
Allometric Growth Pattern and Hunger Tolerance of Hemibarbus maculatus Bleeker Larvae

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fish Rearing and Sampling
2.2. Measurements
2.3. Data Analysis
3. Results
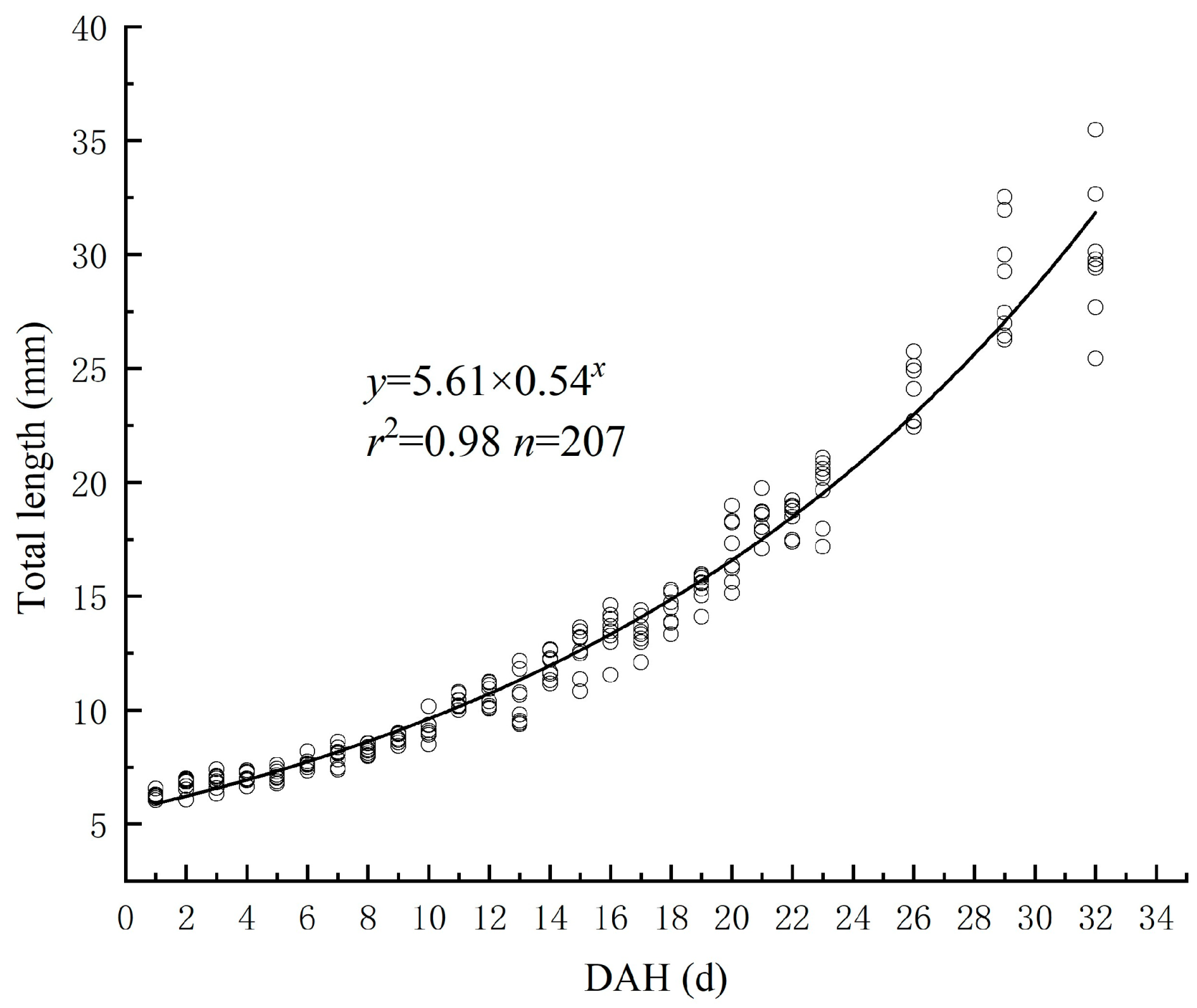
3.1. Relationship between TL and DAH
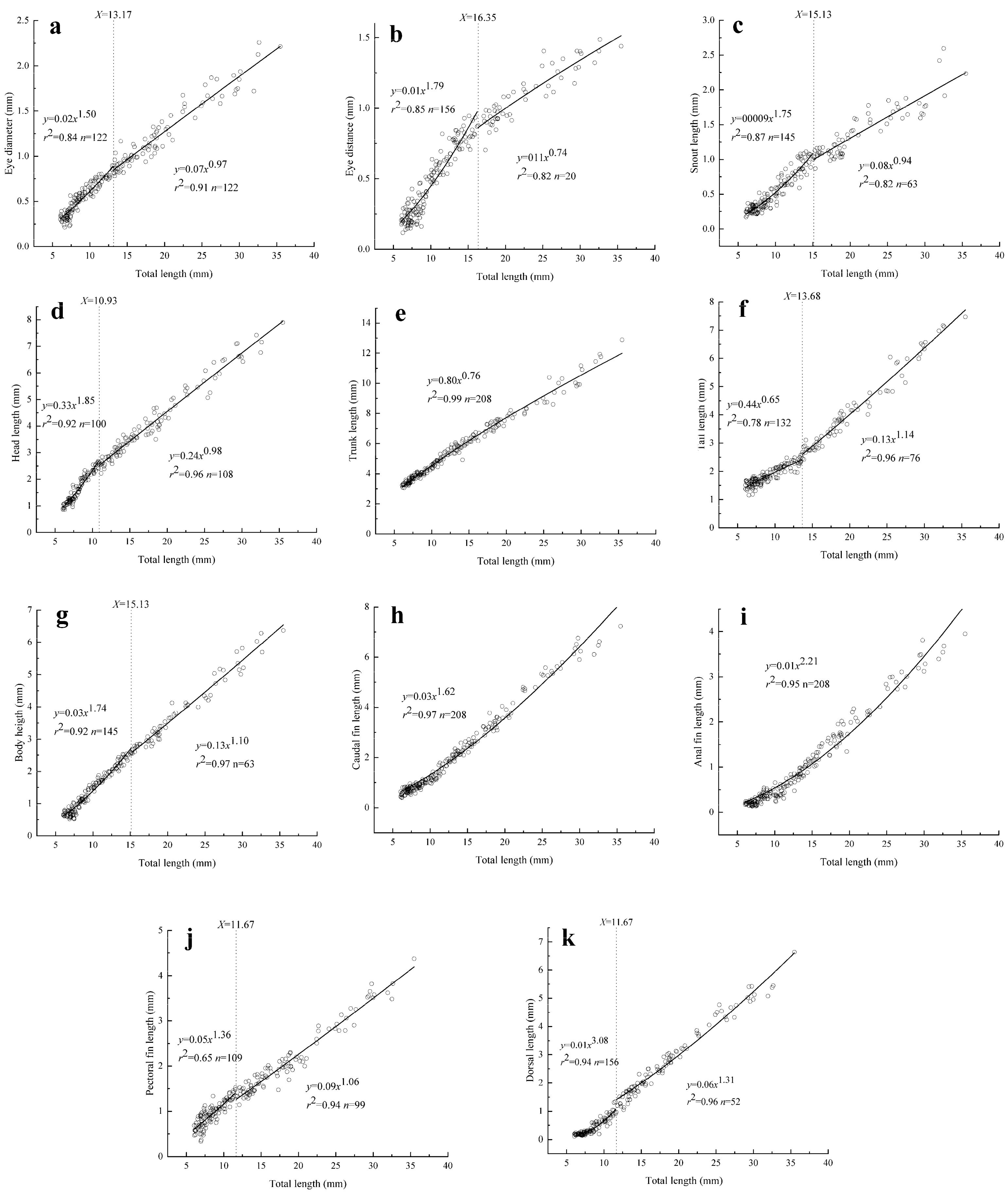
3.2. Allometric growth
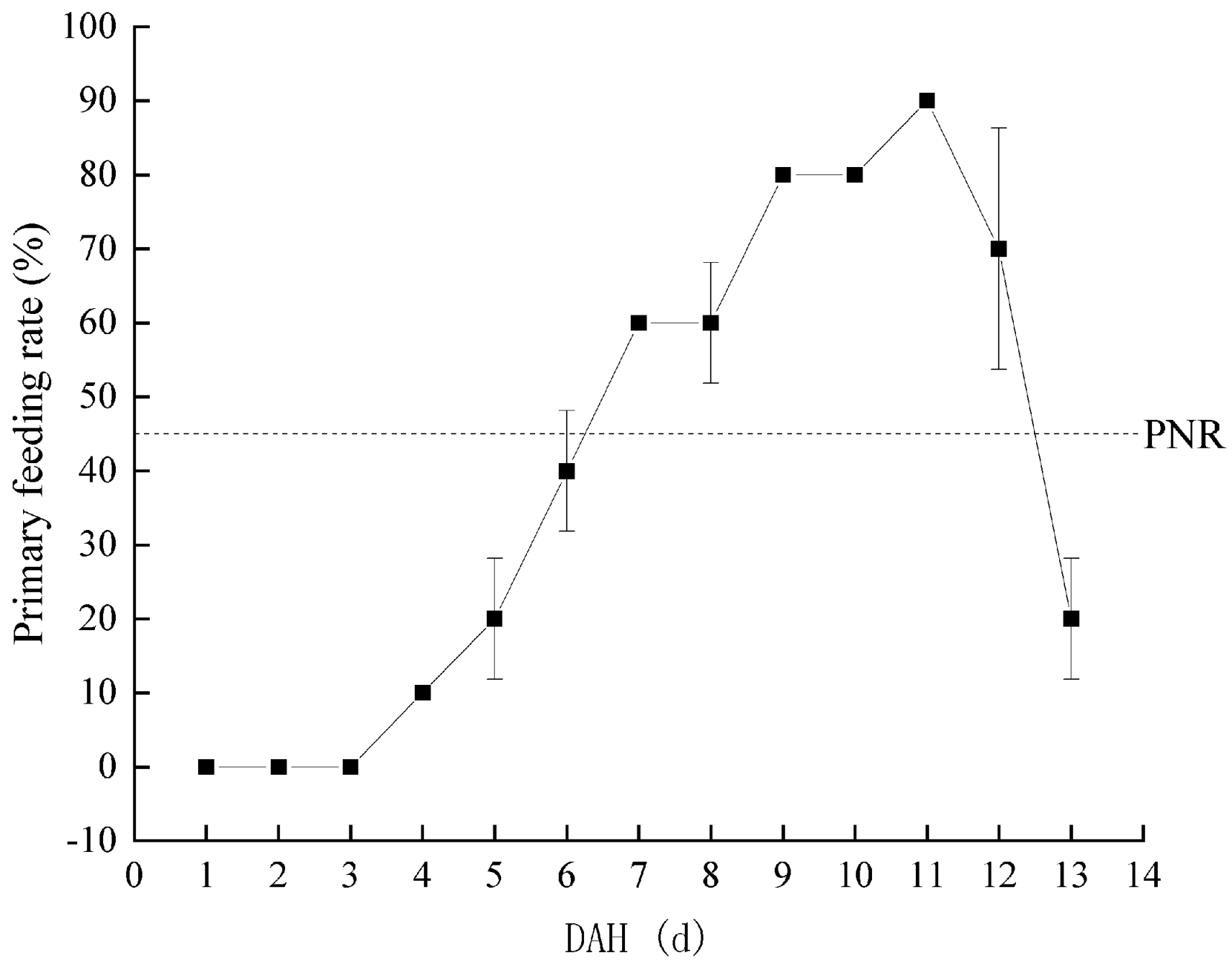
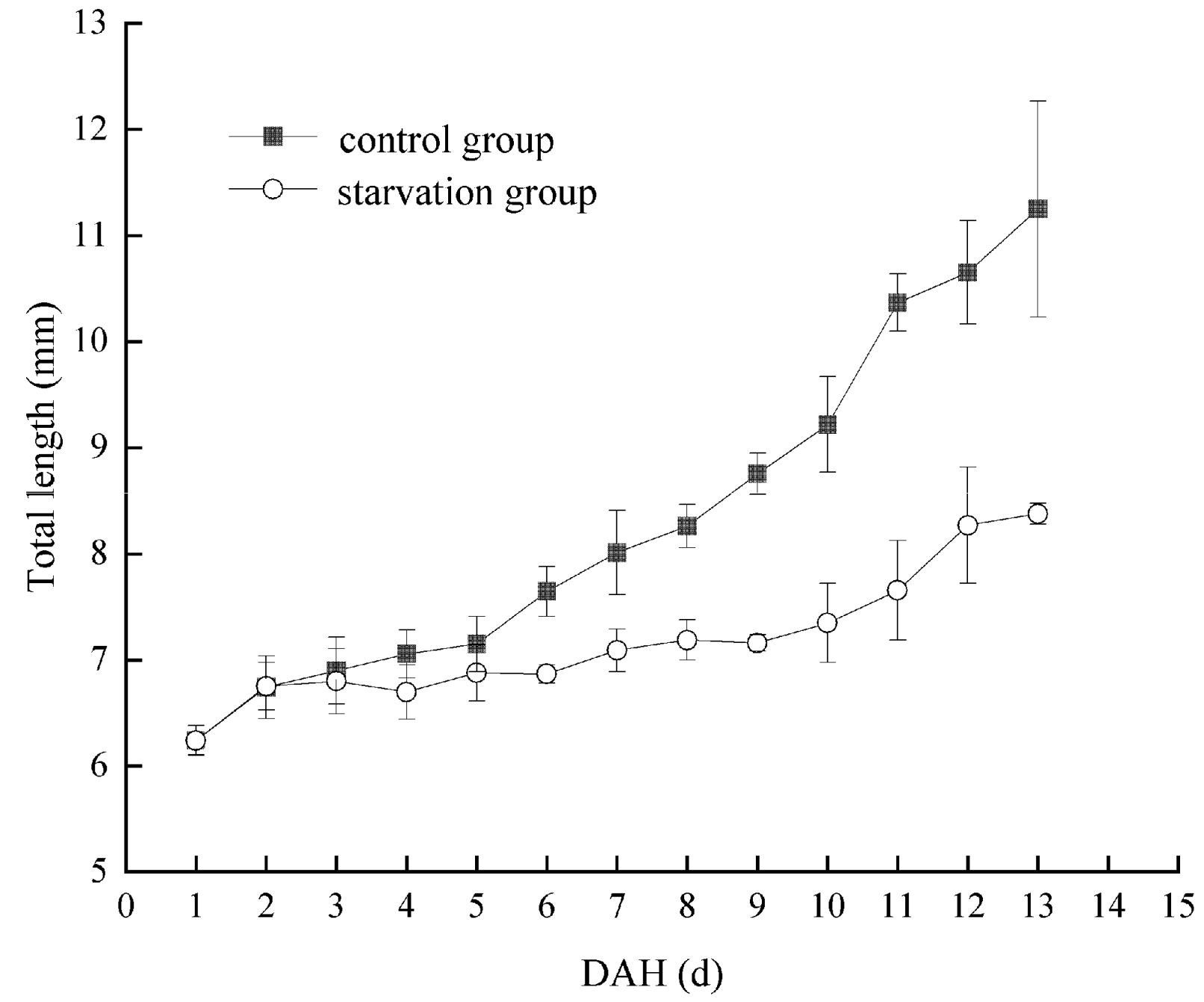
3.3. Effects of Starvation on Feeding and Growth of Larvae of Hemibarbus maculatus
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bao, B.L.; Su, J.X.; Yin, M.C. Effect of delayed feeding on feeding ability, survival and growth of red seabream and olive flounder larvae during early development. J. Fish. China 1998, 22, 33–38. [Google Scholar]
- Song, Y.Q.; Cheng, F.; Zhao, S.S.; Xie, S.G. Ontogenetic development and otolith microstructure in the larval and juvenile stages of mandarin fish Siniperca chuatsi. Ichthyol. Res. 2019, 66, 57–66. [Google Scholar] [CrossRef]
- Huxley, J.S.; Tessier, G. Terminology of relative growth. Nature 1936, 137, 780–781. [Google Scholar] [CrossRef]
- Osse, J.; Boogaart, J.; Snik, G.; Sluys, L. Priorities during early growth of fish larvae. Aquaculture 1997, 155, 249–258. [Google Scholar] [CrossRef]
- Choo, C.K.; Liew, H.C. Morphological development and allometric growth patterns in the juvenile seahorse Hippocampus kuda Bleeker. J. Fish Biol. 2010, 69, 426–445. [Google Scholar] [CrossRef]
- Ma, J.; Zhang, L.Z.; Zhuang, P.; Zhang, T.; Feng, G.P.; Zhao, F. Development and allometric growth of larval Acipenser schrenckii. Chin. J. Appl. Ecol. 2007, 18, 2875–2882. [Google Scholar]
- Zhuang, P.; Song, C.; Zhang, L.Z.; Zhang, T.; Huang, X.R.; Wang, B. Allometric growth of artificial bred siberian sturgeon acipenser baeri larvae and juveniles. Chin. J. Ecol. 2009, 28, 681–687. [Google Scholar]
- Wang, Y.F.; Xiao, Z.Z.; Liu, Q.H.; Zhai, J.M.; Pang, Z.F.; Ma, W.H.; Ma, D.Y.; Xu, S.H.; Xiao, Y.S.; Li, J. Allometric growth pattern during early ontogeny of spotted knifejaw (Oplegnathus punctatus). Mar. Sci. 2016, 40, 43–48. [Google Scholar] [CrossRef]
- Van Sink, G.M.J.; Van den Boogaart, J.G.M.; Osse, J.W.M. Larval growth patterns in Cyprinus carpio and Clarias gariepinus with attention to the finfold. J. Fish Biol. 1997, 50, 1339–1352. [Google Scholar] [CrossRef]
- Herbing, I. Development of feeding structures in larval fish with different life histories: Winter flounder and Atlantic cod. J. Fish Biol. 2001, 59, 767–782. [Google Scholar] [CrossRef]
- Nogueira, L.B.; Godinho, A.L.; Godinho, H.P. Early development and allometric growth in hatchery-reared characin Brycon orbignyanus. Aquac. Res. 2012, 45, 1004–1011. [Google Scholar] [CrossRef]
- Gao, L.; Duan, M.; Cheng, F.; Xie, S.G. Ontogenetic development in the morphology and behavior of loach (Misgurnus anguillicaudatus) during early life stages. Chin. J. Oceanol. Limnol. 2014, 32, 973–981. [Google Scholar] [CrossRef]
- He, T.; Xiao, Z.Z.; Liu, Q.H.; Li, J. Allometric growth in rock bream larvae (Oplegnathus fasciatus Temminck et Schlegel 1844). J. Fish. China 2012, 36, 1242–1248. [Google Scholar] [CrossRef]
- Liu, Y.; Cheng, D.; Zhou, S.; Ma, Z.; Hu, J.; Yang, R. Study on the Allometric Growth of Larvae and Juvenile Lates calarifer. Mar. Fish. 2018, 40, 179–188. [Google Scholar] [CrossRef]
- He, Y.F.; Wu, X.B.; Zhu, Y.J.; Yang, D.G.; Zhan, H.X. Allometric growth pattern of Percocypris pingi larvae. Chin. J. Zool. 2013, 48, 8–15. [Google Scholar] [CrossRef]
- Xi, D. Growth Pattern and Cannibalism in Black Rockfish (Sebastes schlegelii) during Early Development Stage; Ocean University of China: Qingdao, China, 2014. [Google Scholar]
- Zhang, J.M.; Yao, J.Z.; Jiang, W.; Guo, W.T. Experimental starvation on Percocypris pingi larvae and determination of the point of no return. J. South. Agric. 2020, 51, 2565–2572. [Google Scholar] [CrossRef]
- Shi, H.L.; Lou, B.; Mao, G.M.; Luo, J.A. Study on feeding and growth of larval Zhoushan flounder (Paralichthys olivaceus). South China Fish. Sci. 2008, 4, 21–25. [Google Scholar] [CrossRef]
- Zhang, Y.Q.; Yin, J.S.; Du, J.; Jia, Z.H.; Tong, G.X. Experiment starvation on Hucho taimen and definition of the point of no return. Acta Hydrobiol. Sin. 2009, 33, 945–950. [Google Scholar] [CrossRef]
- Li, Y.Q.; Jiang, Z.Q.; Sun, Y.; Mao, M.G.; Meng, X.K. Experimental starvation on Gadus macrocephalus and definition of the point of no return. Acta Ecol. Sin. 2014, 34, 3873–3878. [Google Scholar] [CrossRef]
- Yang, Y.K.; Lin, H.Z.; Wang, Z.Y.; Xie, Y.J.; Guo, Y.H.; Huang, X.L.; Li, T. Experimental starvation of Acanthopagrus schlegelii larvae, and definition of the point of no return. Prog. Fish. Sci. 2020, 41, 87–94. [Google Scholar] [CrossRef]
- Wu, Y.A.; Li, H.; Liao, F.C.; Yang, X.; Xie, Z.G. Fish Chronicles of Hunan; Science Press: Beijing, China, 2021. [Google Scholar]
- Zhu, G.Y. Effect of Dietary Lipid (Energy) to Protein Ratio and Lipid Sources on Growth, Physiological Function and Fatty Acid Composion of Hemibarbus maculatus. Ph.D. Thesis, Suzhou University, Su Zhou, China, 2009. [Google Scholar] [CrossRef]
- Li, H.; Yang, X.; Wu, Y.; Yuan, X.; Liang, Z.; Tian, X.; Ge, H.; Liao, F.; Chen, B.; Liu, J. Assessment of age, growth and stock for Hemibarbus maculatus in Dongting Lake. Freshw. Fish. 2020, 50, 60–67. [Google Scholar]
- Gisbert, E. Early development and allometric growth patterns in Siberian sturgeon and their ecological significance. J. Fish Biol. 1999, 54, 852–862. [Google Scholar] [CrossRef]
- Lu, H.; Li, Z.; Fang, M.; Tong, X. A primary study on the biological characteristics and artificial breeding of Hemibarbus maculatus Bleeker in pond. J. Zhejiang Ocean. Univ. 2004, 23, 211–213. [Google Scholar] [CrossRef]
- Yang, Q.; Ma, Z.; Cheng, D.; Jiang, S.; Li, Y.; Chen, M. Allometric growth in larvae and juvenile golden pompano (Trachinotus ovatus). Fish Sci. 2017, 36, 259–266. [Google Scholar] [CrossRef]
- Xu, B.; Li, D.; Wei, K.; Zhu, X.; Xu, J.; Ma, B. Allometric Growth Patterns and Ontogenetic Development during Early Larval Stages of Schizothorax waltoni Regan and Percocypris retrodorslis in Southwest China. Water 2023, 15, 824. [Google Scholar] [CrossRef]
- Ping, H.L.; Yu, F.P.; Zhang, T.; He, J.Y.; Shi, H.L.; Lu, B. Morphological development and allometric growth pattern of Konosirus Punctatus at larvae and juvenile stages. Acta Hydrobiol. Sin. 2019, 43, 1021–1033. [Google Scholar] [CrossRef]
- Fuiman, L.A. Growth gradients in fish larvae. J. Fish Biol. 1983, 23, 117–123. [Google Scholar] [CrossRef]
- Shea, E.; Vecchione, M. Quantification of ontogenetic discontinuities in three species of oegopsid squids using model II piecewise linear regression. Mar. Biol. 2002, 140, 971–979. [Google Scholar] [CrossRef]
- Wisely, S.M.; Santymire, R.M.; Livieri, T.M.; Marinari, P.E.; Kreeger, J.S.; Wildt, D.E.; Howard, J. Environment influences morphology and development for in situ and ex situ populations of the black-footed ferret (Mustela nigripes). Anim. Conserv. 2005, 8, 321–328. [Google Scholar] [CrossRef]
- Gisbert, E.; Asgari, R.; Rafiee, G.; Agh, N.; Eagderi, S.; Eshaghzadeh, H.; Alcaraz, C. Early development and allometric growth patterns of beluga Huso (Linnaeus, 1758). J. Appl. Ichthyol. 2015, 30, 1264–1272. [Google Scholar] [CrossRef]
- Chen, D.C.; Ma, Z.H.; Jiang, S.G. Allometric growth in larvae and juvenile crimson snapper Lutjanus erythopterus. Acta Hydrobiol. Sin. 2017, 41, 206–213. [Google Scholar] [CrossRef]
- Song, H.J.; Liu, W.; Wang, J.L.; Tang, F.J. Allometric growth during yolk-sac larvae of chum salmon (Oncorhynchus keta Walbaum) and consequent ecological significance. Acta Hydrobiol. Sin. 2013, 37, 329–335. [Google Scholar] [CrossRef]
- Xie, M.; Wu, H.; Song, R.; Xiang, J.; Zhou, J.; Li, H.; Zeng, G.Q.; Li, S.M.; Xiang, J.G. Allometric growth pattern and point of no return for starvation of Furong crucian carp (Carassius auratus Furong carp ♀ × Cyprinus carpio red crucian carp ♂). J. South. Agric. 2023, 54, 1253–1262. [Google Scholar] [CrossRef]
- Shan, X.J.; Dou, S.Z. Allometric growth of croaker (Miichthys miiuy) larvae and juveniles and its ecological implication. Oceanol. Limnol. Sin. 2009, 40, 714–719. [Google Scholar] [CrossRef]
- Lin, Y.Z.; Yu, D.D.; Wem, H.S.; Li, J.F.; Mu, W.J.; Liu, Q.; Zhao, J.L.; Ren, Y.Y.; Zhang, Y.C. Morphometrics development of Ovoviviparous Sebastes Schlegeli Hilgendorf larvae and juvenile. Trans. Oceanol. Limnol. 2014, 2, 51–58. [Google Scholar]
- Peña, R.; Dumas, S. Development and allometric growth patterns during early larval stages of the spotted sand bass Paralabrax maculatofasciatus (Percoidei: Serranidae). Sci. Mar. 2009, 73, 183–189. [Google Scholar] [CrossRef]
- Hu, J.; Zhou, S.J.; Yang, X.; Chen, X.; Yang, Q.B.; Ma, Z.H. Ontogenetic allometry in larval and juvenile Clown Anemone fish Amphiprion ocellaris. Acta Hydrobiol. Sin. 2020, 44, 844–852. [Google Scholar] [CrossRef]
- Yang, Y.K.; Yu, W.; Lin, H.Z.; Li, T.; Huang, X.L.; Huang, Z.; Qi, C.L. Experimental starvation of Plectropomus leopardus larvae and study of point of no return. South China Fish. Sci. 2017, 13, 90–96. [Google Scholar] [CrossRef]
- Gao, X.Q.; Hong, L.; Liu, Z.F.; Guo, Z.L.; Wang, Y.H.; Lei, J.L. The study of allometric growth pattern of american shad larvae and juvenile (Alosa Sapidissima). Acta Hydrobiol. Sin. 2015, 39, 638–644. [Google Scholar] [CrossRef]
- Johnston, I.A.; Hall, T.E. Mechanisms of muscle development and responses to temperature change in fish larvae. In The Development of Form and Function in Fishes and the Question of Larval Adaptation; Govoni, J.J., Ed.; Symposium 40; American Fisheries Society: Bethesda, MD, USA, 2004; pp. 85–116. [Google Scholar]
- Kamler, E. Early Life History of Fish: An Energetics Approach; Springer: Berlin/Heidelberg, Germany, 1992. [Google Scholar] [CrossRef]
- Shan, X.J.; Dou, S.Z. Effect of delayed first feeding on growth, survival and biochemical composition of Croaker Michthys Miiuy Larvae. Oceanol. Limnol. Sin. 2008, 39, 14–23. [Google Scholar]
- Huang, H.; Yang, N.; Zhang, X. Experimental starvation on Cichlasoma managuense larvae and determination of point of no return. South China Fish. Sci. 2012, 8, 43–50. [Google Scholar] [CrossRef]
- Chen, S.Y.; Zhang, Z.Y.; Xu Ji, J.G.; Wu, G.J.; Jia, C.F.; Zhu, F.; Meng, Q. Study on larval early development and point of no return of hybrid F2 of Acanthopagrus schiegelii (♀) × Pagrus major (♂). South China Fish. Sci. 2018, 14, 122–128. [Google Scholar] [CrossRef]
- Li, C.J.; Xu, Z.C.; Zhang, Z.Z.; Wang, H.; Ling, Q.F. Starvation and point of no return of Squaliobarbus curriculus larvae. J. Yangzhou Univ. (Agric. Life Sci. Ed.) 2016, 37, 65–70. [Google Scholar] [CrossRef]
- Peng, Z.L.; Liu, M.H.; Luo, H.Z.; Fu, R.B.; Zhang, F.P.; Mao, Z.Z. Starvation and point of no return in Striped Knifejaw Oplegnathus fasciatus larvae. Fish. Sci. 2010, 29, 152–155. [Google Scholar]
- Xu, X.J.; Ou, Y.J.; Li, J.E. Effect of delayed feeding on foraging, survival and growth of ovate pompano (Trachinotus ovatus) larvae during early development stage. South China Fish. Sci. 2010, 6, 37–41. [Google Scholar] [CrossRef]
- Li, Q.; Li, M.J.; Zhou, C.J.; Chen, X.J. Effects of starvation on feeding and growth of the larvae of Onychostoma sima. Freshw. Fish. 2009, 39, 32–37. [Google Scholar] [CrossRef]
- Duan, G.Q.; Jiang, H.; Hu, W.; Ling, J.; Hu, Y.T. Experimental starvation on Monopterus albus larvae and determination of the point of no return. J. South. Agric. 2013, 44, 1036–1040. [Google Scholar] [CrossRef]





Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Xie, M.; Wang, P.; Deng, Q.; Liu, S.; Zhang, Z.; Wu, H.; Xiang, J.; Zhou, J.; Yang, X.; Song, R.; et al. Allometric Growth Pattern and Hunger Tolerance of Hemibarbus maculatus Bleeker Larvae. Biology 2024, 13, 164. https://doi.org/10.3390/biology13030164
Xie M, Wang P, Deng Q, Liu S, Zhang Z, Wu H, Xiang J, Zhou J, Yang X, Song R, et al. Allometric Growth Pattern and Hunger Tolerance of Hemibarbus maculatus Bleeker Larvae. Biology. 2024; 13(3):164. https://doi.org/10.3390/biology13030164
Chicago/Turabian StyleXie, Min, Pengpeng Wang, Qi Deng, Si Liu, Zhou Zhang, Hao Wu, Jing Xiang, Jie Zhou, Xin Yang, Rui Song, and et al. 2024. "Allometric Growth Pattern and Hunger Tolerance of Hemibarbus maculatus Bleeker Larvae" Biology 13, no. 3: 164. https://doi.org/10.3390/biology13030164