
The Regulation of Reverse Cholesterol Transport and Cellular Cholesterol Homeostasis by MicroRNAs


Abstract
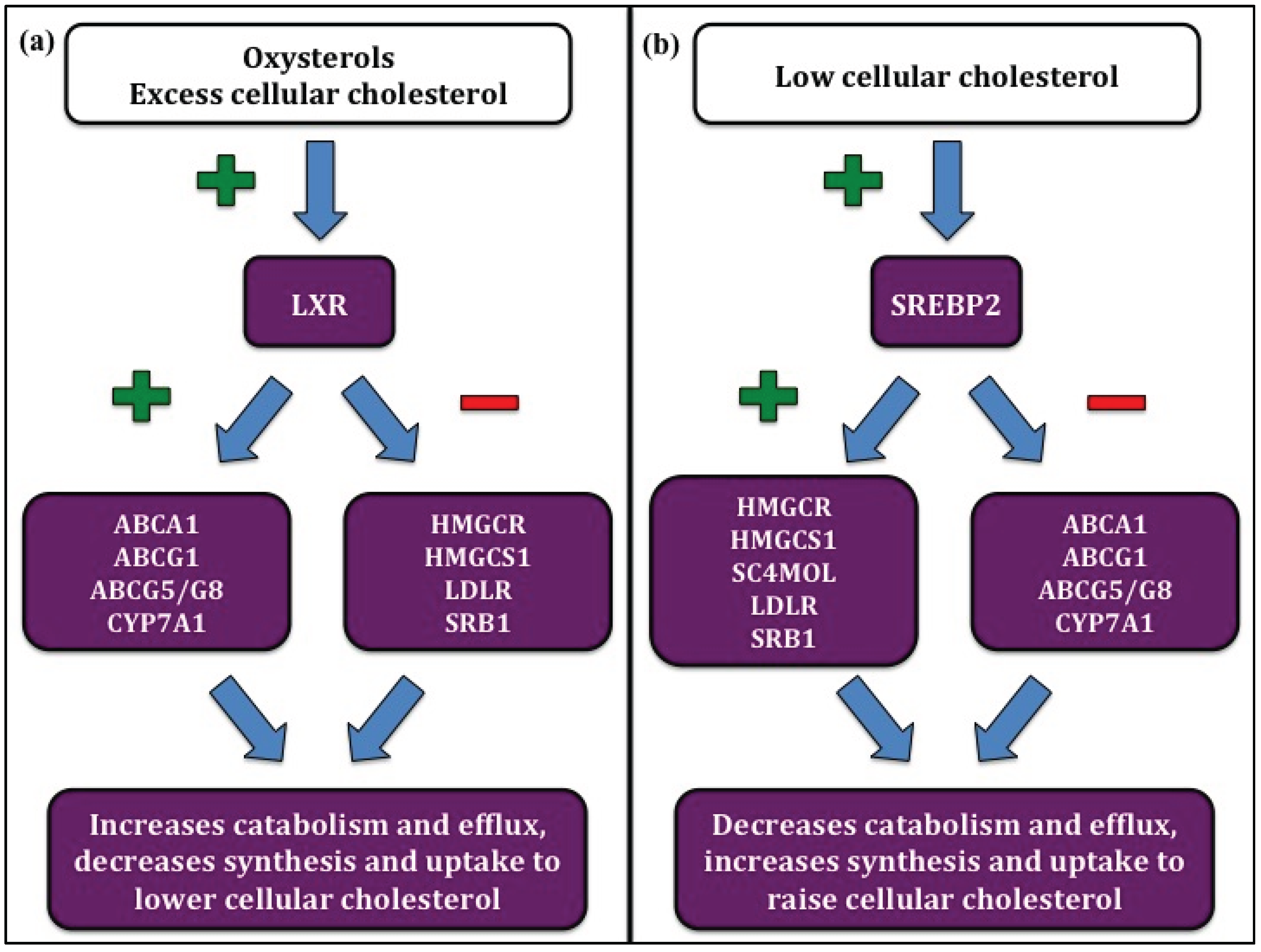
:1. Introduction

2. Regulation of Plasma Lipoproteins by miRNAs
3. Regulation of Cellular Cholesterol Homeostasis by miRNAs
4. Regulation of Cholesterol Synthesis by miRNAs
5. Regulation of Macrophage Cholesterol Uptake by miRNAs
6. Regulation of Cholesterol Esterification by miRNAs
7. Regulation of Cholesterol Efflux by miRNAs
7.1. Regulation of ABCA1 by miRNAs
7.2. Regulation of ABCG1 by miRNAs
8. Regulation of Hepatic HDL by miRNAs
9. Regulation of Cholesterol Catabolism by miRNAs

{kind=link}
| miRNA | Protein target(s) | Regulatory Action | Clinical Implications | References |
|---|---|---|---|---|
| miR-1 | LXRα* | Directly suppresses LXR in vitro | May promote an increase in cellular cholesterol | [38] |
| miR-9 | ACAT1* | Directly suppresses ACAT1 and esterification of cholesterol in macrophages | Overexpression may promote macrophage cholesterol efflux and reduce foam cell formation | [47] |
| miR-10b | ABCA1* ABCG1* | Directly represses ABCA1 and ABCG1 expression and decreases macrophage cholesterol efflux | Can be suppressed by dietary anthocyanins, leading to increased macrophage cholesterol efflux and lesion regression | [63] |
| miR-19b | ABCA1* | Directly suppresses ABCA1 and decreases cholesterol efflux to ApoA1; increases atherosclerotic lesion area and severity | Inhibition may increase macrophage ABCA1, promoting cholesterol efflux and lesion regression | [53] |
| miR-26 | ABCA1* ARL7 | Activated by LXR to suppress both proteins, decreasing macrophage cholesterol efflux | Inhibition may increase macrophage ABCA1, promoting cholesterol efflux and lesion regression | [58] |
| miR-27a/b | ABCA1* ABCG1 ACAT1* CD36 LPL* | Directly suppresses ABCA1, indirectly suppresses ABCG1, and reduces cholesterol efflux. Reduces macrophage cholesterol uptake by suppressing LPL (directly) and CD36 (indirectly), and ACAT1 (directly) | Inhibition of miR-27 would promote macrophage cholesterol efflux, but may also increase macrophage cholesterol uptake and retention | [46] |
| ANGPTL3* | Directly suppresses ANGPTL3, leading to decreased EL activity | Overexpression may provide a means of increasing plasma HDL | [28] | |
| miR-30c | MTP* ApoB | Directly suppresses MTP expression, leading to reduced ApoB secretion and formation of TRL | No hepatic steatosis, reduced TRL, fewer and smaller aortic lesions | [27] |
| miR-33a/b | ABCA1* | Directly suppresses ABCA1 mRNA and protein expression, decreases cholesterol efflux to ApoA1, and decreases HDL | miR-33 inhibition may increase cholesterol efflux and plasma HDL and promote regression of atherosclerotic lesions | [55,56,62,69,70] |
| ABCG5/G8 ATP8B1* BSEP* | Directly suppresses ATP8B1 and BSEP, indirectly suppresses ABCG5/G8 | Inhibition of miR-33 may counteract the known hepatotoxic side effects of statins | [66,67] | |
| CYP7A1* CYP8B1* NTCP* | Suppresses CYP7A1, CYP8B1, and NTCP, and decreases bile acid pool size | miR-33 inhibition may increase conversion of cholesterol to bile acids, reducing plasma TC | [66] | |
| miR-33-3p | ABCA1 ABCG1 | Indirectly suppresses expression of ABCA1 and ABCG1, decreasing cholesterol efflux | miR-33* inhibition may promote cholesterol efflux and lesion regression | [6] |
| miR-96 | SRB1* | Directly suppresses hepatic SRB1 expression but increases HDL uptake | Overexpression increases hepatic HDL uptake | [32] |
| miR-122 | ACC1* ACC2* FASN* SCD1* | Directly upregulates expression of key lipogenic genes, therefore elevating plasma cholesterol | Inhibition of miR-122 may reduce lipogenesis and subsequent release of TRL | [26] |
| miR-128 | ABCA1* ABCG1* HMGCoA-R HMGCoA-S1 LDLR SREBP | Directly suppresses ABCA1, and ABCG1, and increases SREBP2 and its downstream genes to increase cellular cholesterol | Inhibition may decrease cellular cholesterol and promote cholesterol efflux | [49] |
| miR-144-5p/miR-144-3p | ABCA1* | Directly suppresses ABCA1 expression, plasma HDL, and increases atherosclerotic burden | Inhibition of miR-144/miR-144* may increase ABCA1 and promote cholesterol efflux from lesional macrophages | [5,54] |
| miR-145 | ABCA1* | Directly suppresses ABCA1 and decreases cholesterol efflux to ApoA1 | Inhibition may increase hepatic ABCA1 and promote HDL biogenesis | [50] |
| miR-185 | SREBP2* | Directly suppresses expression of SREBP2 and its downstream genes to increase cellular cholesterol | miR-185 overexpression may decrease cholesterol biosynthesis | [35] |
| SRB1* | Negatively regulates hepatic SRB1 expression and HDL uptake | Overexpression reduces HDL uptake, which may promote development of dysfunctional HDL | [32] | |
| miR-197 | HMGCoA-R HMGCoA-S1 IDI1 | Indirectly increases cholesterol synthesis by binding to transcription factor FOXJ2 | Suppression may decrease cholesterol synthesis | [42] |
| miR-206 | LXRα* | Directly suppresses LXR in vitro | May promote an increase in cellular cholesterol | [38] |
| miR-223 | ABCA1 CYP7A1 HMGCoA-S1* SC4MOL* | Indirectly increases ABCA1 and CYP7A1, and decreases cholesterol synthesis | miR-223 agonists may promote cholesterol efflux and HDL formation and reduce cellular cholesterol | [33] |
| SRB1* | Negatively regulates hepatic SRB1 expression and HDL uptake | Overexpression reduces HDL uptake, which may promote development of dysfunctional HDL | [32,33] | |
| miR-302a | ABCA1* | Directly represses ABCA1 expression and reduces cholesterol efflux to ApoA1 | Inhibition increases hepatic and macrophage ABCA1, promoting cholesterol efflux and lesion regression | [51] |
| miR-378 | ABCG1* | Directly decreases expression and attenuates cholesterol efflux to HDL | Antagonism of miR-378 increases ABCG1 and promotes macrophage cholesterol efflux to HDL | [64] |
| miR-613 | ABCA1* | Directly suppresses ABCA1, attenuating macrophage cholesterol efflux to ApoA1 | Suppression increases macrophage cholesterol efflux and may promote lesion regression | [61] |
| LXRα* | Suppresses LXRα as part of an autofeedback loop to maintain cellular cholesterol homeostasis | May be a target for the modulation of cellular cholesterol homeostasis and LXR-target genes | [36,61] | |
| miR-758 | ABCA1* | Directly suppresses ABCA1 and attenuates cholesterol efflux to ApoA1 | Suppression increases macrophage and hepatic ABCA1 and may promote cholesterol efflux | [48] |
10. Dietary Influence on miRNA Expression
11. Conclusions
Author Contributions
Conflicts of Interest
References
- Bartel, D.P. Review microRNAs : Target recognition and regulatory functions. Cell 2009, 136, 215–233. [Google Scholar] [CrossRef] [PubMed]
- Lee, Y.; Jeon, K.; Lee, J.T.; Kim, S.; Kim, V.N. MicroRNA maturation: Stepwisee processing and subcellular localization. EMBO J. 2002, 21, 4663–4670. [Google Scholar] [CrossRef] [PubMed]
- Peters, L.; Meister, G. Argonaute proteins: Mediators of RNA silencing. Mol. Cell 2007, 26, 611–623. [Google Scholar] [CrossRef] [PubMed]
- Meister, G.; Landthaler, M.; Patkaniowska, A.; Dorsett, Y.; Teng, G.; Tuschl, T. Human Argonaute2 mediates RNA cleavage targeted by miRNAs and siRNAs. Mol. Cell 2004, 15, 185–197. [Google Scholar] [CrossRef] [PubMed]
- Hu, Y.W.; Hu, Y.R.; Zhao, J.Y.; Li, S.F.; Ma, X.; Wu, S.G.; Lu, J.B.; Qiu, Y.R.; Sha, Y.H.; Wang, Y.C.; et al. An agomir of miR-144-3p accelerates plaque formation through impairing reverse cholesterol transport and promoting pro-inflammatory cytokine production. PLoS ONE 2014, 9, e94997. [Google Scholar] [CrossRef] [PubMed]
- Goedeke, L.; Vales-Lara, F.M.; Fenstermaker, M.; Cirera-Salinas, D.; Chamorro-Jorganes, A.; Ramírez, C.M.; Mattison, J.A.; de Cabo, R.; Suárez, Y.; Fernández-Hernando, C. A regulatory role for microRNA 33* in controlling lipid metabolism gene expression. Mol. Cell. Biol. 2013, 33, 2339–2352. [Google Scholar] [CrossRef] [PubMed]
- Sayed, D.; Abdellatif, M. MicroRNAs in development and disease. Physiol. Rev. 2011, 91, 827–887. [Google Scholar] [CrossRef] [PubMed]
- Jansson, M.D.; Lund, A.H. MicroRNA and cancer. Mol. Oncol. 2012, 6, 590–610. [Google Scholar] [CrossRef] [PubMed]
- Novák, J.; Bienertová-Vašků, J.; Kára, T.; Novák, M. MicroRNAs involved in the lipid metabolism and their possible implications for atherosclerosis development and treatment. Mediators Inflamm. 2014. [Google Scholar] [CrossRef] [PubMed]
- Friedman, R.C.; Farh, K.K.H.; Burge, C.B.; Bartel, D.P. Most mammalian mRNAs are conserved targets of microRNAs. Genome Res. 2009, 19, 92–105. [Google Scholar] [CrossRef] [PubMed]
- Grundy, S.M.; Cleeman, J.I.; Bairey Merz, C.N.; Brewer, H.B.; Clark, L.T.; Hunninghake, D.B.; Pasternak, R.C.; Smith, S.C.; Stone, N.J. Implications of recent clinical trials for the National Cholesterol Education Program Adult Treatment Panel III guidelines. Circulation 2004, 110, 227–239. [Google Scholar] [CrossRef] [PubMed]
- Go, A.S.; Mozaffarian, D.; Roger, V.L.; Benjamin, E.J.; Berry, J.D.; Blaha, M.J.; Dai, S.; Ford, E.S.; Fox, C.S.; Franco, S.; et al. Heart disease and stroke statistics-2014 update: A report from the American Heart Association. Circulation 2014, 129, e28–e292. [Google Scholar] [CrossRef] [PubMed]
- Wetterau, J.R.; Lin, M.C.; Jamil, H. Microsomal triglyceride transfer protein. Biochim. Biophys. Acta 1997, 1345, 136–150. [Google Scholar] [CrossRef]
- Nilsson-Ehle, P.; Garfinkel, A.S.; Schotz, M.C. Lipolytic enzymes and plasma lipoprotein metabolism. Annu. Rev. Biochem. 1980, 49, 667–693. [Google Scholar] [CrossRef] [PubMed]
- Havel, R.J. The formation of LDL: Mechanisms and regulation. J. Lipid Res. 1984, 25, 1570–1576. [Google Scholar] [PubMed]
- Pentikäinen, M.O.; Öörni, K.; Ala-Korpela, M.; Kovanen, P.T. Modified LDL-trigger of atherosclerosis and inflammation in the arterial intima. J. Intern. Med. 2000, 247, 359–370. [Google Scholar] [CrossRef] [PubMed]
- Annema, W.; Tietge, U.J. Regulation of reverse cholesterol transport—A comprehensive appraisal of available animal studies. Nutr. Metab. 2012. [Google Scholar] [CrossRef] [PubMed]
- Hill, S.A.; McQueen, M.J. Reverse cholesterol transport—A review of the process and its clinical implications. Clin. Biochem. 1997, 30, 517–525. [Google Scholar] [CrossRef]
- Shimano, H. Sterol regulatory element-binding proteins (SREBPs): Transcriptional regulators of lipid synthetic genes. Prog. Lipid Res. 2001, 40, 439–452. [Google Scholar] [CrossRef]
- Horton, J.D.; Goldstein, J.L.; Brown, M.S. SREBPs: Activators of the complete program of cholesterol and fatty acid synthesis in the liver. J. Clin. Invest. 2002, 109, 1125–1131. [Google Scholar] [CrossRef] [PubMed]
- Maxwell, K.N.; Soccio, R.E.; Duncan, E.M.; Sehayek, E.; Breslow, J.L. Novel putative SREBP and LXR target genes identified by microarray analysis in liver of cholesterol-fed mice. J. Lipid Res. 2003, 44, 2109–2119. [Google Scholar] [CrossRef] [PubMed]
- Grefhorst, A.; Oosterveer, M.H.; Brufau, G.; Boesjes, M.; Kuipers, F.; Groen, A.K. Pharmacological LXR activation reduces presence of SR-B1 in liver membranes contributing to LXR-mediated induction of HDL-cholesterol. Atherosclerosis 2012, 222, 382–389. [Google Scholar] [CrossRef] [PubMed]
- Birjmohun, R.S.; Hutten, B.A.; Kastelein, J.J.P.; Stroes, E.S.G. Increasing HDL cholesterol with extended- release nicotinic acid: From promise to practice. Evaluation 2007, 62, 2005–2007. [Google Scholar]
- Taylor, B.A.; Thompson, P.D. Muscle-related side-effects of statins. Curr. Opin. Lipidol. 2015, 26, 221–227. [Google Scholar] [CrossRef] [PubMed]
- Chang, J.; Nicolas, E.; Marks, D.; Sander, C.; Lerro, A.; Buendia, M.A.; Xu, C.; Mason, W.S.; Moloshok, T.; Bort, R.; et al. MiR-122, a mammalian liver-specific microRNA, is processed from hcr mRNA and may downregulate the high affinity cationic amino acid transporter CAT-1. RNA Biol. 2004, 1, 106–113. [Google Scholar] [CrossRef] [PubMed]
- Esau, C.; Davis, S.; Murray, S.F.; Yu, X.X.; Pandey, S.K.; Pear, M.; Watts, L.; Booten, S.L.; Graham, M.; McKay, R.; et al. MiR-122 regulation of lipid metabolism revealed by in vivo antisense targeting. Cell Metab. 2006, 3, 87–98. [Google Scholar] [CrossRef] [PubMed]
- Soh, J.; Iqbal, J.; Queiroz, J.; Fernandez-Hernando, C.; Hussain, M.M. MicroRNA-30c reduces hyperlipidemia and atherosclerosis in mice by decreasing lipid synthesis and lipoprotein secretion. Nat. Med. 2013, 19, 892–900. [Google Scholar] [CrossRef] [PubMed]
- Vickers, K.C.; Shoucri, B.M.; Levin, M.G.; Wu, H.; Pearson, D.S.; Osei-Hwedieh, D.; Collins, F.S.; Remaley, A.T.; Sethupathy, P. MicroRNA-27b is a regulatory hub in lipid metabolism and is altered in dyslipidemia. Hepatology 2013, 57, 533–542. [Google Scholar] [CrossRef] [PubMed]
- Jaye, M.; Krawiec, J. Endothelial lipase and HDL metabolism. Curr. Opin. Lipidol. 2004, 15, 183–189. [Google Scholar] [CrossRef] [PubMed]
- Vickers, K.C.; Palmisano, B.T.; Shoucri, B.M.; Shamburek, R.D.; Remaley, A.T. MicroRNAs are transported in plasma and delivered to recipient cells by high-density lipoproteins. Nat. Cell Biol. 2011, 13, 423–433. [Google Scholar] [CrossRef] [PubMed]
- Wagner, J.; Riwanto, M.; Besler, C.; Knau, A.; Fichtlscherer, S.; Röxe, T.; Zeiher, A.M.; Landmesser, U.; Dimmeler, S. Characterization of levels and cellular transfer of circulating lipoprotein-bound microRNAs. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 1392–1400. [Google Scholar] [CrossRef] [PubMed]
- Wang, L.; Jia, X.J.; Jiang, H.J.; Du, Y.; Yang, F.; Si, S.Y.; Hong, B. MicroRNAs 185, 96, and 223 repress selective high-density lipoprotein cholesterol uptake through posttranscriptional inhibition. Mol. Cell. Biol. 2013, 33, 1956–1964. [Google Scholar] [CrossRef] [PubMed]
- Vickers, K.C.; Landstreet, S.R.; Levin, M.G.; Shoucri, B.M.; Toth, C.L.; Taylor, R.C.; Palmisano, B.T.; Tabet, F.; Cui, H.L.; Rye, K.A.; et al. MicroRNA-223 coordinates cholesterol homeostasis. Proc. Natl. Acad. Sci. USA 2014, 111, 14518–14523. [Google Scholar] [CrossRef] [PubMed]
- Jeon, T., II; Esquejo, R.M.; Roqueta-Rivera, M.; Phelan, P.E.; Moon, Y.A.; Govindarajan, S.S.; Esau, C.C.; Osborne, T.F. An SREBP-responsive microRNA operon contributes to a regulatory loop for intracellular lipid homeostasis. Cell Metab. 2013, 18, 51–61. [Google Scholar] [CrossRef] [PubMed]
- Yang, M.; Liu, W.; Pellicane, C.; Sahyoun, C.; Joseph, B.K.; Gallo-Ebert, C.; Donigan, M.; Pandya, D.; Giordano, C.; Bata, A.; et al. Identification of miR-185 as a regulator of de novo cholesterol biosynthesis and low density lipoprotein uptake. J. Lipid Res. 2014, 55, 226–238. [Google Scholar] [CrossRef] [PubMed]
- Ou, Z.; Wada, T.; Gramignoli, R.; Li, S.; Strom, S.C.; Huang, M.; Xie, W. MicroRNA hsa-miR-613 targets the human LXRα gene and mediates a feedback loop of LXRα autoregulation. Mol. Endocrinol. 2011, 25, 584–596. [Google Scholar] [CrossRef] [PubMed]
- Zelcer, N.; Hong, C.; Boyadjian, R.; Tontonos, P. LXR Regulates cholesterol uptake through idol-dependent ubiquitination of the LDL Receptor. Science 2009, 325, 100–104. [Google Scholar] [CrossRef] [PubMed]
- Zhong, D.; Huang, G.; Zhang, Y.; Zeng, Y.; Xu, Z.; Zhao, Y.; He, X.; He, F. MicroRNA-1 and microRNA-206 suppress LXRα-induced lipogenesis in hepatocytes. Cell. Signal. 2013, 25, 1429–1437. [Google Scholar] [CrossRef] [PubMed]
- Clevenger, C.V. Roles and regulation of stat family transcription factors in human breast cancer. Am. J. Pathol. 2004, 165, 1449–1460. [Google Scholar] [CrossRef]
- Mitchell, T.J.; John, S. Signal transducer and activator of transcription (STAT) signalling and T-cell lymphomas. Immunology 2005, 114, 301–312. [Google Scholar] [CrossRef] [PubMed]
- Dubey, R.; Chhabra, R.; Saini, N. Small interfering RNA against transcription factor STAT6 leads to increased cholesterol synthesis in lung cancer cell lines. PLoS ONE 2011, 6, e28509. [Google Scholar] [CrossRef] [PubMed]
- Dubey, R.; Saini, N. STAT6 silencing up-regulates cholesterol synthesis via miR-197/FOXJ2 axis and induces ER stress-mediated apoptosis in lung cancer cells. Biochim. Biophys. Acta Gene Regul. Mech. 2015, 1849, 32–43. [Google Scholar] [CrossRef] [PubMed]
- Ji, A.; Meyer, J.M.; Cai, L.; Akinmusire, A.; de Beer, M.C.; Webb, N.R.; van der Westhuyzen, D.R. Scavenger receptor SR-BI in macrophage lipid metabolism Ailing. Atherosclerosis 2011, 217, 106–112. [Google Scholar] [CrossRef] [PubMed]
- Collot-Teixeira, S.; Martin, J.; McDermott-Roe, C.; Poston, R.; McGregor, J.L. CD36 and macrophages in atherosclerosis. Cardiovasc. Res. 2007, 75, 468–477. [Google Scholar] [CrossRef] [PubMed]
- Mead, J.R.; Ramji, D.P. The pivotal role of lipoprotein lipase in atherosclerosis. Arterioscler. Thromb. 2002, 55, 261–269. [Google Scholar] [CrossRef]
- Zhang, M.; Wu, J.F.; Chen, W.J.; Tang, S.L.; Mo, Z.C.; Tang, Y.Y.; Li, Y.; Wang, J.L.; Liu, X.Y.; Peng, J.; et al. MicroRNA-27a/b regulates cellular cholesterol efflux, influx and esterification/hydrolysis in THP-1 macrophages. Atherosclerosis 2014, 234, 54–64. [Google Scholar] [CrossRef] [PubMed]
- Xu, J.; Hu, G.; Lu, M.; Xiong, Y.; Li, Q.; Chang, C.C.Y.; Song, B.; Chang, T.; Li, B. MiR-9 reduces human acyl-coenzyme A: Cholesterol acyltransferase-1 to decrease THP-1 macrophage-derived foam cell formation. Acta Biochim. Biophys. Sin. 2013, 45, 953–962. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, C.M.; Dávalos, A.; Goedeke, L.; Salerno, A.G.; Warrier, N.; Cirera-Salinas, D.; Suárez, Y.; Fernández-Hernando, C. MicroRNA-758 regulates cholesterol efflux through posttranscriptional repression of ATP-binding cassette transporter A1. Arterioscler. Thromb. Vasc. Biol. 2011, 31, 2707–2714. [Google Scholar] [CrossRef] [PubMed]
- Adlakha, Y.K.; Khanna, S.; Singh, R.; Singh, V.P.; Agrawal, A.; Saini, N. Pro-apoptotic miRNA-128-2 modulates ABCA1, ABCG1 and RXRα expression and cholesterol homeostasis. Cell Death Dis. 2013. [Google Scholar] [CrossRef] [PubMed]
- Kang, M.H.; Zhang, L.H.; Wijesekara, N.; de Haan, W.; Butland, S.; Bhattacharjee, A.; Hayden, M.R. Regulation of ABCA1 protein expression and function in hepatic and pancreatic islet cells by miR-145. Arterioscler. Thromb. Vasc. Biol. 2013, 33, 2724–2732. [Google Scholar] [CrossRef] [PubMed]
- Meiler, S.; Baumer, Y.; Toulmin, E.; Seng, K.; Boisvert, W.A. MicroRNA 302a is a novel modulator of cholesterol homeostasis and atherosclerosis. Arterioscler. Thromb. Vasc. Biol. 2014, 35, 323–331. [Google Scholar] [CrossRef] [PubMed]
- Langmann, T.; Porsch-Özcürümez, M.; Heimerl, S.; Probst, M.; Moehle, C.; Taher, M.; Borsukova, H.; Kielar, D.; Kaminski, W.E.; Dittrich-Wengenroth, E.; et al. Identification of sterol-independent regulatory elements in the human ATP-binding cassette transporter A1 promoter. Role of Sp1/3, E-box binding factors, and an oncostatin M-responsive element. J. Biol. Chem. 2002, 277, 14443–14450. [Google Scholar] [CrossRef] [PubMed]
- Lv, Y.C.; Tang, Y.Y.; Peng, J.; Zhao, G.J.; Yang, J.; Yao, F.; Ouyang, X.P.; He, P.P.; Xie, W.; Tan, Y.L.; et al. MicroRNA-19b promotes macrophage cholesterol accumulation and aortic atherosclerosis by targeting ATP-binding cassette transporter A1. Atherosclerosis 2014, 236, 215–226. [Google Scholar] [CrossRef] [PubMed]
- De Aguiar Vallim, T.Q.; Tarling, E.J.; Kim, T.; Civelek, M.; Baldán, Á.; Esau, C.; Edwards, P.A. MicroRNA-144 regulates hepatic ABCA1 and plasma HDL following activation of the nuclear receptor FXR. Circ. Res. 2013, 112, 1602–1612. [Google Scholar] [CrossRef] [PubMed]
- Rayner, K.J.; Sheedy, F.J.; Esau, C.C.; Hussain, F.N.; Temel, R.E.; Parathath, S.; van Gils, J.M.; Rayner, A.J.; Chang, A.N.; Suarez, Y.; et al. Antagonism of miR-33 in mice promotes reverse cholesterol transport and regression of atherosclerosis. J. Clin. Invest. 2011, 121, 2921–2931. [Google Scholar] [CrossRef] [PubMed]
- Najafi-Shoushtari, S.H.; Kristo, F.; Li, Y.; Shioda, T.; Cohen, D.E.; Gerszten, R.E.; Näär, A.M. MicroRNA-33 and the SREBP host genes cooperate to control cholesterol homeostasis. Science 2010, 328, 1566–1569. [Google Scholar] [CrossRef] [PubMed]
- Marquart, T.J.; Allen, R.M.; Ory, D.S.; Baldán, A. MiR-33 links SREBP-2 induction to repression of sterol transporters. Proc. Natl. Acad. Sci. USA. 2010, 107, 12228–12232. [Google Scholar] [CrossRef] [PubMed]
- Sun, D.; Zhang, J.; Xie, J.; Wei, W.; Chen, M.; Zhao, X. MiR-26 controls LXR-dependent cholesterol efflux by targeting ABCA1 and ARL7. FEBS Lett. 2012, 586, 1472–1479. [Google Scholar] [CrossRef] [PubMed]
- Engel, T.; Lueken, A.; Bode, G.; Hobohm, U.; Lorkowski, S.; Schlueter, B.; Rust, S.; Cullen, P.; Pech, M.; Assmann, G.; et al. ADP-ribosylation factor (ARF)-like 7 (ARL7) is induced by cholesterol loading and participates in apolipoprotein AI-dependent cholesterol export. FEBS Lett. 2004, 566, 241–246. [Google Scholar] [CrossRef] [PubMed]
- Ramirez, C.M.; Rotllan, N.; Vlassov, A.V.; Davalos, A.; Li, M.; Goedeke, L.; Aranda, J.F.; Cirera-Salinas, D.; Araldi, E.; Salerno, A.G.; et al. Control of cholesterol metabolism and plasma HDL levels by miRNA-144. Circ. Res. 2013, 112, 1592–1601. [Google Scholar] [CrossRef] [PubMed]
- Zhao, R.; Feng, J.; He, G. MiR-613 regulates cholesterol efflux by targeting LXRα and ABCA1 in PPARγ activated THP-1 macrophages. Biochem. Biophys. Res. Commun. 2014, 448, 329–334. [Google Scholar] [CrossRef] [PubMed]
- Tarling, E.J.; Ahn, H.; de Aguiar Vallim, T.Q. The nuclear receptor FXR uncouples the actions of miR-33 from SREBP-2. Arterioscler. Thromb. Vasc. Biol. 2015, 35, 787–795. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Xia, M.; Yan, X.; Li, D.; Wang, L.; Xu, Y.; Jin, T.; Ling, W. Gut microbiota metabolism of anthocyanin promotes reverse cholesterol transport in mice via repressing miRNA-10b. Circ. Res. 2012, 111, 967–981. [Google Scholar] [CrossRef] [PubMed]
- Wang, D.; Yan, X.; Xia, M.; Yang, Y.; Li, D.; Li, X.; Song, F.; Ling, W. Coenzyme Q10 promotes macrophage cholesterol efflux by regulation of the activator protein-1/microRNA-378/ATP-binding cassette transporter G1-signaling pathway. Arterioscler. Thromb. Vasc. Biol. 2014, 34, 1860–1870. [Google Scholar] [CrossRef] [PubMed]
- Rayner, K.J.; Suárez, Y.; Dávalos, A.; Parathath, S.; Fitzgerald, M.L.; Tamehiro, N.; Fisher, E.A.; Moore, K.J.; Fernández-hernando, C. MiR-33 contributes to the regulation of cholesterol homeostasis. Science 2010, 328, 1570–1573. [Google Scholar] [CrossRef] [PubMed]
- Li, T.; Francl, J.M.; Boehme, S.; Chiang, J.Y.L. Regulation of cholesterol and bile acid homeostasis by the CYP7A1/SREBP2/miR-33a axis. Hepatology 2013, 58, 1111–1121. [Google Scholar] [CrossRef] [PubMed]
- Allen, R.M.; Marquart, T.J.; Albert, C.J.; Suchy, F.J.; Wang, D.Q.H.; Ananthanarayanan, M.; Ford, D. A.; Baldán, Á. MiR-33 controls the expression of biliary transporters, and mediates statin- and diet-induced hepatotoxicity. EMBO Mol. Med. 2012, 4, 882–895. [Google Scholar] [CrossRef] [PubMed]
- Niesor, E.J.; Schwartz, G.G.; Perez, A.; Stauffer, A.; Durrwell, A.; Bucklar-Suchankova, G.; Benghozi, R.; Abt, M.; Kallend, D. Statin-Induced decrease in ATP-binding cassette transporter A1 expression via microRNA33 induction may counteract cholesterol efflux to high-density lipoprotein. Cardiovasc. Drugs Ther. 2015, 29, 7–14. [Google Scholar] [CrossRef] [PubMed]
- Gerin, I.; Clerbaux, L.A.; Haumont, O.; Lanthier, N.; Das, A.K.; Burant, C.F.; Leclercq, I.A.; MacDougald, O.A.; Bommer, G.T. Expression of miR-33 from an SREBP2 intron inhibits cholesterol export and fatty acid oxidation. J. Biol. Chem. 2010, 285, 33652–33661. [Google Scholar] [CrossRef] [PubMed]
- Horie, T.; Nishino, T.; Baba, O.; Kuwabara, Y.; Nakao, T.; Nishiga, M.; Usami, S.; Izuhara, M.; Sowa, N.; Yahagi, N.; et al. MicroRNA-33 regulates sterol regulatory element-binding protein 1 expression in mice. Nat. Commun. 2013. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jeon, T., II; Park, J.W.; Ahn, J.; Jung, C.H.; Ha, T.Y. Fisetin protects against hepatosteatosis in mice by inhibiting miR-378. Mol. Nutr. Food Res. 2013, 57, 1931–1937. [Google Scholar] [CrossRef] [PubMed]
- Milenkovic, D.; Deval, C.; Gouranton, E.; Landrier, J.F.; Scalbert, A.; Morand, C.; Mazur, A. Modulation of miRNA expression by dietary polyphenols in apoE deficient mice: A new mechanism of the action of polyphenols. PLoS ONE 2012, 7, e29837. [Google Scholar] [CrossRef] [PubMed]
- Iliopoulos, D.; Drosatos, K.; Hiyama, Y.; Goldberg, I.J.; Zannis, V.I. MicroRNA-370 controls the expression of microRNA-122 and Cpt1α and affects lipid metabolism. J. Lipid Res. 2010, 51, 1513–1523. [Google Scholar] [CrossRef] [PubMed]
- Crane, F.L. Biochemical functions of coenzyme Q10. J. Am. Coll. Nutr. 2001, 20, 591–598. [Google Scholar] [CrossRef] [PubMed]
- Parra, P.; Serra, F.; Palou, A. Expression of adipose MicroRNAs is sensitive to dietary conjugated linoleic acid treatment in mice. PLoS ONE 2010, 5, e13005. [Google Scholar] [CrossRef] [PubMed]
© 2015 by the authors; licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
DiMarco, D.M.; Fernandez, M.L. The Regulation of Reverse Cholesterol Transport and Cellular Cholesterol Homeostasis by MicroRNAs. Biology 2015, 4, 494-511. https://doi.org/10.3390/biology4030494
DiMarco DM, Fernandez ML. The Regulation of Reverse Cholesterol Transport and Cellular Cholesterol Homeostasis by MicroRNAs. Biology. 2015; 4(3):494-511. https://doi.org/10.3390/biology4030494
Chicago/Turabian StyleDiMarco, Diana M., and Maria Luz Fernandez. 2015. "The Regulation of Reverse Cholesterol Transport and Cellular Cholesterol Homeostasis by MicroRNAs" Biology 4, no. 3: 494-511. https://doi.org/10.3390/biology4030494