Oxidative Stress in Male Infertility: Causes, Effects in Assisted Reproductive Techniques, and Protective Support of Antioxidants



1
Biotechnology of Animal and Human Reproduction (TechnoSperm), Institute of Food and Agricultural Technology, University of Girona, E-17003 Girona, Spain
2
Unit of Cell Biology, Department of Biology, Faculty of Sciences, University of Girona, E-17003 Girona, Spain
*
Authors to whom correspondence should be addressed.
Biology 2020, 9(4), 77; https://doi.org/10.3390/biology9040077
Submission received: 25 March 2020
/
Revised: 8 April 2020
/
Accepted: 9 April 2020
/
Published: 10 April 2020
(This article belongs to the Special Issue Oxidative Stress in Gametes and Embryos)
{kind=link}
{kind=link}
Abstract
:The spermatozoon is a highly specialized cell, whose main function is the transport of the intact male genetic material into the oocyte. During its formation and transit throughout male and female reproductive tracts, sperm cells are internally and externally surrounded by reactive oxygen species (ROS), which are produced from both endogenous and exogenous sources. While low amounts of ROS are known to be necessary for crucial physiological sperm processes, such as acrosome reaction and sperm–oocyte interaction, high levels of those species underlie misbalanced antioxidant-oxidant molecules, generating oxidative stress (OS), which is one of the most damaging factors that affect sperm function and lower male fertility potential. The present work starts by reviewing the different sources of oxidative stress that affect sperm cells, continues by summarizing the detrimental effects of OS on the male germline, and discusses previous studies addressing the consequences of these detrimental effects on natural pregnancy and assisted reproductive techniques effectiveness. The last section is focused on how antioxidants can counteract the effects of ROS and how sperm fertilizing ability may benefit from these agents.
1. Introduction
Human infertility is recognized as a disease by the World Health Organization (WHO) and has nowadays an estimated prevalence of 7–15%, with 50–80 million people being affected around the world [1,2]. These high ranks of incidence exemplify the differences between developed and developing countries, the latter being the ones with higher rates, a fact that has been associated to lower familiar income and less health-care accessibility [3,4]. Despite these worrying numbers, mounting evidence from surveys and comparative studies suggest that infertility grows year over year [4,5]. In this scenario, and despite the high relevance given to the female factor in the nineties, it is now well accepted that dysfunctions can also affect spermatozoa, so that a male factor is thought to be present in half of the infertile couples, being the only or main cause of infertility in about 20–30% of cases [6,7,8]. For this reason, research focused on the male factor has been increasing over the last decades, revealing new knowledge about causes and treatments of infertility. Although significant advances on understanding the etiology of male infertility have been made, little is still known about the underlying physiopathology in a high number of particular cases. In most couples, attributing a single cause to infertility is difficult since there is usually a multifactorial, sometimes unknown, pathology. Recent research has come up with different indicators and biomarkers that are able to explain male infertility, such as some specific related genes, concrete epigenetic profiles, and several mRNAs that are responsible for poor sperm quality and/or embryo development [9,10,11]. On the other hand, anatomical affectations such as vas deferens agenesia, hormonal issues causing azoospermia, or varicocele are also factors that help diagnose infertility in the male [12,13]. Moreover, increasing male age, environmental pollution, diet, heat stress, and obesity are also known to be causes for a reduction of sperm quality. In these latter ones, however, it is difficult to establish their precise incidence in the physiopathology of a single male, despite the efforts in defining ranks or risk areas for each parameter. In a scenario where a human male is exposed to some of these exogenous affectations, an increase in oxidative stress is usually observed [14]. An excess of reactive oxygen species (ROS) can cause unbalance with the antioxidant capacity, reaching pathological levels and originating fertility issues. Infertility caused by oxidative stress has been widely studied for years, and multiple studies analyzing its clinical effect have been performed, in relation to both natural fertility and assisted reproductive techniques. Therefore, the aims of the present review are (a) to summarize the causes and consequences of oxidative stress in male fertility; (b) to provide an update about the clinical association between oxidative stress and fertility rates; and (c) to discuss the role of antioxidants in ameliorating fertility outcomes.
2. Sources of Oxidative Stress with Potential Effects to the Male Germline
Oxidative stress is related to the disbalance of oxidant molecules in cells. Reactive oxygen species refer to oxygen-derivate molecules that are highly reactive due to their free electrons or radicals. These group of molecules, which have a half-life of nanoseconds, include superoxide (·O2−), hydrogen peroxide (H2O2), proxyl radical (·R OO), or hydroxyl (·OH−). Less common but also present in sperm cells are the reactive nitrogen species (RNS), which include nitric oxide (·NO), dinitrogen trioxide (N2O3), and peroxinitrite (ONOO-). The presence of oxidative stress affecting sperm function has been consistently reported since the late forties and early fifties, when different authors pointed out that hydrogen peroxide exerted a detrimental impact on sperm [15]. Nowadays, this damage is irrefutable and, over the last decades, multiple investigations have been oriented towards identifying the possible sources of ROS in order to understand their action mechanism and their consequences. This knowledge has allowed the discovery of new treatment options for human patients, thus improving their reproductive chances. Regarding the male germline, reactive oxygen species can be produced either endogenous or exogenous. From here on, we review these two ways of producing ROS that, despite being necessary at low levels, contribute to a pathological state at high levels.
2.1. Endogenous Sources of ROS
One should note that the spermatozoon itself is a source of ROS due to its metabolic activity, and that other immature sperm cells present in the semen are also important sources of free radicals. While disbalances of ROS have disruptive effects that will be described below in other sections, low levels of ROS are known to be necessary for the sperm cell to perform natural functions. For instance, nitric oxide and hydrogen peroxide are necessary compounds to achieve capacitation, enabling acrosome reaction, which is controlled by reactive oxygen species [16,17]. Moreover, sperm hyperactivation or its interaction with the oocyte is mediated by ROS, thereby being also essential for achieving their fertilizing ability [18,19] (Figure 1). For these reasons, after spermatogenesis and epididymal maturation sperm cells have low levels of oxidative radicals.
Endogenous free radicals can be generated as a by-product of cell metabolism, or by enzymatic activity. Sperm metabolism is a major source of ROS (Figure 1). The activity of mitochondria, which are located in the mid-piece, is, together with glycolysis, essential for sperm motility and capacitation. These mitochondria generate ATP through respiratory electron chain and oxidative phosphorylation, which are based on transferring electrons from inner mitochondrial membrane complexes to oxygen and on pumping of protons to the intermembrane space. These protons are ultimately used to synthesize ATP via complex V (ATP synthase). In this scenario, ROS are by-products of the electron chain activity and are especially generated at complexes I and III, which mainly release superoxide and hydroxyl radicals into the matrix and the intermembrane space [20]. These free radicals are then converted into hydrogen peroxide by superoxide dismutase.
During spermatogenesis, release of sperm with retained cytoplasm usually occurs. Retention of cytoplasm leads to non-functional and immature sperm which retain glucose-6-phosphate dehydrogenase (G6PDH) that enables the production of intracellular β-nicotinamide adenine dinucleotide phosphate (NADPH). This intracellular NADPH is processed via NADPH oxidase which regenerates NADPH to NADP converting O2 to superoxide, which is converted to hydrogen peroxide at the intermembrane space by the superoxide dismutase [21,22].
The hydrogen peroxide generated through these two different ways is scavenged by glutatione peroxidase or glutathione-s-reductase, which use reduced glutathione (GSH) as an electrone donor. Reduced glutathione is maintained by an ATP-consuming process, through glutathione synthetase (GSS) or glutathione reductase (GSR), which transforms oxidized glutathione (GSSG) into reduced glutathione in a NADPH dependent way [23,24].
2.2. Exogenous Sources of ROS
2.2.1. Varicocele
Varicocele is a dilatation of the pampiniform plexus of the spermatic cord that is considered the most common correctable cause of male infertility. The incidence of varicocele in the general population is about 15%, and different studies have determined that its incidence among infertile men is about 35–44%, these figures increasing up to 45–81% in the case of secondary infertility [12]. The dilatation of varicose veins coupled to insufficient venous valves cause a blood reflux and an increase in the blood pressure to the vein wall; this increases testis temperature, which usually has to be 2 °C lower than that of the body. These two varicocele consequences lead to an increase of reactive oxygen species and a reduction of antioxidant capacity [25] (Figure 1). Varicoceles are divided into different grades (subclinical and clinical, with grades 1, 2, or 3) according to their clinical features. It is known that their effect is higher as grade increases, and several studies have pointed out that varicocelectomy, the most common procedure for varicocele correction, is an effective method to reduce sperm OS and sperm DNA damage in clinical varicoceles [26,27].
2.2.2. Infections and Leucocytospermia
Immune response against infections cause inflammation of tissues to promote leucocyte infiltration. Leukocytes are an important source of oxidative stress, and it has been described that a single leukocyte produces 1000 times more ROS than a single spermatozoon, via increasing NADPH production [28] (Figure 1). Although every ejaculate contains a certain number of leukocytes, leucocytospermia consists of an increase in that number and usually results from infections [29]. While the increase in ROS has been reported to affect both leukospermic and non-leukospermic patients, it is apparent that the extent at which patients with augmented leukocyte counts suffer from higher oxidative stress is higher [30]. This increment has been demonstrated to be associated to a detrimental impact on different conventional semen parameters, such as motility, morphology, and concentration [31,32,33].
2.2.3. Alcohol and Tobacco
It is well known that consumption of alcohol, tobacco, and different recreational drugs contribute to serious, negative effects on the organism. Regarding alcohol, a study in rats concluded that continued alcohol intake causes a decrease in testicular reduced glutathione concentration, a decrease in testicular superoxide dismutase activity, an increase in testicular malondialdehyde concentration, and an increase in sperm DNA damage. In addition, fertility rates of male rats ingesting alcohol were demonstrated to be lower than those of the control group [34]. Other studies reported similar results, describing detrimental effects on mitochondria, with a significant increase in ROS generation, and observing epigenetic modifications in the germline [35,36]. In addition, different studies have shown that chronic ethanol intake has been related to a decrease of cell proliferation in testes; to the induction of testicular apoptosis, increasing the Fas ligand and upregulating p53 gene expression; and to epididymal damage [37,38,39]. A deregulation of the apoptotic response at testicular level has been described as an issue caused by oxidative stress, and named as abortive apoptosis [40,41]. Moreover, autophagy can be also activated as a protective role to cooperate with apoptosis during spermatogenesis [42,43]
On the other hand, tobacco smokers are exposed to thousands of chemicals, which are demonstrated to be carcinogenic and the cause of several diseases that may lead to death. Most of these chemicals are demonstrated to increase free radicals and ROS coupled to a reduction of antioxidant activity, which leads to a higher rate of sperm DNA fragmentation and loss of sperm motility [44,45] (Figure 1).
2.2.4. Physical Exercise and Heat Stress
Exercise is beneficial for different aspects of health. In the case of reproductive physiology, while improved fertility potential has been identified in animals exposed to regular exercise [46] and sedentary men have been reported to present worse sperm quality [47,48], other studies have shown that exercise has no improving effect on conventional spermiogram parameters [49]. In addition, intense physical exercise or cycling is considered to be detrimental for fertility, and both an increase in oxidative stress and a reduction in sperm motility were observed in studies focusing on these types of exercise [50,51]. This increment in oxidative stress and the reduction in sperm quality could also be related to an increase in testicular temperature (Figure 1). Scrotal temperature is known to be 2 °C lower than that the body core temperature, and increases in this value have been shown to impair sperm quality. In fact, it has been reported that every 1 °C of increase correlates to a 14% drop in sperm production [52]. In the same way, some reports have shown that habits such as prolonged car sitting, taking regular sauna bath, or wearing tight fitting underwear can have an impact on testis heat stress, reducing sperm quality [53].
2.2.5. Radiations and Pollution
Radiations can be divided into ionizing and non-ionizing. Non-ionizing radiation is the most used by human beings, and one can find in this category from cell phones, which use extremely low frequency, to microwave ovens and radars, which are in the radio frequency range [54]. The effect of non-ionizing radiations produced by mobile phones, microwaves or WIFI devices to sperm oxidative stress and its fertilizing ability is a topic of increasing interest among the scientific community. Different multiple in vitro and in vivo studies aiming at addressing this issue by testing sperm from animal models and human beings have shown that mobile phone’s radiation causes an increase in reactive oxygen species that induces lipid peroxidation and a decline in the antioxidant capacity, induced, amongst others, by the decrease in reduced glutathione levels [55,56,57,58]. Moreover, other studies have associated laptop computer WIFI to increases of DNA damage and decreases of sperm parameters, such as motility, count, and morphology [59,60]. In summary, although non-ionizing radiations are not able to cause DNA alterations directly, they have an indirect potential of affecting fertility through increasing pro-oxidant molecules (Figure 1).
X-rays, γ-rays, and α-particles are ionizing radiations, which are rather more dangerous than the non-ionizing ones at different health levels. Exposure of cells to ionizing radiations increase ROS generation and induce their senescence [61]. Radiation induces direct DNA breaks and potentially affects proteins and membranes through increased ROS levels (Figure 1). In fact, serious health problems, such as different types of cancer, can arise from the exposure to ionizing radiation; in this context, it is worth noting that, due to their lack of antioxidant defense, sperm cells are especially vulnerable [62].
Environmental pollution has been found to increase ROS generation and lead to a reduction in sperm quality. For instance, on the one hand and according to studies analyzing semen from human males exposed to traffic pollution, car smoke pollutants potentially reduce men fertility [63] by affecting membrane lipids, generating DNA damage, and even changing expression patterns of proteins involved in spermatogenesis [64,65,66]. On the other hand, continuous exposure to phthalate derivatives have been correlated to increases in reactive oxygen species and decreases in enzymatic and non-enzymatic antioxidants in animal models [67], and in the seminal plasma of infertile patients involved in assisted reproductive programs [68]. While these are only two examples of how air pollutants may affect men fertility, strong evidence is still required to support the urgent need of reducing environmental toxicants to improve reproductive efficiency [69].
3. Effects of Oxidative Stress to Sperm Components
All the aforementioned causes have been related to increases in intracellular ROS levels, and the particular and specific organization of the sperm cell makes it especially vulnerable to these increments. The sperm cell does not present a DNA repair machinery since it is transcriptionally silent and possesses either no or poor translational activity [70]. Moreover, at the end of spermiogenesis, testicular spermatozoa lose their cytoplasm, leaving their DNA highly vulnerable to ROS. Therefore, neither cytosolic antioxidant enzymes, such as glutathione peroxidase (GPX), catalase (CAT) and superoxide dismutase (SOD), which are the most common cytosolic ROS-scavengers, nor the Base Excision Repair pathway (BER) are present in sperm cells [70]. Regarding the BER pathway, only an active form of 8-oxoguanine DNA glycosylase (OGG1) enzyme, which excises the oxidized base 8-hydroxy-2′-deoxyguanosine (8OHdG), is left [23]. Since downstream enzymes involved in the BER are missing, the process is stopped after OGG1-excission. For this reason, one can usually find ejaculated sperm with damaged DNA. However, evolution has endowed the oocyte with enzymes devoted to paternal DNA repair after fertilization [71], which unequivocally underpins the collaboration between male and female gametes during the first stages of embryo development.
The aforementioned exposure of sperm cells to different sources of ROS, which are free to travel across the membrane and between mid-piece and the nucleus, and their inability to scavenge ROS make sperm cells very susceptible to damage. This fact leads to different affectation grades, especially targeting their membranes and nuclei, inducing lipid peroxidation, protein alterations, and DNA damage.
3.1. Lipid Peroxidation
Sperm plasma membrane has a particular susceptibility to oxidative damage, as it contains high amounts of polyunsaturated fatty acids, which present multiple double bonds, the decohexaenoic acid (DHA) being the most representative one [70,72,73]. These membrane molecules are reactive to oxygen radicals, producing highly reactive lipid aldehydes, like malondialdehyde, that have the potential of causing DNA damage and modify proteins. It has been described that these aldehydes inhibit some antioxidant enzymes like G6PDH which, in turn, reduce the activity of glutathione peroxidase [18,23]. Lipid peroxidation alters membrane fluidity and permeability, which results in sperm motility loss and in a reduced sperm ability to interact with the oocyte [23].
3.2. Protein Modifications
Proteins are the target of redox reactions, which can activate or inactivate their functionality. Depending on the type of ROS, proteins can be altered through thiol oxidation, tyrosine nitration, sulfonation, or glutathionylation. Thiol oxidation is the most common protein modification that targets glycolytic and Krebs cycle enzymes, which leads to a reduction in the efficiency of ATP production; targets α-tubulin, which impairs microtubule polymerization; and targets protamines, which affect chromatin remodeling during spermiogenesis [74].
3.3. Sperm DNA Damage
Sperm DNA fragmentation is the other major effect directly or indirectly caused by oxidative stress. In contact to sperm DNA, ROS cause base modifications, such as 8-OH-guanine and 8-OH-2′-deoxyguanosine which, after being excised by OGG1 enzyme, generate an abasic site that gives place to a single-strand DNA break. Since, as aforementioned, spermatozoa are devoid of enzymes allowing the continuation of the BER pathway and the silencing of sperm chromatin, this break site cannot be repaired by the sperm cell itself and, thus, sperm with fragmented DNA are observed in the ejaculate [70].
Oxidative stress can also promote nuclear decondensation, increasing the DNA susceptibility to be damaged by free radicals, which will thus have easier access to the entire sperm genome. Related with this, incubation of spermatozoa from different animals with H2O2 indicates that DNA breaks occur in a dose-dependent manner; at a final concentration of 10 mM, H2O2 affects around 60% of sperm DNA [75]. In this context, however, it is worth mentioning that detecting these DNA breaks and its type heavily relies upon the sensitivity of the technique used to evaluate sperm DNA fragmentation (i.e., SCSA, SCDt, TUNEL, neutral Comet, alkaline Comet, etc.), which leads to different correlations to clinical outcomes [76].
4. Effects of Oxidative Stress on Fertility Treatments
4.1. Natural Pregnancy
A disbalance of oxidative stress is usually the cause of different alterations heading, in the end, to DNA fragmentation, protein and lipid modifications, and affectations in sperm count, motility, and morphology. Although it is difficult to establish which percentages of infertile patients have an underlying mechanism of oxidative stress, it is known that it could be present as a factor in asthenozoospermic, teratozospermic, and oligozoospermic patients who present reproductive issues [77,78]. All these parameters have been associated in different studies with a reduction in natural pregnancy, both in animal [79,80] and human studies [76,81,82], supporting the evidence that oxidative stress leads to a reduction in natural fertility rates [77,83].
4.2. Assisted Reproduction Techniques
In a scenario where natural pregnancies are impaired for at least 12 months, assisted reproduction techniques (ART) are available treatments for infertile couples, and include intrauterine insemination (IUI), in vitro fertilization (IVF), and intracytoplasmic sperm injection (ICSI). While the contribution of the male factor infertility to the success rates of different ART has been the source of much debate during the last decade, the following two subsections summarize our current knowledge about the effects of oxidative stress on the effectiveness of those treatments.
4.2.1. Intrauterine Insemination
Three systematic reviews and meta-analyses related to the topic have been identified, with the general conclusion that oxidative stress and sperm DNA damage have negative effects on pregnancy rates after IUI. In the first one, the authors report different studies supporting that sperm DNA damage is related to lower pregnancy rates after IUI, since altered sperm DNA can lead to a decrease in pregnancy rates from 24% to 3% [84]. In the second meta-analysis, similar conclusions were reached from the selected studies included in the systematic review, emphasizing that the extent of the impact of sperm DNA damage on pregnancy rates after IUI is higher when male age is over 30 years old [85]. Finally, a meta-analysis by Belgian researchers including 940 IUI cycles revealed that low DNA damage increases by 3.15 the chances of achieving clinical pregnancy after IUI [86].
4.2.2. IVF and ICSI
Different meta-analyses aimed at addressing the relationship between sperm DNA damage and IVF/ICSI success rates have been conducted. Regarding IVF, separate studies have concluded that sperm DNA damage may have an important influence in preventing clinical pregnancy, increasing miscarriage, or even decreasing embryo quality [84,87,88,89].
As far as ICSI is concerned, data are less clear since whereas some works have reported that sperm DNA damage influences treatment outcomes, such as pregnancy rates and embryo quality [88,89], other meta-analyses have concluded that sperm DNA fragmentation has no influence on ICSI performance [84,90,91].
Inconsistent results from studies analyzing combined IVF and ICSI data are found in the literature, with some meta-analyses supporting that sperm DNA damage causes negative effects on ART [88] and others showing that there is no enough evidence to sustain that assertion [92,93]. These contrasting conclusions may not only be due to the inclusion of different techniques but also to the heterogeneity of different laboratories and populations.
5. Protection against Oxidative Damage and Designed Treatments for Fertility Improvement
The principal mission of the sperm cell is to transport and deliver the intact paternal DNA into the oocyte. For this reason, sperm DNA has evolved to be as much protected as possible from both internal and external sources of damage. This evolution has led to an almost inert state, where histones are replaced in most parts of the genome by protamines, which remain organized in toroids condensing about 50 kilobases of DNA. These toroids are linked to each other by toroid linker regions that are condensed with histones [94]. While the role of histone-condensed regions is currently under debate, recent studies suggest that they could include gene promoter regions, such as CpG-rich islands and satellite repeats [95,96]. In addition, telomeres have also been described to retain a high percentage of histones [97]. Whilst protaminated toroidal regions have been shown to be resilient against nuclease activity and histone-condensed regions seem to be more susceptible to DNA damage, incubation of sperm with H2O2 indicates that oxidative damage can induce DNA breaks in both regions [98].
5.1. Seminal Plasma
Due to the loss of the most part of the cytoplasm and the lack of transcription and translation potential, the sperm cell is devoid or has very low levels of antioxidant enzymes in the cytosol. In order to avoid the disbalance of ROS generating oxidative stress, the low antioxidant activity of mature spermatozoa has to be compensated by the antioxidant capacity of seminal plasma [99]. As a result of this atoning role, seminal plasma has evolved as one of the most known antioxidant fluids, estimating that its antioxidant power is more than 10 times higher than that of the blood [100]. This includes both antioxidant enzymes, such as CAT, SOD, glutathione-s-transferase (GST), GPX and GSR, and small free-radical scavengers, such as vitamin C, polyphenols, carotenoids, and coenzyme Q10 [100,101,102]. Taking into account both pro-oxidant and antioxidant activities, it seems reasonable to suggest that measuring redox balance may provide a broader picture on the relationship between sperm oxidative stress and male infertility [70], this balance being altered either by the increase of ROS or by the decrease of antioxidant activity.
5.2. Varicocelectomy
As mentioned before in Section 2, varicocele is a pathology that induces an increase of oxidative stress in the male reproductive system. The surgical treatment of varicocele has been demonstrated to be an effective method to remove the alterations caused by the affected veins, as blocking blood reflux prevents temperature increases. The effects observed after the recovery from surgery include a decrease in reactive oxygen species and an increase in total antioxidant capacity, which comes from augmenting the levels of antioxidant enzymes and molecules [27,103,104]. As a consequence, DNA damage is reduced after varicocelectomy [105]. In addition, a placebo-controlled, double-blind trial showed that co-treatment with melatonin as an antioxidant may improve the results obtained following surgery [106].
5.3. Antioxidants
Since not only does an increase in oxidative stress reduce sperm quality but also the likelihood of achieving pregnancy (either natural or through ART), the use of exogenous antioxidants has been proposed to balance the ROS:antioxidant ratio and increase the sperm quality. In the literature, several non-enzymatic antioxidants, such as arginine, carnitine, carotenoids, coenzyme Q10, cysteine, reduced glutathione, micronutrients like selenium or zinc, vitamin E, vitamin C, myo-inositol, or resveratrol, have been reported to be utilized to treat different diseases [107]. In the case of ART, exogenous antioxidants have been tested for decades, and several works point out to a positive contribution of these supplements on sperm count, motility, and morphology [108,109,110]. Other studies have also proved that antioxidants are useful to ameliorate lipid peroxidation, reduce DNA base modifications (like 8-OHdG) and fragmentation, and increase total antioxidant capacity [111,112,113,114]. Finally, previous research has also demonstrated that adding cryopreserved sperm with antioxidants increases their quality [115,116,117].
5.3.1. Use of Antioxidants for Natural Pregnancy
Given that sperm quality has been associated to higher success in achieving natural pregnancy, one could suggest that improving sperm function and survival through antioxidant supplementation should cause a positive effect on fertilization success (Figure 2). However, at present, only few studies with randomized and placebo controls prove that association. The latest review of the Cochrane Library including high quality studies concludes that the antioxidant intake by male patients is associated to an increase in live birth rates from 12% to 14–26%, representing between 1.2- and 2.1-fold increase. The same meta-analysis included eleven studies analyzing natural pregnancy and showed that antioxidant supplementation increases pregnancy rates from 7% to 12–26%, representing between 1.7- and 3.7-fold increase [107]. While these results seem to be promising, the authors of that study warned about the increase in miscarriage rates (from 2% to 13%) following antioxidant supplementation. This detrimental effect, however, was concluded from few studies (only three) meeting the quality criteria (randomized, prospective, and double-blinded) [107].
5.3.2. Use of Antioxidants for Intrauterine Insemination
The use of antioxidants with patients undergoing ART is usual. Nevertheless, the potential benefits have been the subject of controversy in the field. Although it seems a common finding that antioxidants have a positive effect on sperm quality parameters, the very low amount of high quality studies foster the controversy on whether or not their use increases male fertility in the case of IUI (Figure 2). For instance, a randomized controlled trial using N–acelylcysteine and a randomized double-blind trial using astaxanthin as antioxidants in couples performing IUI observed an increase in pregnancy rates compared to controls [118,119]. In contrast, a multicenter, double-blind, randomized, placebo-controlled trial published in 2020, and conducted in the United States between 2015 and 2018 concluded that an antioxidant formulation containing vitamins C, E, selenium, L-Carnitine, zinc, folic acid, and lycopene had no effect on sperm quality parameters, pregnancy, or live birth rates [120]. The limitation of these two studies, however, was the low number of patients.
5.3.3. Use of Antioxidants for IVF/ICSI
While randomized and controlled trials are much warranted in this realm, few studies have analyzed the effects of antioxidants on pregnancy rates after IVF or ICSI, usually with small sample sizes, thus reinforcing the need of large studies. A prospective, randomized, double-blind placebo-controlled study performed in Australia between 2004 and 2006 conducted with 60 patients found that a mix of commercial antioxidants (which included lycopene, Vitamins E and C, zinc, selenium, folate, and garlic) taken orally during three months prior to IVF increased both pregnancy and implantation rates [121]. A non-controlled study showed that vitamin E intake during one or three months increased fertilization rates per cycle after IVF regardless of intake time [122]. Furthermore, another non-controlled study conducted with 38 patients presenting a previous failed ICSI cycle and high sperm DNA fragmentation reported that administrating vitamins C and E orally for two months prior to the second ICSI attempt improved clinical pregnancy and implantation rates compared to the first ICSI attempt [123].
6. Conclusions
Despite the vital function of ROS during fertilization, an increase in those species causes oxidative stress and detrimentally affects sperm function. The origin of this increase arises from different endogenous and exogenous sources, such as sperm metabolism or infections, respectively. It is well known that ROS cause a reduction in sperm count and motility, protein alterations, lipid peroxidation, and DNA fragmentation, amongst others. These affectations to sperm quality lead to lower natural pregnancy rates and decline IUI and IVF success. Regarding ICSI, more studies are needed to elucidate which the actual impact of high ROS levels in sperm is. Finally, while using exogenous antioxidants as a method to counteract the adverse effects of oxidative stress has been proven to be effective for natural pregnancy, randomized, double-blind, prospective, and placebo-controlled studies are still needed to demonstrate their effectiveness in assisted reproductive techniques.
Author Contributions
Conceptualization, J.R.-M. and M.Y.; methodology, J.R.-M.; formal analysis, J.R.-M.; investigation, J.R.-M.; resources, M.Y.; writing—original draft preparation, J.R.-M.; writing—review and editing, M.Y.; supervision, J.R.-M. and M.Y.; project administration, J.R.-M. and M.Y.; funding acquisition, M.Y. All authors have read and agreed to the published version of the manuscript.
Funding
This research was funded by the Ministry of Science, Innovation and Universities, Spain (Grants: RYC-2014-15581) and the Regional Government of Catalonia, Spain (2017-SGR-1229).
Acknowledgments
Conflicts of Interest
Authors declare no personal conflicts of interest.
References
- Louis, J.F.; Thoma, M.E.; Sørensen, D.N.; McLain, A.C.; King, R.B.; Sundaram, R.; Keiding, N.; Buck Louis, G.M. The prevalence of couple infertility in the United States from a male perspective: Evidence from a nationally representative sample. Andrology 2013, 1, 741–748. [Google Scholar] [CrossRef] [PubMed]
- Datta, J.; Palmer, M.J.; Tanton, C.; Gibson, L.J.; Jones, K.G.; Macdowall, W.; Glasier, A.; Sonnenberg, P.; Field, N.; Mercer, C.H.; et al. Prevalence of infertility and help seeking among 15000 women and men. Hum. Reprod. 2016, 31, 2108–2118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Polis, C.B.; Cox, C.M.; Tunçalp, Ö.; McLain, A.C.; Thoma, M.E. Estimating infertility prevalence in low-to-middle-income countries: An application of a current duration approach to Demographic and Health Survey data. Hum. Reprod. 2017, 32, 1064–1074. [Google Scholar] [CrossRef] [PubMed]
- Mascarenhas, M.N.; Flaxman, S.R.; Boerma, T.; Vanderpoel, S.; Stevens, G.A. National, Regional, and Global Trends in Infertility Prevalence Since 1990: A Systematic Analysis of 277 Health Surveys. PLoS Med. 2012, 9, e1001356. [Google Scholar] [CrossRef]
- Bushnik, T.; Cook, J.L.; Yuzpe, A.A.; Tough, S.; Collins, J. Estimating the prevalence of infertility in Canada. Hum. Reprod. 2012, 27, 738–746. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, A.; Mulgund, A.; Hamada, A.; Chyatte, M.R. A unique view on male infertility around the globe. Reprod. Biol. Endocrinol. 2015, 13, 37. [Google Scholar] [CrossRef] [Green Version]
- Mehra, B.L.; Skandhan, K.P.; Prasad, B.S.; Pawankumar, G.; Singh, G.; Jaya, V. Male infertility rate: A retrospective study. Urologia 2018, 85, 22–24. [Google Scholar] [CrossRef]
- Vander Borght, M.; Wyns, C. Fertility and Infertility: Definition and Epidemiology. Clin. Biochem. 2018, 62, 2–10. [Google Scholar] [CrossRef]
- Krausz, C.; Escamilla, A.R.; Chianese, C. Genetics of male infertility: From research to clinic. Reproduction 2015, 150, R159–R174. [Google Scholar] [CrossRef]
- Sharma, P.; Ghanghas, P.; Kaushal, N.; Kaur, J.; Kaur, P. Epigenetics and oxidative stress: A twin-edged sword in spermatogenesis. Andrologia 2019, 51, e13432. [Google Scholar] [CrossRef]
- Jodar, M.; Selvaraju, S.; Sendler, E.; Diamond, M.P.; Krawetz, S.A. Reproductive Medicine Network The presence, role and clinical use of spermatozoal RNAs. Hum. Reprod. Update 2013, 19, 604–624. [Google Scholar] [CrossRef] [PubMed]
- Jensen, C.F.S.; Østergren, P.; Dupree, J.M.; Ohl, D.A.; Sønksen, J.; Fode, M. Varicocele and male infertility. Nat. Rev. Urol. 2017, 14, 523–533. [Google Scholar] [CrossRef] [PubMed]
- Soraggi, S.; Riera, M.; Rajpert-De Meyts, E.; Schierup, M.H.; Almstrup, K. Evaluating genetic causes of azoospermia: What can we learn from a complex cellular structure and single-cell transcriptomics of the human testis? Hum. Genet. 2020, 1–9. [Google Scholar] [CrossRef] [PubMed]
- Bui, A.D.; Sharma, R.; Henkel, R.; Agarwal, A. Reactive oxygen species impact on sperm DNA and its role in male infertility. Andrologia 2018, 50, e13012. [Google Scholar] [CrossRef]
- Tosic, J.; Walton, A. Metabolism of spermatozoa. The formation and elimination of hydrogen peroxide by spermatozoa and effects on motility and survival. Biochem. J. 1950, 47, 199–212. [Google Scholar] [CrossRef] [Green Version]
- Lamirande, E.; Harakat, A.; Gagnon, C. Human Sperm Capacitation Induced by Biological Fluids and Progesterone, but Not by NADH or NADPH, Is Associated with the Production of Superoxide Anion. J. Androl. 1998, 19, 215–225. [Google Scholar]
- Herrero, M.B.; de Lamirande, E.; Gagnon, C. Nitric Oxide Regulates Human Sperm Capacitation and Protein-Tyrosine Phosphorylation in Vitro. Biol. Reprod. 1999, 61, 575–581. [Google Scholar] [CrossRef]
- Barati, E.; Nikzad, H.; Karimian, M. Oxidative stress and male infertility: Current knowledge of pathophysiology and role of antioxidant therapy in disease management. Cell. Mol. Life Sci. 2020, 77, 93–113. [Google Scholar] [CrossRef]
- Griveau, J.F.; Lannou, D.L. Reactive oxygen species and human spermatozoa: Physiology and pathology. Int. J. Androl. 1997, 20, 61–69. [Google Scholar] [CrossRef]
- Camello-Almaraz, C.; Gomez-Pinilla, P.J.; Pozo, M.J.; Camello, P.J. Mitochondrial reactive oxygen species and Ca2+ signaling. Am. J. Physiol. Cell Physiol. 2006, 291, C1082–C10828. [Google Scholar] [CrossRef] [Green Version]
- Aitken, R.J.; Buckingham, D.W.; Carreras, A.; Irvine, D.S. Superoxide dismutase in human sperm suspensions: Relationship with cellular composition, oxidative stress, and sperm function. Free Radic. Biol. Med. 1996, 21, 495–504. [Google Scholar] [CrossRef]
- Said, T.M.; Agarwal, A.; Sharma, R.K.; Mascha, E.; Sikka, S.C.; Thomas, A.J. Human sperm superoxide anion generation and correlation with semen quality in patients with male infertility. Fertil. Steril. 2004, 82, 871–877. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Gibb, Z.; Baker, M.A.; Drevet, J.; Gharagozloo, P. Causes and consequences of oxidative stress in spermatozoa. Reprod. Fertil. Dev. 2016, 28, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Guerriero, G.; Trocchia, S.; Abdel-Gawad, F.K.; Ciarcia, G. Roles of Reactive Oxygen Species in the Spermatogenesis Regulation. Front. Endocrinol. 2014, 5, 56. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzyściak, W. Activity of selected aromatic amino acids in biological systems. Acta Biochim. Pol. 2011, 58, 461–466. [Google Scholar] [CrossRef] [Green Version]
- Mehta, A.; Goldstein, M. Microsurgical varicocelectomy: A review. Asian J. Androl. 2013, 15, 56–60. [Google Scholar] [CrossRef] [Green Version]
- Kroese, A.C.; de Lange, N.M.; Collins, J.; Evers, J.L. Surgery or embolization for varicoceles in subfertile men. Cochrane Database Syst. Rev. 2012. [Google Scholar] [CrossRef] [Green Version]
- Plante, M.; de Lamirande, E.; Gagnon, C. Reactive oxygen species released by activated neutrophils, but not by deficient spermatozoa, are sufficient to affect normal sperm motility. Fertil. Steril. 1994, 62, 387–393. [Google Scholar] [CrossRef]
- WHO. WHO Laboratory Manual for the Examination and Precesssing of Human Semen, 5th ed.; World Heal Organ Press: Geneva, Switzerland, 2010. [Google Scholar]
- Henkel, R. The impact of oxidants on sperm function. Andrologia 2005, 37, 205–206. [Google Scholar] [CrossRef]
- Lobascio, A.M.; De Felici, M.; Anibaldi, M.; Greco, P.; Minasi, M.G.; Greco, E. Involvement of seminal leukocytes, reactive oxygen species, and sperm mitochondrial membrane potential in the DNA damage of the human spermatozoa. Andrology 2015, 3, 265–270. [Google Scholar] [CrossRef]
- Seshadri, S.; Flanagan, B.; Vince, G.; Lewis Jones, D.I. Leucocyte subpopulations in the seminal plasma and their effects on fertilisation rates in an IVF cycle. Andrologia 2012, 44, 396–400. [Google Scholar] [CrossRef] [PubMed]
- Saleh, R.A.; Agarwal, A.; Kandirali, E.; Sharma, R.K.; Thomas, A.J.; Nada, E.A.; Evenson, D.P.; Alvarez, J.G. Leukocytospermia is associated with increased reactive oxygen species production by human spermatozoa. Fertil. Steril. 2002, 78, 1215–1224. [Google Scholar] [CrossRef]
- Akang, E.N.; Oremosu, A.A.; Osinubi, A.A.; James, A.B.; Biose, I.J.; Dike, S.I.; Idoko, K.M. Alcohol-induced male infertility: Is sperm DNA fragmentation a causative? J. Exp. Clin. Anat. 2017, 16, 53. [Google Scholar]
- Manzo-Avalos, S.; Saavedra-Molina, A. Cellular and Mitochondrial Effects of Alcohol Consumption. Int. J. Environ. Res. Public Health 2010, 7, 4281–4304. [Google Scholar] [CrossRef] [Green Version]
- Rompala, G.R.; Homanics, G.E. Intergenerational Effects of Alcohol: A Review of Paternal Preconception Ethanol Exposure Studies and Epigenetic Mechanisms in the Male Germline. Alcohol. Clin. Exp. Res. 2019, 43, 1032–1045. [Google Scholar] [CrossRef]
- Zhu, Q.; Meisinger, J.; Emanuele, N.V.; Emanuele, M.A.; LaPaglia, N.; Van Thiel, D.H. Ethanol exposure enhances apoptosis within the testes. Alcohol. Clin. Exp. Res. 2000, 24, 1550–1556. [Google Scholar] [CrossRef]
- Koh, P.O.; Kim, M.O. Ethanol Exposure Decreases Cell Proliferation and Increases Apoptosis in Rat Testes. J. Vet. Med. Sci. 2006, 68, 1013–1017. [Google Scholar] [CrossRef] [Green Version]
- Sadeghzadeh, M.; Shirpoor, A.; Naderi, R.; Kheradmand, F.; Gharalari, F.H.; Samadi, M.; Khalaji, N.; Gharaaghaji, R. Long-term ethanol consumption promotes changes in β-defensin isoform gene expression and induces structural changes and oxidative DNA damage to the epididymis of rats. Mol. Reprod. Dev. 2019, 86, 624–631. [Google Scholar] [CrossRef]
- Aitken, R.J.; Baker, M.A. Causes and consequences of apoptosis in spermatozoa; contributions to infertility and impacts on development. Int. J. Dev. Biol. 2013, 57, 265–272. [Google Scholar] [CrossRef] [Green Version]
- Sakkas, D.; Alvarez, J.G. Sperm DNA fragmentation: Mechanisms of origin, impact on reproductive outcome, and analysis. Fertil. Steril. 2010, 93, 1027–1036. [Google Scholar] [CrossRef]
- Aparicio, I.M.; Espino, J.; Bejarano, I.; Gallardo-Soler, A.; Campo, M.L.; Salido, G.M.; Pariente, J.A.; Peña, F.J.; Tapia, J.A. Autophagy-related proteins are functionally active in human spermatozoa and may be involved in the regulation of cell survival and motility. Sci. Rep. 2016, 6, 33647. [Google Scholar] [CrossRef] [PubMed]
- Zhang, M.; Jiang, M.; Bi, Y.; Zhu, H.; Zhou, Z.; Sha, J. Autophagy and Apoptosis Act as Partners to Induce Germ Cell Death After Heat Stress in Mice. PLoS ONE 2012, 7, e41412. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rehman, R.; Zahid, N.; Amjad, S.; Baig, M.; Gazzaz, Z.J. Relationship Between Smoking Habit and Sperm Parameters Among Patients Attending an Infertility Clinic. Front. Physiol. 2019, 10, 1356. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dai, J.-B.; Wang, Z.-X.; Qiao, Z.-D. The hazardous effects of tobacco smoking on male fertility. Asian J. Androl. 2015, 17, 954–960. [Google Scholar]
- Santos, M.; Rodríguez-González, G.L.; Ibáñez, C.; Vega, C.C.; Nathanielsz, P.W.; Zambrano, E. Adult exercise effects on oxidative stress and reproductive programming in male offspring of obese rats. Am. J. Physiol. Integr. Comp. Physiol. 2015, 308, R219–R225. [Google Scholar] [CrossRef] [Green Version]
- Vaamonde, D.; Da Silva-Grigoletto, M.E.; García-Manso, J.M.; Barrera, N.; Vaamonde-Lemos, R. Physically active men show better semen parameters and hormone values than sedentary men. Eur. J. Appl. Physiol. 2012, 112, 3267–3273. [Google Scholar] [CrossRef]
- Gaskins, A.J.; Mendiola, J.; Afeiche, M.; Jørgensen, N.; Swan, S.H.; Chavarro, J.E. Physical activity and television watching in relation to semen quality in young men. Br. J. Sports Med. 2015, 49, 265–270. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg, M.L.; Kim, S.; Chen, Z.; Sundaram, R.; Schisterman, E.F.; Louis, G.M.B. The relationship between male BMI and waist circumference on semen quality: Data from the LIFE study. Hum. Reprod. 2015, 30, 493–494. [Google Scholar] [CrossRef] [Green Version]
- Hajizadeh Maleki, B.; Tartibian, B.; Chehrazi, M. The effects of three different exercise modalities on markers of male reproduction in healthy subjects: A randomized controlled trial. Reproduction 2017, 153, 157–174. [Google Scholar] [CrossRef] [Green Version]
- Mastaloudis, A.; Leonard, S.W.; Traber, M.G. Oxidative stress in athletes during extreme endurance exercise. Free Radic. Biol. Med. 2001, 31, 911–922. [Google Scholar] [CrossRef]
- Durairajanayagam, D.; Agarwal, A.; Ong, C. Causes, effects and molecular mechanisms of testicular heat stress. Reprod. Biomed. Online 2015, 30, 14–27. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Parazzini, F.; Marchini, M.; Luchini, L.; Tozzi, L.; Mezzopane, R.; Fedele, L. Tight underpants and trousers and risk of dyspermia. Int. J. Androl. 1995, 18, 137–140. [Google Scholar] [CrossRef] [PubMed]
- Kesari, K.K.; Agarwal, A.; Henkel, R. Radiations and male fertility. Reprod. Biol. Endocrinol. 2018, 16, 118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kumar, S.; Nirala, J.P.; Behari, J.; Paulraj, R. Effect of electromagnetic irradiation produced by 3G mobile phone on male rat reproductive system in a simulated scenario. Indian J. Exp. Biol. 2014, 52, 890–897. [Google Scholar] [PubMed]
- Agarwal, A.; Desai, N.R.; Makker, K.; Varghese, A.; Mouradi, R.; Sabanegh, E.; Sharma, R. Effects of radiofrequency electromagnetic waves (RF-EMW) from cellular phones on human ejaculated semen: An in vitro pilot study. Fertil. Steril. 2009, 92, 1318–1325. [Google Scholar] [CrossRef] [PubMed]
- Mailankot, M.; Kunnath, A.P.; Jayalekshmi, H.; Koduru, B.; Valsalan, R. Radio frequency electromagnetic radiation (RF-EMR) from GSM (0.9/1.8GHz) mobile phones induces oxidative stress and reduces sperm motility in rats. Clinics 2009, 64, 561–565. [Google Scholar] [CrossRef] [Green Version]
- Meena, R.; Kumari, K.; Kumar, J.; Rajamani, P.; Verma, H.N.; Kesari, K.K. Therapeutic approaches of melatonin in microwave radiations-induced oxidative stress-mediated toxicity on male fertility pattern of Wistar rats. Electromagn. Biol. Med. 2014, 33, 81–91. [Google Scholar] [CrossRef]
- Avendano, C.; Mata, A.; Sarmiento, C.A.S.; Doncel, G.F. Use of Laptop Computers Connected to Internet Through Wi-Fi Decreases Human Sperm Motility and Increases Sperm DNA Fragmentation. Fertil. Steril. 2012, 97, 39–44. [Google Scholar] [CrossRef]
- Kumar, S.; Behari, J.; Sisodia, R. Influence of electromagnetic fields on reproductive system of male rats. Int. J. Radiat. Biol. 2013, 89, 147–154. [Google Scholar] [CrossRef]
- Loseva, O.; Shubbar, E.; Haghdoost, S.; Evers, B.; Helleday, T.; Harms-Ringdahl, M. Chronic Low Dose Rate Ionizing Radiation Exposure Induces Premature Senescence in Human Fibroblasts that Correlates with up Regulation of Proteins Involved in Protection against Oxidative Stress. Proteomes 2014, 2, 341–362. [Google Scholar] [CrossRef] [Green Version]
- Einor, D.; Bonisoli-Alquati, A.; Costantini, D.; Mousseau, T.A.; Møller, A.P. Ionizing radiation, antioxidant response and oxidative damage: A meta-analysis. Sci. Total Environ. 2016, 548–549, 463–471. [Google Scholar] [CrossRef] [PubMed]
- De Rosa, M.; Zarrilli, S.; Paesano, L.; Carbone, U.; Boggia, B.; Petretta, M.; Maisto, A.; Cimmino, F.; Puca, G.; Colao, A.; et al. Traffic pollutants affect fertility in men. Hum. Reprod. 2003, 18, 1055–1061. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carré, J.; Gatimel, N.; Moreau, J.; Parinaud, J.; Léandri, R. Does air pollution play a role in infertility? A systematic review. Environ. Health 2017, 16, 82. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rubes, J.; Selevan, S.G.; Evenson, D.P.; Zudova, D.; Vozdova, M.; Zudova, Z.; Robbins, W.A.; Perreault, S.D. Episodic air pollution is associated with increased DNA fragmentation in human sperm without other changes in semen quality. Hum. Reprod. 2005, 20, 2776–2783. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rengaraj, D.; Kwon, W.-S.; Pang, M.-G. Effects of Motor Vehicle Exhaust on Male Reproductive Function and Associated Proteins. J. Proteome Res. 2015, 14, 22–37. [Google Scholar] [CrossRef]
- Bahrami, N.; Goudarzi, M.; Hosseinzadeh, A.; Sabbagh, S.; Reiter, R.J.; Mehrzadi, S. Evaluating the protective effects of melatonin on di(2-ethylhexyl) phthalate-induced testicular injury in adult mice. Biomed. Pharmacother. 2018, 108, 515–523. [Google Scholar] [CrossRef]
- Al-Saleh, I.; Coskun, S.; Al-Doush, I.; Al-Rajudi, T.; Al-Rouqi, R.; Abduljabbar, M.; Al-Hassan, S. Exposure to phthalates in couples undergoing in vitro fertilization treatment and its association with oxidative stress and DNA damage. Environ. Res. 2019, 169, 396–408. [Google Scholar] [CrossRef]
- Lafuente, R.; García-Blàquez, N.; Jacquemin, B.; Checa, M.A. Outdoor Air Pollution and Sperm Quality. Fertil. Steril. 2016, 106, 880–896. [Google Scholar] [CrossRef] [Green Version]
- Drevet, J.R.; Aitken, R.J. Oxidative damage to sperm DNA: Attack and defense. In Advances in Experimental Medicine and Biology; Springer: Cham, Switzerland, 2019; Volume 1166, pp. 107–117. [Google Scholar]
- Aitken, R.J.; Smith, T.B.; Jobling, M.S.; Baker, M.A.; De Iuliis, G.N. Oxidative stress and male reproductive health. Asian J. Androl. 2014, 16, 31–38. [Google Scholar] [CrossRef]
- Agarwal, A.; Makker, K.; Sharma, R. Clinical relevance of oxidative stress in male factor infertility: An update. Am. J. Reprod. Immunol. 2008, 59, 2–11. [Google Scholar] [CrossRef]
- Lenzi, A.; Gandini, L.; Picardo, M.; Tramer, F.; Sandri, G.; Panfili, E. Lipoperoxidation damage of spermatozoa polyunsaturated fatty acids (PUFA): Scavenger mechanisms and possible scavenger therapies. Front. Biosci. 2000, 5, E1–E15. [Google Scholar] [PubMed] [Green Version]
- O’Flaherty, C.; Matsushita-Fournier, D. Reactive oxygen species and protein modifications in spermatozoa. Biol. Reprod. 2017, 97, 577–585. [Google Scholar] [CrossRef] [PubMed]
- Villani, P.; Eleuteri, P.; Grollino, M.G.; Rescia, M.; Altavista, P.; Spanò, M.; Pacchierotti, F.; Cordelli, E. Sperm DNA fragmentation induced by DNAse I and hydrogen peroxide: An in vitro comparative study among different mammalian species. Reproduction 2010, 140, 445–452. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ribas-Maynou, J.; Benet, J. Single and double strand sperm DNA damage: Different reproductive effects on male fertility. Genes 2019, 10, 105. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dorostghoal, M.; Kazeminejad, S.R.; Shahbazian, N.; Pourmehdi, M.; Jabbari, A. Oxidative stress status and sperm DNA fragmentation in fertile and infertile men. Andrologia 2017, 49, e12762. [Google Scholar] [CrossRef]
- Ammar, O.; Haouas, Z.; Hamouda, B.; Hamdi, H.; Hellara, I.; Jlali, A.; Cheikh, H.B.; Mehdi, M. Relationship between sperm DNA damage with sperm parameters, oxidative markers in teratozoospermic men. Eur. J. Obstet. Gynecol. Reprod. Biol. 2019, 233, 70–75. [Google Scholar] [CrossRef]
- Gawecka, J.E.; Marh, J.; Ortega, M.; Yamauchi, Y.; Ward, M.A.; Ward, W.S. Mouse zygotes respond to severe sperm DNA damage by delaying paternal DNA replication and embryonic development. PLoS ONE 2013, 8, e56385. [Google Scholar] [CrossRef] [Green Version]
- Toyoshima, M.; Shimura, T.; Adiga, S.-K.; Taga, M.; Shiraishi, K.; Inoue, M.; Yuan, Z.-M.; Niwa, O. Transcription-independent suppression of DNA synthesis by p53 in sperm-irradiated mouse zygotes. Oncogene 2005, 24, 3229–3235. [Google Scholar] [CrossRef] [Green Version]
- Lewis, S.E.M.; Aitken, R.J. DNA damage to spermatozoa has impacts on fertilization and pregnancy. Cell Tissue Res. 2005, 322, 33–41. [Google Scholar] [CrossRef]
- Simon, L.; Lewis, S.E.M. Sperm DNA damage or progressive motility: Which one is the better predictor of fertilization in vitro? Syst. Biol. Reprod. Med. 2011, 57, 133–138. [Google Scholar] [CrossRef]
- Barbato, V.; Talevi, R.; Braun, S.; Merolla, A.; Sudhakaran, S.; Longobardi, S.; Gualtieri, R. Supplementation of sperm media with zinc, D-aspartate and co-enzyme Q10 protects bull sperm against exogenous oxidative stress and improves their ability to support embryo development. Zygote 2017, 25, 168–175. [Google Scholar] [CrossRef] [PubMed]
- Zini, A. Are sperm chromatin and DNA defects relevant in the clinic? Syst. Biol. Reprod. Med. 2011, 57, 78–85. [Google Scholar] [CrossRef] [PubMed]
- Chen, Q.; Zhao, J.-Y.; Xue, X.; Zhu, G.-X. The association between sperm DNA fragmentation and reproductive outcomes following intrauterine insemination, a meta analysis. Reprod. Toxicol. 2019, 86, 50–55. [Google Scholar] [CrossRef] [PubMed]
- Sugihara, A.; Van Avermaete, F.; Roelant, E.; Punjabi, U.; De Neubourg, D. The role of sperm DNA fragmentation testing in predicting intra-uterine insemination outcome: A systematic review and meta-analysis. Eur. J. Obstet. Gynecol. Reprod. Biol. 2020, 244, 8–15. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Deng, C.; Li, T.; Xie, Y.; Guo, Y.; Yang, Q.; Liang, X.; Deng, C.; Liu, G. Sperm DNA fragmentation index influences assisted reproductive technology outcome: A systematic review and meta-analysis combined with a retrospective cohort study. Andrologia 2019, 51, e13263. [Google Scholar] [CrossRef] [PubMed]
- Simon, L.; Zini, A.; Dyachenko, A.; Ciampi, A.; Carrell, D.T. A systematic review and meta-analysis to determine the effect of sperm DNA damage on in vitro fertilization and intracytoplasmic sperm injection outcome. Asian J. Androl. 2017, 19, 80–90. [Google Scholar]
- Zhao, J.; Zhang, Q.; Wang, Y.; Li, Y. Whether sperm deoxyribonucleic acid fragmentation has an effect on pregnancy and miscarriage after in vitro fertilization/intracytoplasmic sperm injection: A systematic review and meta-analysis. Fertil. Steril. 2014, 102, 998–1005.e8. [Google Scholar] [CrossRef]
- Osman, A.; Alsomait, H.; Seshadri, S.; El-Toukhy, T.; Khalaf, Y. The effect of sperm DNA fragmentation on live birth rate after IVF or ICSI: A systematic review and meta-analysis. Reprod. Biomed. Online 2015, 30, 120–127. [Google Scholar] [CrossRef] [Green Version]
- Cissen, M.; Van Wely, M.; Scholten, I.; Mansell, S.; De Bruin, J.P.; Mol, B.W.; Braat, D.; Repping, S.; Hamer, G. Measuring Sperm DNA Fragmentation and Clinical Outcomes of Medically Assisted Reproduction: A Systematic Review and Meta-Analysis. PLoS ONE 2016, 11, e0165125. [Google Scholar] [CrossRef] [Green Version]
- Halpern, J.A.; Schlegel, P.N. Should A Couple with Failed In Vitro Fertilization/Intracytoplasmic Sperm Injection and Increased Sperm DNA Fragmentation Use Testicular Sperm for the Next Cycle? Eur. Urol. Focus 2018, 4, 299–300. [Google Scholar] [CrossRef]
- Zhang, Z.; Zhu, L.; Jiang, H.; Chen, H.; Chen, Y.; Dai, Y. Sperm DNA fragmentation index and pregnancy outcome after IVF or ICSI: A meta-analysis. J. Assist. Reprod. Genet. 2015, 32, 17–26. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ward, W.S. Function of sperm chromatin structural elements in fertilization and development. Mol. Hum. Reprod. 2010, 16, 30–36. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshida, K.; Muratani, M.; Araki, H.; Miura, F.; Suzuki, T.; Dohmae, N.; Katou, Y.; Shirahige, K.; Ito, T.; Ishii, S. Mapping of histone-binding sites in histone replacement-completed spermatozoa. Nat. Commun. 2018, 9, 3885. [Google Scholar] [CrossRef] [PubMed]
- Yamaguchi, K.; Hada, M.; Fukuda, Y.; Inoue, E.; Makino, Y.; Katou, Y.; Shirahige, K.; Okada, Y. Re-evaluating the Localization of Sperm-Retained Histones Revealed the Modification-Dependent Accumulation in Specific Genome Regions. Cell Rep. 2018, 23, 3920–3932. [Google Scholar] [CrossRef] [PubMed]
- Mudrak, O.; Tomilin, N.; Zalensky, A. Chromosome architecture in the decondensing human sperm nucleus. J. Cell Sci. 2005, 118, 4541–4550. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- González-Rojo, S.; Fernández-Díez, C.; Lombó, M.; Herráez, M.P. Distribution of DNA damage in the sperm nucleus: A study of zebrafish as a model of histone-packaged chromatin. Theriogenology 2018, 122, 109–115. [Google Scholar] [CrossRef] [PubMed]
- Aitken, R.J.; Jones, K.T.; Robertson, S.A. Reactive oxygen species and sperm function–in sickness and in health. J. Androl. 2012, 33, 1096–1106. [Google Scholar] [CrossRef]
- Rhemrev, J.P.T.; Van Overveld, F.W.; Haenen, G.R.; Teerlink, T.; Bast, A.; Vermeiden, J.P. Quantification of the Nonenzymatic Fast and Slow TRAP in a Postaddition Assay in Human Seminal Plasma and the Antioxidant Contributions of Various Seminal Compounds. J. Androl. 2000, 21, 913–920. [Google Scholar]
- Lazzarino, G.; Listorti, I.; Bilotta, G.; Capozzolo, T.; Amorini, A.M.; Longo, S.; Caruso, G.; Lazzarino, G.; Tavazzi, B.; Bilotta, P. Water- and Fat-Soluble Antioxidants in Human Seminal Plasma and Serum of Fertile Males. Antioxidants 2019, 8, 96. [Google Scholar] [CrossRef] [Green Version]
- O’Flaherty, C. Peroxiredoxins: Hidden players in the antioxidant defence of human spermatozoa. Basic Clin. Androl. 2014, 24, 4. [Google Scholar] [CrossRef] [Green Version]
- Abdelbaki, S.A.; Sabry, J.H.; Al-Adl, A.M.; Sabry, H.H. The impact of coexisting sperm DNA fragmentation and seminal oxidative stress on the outcome of varicocelectomy in infertile patients: A prospective controlled study. Arab J. Urol. 2017, 15, 131–139. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Baazeem, A.; Belzile, E.; Ciampi, A.; Dohle, G.; Jarvi, K.; Salonia, A.; Weidner, W.; Zini, A. Varicocele and Male Factor Infertility Treatment: A New Meta-Analysis and Review of the Role of Varicocele Repair. Eur. Urol. 2011, 60, 796–808. [Google Scholar] [CrossRef] [PubMed]
- Lara-Cerrillo, S.; Gual-Frau, J.; Benet, J.; Abad, C.; Prats, J.; Amengual, M.J.; Ribas-Maynou, J.; García-Peiró, A. Microsurgical varicocelectomy effect on sperm telomere length, DNA fragmentation and seminal parameters. Hum. Fertil. 2020, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Lu, X.-L.; Liu, J.-J.; Li, J.-T.; Yang, Q.-A.; Zhang, J.-M. Melatonin therapy adds extra benefit to varicecelectomy in terms of sperm parameters, hormonal profile and total antioxidant capacity: A placebo-controlled, double-blind trial. Andrologia 2018, 50, e13033. [Google Scholar] [CrossRef] [PubMed]
- Smits, R.M.; Mackenzie-Proctor, R.; Yazdani, A.; Stankiewicz, M.T.; Jordan, V.; Showell, M.G. Antioxidants for male subfertility. Cochrane Database Syst. Rev. 2019, 3, CD007411. [Google Scholar] [CrossRef]
- Gharagozloo, P.; Aitken, R.J. The role of sperm oxidative stress in male infertility and the significance of oral antioxidant therapy. Hum. Reprod. 2011, 26, 1628–1640. [Google Scholar] [CrossRef] [Green Version]
- Ardestani Zadeh, A.; Arab, D.; Kia, N.S.; Heshmati, S.; Amirkhalili, S.N. The role of Vitamin E-Selenium-Folic Acid Supplementation in Improving Sperm Parameters After Varicocelectomy: A Randomized Clinical Trial. Urol. J. 2019, 16, 495–500. [Google Scholar]
- Nouri, M.; Amani, R.; Nasr-Esfahani, M.; Tarrahi, M.J. The effects of lycopene supplement on the spermatogram and seminal oxidative stress in infertile men: A randomized, double-blind, placebo-controlled clinical trial. Phytother. Res. 2019, 33, 3203–3211. [Google Scholar] [CrossRef]
- Banday, M.N.; Lone, F.A.; Rasool, F.; Rashid, M.; Shikari, A. Use of antioxidants reduce lipid peroxidation and improve quality of crossbred ram sperm during its cryopreservation. Cryobiology 2017, 74, 25–30. [Google Scholar] [CrossRef]
- Gual-Frau, J.; Abad, C.; Amengual, M.J.; Hannaoui, N.; Checa, M.A.; Ribas-Maynou, J.; Lozano, I.; Nikolaou, A.; Benet, J.; García-Peiró, A.; et al. Oral antioxidant treatment partly improves integrity of human sperm DNA in infertile grade i varicocele patients. Hum. Fertil. 2015, 18, 225–229. [Google Scholar] [CrossRef]
- Negri, L.; Benaglia, R.; Monti, E.; Morenghi, E.; Pizzocaro, A.; Levi Setti, P.E. Effect of superoxide dismutase supplementation on sperm DNA fragmentation. Arch. Ital. Urol. Androl. 2017, 89, 212–218. [Google Scholar] [CrossRef] [PubMed]
- Martínez-Soto, J.C.; Domingo, J.C.; Cordobilla, B.; Nicolás, M.; Fernández, L.; Albero, P.; Gadea, J.; Landeras, J. Dietary supplementation with docosahexaenoic acid (DHA) improves seminal antioxidant status and decreases sperm DNA fragmentation. Syst. Biol. Reprod. Med. 2016, 62, 387–395. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, J.; Barranco, I.; Tvarijonaviciute, A.; Molina, M.F.; Martinez, E.A.; Rodriguez-Martinez, H.; Parrilla, I.; Roca, J. Seminal plasma antioxidants are directly involved in boar sperm cryotolerance. Theriogenology 2018, 107, 27–35. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Amidi, F.; Pazhohan, A.; Shabani Nashtaei, M.; Khodarahmian, M.; Nekoonam, S. The role of antioxidants in sperm freezing: A review. Cell Tissue Bank. 2016, 17, 745–756. [Google Scholar] [CrossRef]
- Martínez-Soto, J.C.; Landeras, J.; Gadea, J. Spermatozoa and seminal plasma fatty acids as predictors of cryopreservation success. Andrology 2013, 1, 365–375. [Google Scholar] [CrossRef] [Green Version]
- Attallah, D.; El-Nashar, I.H.; Mahmoud, R.; Shaaban, O.M.; Salman, S.A. N-acytelcysteine prior to intrauterine insemination in couples with isolated athenozospermia: A randomized controlled trial. Fertil. Steril. 2013, 100, S462. [Google Scholar] [CrossRef]
- Comhaire, F.H.; Kunnen, M. Factors affecting the probability of conception after treatment of subfertile men with varicocele by transcatheter embolization with Bucrylate. Fertil. Steril. 1985, 43, 781–786. [Google Scholar] [CrossRef]
- Steiner, A.Z.; Hansen, K.R.; Barnhart, K.T.; Cedars, M.I.; Legro, R.S.; Diamond, M.P.; Krawetz, S.A.; Usadi, R.; Baker, V.L.; Coward, R.M.; et al. The effect of antioxidants on male factor infertility: The Males, Antioxidants, and Infertility (MOXI) randomized clinical trial. Fertil. Steril. 2020, 113, 552–560.e3. [Google Scholar] [CrossRef]
- Tremellen, K.; Miari, G.; Froiland, D.; Thompson, J. A randomised control trial examining the effect of an antioxidant (Menevit) on pregnancy outcome during IVF-ICSI treatment. Aust. N. Z. J. Obstet. Gynaecol. 2007, 47, 216–221. [Google Scholar] [CrossRef]
- Geva, E.; Bartoov, B.; Zabludovsky, N.; Lessing, J.B.; Lerner-Geva, L.; Amit, A. The effect of antioxidant treatment on human spermatozoa and fertilization rate in an in vitro fertilization program. Fertil. Steril. 1996, 66, 430–434. [Google Scholar] [CrossRef]
- Greco, E.; Romano, S.; Iacobelli, M.; Ferrero, S.; Baroni, E.; Minasi, M.G.; Ubaldi, F.; Rienzi, L.; Tesarik, J. ICSI in cases of sperm DNA damage: Beneficial effect of oral antioxidant treatment. Hum. Reprod. 2005, 20, 2590–2594. [Google Scholar] [CrossRef] [PubMed] [Green Version]
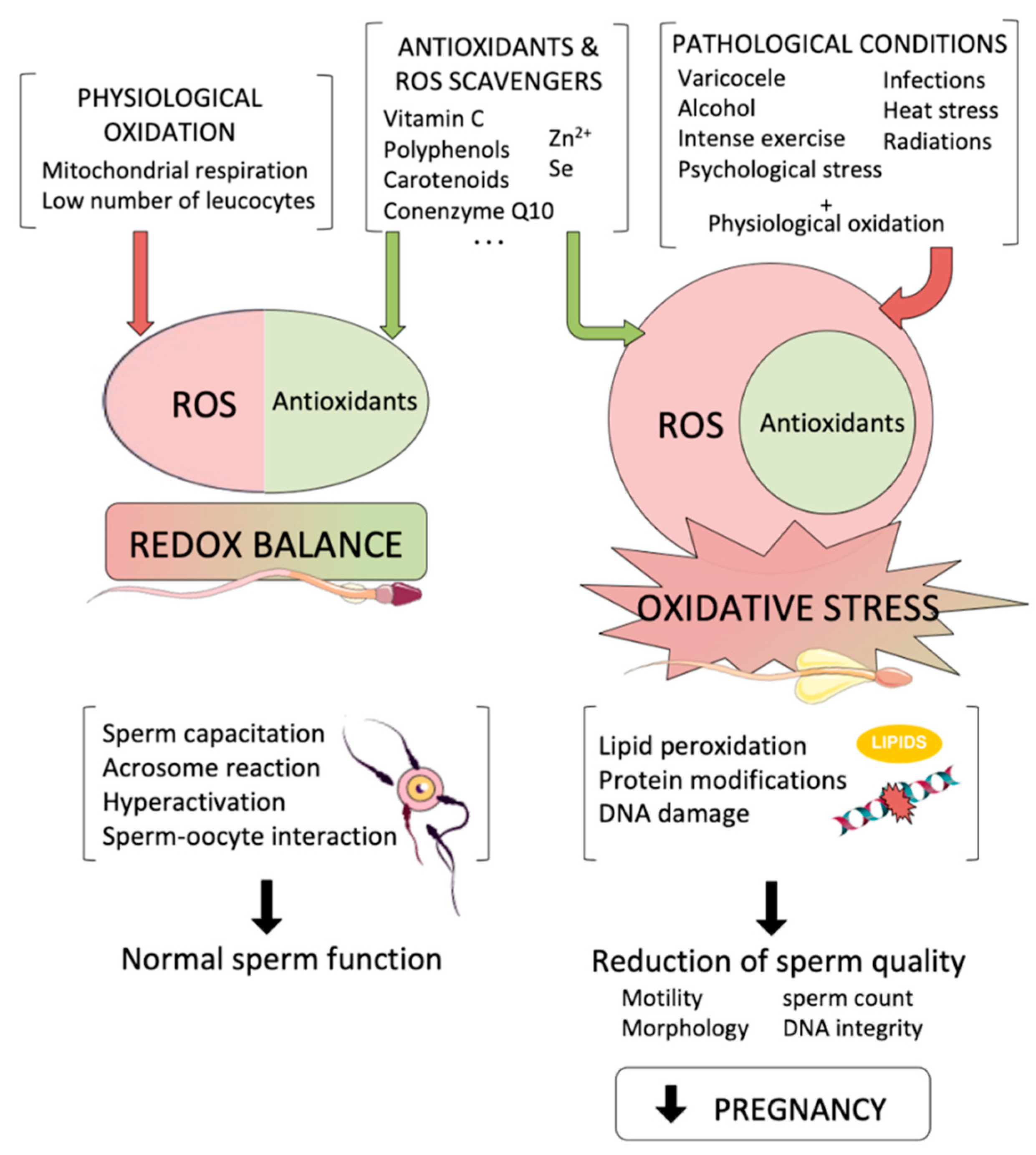
Figure 1.
Reactive oxygen species (ROS) are by-products that are necessary for essential sperm functions. On the left, a redox balance is achieved when physiological oxidation is compensated by antioxidants, so that physiological oxidation allows sperm to perform their normal functions. However, when pathological conditions lead to an increase in intracellular ROS levels (right), oxidative stress affects sperm cells causing a reduction in sperm quality that leads to a reduction in pregnancy achievement.
Figure 1.
Reactive oxygen species (ROS) are by-products that are necessary for essential sperm functions. On the left, a redox balance is achieved when physiological oxidation is compensated by antioxidants, so that physiological oxidation allows sperm to perform their normal functions. However, when pathological conditions lead to an increase in intracellular ROS levels (right), oxidative stress affects sperm cells causing a reduction in sperm quality that leads to a reduction in pregnancy achievement.

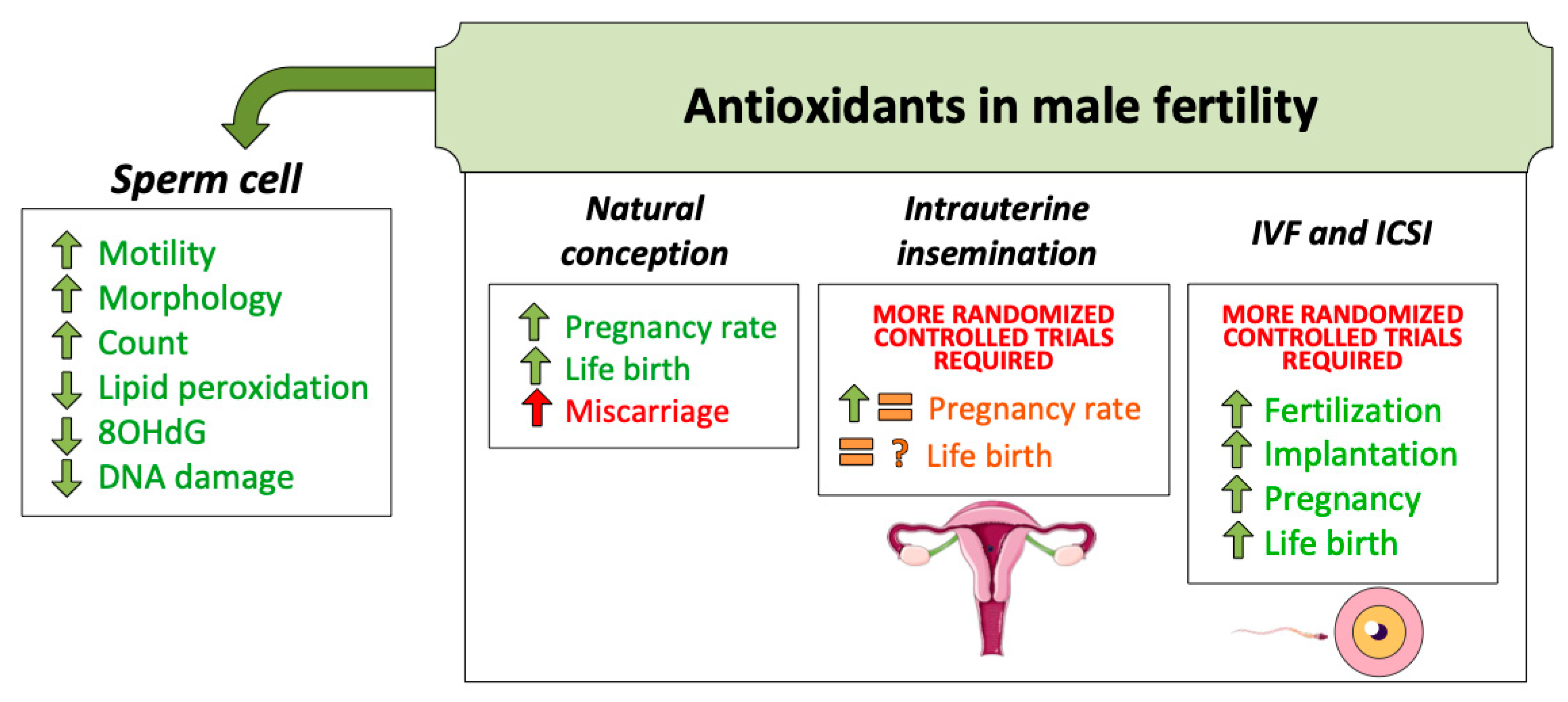
Figure 2.
Antioxidant treatments for male fertility have been used for decades. Until the present moment, evidence supports its use to achieve better sperm quality. This, in turn, increases natural conception success. Regarding assisted reproduction techniques (ART), more randomized and controlled trials are necessary to confirm the benefit of oral antioxidant intake.
Figure 2.
Antioxidant treatments for male fertility have been used for decades. Until the present moment, evidence supports its use to achieve better sperm quality. This, in turn, increases natural conception success. Regarding assisted reproduction techniques (ART), more randomized and controlled trials are necessary to confirm the benefit of oral antioxidant intake.

© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Ribas-Maynou, J.; Yeste, M. Oxidative Stress in Male Infertility: Causes, Effects in Assisted Reproductive Techniques, and Protective Support of Antioxidants. Biology 2020, 9, 77. https://doi.org/10.3390/biology9040077
AMA Style
Ribas-Maynou J, Yeste M. Oxidative Stress in Male Infertility: Causes, Effects in Assisted Reproductive Techniques, and Protective Support of Antioxidants. Biology. 2020; 9(4):77. https://doi.org/10.3390/biology9040077
Chicago/Turabian StyleRibas-Maynou, Jordi, and Marc Yeste. 2020. "Oxidative Stress in Male Infertility: Causes, Effects in Assisted Reproductive Techniques, and Protective Support of Antioxidants" Biology 9, no. 4: 77. https://doi.org/10.3390/biology9040077
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.