to FMRP.
Booy et al.’s work offers potential for new regulatory systems of BC RNA function in neurons [73]. Among newly discovered binding proteins, poly C binding protein 2 (PCBP2) has been shown to control the translational inhibition of BC200 RNA in vitro [74]. Further research is needed to see whether other interactions have regulatory effects and whether these regulators also work in real neurons.
6. Neuronal Diseases Related to BC RNAs
BC RNAs are associated with several neuronal diseases (Table 1). In 2007, Mus and colleagues reported that BC200 RNA was associated with Alzheimer’s disease (AD) [34]. They investigated BC200 RNA expression levels in Brodmann’s area 9, known to be severely affected in AD, at different ages. They found that, in a normal brain, expression of BC200 RNA gradually decreases with age. However, they confirmed that BC200 RNA levels were increased by ≈2-fold in the brains of AD patients of a similar age. The degree of abnormal expression increased in proportion to the clinical dementia rating. Additionally, BC200 RNAs in patients’ brains were abnormally distributed, showing high-intensity clusters in the soma. However, how BC200 RNA expression is promoted in AD and how it affects the pathogenesis of AD are not known.
Fragile X syndrome (FXS) is a congenital brain disease characterized by a wide range of neuropsychiatric manifestations, including mental retardation, hyperactivity, and autism [75]. The main cause is decreased expression of FMRP protein owing to a mutation in the FMR1 gene on the X chromosome [76]. Independently of BC RNAs, FMRP contributes to building synaptic plasticity by regulating translation of dendritic mRNAs [77]. Although normal people have between 6–54 CGG repeats in the FMR1 gene, FXS patients have over 200 CGG repeats, which reduces the expression of FMRP protein and causes brain dysfunction [78]. BC1-KO mice show abnormal neuronal transmission patterns that are similar to those of FMRP-KO mice [61,79], suggesting that the loss of the BC RNAs function may have a similar pathological effect.
In addition, CGG repeats of the FMR1 gene have been reported to directly affect the function of BC RNAs. People with 55-200 CGG repeats inserted into the FMR1 gene are classified as premutation [78]. Young premutation carriers may present with cognitive disturbances and aged carriers may develop fragile X-associated tremor/ataxia syndrome (FXTAS), a neurodegenerative disorder; however, detailed pathogenesis is not yet well understood [80]. Muslimov et al. established and analyzed CGG knock-in mice [81]. They found that most of the BC1 RNA remained near the nucleus in neurons and confirmed that these neurons were hyper-excited by signaling and caused cognitive decline. CGG repeat motif strongly bound to hnRNP A2, inhibiting the localization of BC RNAs by weakening the interaction between hnRNP A2-BC RNAs. Through these observations, they suggested that the mislocalization of BC RNAs by CGG repeats could be the cause of fragile X premutation disorders.
Muslimov and colleagues recently reported that the mislocalization of BC RNAs is also associated with SLE [38]. Researchers confirmed the production of antibodies that recognize BC200 RNA in SLE patients and named them anti-BC abs. They found that anti-BC abs are not detectable in normal cells or in other autoimmune diseases. Anti-BC abs bind to the 5′ stem-loop of BC200 RNA and inhibit the binding of hnRNP A2, resulting in a defect in dendritic localization. Finally, they confirmed that administration of anti-BC abs to normal mice resulted in phenotypic defects, such as epileptic-induced responses and impaired cognitive function.
7. BC RNAs in Cancers
In 1997, abnormal expression of BC200 RNA was first detected in several cancer tissues, including breast, esophagus, lung, ovary, parathyroid, and tongue [43]. In 2004, more invasive cancer cells were found to express higher levels of BC200 RNA, showing the possibility of contributing to cancer development [82]. The detailed mechanism had not been well understood for a long time, but has recently begun to be proposed by some studies [44,63,83].
In 2016, Singh and colleagues constructed BC200 knock-out cell lines by the clustered regularly interspaced short palindromic repeats/CRISPR associated protein 9 (CRISPR/Cas9) system [44]. They observed that the deletion of the BC200 gene inhibits cell growth by activating the apoptosis of the cancer cells. They showed that BC200 RNA partially binds to B-cell lymphoma-extra (Bcl-x) mRNA, inhibiting splicing to Bcl-xS, the apoptosis-promoting factor. However, further studies are required that verifythe real interaction between BC200 RNA and Bcl-x mRNA in vivo and show how cancer BC RNAs, mainly located in the cytoplasm in neurons, work in the nucleus in cancer cells.
In 2017, Shin et al. investigated the effect of BC200 RNA on genome-wide expression profiling of the cervical cancer cell line HeLa [63]. As a result, they found that expressions of 29 genes are altered by BC200 RNA knockdown. Among them, the expression of S100A11, previously identified as the cell mobility activating factor, was significantly reduced. The researchers showed that BC200 RNA promotes cell mobility of HeLa cells by stabilizing S100A11 mRNA and promoting its expression. However, it is not yet known how BC200 RNA enhances the stability of S100A11 mRNA. Additionally, some studies suggest that BC200 RNA may promote the development of cancer cells by inhibiting tumor-suppressive miRNAs [84,85].
The study of BC RNAs in cancer has a relatively shorter history. Although more detailed research on mechanisms is required, it is interesting to have the potential shown for BC200 RNA to act as a more diverse mechanism within cells compared to previous studies in neurons. Future studies need to confirm whether these mechanisms are also preserved in neurons. Although the environment of BC RNAs varies dramatically from cell to cell, neurons and cancer cells may share some of the same binding proteins and functions of BC RNAs. The BC RNAs of neurons have the potential to regulate gene expression through more complex mechanisms than have been identified to date.
8. Conclusions
If a protein is the object that performs a physical function in a cell, ncRNAs are regulators that control this function so that it occurs at the right time and place [11]. The proportion of ncRNAs in the genome increases as biological complexity increases [86], suggesting that the higher-order functions of complex organisms may be more dependent on the number of ncRNAs than on the number of proteins. The human brain performs the highest-level functions of all organs in the body. The process of recognizing, interpreting, and responding to the diverse situations encountered requires the most sophisticated control systems in all living organisms. It is known that various classes of neuronal ncRNAs actively contribute to this regulation [20,64,87,88,89].
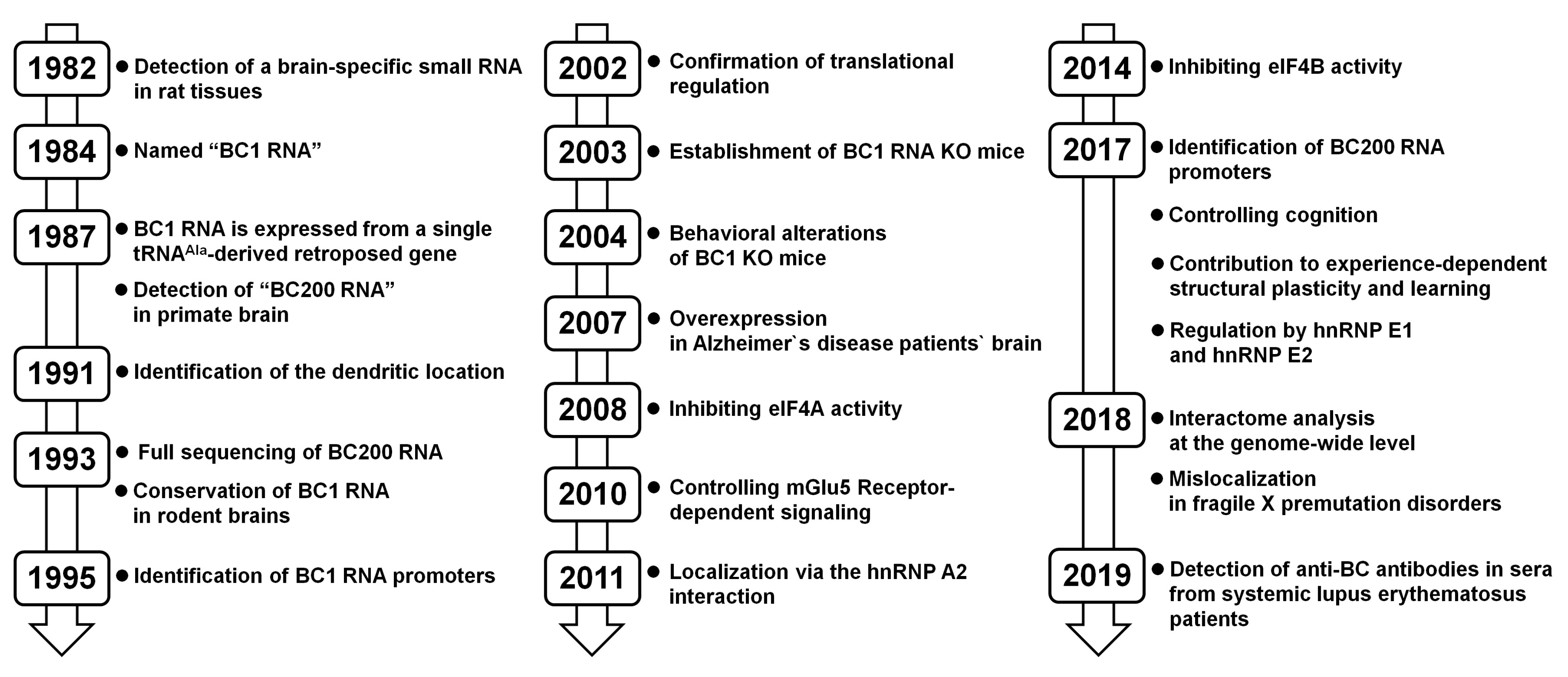
The long history of BC RNA research in the neuron is divided into three stages (Figure 3). In the first stage, early research examined the expression characteristics of BC RNAs. RNA expression analyses of each tissue revealed that BC1 RNA was specifically expressed in rat brain and BC200 RNA was expressed in the human brain. In the second stage, in vitro and cell-based experiments revealed the molecular function of BC RNAs in neurons. The binding partners of BC RNAs were identified and their interactions were shown to exert potent translation-inhibitory functions in post-synaptic regions. In the third stage, the physiological functions of BC RNAs in neurons were explored. Behavioral experiments using BC1-KO mice showed that BC RNAs are involved in higher-level brain functions, such as learning and cognition. Combined with molecular-level functional studies, this research showed that BC RNAs play an important role in the formation of neuronal plasticity involved in signal-dependent activation, while maintaining a translational balance in dendrite regions.
This flow of studies provides important inspiration for future research of BC RNAs and other ncRNAs in neurons. An enormous number of ncRNAs expressed in the central nervous system have still remained in the first stage [24,27,90]. Multifaceted functional analyses of neuron-specific transcripts are needed. To this end, an analysis of genome-wide effects is essential, and it will be necessary to establish animal models for the analysis of physiological functions. Recent advances in sequencing analysis and genome editing technology using CRISPR/Cas9 are accelerating these studies.
Funding
We are supported by the National Research Foundation of Korea (NRF) grants funded by the Korean government (MSIT) (no. NRF-2017R1A6A3A11031308; no. NRF-2018R1A5A1025208; no. NRF-2019R1H1A2039730).
Acknowledgments
The authors are grateful to Jürgen Brosius for his comments on the manuscript.
Conflicts of Interest
The authors declare no conflict of interest.
Abbreviations
ncRNA, noncoding RNA; BC RNA, brain cytoplasmic RNA; pol III, polymerase III; MEK/ERK, mitogen-activated protein kinase kinase/extracellular signal-regulated kinase; TBP, TATA-binding protein; ChIP, chromatin immunoprecipitation; eIF, eukaryotic initiation factor; PABP, poly A binding protein; IRES, internal ribosome entry site; KO, knock-out; mGluR, metabotropic glutamate receptor; hnRNP, heterogeneous ribonucleoprotein particle; FMRP, fragile X mental retardation protein; PCBP, poly C binding protein; AD, Alzheimer’s disease; FXS, fragile X syndrome; FXTAS, fragile X-associated tremor/ataxia syndrome; SLE, systemic lupus erythematosus; CRISPR/Cas9, clustered regularly interspaced short palindromic repeats/CRISPR associated protein 9; Bcl-x, B-cell lymphoma-extra
References
- Liang, F.; Holt, I.; Pertea, G.; Karamycheva, S.; Salzberg, S.L.; Quackenbush, J. Gene index analysis of the human genome estimates approximately 120,000 genes. Nat. Genet. 2000, 25, 239–240. [Google Scholar] [CrossRef] [PubMed]
- Salzberg, S.L. Open questions: How many genes do we have? BMC Biol. 2018, 16, 1–3. [Google Scholar] [CrossRef] [PubMed]
- Lander, S.; Linton, L.M.; Birren, B.; Nusbaum, C.; Zody, M.C.; Baldwin, J.; Devon, K.; Dewar, K.; Doyle, M.; FitzHugh, W.; et al. Initial sequencing and analysis of the human genome International Human Genome Sequencing Consortium. Nature 2001, 409, 860–921. [Google Scholar]
- Dinger, M.E.; Amaral, P.P.; Mercer, T.R.; Mattick, J.S. Pervasive transcription of the eukaryotic genome: Functional indices and conceptual implications. Brief. Funct. Genom. Proteom. 2009, 8, 407–423. [Google Scholar] [CrossRef] [PubMed]
- Clark, M.B.; Amaral, P.P.; Schlesinger, F.J.; Dinger, M.E.; Taft, R.J.; Rinn, J.L.; Ponting, C.P.; Stadler, P.F.; Morris, K.V.; Morillon, A.; et al. The reality of pervasive transcription. PLoS Biol. 2011, 9, 5–10. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hangauer, M.J.; Vaughn, I.W.; McManus, M.T. Pervasive Transcription of the Human Genome Produces Thousands of Previously Unidentified Long Intergenic Noncoding RNAs. PLoS Genet. 2013, 9, e1003569. [Google Scholar] [CrossRef] [PubMed]
- Jacquier, A. The complex eukaryotic transcriptome: Unexpected pervasive transcription and novel small RNAs. Nat. Rev. Genet. 2009, 10, 833–844. [Google Scholar] [CrossRef]
- Iyer, M.K.; Niknafs, Y.S.; Malik, R.; Singhal, U.; Sahu, A.; Hosono, Y.; Barrette, T.R.; Prensner, J.R.; Evans, J.R.; Zhao, S.; et al. The landscape of long noncoding RNAs in the human transcriptome. Nat. Genet. 2015, 47, 199–208. [Google Scholar] [CrossRef]
- Struhl, K. Transcriptional noise and the fidelity of initiation by RNA polymerase II. Nat. Struct. Mol. Biol. 2007, 14, 103–105. [Google Scholar] [CrossRef]
- Ebisuya, M.; Yamamoto, T.; Nakajima, M.; Nishida, E. Ripples from neighbouring transcription. Nat. Cell Biol. 2008, 10, 1106–1113. [Google Scholar] [CrossRef]
- Storz, G.; Altuvia, S.; Wassarman, K.M. An Abundance of RNA Regulators. Annu. Rev. Biochem. 2005, 74, 199–217. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.Y.; Johnson, R.; Stanton, L.W. Human long non-coding RNAs promote pluripotency and neuronal differentiation by association with chromatin modifiers and transcription factors. EMBO J. 2012, 31, 522–533. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, Y.G.; Satpathy, A.T.; Chang, H.Y. Gene regulation in the immune system by long noncoding RNAs. Nat. Immunol. 2017, 18, 962–972. [Google Scholar] [CrossRef]
- Volders, P.J.; Anckaert, J.; Verheggen, K.; Nuytens, J.; Martens, L.; Mestdagh, P.; Vandesompele, J. Lncipedia 5: Towards a reference set of human long non-coding rnas. Nucleic Acids Res. 2019, 47, D135–D139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shearwin, K.E.; Callen, B.P.; Egan, J.B. Transcriptional interference—A crash course. Trends Genet. 2005, 21, 339–345. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Herculano-Houzel, S. The human brain in numbers: A linearly scaled-up primate brain. Front. Hum. Neurosci. 2009, 3, 31. [Google Scholar] [CrossRef] [Green Version]
- Raichle, M.E.; MacLeod, A.M.; Snyder, A.Z.; Powers, W.J.; Gusnard, D.A.; Shulman, G.L. A default mode of brain function. Proc. Natl. Acad. Sci. USA 2001, 98, 676–682. [Google Scholar] [CrossRef] [Green Version]
- Biswal, B.B.; Mennes, M.; Zuo, X.N.; Gohel, S.; Kelly, C.; Smith, S.M.; Beckmann, C.F.; Adelstein, J.S.; Buckner, R.L.; Colcombe, S.; et al. Toward discovery science of human brain function. Proc. Natl. Acad. Sci. USA 2010, 107, 4734–4739. [Google Scholar] [CrossRef] [Green Version]
- Hsieh, J.; Gage, F.H. Epigenetic control of neural stem cell fate. Curr. Opin. Genet. Dev. 2004, 14, 461–469. [Google Scholar] [CrossRef]
- Cao, X.; Yeo, G.; Muotri, A.R.; Kuwabara, T.; Gage, F.H. Noncoding RNAs in the mammalian central nervous system. Annu. Rev. Neurosci. 2006, 29, 77–103. [Google Scholar] [CrossRef] [Green Version]
- Clark, B.S.; Blackshaw, S. Long non-coding RNA-dependent transcriptional regulation in neuronal development and disease. Front. Genet. 2014, 5, 164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rajman, M.; Schratt, G. MicroRNAs in neural development: From master regulators to fine-tuners. Development 2017, 144, 2310–2322. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, Z.; Li, Z. miRNAs in synapse development and synaptic plasticity. Curr. Opin. Neurobiol. 2017, 45, 24–31. [Google Scholar] [CrossRef] [PubMed]
- Ng, S.Y.; Lin, L.; Soh, B.S.; Stanton, L.W. Long noncoding RNAs in development and disease of the central nervous system. Trends Genet. 2013, 29, 461–468. [Google Scholar] [CrossRef] [PubMed]
- Hussain, S.; Bashir, Z.I. The epitranscriptome in modulating spatiotemporal RNA translation in neuronal post-synaptic function. Front. Cell. Neurosci. 2015, 9, 420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ramos, A.D.; Andersen, R.E.; Liu, S.J.; Nowakowski, T.J.; Hong, S.J.; Gertz, C.C.; Salinas, R.D.; Zarabi, H.; Kriegstein, A.R.; Lim, D.A. The long noncoding RNA Pnky regulates neuronal differentiation of embryonic and postnatal neural stem cells. Cell Stem Cell 2015, 16, 439–447. [Google Scholar] [CrossRef] [Green Version]
- Mercer, T.R.; Dinger, M.E.; Sunkin, S.M.; Mehler, M.F.; Mattick, J.S. Specific expression of long noncoding RNAs in the mouse brain. Proc. Natl. Acad. Sci. USA 2008, 105, 716–721. [Google Scholar] [CrossRef] [Green Version]
- Brosius, J.; Raabe, C.A. What is an RNA? A top layer for RNA classification. RNA Biol. 2016, 13, 140–144. [Google Scholar] [CrossRef] [Green Version]
- Sutcliffe, J.G.; Milnert, R.J.; Bloomt, F.E.; Lerner, R.A. Common 82-nucleotide sequence unique to brain RNA. Proc. Natl. Acad. Sci. USA 1982, 79, 4942–4946. [Google Scholar] [CrossRef] [Green Version]
- Sutcliffe, J.R.; Milner, J.; Gottesfeld, R.L. Identifier sequences are transcribed specifically in brain. Nature 1984, 308, 237–241. [Google Scholar] [CrossRef]
- Watson, J.B.; Sutcliffe, J.G. Primate brain-specific cytoplasmic transcript of the Alu repeat family. Mol. Cell. Biol. 1987, 7, 3324–3327. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacoangeli, A.; Tiedge, H. Translational control at the synapse: Role of RNA regulators. Trends Biochem. Sci. 2013, 38, 47–55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacoangeli, A.; Dosunmu, A.; Eom, T.; Stefanov, D.G.; Tiedge, H. Regulatory BC1 RNA in cognitive control. Learn. Mem. 2017, 24, 267–277. [Google Scholar] [CrossRef] [Green Version]
- Mus, E.; Hof, P.R.; Tiedge, H. Dendritic BC200 RNA in aging and in Alzheimer s disease. Proc. Natl. Acad. Sci. USA 2007, 104, 10679–10684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- DeChiara, T.M.; Brosius, J. Neural BC1 RNA: cDNA clones reveal nonrepetitive sequence content. Proc. Natl. Acad. Sci. USA 1987, 84, 2624–2628. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martignetti, J.A.; Brosius, J. Neural BC1 RNA as an evolutionary marker: Guinea pig remains a rodent. Proc. Natl. Acad. Sci. USA 1993, 90, 9698–9702. [Google Scholar] [CrossRef] [Green Version]
- Samson, J.; Cronin, S.; Dean, K. BC200 (BCYRN1)—The shortest, long, non-coding RNA associated with cancer. Non-Coding RNA Res. 2018, 3, 131–143. [Google Scholar] [CrossRef]
- Muslimov, I.A.; Iacoangeli, A.; Eom, T.; Ruiz, A.; Lee, M.; Stephenson, S.; Ginzler, E.M.; Tiedge, H. Neuronal BC RNA transport impairments caused by systemic lupus erythematosus autoantibodies. J. Neurosci. 2019, 39, 7759–7777. [Google Scholar] [CrossRef]
- Rozhdestvensky, T.S.; Kopylov, A.M.; Brosius, J.; Hüttenhofer, A. Neuronal BC1 RNA structure: Evolutionary conversion of a tRNAAla domain into an extended stem-loop structure. Rna 2001, 7, 722–730. [Google Scholar] [CrossRef] [Green Version]
- Khanam, T.; Rozhdestvensky, T.S.; Bundman, M.; Galiveti, C.R.; Handel, S.; Sukonina, V.; Jordan, U.; Brosius, J.; Skryabin, B.V. Two primate-specific small non-protein-coding RNAs in transgenic mice: Neuronal expression, subcellular localization and binding partners. Nucleic Acids Res. 2007, 35, 529–539. [Google Scholar] [CrossRef] [Green Version]
- Kim, Y.; Lee, J.; Shin, H.; Jang, S.; Kim, S.C.; Lee, Y. Biosynthesis of brain cytoplasmic 200 RNA. Sci. Rep. 2017, 7, 6884. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martignetti, J.A.; Brosius, J. BC1 RNA: Transcriptional analysis of a neural cell-specific RNA polymerase III transcript. Mol. Cell. Biol. 1995, 15, 1642–1650. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chen, W.; Bocker, W.; Brosius, J.; Tiedge, H. Expression of neural BC200 RNA in human tumours. J. Pathol. 1997, 183, 345–351. [Google Scholar] [CrossRef]
- Singh, R.; Gupta, S.C.; Peng, W.X.; Zhou, N.; Pochampally, R.; Atfi, A.; Watabe, K.; Lu, Z.; Mo, Y.Y. Regulation of alternative splicing of Bcl-x by BC200 contributes to breast cancer pathogenesis. Cell Death Dis. 2016, 7, e2262. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hu, T.; Lu, Y.R. BCYRN1, a c-MYC-activated long non-coding RNA, regulates cell metastasis of non-small-cell lung cancer. Cancer Cell Int. 2015, 15, 36. [Google Scholar] [CrossRef] [Green Version]
- Sonawane, A.R.; Platig, J.; Fagny, M.; Chen, C.Y.; Paulson, J.N.; Lopes-Ramos, C.M.; DeMeo, D.L.; Quackenbush, J.; Glass, K.; Kuijjer, M.L. Understanding Tissue-Specific Gene Regulation. Cell Rep. 2017, 21, 1077–1088. [Google Scholar] [CrossRef] [Green Version]
- Weirauch, M.T.; Yang, A.; Albu, M.; Cote, A.G.; Montenegro-Montero, A.; Drewe, P.; Najafabadi, H.S.; Lambert, S.A.; Mann, I.; Cook, K.; et al. Determination and inference of eukaryotic transcription factor sequence specificity. Cell 2014, 158, 1431–1443. [Google Scholar] [CrossRef] [Green Version]
- Szklarczyk, D.; Franceschini, A.; Wyder, S.; Forslund, K.; Heller, D.; Huerta-Cepas, J.; Simonovic, M.; Roth, A.; Santos, A.; Tsafou, K.P.; et al. STRING v10: Protein-protein interaction networks, integrated over the tree of life. Nucleic Acids Res. 2015, 43, D447–D452. [Google Scholar] [CrossRef]
- Muslimov, I.A.; Banker, G.; Brosius, J.; Tiedge, H. Activity-dependent regulation of dendritic BC1 RNA in hippocampal neurons in culture. J. Cell Biol. 1998, 141, 1601–1611. [Google Scholar] [CrossRef]
- Maekawa, S.; Imamachi, N.; Irie, T.; Tani, H.; Matsumoto, K.; Mizutani, R.; Imamura, K.; Kakeda, M.; Yada, T.; Sugano, S.; et al. Analysis of RNA decay factor mediated RNA stability contributions on RNA abundance. BMC Genom. 2015, 16, 154. [Google Scholar] [CrossRef] [Green Version]
- Wang, H.; Iacoangeli, A.; Popp, S.; Muslimov, I.A.; Imataka, H.; Sonenberg, N.; Lomakin, I.B.; Tiedge, H. Dendritic BC1 RNA: Functional role in regulation of translation initiation. J. Neurosci. 2002, 22, 10232–10241. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Muddashetty, R.; Khanam, T.; Kondrashov, A.; Bundman, M.; Iacoangeli, A.; Kremerskothen, J.; Duning, K.; Barnekow, A.; Hüttenhofer, A.; Tiedge, H.; et al. Poly(A)-binding protein is associated with neuronal BC1 and BC200 ribonucleoprotein particles. J. Mol. Biol. 2002, 321, 433–445. [Google Scholar] [CrossRef]
- Kondrashov, A.V.; Kiefmann, M.; Ebnet, K.; Khanam, T.; Muddashetty, R.S.; Brosius, J. Inhibitory effect of naked neural BC1 RNA or BC200 RNA on eukaryotic in vitro translation systems is reversed by poly(A)-binding protein (PABP). J. Mol. Biol. 2005, 353, 88–103. [Google Scholar] [CrossRef] [PubMed]
- Hinnebusch, A.G.; Ivanov, I.P.; Sonenberg, N. Translational control by 5′-untranslated regions of eukaryotic mRNAs. Science 2016, 352, 1413–1416. [Google Scholar] [CrossRef] [PubMed]
- Lin, D.; Pestova, T.V.; Hellen, C.U.T.; Tiedge, H. Translational Control by a Small RNA: Dendritic BC1 RNA Targets the Eukaryotic Initiation Factor 4A Helicase Mechanism. Mol. Cell. Biol. 2008, 28, 3008–3019. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eom, T.; Muslimov, I.A.; Tsokas, P.; Berardi, V.; Zhong, J.; Sacktor, T.C.; Tiedge, H. Neuronal BC RNAs cooperate with eIF4B to mediate activity-dependent translational control. J. Cell Biol. 2014, 207, 237–252. [Google Scholar] [CrossRef]
- Eom, T.; Berardi, V.; Zhong, J.; Risuleo, G.; Tiedge, H. Dual nature of translational control by regulatory BC RNAs. Mol. Cell. Biol. 2011, 31, 4538–4549. [Google Scholar] [CrossRef] [Green Version]
- Skryabin, B.V.; Sukonina, V.; Jordan, U.; Lewejohann, L.; Sachser, N.; Muslimov, I.; Tiedge, H.; Brosius, J. Neuronal Untranslated BC1 RNA: Targeted Gene Elimination in Mice. Mol. Cell. Biol. 2003, 23, 6435–6441. [Google Scholar] [CrossRef] [Green Version]
- Lewejohann, L.; Skryabin, B.V.; Sachser, N.; Prehn, C.; Heiduschka, P.; Thanos, S.; Jordan, U.; Dell’Omo, G.; Vyssotski, A.L.; Pleskacheva, M.G.; et al. Role of a neuronal small non-messenger RNA: Behavioural alterations in BC1 RNA-deleted mice. Behav. Brain Res. 2004, 154, 273–289. [Google Scholar] [CrossRef] [Green Version]
- Zhong, J.; Chuang, S.-C.; Bianchi, R.; Zhao, W.; Lee, H.; Fenton, A.A.; Wong, R.K.S.; Tiedge, H. BC1 regulation of metabotropic glutamate receptor-mediated neuronal excitability. J. Neurosci. 2009, 29, 9977–9986. [Google Scholar] [CrossRef] [Green Version]
- MacCarrone, M.; Rossi, S.; Bari, M.; De Chiara, V.; Rapino, C.; Musella, A.; Bernardi, G.; Bagni, C.; Centonze, D. Abnormal mGlu 5 receptor/endocannabinoid coupling in mice lacking FMRP and BC1 RNA. Neuropsychopharmacology 2010, 35, 1500–1509. [Google Scholar] [CrossRef] [PubMed]
- Briz, V.; Restivo, L.; Pasciuto, E.; Juczewski, K.; Mercaldo, V.; Lo, A.C.; Baatsen, P.; Gounko, N.V.; Borreca, A.; Girardi, T.; et al. The non-coding RNA BC1 regulates experience-dependent structural plasticity and learning. Nat. Commun. 2017, 8, 293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Shin, H.; Lee, J.; Kim, Y.; Jang, S.; Lee, Y.; Kim, S.; Lee, Y. Knockdown of BC200 RNA expression reduces cell migration and invasion by destabilizing mRNA for calcium-binding protein S100A11. RNA Biol. 2017, 14, 1418–1430. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tiedge, H.; Fremeau, R.T.; Weinstock, P.H.; Arancio, O.; Brosius, J. Dendritic location of neural BC1 RNA. Proc. Natl. Acad. Sci. USA 1991, 88, 2093–2097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Di Liegro, C.M.; Schiera, G.; Di Liegro, I. Regulation of mRNA transport, localizationand translation in the nervous system of mammals (Review). Int. J. Mol. Med. 2014, 33, 747–762. [Google Scholar] [CrossRef] [Green Version]
- Shan, J.; Munro, T.P.; Barbarese, E.; Carson, J.H.; Smith, R. A molecular mechanism for mRNA trafficking in neuronal dendrites. J. Neurosci. 2003, 23, 8859–8866. [Google Scholar] [CrossRef] [Green Version]
- Tiedge, H.; Chen, W.; Brosius, J. Primary structure, neural-specific expression, and dendritic location of human BC200 RNA. J. Neurosci. 1993, 13, 2382–2390. [Google Scholar] [CrossRef]
- Tiedge, H. K-turn motifs in spatial RNA coding. RNA Biol. 2006, 3, 133–139. [Google Scholar] [CrossRef]
- Muslimov, I.A.; Patel, M.V.; Rose, A.; Tiedge, H. Spatial code recognition in neuronal RNA targeting: Role of RNA-hnRNP A2 interactions. J. Cell Biol. 2011, 194, 441–457. [Google Scholar] [CrossRef] [Green Version]
- Cristofanilli, M.; Iacoangeli, A.; Muslimov, I.A.; Tiedge, H. Neuronal BC1 RNA: Microtubule-dependent dendritic delivery. J. Mol. Biol. 2006, 356, 1118–1123. [Google Scholar] [CrossRef] [Green Version]
- Zalfa, F.; Giorgi, M.; Primerano, B.; Moro, A.; Di Penta, A.; Reis, S.; Oostra, B.; Bagni, C. The Fragile X syndrome protein FMRP associates with BC1 RNA and regulates the translation of specific mRNAs at synapses. Cell 2003, 112, 317–327. [Google Scholar] [CrossRef] [Green Version]
- Iacoangeli, A.; Rozhdestvensky, T.S.; Dolzhanskaya, N.; Tournier, B.; Schütt, J.; Brosius, J.; Denman, R.B.; Khandjian, E.W.; Kindler, S.; Tiedge, H. On BC1 RNA and the fragile X mental retardation protein. Proc. Natl. Acad. Sci. USA 2008, 105, 734–739. [Google Scholar] [CrossRef] [Green Version]
- Booy, E.P.; McRae, E.K.; Ezzati, P.; Choi, T.; Gussakovsky, D.; McKenna, S.A. Comprehensive analysis of the BC200 ribonucleoprotein reveals a reciprocal regulatory function with CSDE1/UNR. Nucleic Acids Res. 2018, 46, 11575–11591. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, S.; Shin, H.; Lee, J.; Kim, Y.; Bak, G.; Lee, Y. Regulation of BC200 RNA-mediated translation inhibition by hnRNP E1 and E2. FEBS Lett. 2017, 591, 393–405. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Huber, K.M.; Gallagher, S.M.; Warren, S.T.; Bear, M.F. Altered synaptic plasticity in a mouse model of fragile X mental retardation. Proc. Natl. Acad. Sci. USA 2002, 99, 7746–7750. [Google Scholar] [CrossRef] [Green Version]
- Verheij, C.; Bakker, C.E.; de Graaff, E.; Keulemans, J.; Willemsen, R.; Verkerk, A.J.M.H.; Galjaard, H.; Reuser, A.J.J.; Hoogeveen, A.T.; Oostra, B.A. Characterization and localization of the FMR-1 gene product. Nature 1993, 363, 722–724. [Google Scholar] [CrossRef]
- Zhong, J.; Chuang, S.C.; Bianchi, R.; Zhao, W.; Paul, G.; Thakkar, P.; Liu, D.; Fenton, A.; Wong, R.K.S.; Tiedge, H. Regulatory BC1 RNA and the fragile X mental retardation protein: Convergent functionality in brain. PLoS ONE 2010, 5, e15509. [Google Scholar] [CrossRef] [Green Version]
- Jacquemont, S.; Hagerman, R.J.; Hagerman, P.J.; Leehey, M.A. Fragile-X syndrome and fragile X-associated tremor/ataxia syndrome: Two faces of FMR1. Lancet Neurol. 2007, 6, 45–55. [Google Scholar] [CrossRef]
- Centonze, D.; Rossi, S.; Mercaldo, V.; Napoli, I.; Ciotti, M.T.; De Chiara, V.; Musella, A.; Prosperetti, C.; Calabresi, P.; Bernardi, G.; et al. Abnormal Striatal GABA Transmission in the Mouse Model for the Fragile X Syndrome. Biol. Psychiatry 2008, 63, 963–973. [Google Scholar] [CrossRef]
- Nelson, D.L.; Orr, H.T.; Warren, S.T. Review the Unstable Repeats—Three Evolving Faces of Neurological Disease. Neuron 2013, 77, 825–843. [Google Scholar] [CrossRef] [Green Version]
- Muslimov, I.A.; Eom, T.; Iacoangeli, A.; Chuang, S.; Renate, K.; Willemsen, R.; Stefanov, D.G.; Wong, R.K.S.; Tiedge, H. BC RNA Mislocalization in the Fragile X Premutation. eNeuro 2018, 5. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iacoangeli, A.; Lin, Y.; Morley, E.J.; Muslimov, I.A.; Bianchi, R.; Reilly, J.; Weedon, J.; Diallo, R.; Böcker, W.; Tiedge, H. BC200 RNA in invasive and preinvasive breast cancer. Carcinogenesis 2004, 25, 2125–2133. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.; Kim, Y.; Kim, M.; Lee, Y. BC200 RNA: An Emerging Therapeutic Target and Diagnostic Marker for Human Cancer. Mol. Cells 2018, 41, 993–999. [Google Scholar] [PubMed]
- Peng, J.; Hou, F.; Feng, J.; Xu, S.X.; Meng, X.Y. Long non-coding RNA BCYRN1 promotes the proliferation and metastasis of cervical cancer via targeting microRNA-138 in vitro and in vivo. Oncol. Lett. 2018, 15, 5809–5818. [Google Scholar] [CrossRef] [PubMed]
- Zhai, H.; Li, Y. BCYRN1 is correlated with progression and prognosis in gastric cancer. Biosci. Rep. 2019, 39, BSR20190505. [Google Scholar] [CrossRef] [Green Version]
- Taft, R.; Mattick, J. Increasing biological complexity is positively correlated with the relative genome-wide expansion of non-protein-coding DNA sequences. Genome Biol. 2003, 5, 1. [Google Scholar] [CrossRef] [Green Version]
- Rogaev, E.I. Small RNAs in Human Brain Development and Disorders. Biochemistry 2005, 70, 1404–1407. [Google Scholar] [CrossRef]
- Ravasi, T.; Suzuki, H.; Pang, K.C.; Katayama, S.; Furuno, M.; Okunishi, R.; Fukuda, S.; Ru, K.; Frith, M.C.; Gongora, M.M.; et al. Experimental validation of the regulated expression of large numbers of non-coding RNAs from the mouse genome. Genome Res. 2006, 16, 11–19. [Google Scholar] [CrossRef] [Green Version]
- Kim, J.; Krichevsky, A.; Grad, Y.; Hayes, G.D.; Kosik, K.S.; Church, G.M.; Ruvkun, G. Identification of many microRNAs that copurify with polyribosomes in mammalian neurons. Proc. Natl. Acad. Sci. USA 2004, 101, 322–327. [Google Scholar] [CrossRef] [Green Version]
- Mehler, M.F.; Mattick, J.S. Non-coding RNAs in the nervous system. J. Physiol. 2006, 575, 333–341. [Google Scholar] [CrossRef]
Figure 1.
Brain cytoplasmic RNAs (BC RNAs) in neurons. (A) The journey of BC RNAs in neurons. (B) Promoters of BC RNAs consist of the core motif, TATA-like sequence in the upstream sequence and A-box and B-box in the internal sequence. TATA-binding protein (TBP) recognizes promoter sequences and recruits the polymerase III complex. These processes are regulated by sophisticated neuronal-specific activating pathway. (C) BC RNAs bind with heterogeneous ribonucleoprotein A2 (hnRNP A2) in the soma. RNP complex is migrated into dendrite along the microtubule. (D) Mitogen-activated protein kinase kinase/Extracellular signal-regulated kinase (MEK/ERK) promotes the translation of dendritic mRNA and BC RNAs inhibit excessive translation. BC RNAs are key factors maintaining translation balance in the postsynaptic region.
Figure 1.
Brain cytoplasmic RNAs (BC RNAs) in neurons. (A) The journey of BC RNAs in neurons. (B) Promoters of BC RNAs consist of the core motif, TATA-like sequence in the upstream sequence and A-box and B-box in the internal sequence. TATA-binding protein (TBP) recognizes promoter sequences and recruits the polymerase III complex. These processes are regulated by sophisticated neuronal-specific activating pathway. (C) BC RNAs bind with heterogeneous ribonucleoprotein A2 (hnRNP A2) in the soma. RNP complex is migrated into dendrite along the microtubule. (D) Mitogen-activated protein kinase kinase/Extracellular signal-regulated kinase (MEK/ERK) promotes the translation of dendritic mRNA and BC RNAs inhibit excessive translation. BC RNAs are key factors maintaining translation balance in the postsynaptic region.

Figure 2.
Secondary structures of BC1 RNA and BC200 RNA. (A,B) BC1 RNA and BC200 RNA are functional analogues. Secondary structures of BC1 RNA (A) and BC200 RNA (B) with common functional motifs (the GA kink motif in the red box, the poly A region in the yellow box, and the C-loop motif in the blue box).
Figure 2.
Secondary structures of BC1 RNA and BC200 RNA. (A,B) BC1 RNA and BC200 RNA are functional analogues. Secondary structures of BC1 RNA (A) and BC200 RNA (B) with common functional motifs (the GA kink motif in the red box, the poly A region in the yellow box, and the C-loop motif in the blue box).

Figure 3.
History of BC RNA studies in neurons.


{kind=link}
{kind=link}
{kind=link}
Table 1.
Neuronal diseases associated with BC RNAs.
| Disease | BC RNAs | Description |
|---|---|---|
| Alzheimer’s Disease | Up-regulation | Overexpressed in Brodmann’s area 9 and accumulated in the soma |
| Fragile X Syndrome | Down-regulation | BC1-KO mice are similar to FMRP-KO mice |
| Fragile X Premutation Disorders | Mislocalization | CGG repeats inhibits dendritic localization of BC RNAs |
| Systemic Lupus Erythematosus | Mislocalization | Anti-BC antibodies inhibit dendritic localization of BC RNAs |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Lee, Y.; Lee, H.-S.; Kim, M.; Shin, H. Brain Cytoplasmic RNAs in Neurons: From Biosynthesis to Function. Biomolecules 2020, 10, 313. https://doi.org/10.3390/biom10020313
AMA Style
Lee Y, Lee H-S, Kim M, Shin H. Brain Cytoplasmic RNAs in Neurons: From Biosynthesis to Function. Biomolecules. 2020; 10(2):313. https://doi.org/10.3390/biom10020313
Chicago/Turabian StyleLee, Younghoon, Hee-Seung Lee, Meehyein Kim, and Heegwon Shin. 2020. "Brain Cytoplasmic RNAs in Neurons: From Biosynthesis to Function" Biomolecules 10, no. 2: 313. https://doi.org/10.3390/biom10020313
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.