
Wolbachia Interactions with Diverse Insect Hosts: From Reproductive Modulations to Sustainable Pest Management Strategies
 , and
, and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Wolbachia Biology: Host Ecology, Diversity, and Genomics
3. Wolbachia for Nature’s Collaborations to Control Insect Populations
4. Wolbachia Dynamics in Lepidopteran and Dipteran Insects: Geographic Structure and Male Killing Aspects
5. The Function of Scale Insect Companion Species in Wolbachia Host Fluctuations
6. The Use of Wolbachia in Leafhopper Vectors to Regulate Plant Hopper Populations
7. Future Prospective and Challenges
8. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Bordenstein, S.R.; Theis, K.R. Host biology in light of the microbiome: Ten principles of Holobionts and Hologenomes. PLoS Biol. 2015, 13, e1002226. [Google Scholar] [CrossRef]
- McFall-Ngai, M. Are biologists in ‘‘future shock’’? Symbiosis integrates biology across domains. Nat. Rev. Microbiol. 2008, 6, 789–792. [Google Scholar] [CrossRef]
- Hertig, M.; Wolbach, S.B. Studies on Rickettsia-like micro-organisms in insects. J. Med. Res. 1924, 44, 329–374.7. [Google Scholar]
- Kaur, R.; Shropshire, J.D.; Cross, K.L.; Leigh, B.; Mansueto, A.J.; Stewart, V.; Bordenstein, S.R. Living in the endosymbiotic world of Wolbachia: A centennial review. Cell Host Microbe 2021, 29, 879–893. [Google Scholar] [CrossRef]
- Hurst, G.D.D.; Frost, C.L. Reproductive parasitism: Maternally inherited symbionts in a biparental world. Cold Spring Harb. Perspect. Biol. 2015, 7, a017699. [Google Scholar]
- Bliman, P.A.; Aronna, M.S.; Coelho, F.C.; Da Silva, M.A.H.B. Ensuring successful introduction of Wolbachia in natural populations of Aedes aegypti by means of feedback control. J. Math. Biol. 2018, 76, 1269–1300. [Google Scholar] [CrossRef]
- Kean, J.; Rainey, S.M.; Mcfarlane, M.; Donald, C.L.; Schnettler, E.; Kohl, A.; Pondeville, E. Fighting arbovirus transmission: Natural and engineered control of vector competence in Aedes mosquitoes. Insects 2015, 1, 236–278. [Google Scholar] [CrossRef]
- Iturbe-Ormaetxe, I.; Walker, T.; O’Neill, S.L. Wolbachia and the biological control of mosquito-borne disease. EMBO Rep. 2011, 12, 508–518. [Google Scholar] [CrossRef]
- Aliota, M.T.; Walker, E.C.; Yepes, A.U.; Velez, I.D.; Christensen, B.M.; Osorio, J.E. The wMel strain of Wolbachia reduces transmission of chikungunya virus in Aedes aegypti. PLoS Negl. Trop. Dis. 2016, 10, e0004677. [Google Scholar] [CrossRef] [PubMed]
- Hertig, M. The rickettsia, Wolbachia pipientis and associated inclusions of the mosquito, Culex Pipiens. Parasitology 1936, 28, 453–486. [Google Scholar] [CrossRef]
- Sironi, M.; Bandi, C.; Sacchi, L.; Di, B.S.; Damiani, G.; Genchi, C. Molecular evidence for a close relative of the arthropod endosymbiont Wolbachia in a filarial worm. Mol. Biochem. Parasitol. 1995, 74, 223–227. [Google Scholar] [CrossRef] [PubMed]
- Werren, J.H. Biology of wolbachia. Annu. Rev. Entomol. 1997, 42, 587–609. [Google Scholar] [CrossRef]
- Weinert, L.A.; Araujo-Jnr, E.V.; Ahmed, M.Z.; Welch, J.J. The incidence of bacterial endosymbionts in terrestrial arthropods. Proc. R. Soc. B Biol. Sci. 2015, 282, 20150249. [Google Scholar] [CrossRef]
- Zug, R.; Hammerstein, P. Still a host of hosts for Wolbachia: Analysis of recent data suggests that 40% of terrestrial arthropod species are infected. PLoS ONE 2012, 7, e38544. [Google Scholar] [CrossRef] [PubMed]
- Werren, J.H.; Baldo, L.; Clark, M.E. Wolbachia: Master manipulators of invertebrate biology. Nat. Rev. Microbiol. 2008, 6, 741–751. [Google Scholar] [CrossRef] [PubMed]
- Hoffmann, A.A.; Ross, P.A.; Rašić, G. Wolbachia strains for disease control: Ecological and evolutionary considerations. Evol. Appl. 2015, 8, 751–768. [Google Scholar] [CrossRef] [PubMed]
- Ross, P.A.; Turelli, M.; Hoffmann, A.A. Evolutionary ecology of Wolbachia releases for disease control. Annu. Rev. Genet. 2019, 53, 93–116. [Google Scholar] [CrossRef]
- Charleston, M.A.; Perkins, S.L. Traversing the tangle: Algorithms and applications for cophylogenetic studies. J. Bio. Infor. 2006, 39, 62–71. [Google Scholar] [CrossRef]
- Thompson, J.N. Symbiont-induced speciation. Biol. J. Linn. Soc. 1987, 32, 385–393. [Google Scholar] [CrossRef]
- Fenn, K.; Blaxter, M. Are filarial nematode Wolbachia obligate mutualist symbionts? Trends Ecol. Evol. 2004, 19, 163–166. [Google Scholar] [CrossRef]
- Balvín, O.; Roth, S.; Talbot, B.; Reinhardt, K. Co-speciation in bedbug Wolbachia parallel the pattern in nematode hosts. Sci. Rep. 2018, 8, 8797. [Google Scholar] [CrossRef]
- Shoemaker, D.D.; Machado, C.A.; Molbo, D.; Werren, J.H.; Windsor, D.M.; Herre, E.A. The distribution of Wolbachia in fig wasps: Correlations with host phylogeny, ecology and population structure. Proc. R. Soc. Lond. B Biol. 2002, 269, 2257–2267. [Google Scholar] [CrossRef]
- Frost, C.L.; Fernandez-Marin, H.; Smith, J.E.; Hughes, W.O.H. Multiple gains and losses of Wolbachia symbionts across a tribe of fungus-growing ants. Mol. Ecol. 2010, 19, 4077–4085. [Google Scholar] [CrossRef]
- Ahmed, M.Z.; Breinholt, J.W.; Kawahara, A.Y. Evidence for common horizontal transmission of Wolbachia among butterflies and moths. BMC Evol. Biol. 2016, 16, 118. [Google Scholar] [CrossRef] [PubMed]
- Gerth, M.; Röthe, J.; Bleidorn, C. Tracing horizontal Wolbachia movements among bees (Anthophila): A combined approach using multilocus sequence typing data and host phylogeny. Mol. Ecol. 2013, 22, 6149–6162. [Google Scholar] [CrossRef] [PubMed]
- Sanaei, E.; Charlat, S.; Engelstädter, J. Wolbachia host shifts: Routes, mechanisms, constraints and evolutionary consequences. Biol. Rev. 2021, 96, 433–453. [Google Scholar] [CrossRef] [PubMed]
- Heath, B.D.; Butcher, R.D.J.; Whitfield, W.G.F.; Hubbard, S.F. Horizontal transfer of Wolbachia between phylogenetically distant insect species by a naturally occurring mechanism. Curr. Biol. 2021, 9, 313–316. [Google Scholar] [CrossRef] [PubMed]
- Cooper, B.S.; Vanderpool, D.; Conner, W.R.; Matute, D.R.; Turelli, M. Wolbachia acquisition by drosophila yakuba-clade hosts and transfer of incompatibility loci between distantly related Wolbachia. Genetics 2019, 212, 1399–1419. [Google Scholar] [CrossRef] [PubMed]
- Turelli, M.; Cooper, B.S.; Richardson, K.M.; Ginsberg, P.S.; Peckenpaugh, B.; Antelope, C.X.; Kim, K.J.; May, M.R.; Abrieux, A.; Wilson, D.A.; et al. Rapid global spread of wRi-like Wolbachia across multiple drosophila. Curr. Biol. 2018, 28, 963–971.e8. [Google Scholar] [CrossRef] [PubMed]
- Hughes, G.L.; Rasgon, J.L. Transinfection: A method to investigate Wolbachia–host interactions and control arthropodborne disease. Insect Mol. Biol. 2015, 23, 141–151. [Google Scholar] [CrossRef]
- Ilinsky, Y.; Kosterin, O.E. Molecular diversity of Wolbachia in Lepidoptera: Prevalent allelic content and high recombination of MLST genes. Mol. Phylogenetics Evol. 2017, 109, 164–179. [Google Scholar] [CrossRef]
- Riegler, M.; Charlat, S.; Stauffer, C.; Merçot, H. Wolbachia transfer from Rhagoletis cerasi to Drosophila simulans: Investigating the outcomes of host-symbiont coevolution. Appl. Environ. Microbiol. 2004, 70, 273–279. [Google Scholar] [CrossRef]
- Vavre, F.; Fleury, F.; Lepetit, D.; Fouillet, P.; Boulétreau, M. Phylogenetic evidence for horizontal transmission of Wolbachia in host-parasitoid associations. Mol. Biol. Evol. 1999, 16, 1711–1723. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, M.Z.; Li, S.J.; Xue, X.; Yin, X.J.; Ren, S.X.; Jiggins, F.M.; Greeff, J.M.; Qiu, B.L. The intracellular bacterium Wolbachia uses parasitoid wasps as phoretic vectors for efficient horizontal transmission. PLoS Pathog. 2015, 11, e1004672. [Google Scholar] [CrossRef] [PubMed]
- Cook, J.M.; Butcher, R.D.J. The transmission and effects of Wolbachia bacteria in parasitoids. Res. Popul. Ecol. 1999, 41, 15–28. [Google Scholar] [CrossRef]
- Le Clec’h, W.; Chevalier, F.D.; Genty, L.; Bertaux, J.; Bouchon, D.; Sicard, M. Cannibalism and predation as paths for horizontal passage of Wolbachia between terrestrial isopods. PLoS ONE 2013, 8, e60232. [Google Scholar] [CrossRef] [PubMed]
- Li, S.J.; Ahmed, M.Z.; Lv, N.; Shi, P.Q.; Wang, X.M.; Huang, J.L.; Qiu, B.L. Plantmediated horizontal transmission of Wolbachia between whiteflies. ISME J. 2017, 11, 1019–1028. [Google Scholar] [CrossRef] [PubMed]
- Mouton, L.H.; Henri, M.; Bouletreau Vavre, F. Strain-specifc regulation of intracellular Wolbachia density in multiply infected insects. Mol. Ecol. 2003, 12, 3459–3465. [Google Scholar] [CrossRef] [PubMed]
- Correa, C.C.; Ballard, J.W. Wolbachia gonadal density in female and male Drosophila vary with laboratory adaptation and respond differently to physiological and environmental challenges. J. Invertebr. Pathol. 2012, 111, 197–204. [Google Scholar] [CrossRef] [PubMed]
- López-Madrigal, S.; Duarte, E.H. Titer regulation in arthropod Wolbachia symbioses. FEMS Microbiol. Lett. 2019, 366, 1–9. [Google Scholar] [CrossRef]
- Hanboonsong, Y.; Ritthison, W.; Choosai, C.; Sirithorn, P. Transmission of sugarcane white leaf phytoplasma by Yamatotettix flavovittatus, a new leafhopper vector. J. Econ. Entomol. 2006, 99, 1531–1537. [Google Scholar] [CrossRef]
- Thein, M.M.; Jamjanya, T.; Kobori, Y.; Hanboonsong, Y. Dispersal of the leafhoppers Matsumuratettix hiroglyphicus and Yamatotettix flavovittatus (Homoptera: Cicadellidae), vectors of sugarcane white leaf disease. Appl. Entomol. Zool. 2012, 47, 255–262. [Google Scholar] [CrossRef]
- Youichi, K.; Hanboonsong, Y. Effect of temperature on the development and reproduction of the sugarcane white leaf insect vector, Matsumuratettix hiroglyphicus (Matsumura) (Hemiptera: Cicadellidae). J. Asia Pac. Entomol. 2017, 20, 281–284. [Google Scholar]
- Wangkeeree, J.; Tewaruxsa, P.; Roddee, J.; Hanboonsong, Y. Wolbachia (Rickettsiales: Alphaproteobacteria) infection in the leafhopper vector of sugarcane white leaf disease. J. Insect Sci. 2020, 20, 20. [Google Scholar] [CrossRef] [PubMed]
- Wangkeeree, J.; Suwanchaisri, K.; Roddee, J.; Hanboonsong, Y. Effect of Wolbachia infection states on the life history and reproductive traits of the leafhopper Yamatotettix flavovittatus Matsumura. J. Invertebr. Pathol. 2020, 177, 107490. [Google Scholar] [CrossRef]
- Hoffmann, A.A.; Turelli, M. Unidirectional incompatibility in Drosophila simulans: Inheritance, geographic variation and fitness effects. Genetics 1998, 119, 435444. [Google Scholar] [CrossRef] [PubMed]
- Moran, N.; Dunbar, A.H.E. Sexual acquisition of beneficial symbionts in aphids. Proc. Natl. Acad. Sci. USA 2006, 103, 12803–12806. [Google Scholar] [CrossRef] [PubMed]
- Chafee, M.; Funk, E.; Harrison, D.J.; Bordenstein, R.G. Lateral phage transfer in obligate intracellular bacteria (Wolbachia): Verification from natural populations. Mol. Biol. Evol. 2010, 27, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Saridaki, A.; Bourtzis, K. Wolbachia: More than just a bug in insects genitals. Curr. Opin. Microbiol. 2010, 13, 67–72. [Google Scholar] [CrossRef]
- Schneider, D.; Miller, W.J.; Riegler, M. Arthropods shopping for Wolbachia. In Manipulative Tenants Bacteria Associated with Arthropods; Zchori-Fein, E., Bourtzis, K., Eds.; CRC Press: Boca Raton, Fl, USA, 2011; pp. 149–173. [Google Scholar]
- Yen, J.H.; Barr, A.R. New hypothesis of the cause of cytoplasmic incompatibility in Culex pipiens L. Nature 1971, 232, 657–658. [Google Scholar] [CrossRef]
- LePage, D.; Bordenstein, S. Wolbachia: Can we save lives with a great pandemic? Trends Parasitol. 2013, 29, 385–393. [Google Scholar] [CrossRef]
- Bian, G.; Xu, Y.; Lu, P.; Xie, Y.; Xi, Z. The Endosymbiotic Bacterium Wolbachia Induces Resistance to Dengue Virus in Aedes aegypti. PLoS Pathog. 2013, 9, e1003531. [Google Scholar] [CrossRef] [PubMed]
- Baton, L.A.; Pacidônio, E.C.; Gonçalves, D.d.S.; Moreira, L.A. WFLU: Characterization and evaluation of a native Wolbachia from the mosquito Aedes fluviatilis as a potential vector control agent. PLoS ONE 2013, 8, e59619. [Google Scholar] [CrossRef]
- Duplouy, A.; Iturbe-Ormaetxe, I.; A Beatson, S.; Szubert, J.M.; Brownlie, J.C.; McMeniman, C.J.; A McGraw, E.; Hurst, G.D.D.; Charlat, S.; O’neill, S.L.; et al. Draft Genome Sequence of the Male-Killing Wolbachia Strain wBol1 Reveals Recent Horizontal Gene Transfers from Diverse Sources. BMC Genom. 2013, 14, 20. [Google Scholar] [CrossRef] [PubMed]
- Frentiu, F.D.; Robinson, J.; Young, P.R.; McGraw, E.A.; O’Neill, S.L. Wolbachia-Mediated Resistance to Dengue Virus Infection and Death at the Cellular Level. PLoS ONE 2010, 5, e13398. [Google Scholar] [CrossRef]
- Machkovech, H.M.; Bedford, T.; Suchard, M.A.; Bloom, J.D. Positive Selection in CD8+ T-Cell Epitopes of Influenza Virus Nucleoprotein Revealed by a Comparative Analysis of Human and Swine Viral Lineages. J. Virol. 2013, 87, 7066–7078. [Google Scholar] [CrossRef]
- Pan American Health Organization. Evaluation of Innovative Strategies for Aedes aegypti Control: Challenges for their Introduction and Impact Assessment; PAHO: Washington, DC, USA, 2019. [Google Scholar]
- Audsley, M.D.; Ye, Y.H.; McGraw, E.A. The Microbiome Composition of Aedes aegypti is not Critical for Wolbachia-Mediated Inhibition of Dengue Virus. PLoS Neglected Trop. Dis. 2017, 11, e0005426. [Google Scholar] [CrossRef]
- Klasson, L.; Walker, T.; Sebaihia, M.; Sanders, M.J.; Quail, M.A.; Lord, A.; Sanders, S.; Earl, J.; O’Neill, S.L.; Thomson, N.; et al. Genome evolution of Wolbachia strain wPip from the Culex pipiens group. Mol. Biol. Evol. 2009, 26, 1877–1887. [Google Scholar] [CrossRef]
- Kambris, Z.; Blagborough, A.M.; Pinto, S.B.; Blagrove, M.S.C.; Godfray, H.C.J.; Sinden, R.E.; Sinkins, S.P. Wolbachia Stimulates Immune Gene Expression and Inhibits Plasmodium Development in Anopheles gambiae. PLoS Pathog. 2010, 6, e1001143. [Google Scholar] [CrossRef]
- Chen, H.; Zhang, M.; Hochstrasser, M. The Biochemistry of Cytoplasmic Incompatibility Caused by Endosymbiotic Bacteria. Genes 2020, 8, 852. [Google Scholar] [CrossRef]
- Pan, X.; Zhou, G.; Wu, J.; Bian, G.; Lu, P.; Raikhel, A.S.; Xi, Z. Wolbachia Induces Reactive Oxygen Species-Dependent Activation of the Toll Pathway to Control Dengue Virus in the Mosquito Aedes aegypti. Proc. Natl. Acad. Sci. USA 2012, 109, E23–E31. [Google Scholar] [CrossRef] [PubMed]
- Bordenstein, S.R.; Werren, J.H. Bidirectional Incompatibility among divergent Wolbachia and incompatibility level differences among closely related Wolbachia in Nasonia. Heredity 2007, 99, 278–287. [Google Scholar] [CrossRef]
- Crawford, J.E.; Clarke, D.W.; Criswell, V.; Desnoyer, M.; Cornel, D.; Deegan, B.; Gong, K.; Hopkins, K.C.; Howell, P.; Hyde, J.S.; et al. Efficient production of male Wolbachia-infected Aedes aegypti mosquitoes enables large-scale suppression of wild populations. Nat. Biotechnol. 2020, 38, 482–492. [Google Scholar] [CrossRef] [PubMed]
- Duron, O.; Hurst, G.D. Arthropods and inherited bacteria: From counting the symbionts to understanding how symbionts count. BMC Biol. 2013, 11, 45. [Google Scholar] [CrossRef]
- Hilgenboecker, K.; Hammerstein, P.; Schlattmann, P.; Telschow, A.; Werren, J.H. How many species are infected with Wolbachia?–A statistical analysis of current data. FEMS Microbiol. Lett. 2008, 281, 215–220. [Google Scholar] [CrossRef]
- Gerth, M.; Bleidorn, C. Comparative genomics provides a timeframe for Wolbachia evolution and exposes a recent biotin synthesis operon transfer. Nat. Microbiol. 2016, 2, 16241. [Google Scholar] [CrossRef]
- Harris, H.L.; Brennan, L.J.; Keddie, B.A.; Braig, H.R. Bacterial symbionts in insects: Balancing life and death. Symbiosis 2010, 51, 37–53. [Google Scholar] [CrossRef]
- Sacchi, L.; Genchi, M.; Clementi, E.; Negri, I.; Alma, A.; Ohler, S.; Sassera, D.; Bourtzis, K.; Bandi, C. Bacteriocyte-like cells harbour Wolbachia in the ovary of Drosophila melanogaster (Insecta, Diptera) and Zyginidia pullula (Insecta, Hemiptera). Tissue Cell 2010, 42, 328–333. [Google Scholar] [CrossRef] [PubMed]
- de Oliveira, C.D.; Goncalves, D.S.; Baton, L.A.; Shimabukuro, P.H.; Carvalho, F.D.; Moreira, L.A. Broader prevalence of Wolbachia in insects including potential human disease vectors. Bull. Entomol. Res. 2015, 105, 305–315. [Google Scholar] [CrossRef] [PubMed]
- Kriesner, P.; Conner, W.R.; Weeks, A.R.; Turelli, M.; Hoffmann, A.A. Persistence of a Wolbachia infection frequency cline in Drosophila melanogaster and the possible role of reproductive dormancy. Evolution 2016, 70, 979–997. [Google Scholar] [CrossRef] [PubMed]
- Zhang, K.J.; Han, X.; Hong, X.Y. Various infection status and molecular evidence for horizontal transmission and recombination of Wolbachia and Cardinium among rice planthoppers and related species. Insect Sci. 2013, 20, 329–344. [Google Scholar] [CrossRef]
- Huigens, M.E.; de Almeida, R.P.; Boons, P.A.H.; Luck, R.F.; Stouthamer, R. Natural interspecifc and intraspecifc horizontal transfer of parthenogenesisinducing Wolbachia in Trichogramma wasps. Proc. R. Soc. Lond. B Biol. Sci. 2004, 271, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Gehrer, L.; Vorburger, C. Parasitoids as vectors of facultative bacterial endosymbionts in aphids. Biol. Lett. 2012, 8, 613–615. [Google Scholar] [CrossRef] [PubMed]
- Wang, Z.Y.; Deng, C.; Yun, Y.L.; Jian, C.; Peng, Y. Molecular detection and the phylogenetics of Wolbachia in Chinese spiders (Araneae). J. Arachnol. 2010, 38, 237–241. [Google Scholar] [CrossRef]
- Goya, L.F.; Lanteri, A.A.; Confalonieri, V.A.; Rodriguero, M.S. New host-parasitoid interactions in Naupactus cervinus (Coleoptera, Curculionidae) raise the question of Wolbachia horizontal transmission. Symbiosis 2022, 86, 325–336. [Google Scholar] [CrossRef]
- Schuler, H.; Bertheau, C.; Egan, S.P.; Feder, J.L.; Riegler, M.; Schlick-Steiner, B.C.; Steiner, F.M.; Johannesen, J.; Kern, P.; Tuba, K.; et al. Evidence for a recent horizontal transmission and spatial spread of Wolbachia from endemic Rhagoletis cerasi (Diptera: Tephritidae) to invasive Rhagoletis cingulata in Europe. Mol. Ecol. 2013, 22, 4101–4111. [Google Scholar] [CrossRef]
- Serbina, L.S.; Gajski, D.; Malenovsky, I.; Corretto, E.; Schuler, H.; Dittmer, J. Wolbachia infection dynamics in a natural population of the pear psyllid Cacopsylla pyri (Hemiptera: Psylloidea) across its seasonal generations. Sci. Rep. 2022, 12, 16502. [Google Scholar] [CrossRef] [PubMed]
- Shaikevich, E.; Bogacheva, A.; Rakova, V.; Ganushkina, L.; Ilinsky, Y. Wolbachia symbionts in mosquitoes: Intra- and intersupergroup recombinations, horizontal transmission and evolution. Mol. Phylogenet. Evol. 2019, 134, 24–34. [Google Scholar] [CrossRef]
- Su, Q.H.; Hu, G.W.; Yun, Y.L.; Peng, Y. Horizontal transmission of Wolbachia in Hylyphantes graminicola is more likely via intraspecies than interspecies transfer. Symbiosis 2019, 79, 123–128. [Google Scholar] [CrossRef]
- Lequime, S.; Paul, R.E.; Lambrechts, L. Determinants of arbovirus vertical transmission in mosquitoes. PLoS Pathog. 2016, 12, e1005548. [Google Scholar] [CrossRef]
- Fast, E.M.; Toomey, M.E.; Panaram, K.; Desjardins, D.; Kolaczyk, E.D.; Frydman, H.M. Wolbachia enhance Drosophila stem cell proliferation and target the germline stem cell niche. Science 2011, 334, 990–992. [Google Scholar] [CrossRef]
- Newton, I.L.; Savytskyy, O.; Sheehan, K.B. Wolbachia utilize host actin for efcient maternal transmission in Drosophila melanogaster. PLoS Pathog. 2015, 11, e1004798. [Google Scholar] [CrossRef]
- Bordenstein, S.R.; Rosengaus, R.B. Discovery of a novel Wolbachia Supergroup in Isoptera. Curr. Microbiol. 2005, 51, 393–398. [Google Scholar] [CrossRef]
- Zhou, W.; Rousset, F.; O’Neill, S. Phylogeny and PCR-based classification of Wolbachia strains using wsp gene sequences. Proc. R. Soc. Lond. Ser. B Biol. Sci. 1998, 265, 509–515. [Google Scholar] [CrossRef] [PubMed]
- Jiggins, F.M.; Schulenburg, J.H.G.; Hurst, G.D.D.; Majerus, M.E.N. Recombination confounds interpretations of Wolbachia evolution. Proc. R. Soc. Lond. Ser. B Biol. Sci. 2001, 268, 14231427. [Google Scholar] [CrossRef]
- Werren, J.H.; Bartos, J.D. Recombination in Wolbachia. Curr. Biol. 2001, 11, 431–435. [Google Scholar] [CrossRef] [PubMed]
- Bandi, C.; Bourtzis, K.; Braig, H.; Lo, N.; O’Neill, S.; Stouthamer, R.; Vandekerckove, T.T.M.; Werren, J.H. Wolbachia Nomenclatural System: A Discussion Paper. News and Forums. 2003. Available online: www.wolbachia.sols.uq.edu.au/news.cfm?action=thread&id=25 (accessed on 14 February 2024).
- O’Neill, S.L.; Hoffmann, A.A.; Werren, J.H. Influential Passengers: Inherited Microorganisms and Arthropod Reproduction and Development; Oxford University Press: New York, NY, USA, 1997. [Google Scholar]
- Baldo, L.; Dunning Hotopp, J.C.; Jolley, K.A.; Bordenstein, S.R.; Biber, S.A.; Choudhury, R.R.; Hayashi, C.; Maiden, M.C.J.; Tettelin, H.; Werren, J.H. Multilocus sequence typing system for the endosymbiont Wolbachia pipientis. Appl. Environ. Microbiol. 2006, 72, 7098–7110. [Google Scholar] [CrossRef] [PubMed]
- Bleidorn, C.; Gerth, M. A critical re-evaluation of multilocus sequence typing (MLST) efforts in Wolbachia. FEMS Microbiol. Ecol. 2017, 94, fix163. [Google Scholar] [CrossRef] [PubMed]
- Hotopp, J.C.D.; Slatko, B.E.; Foster, J.M. Targeted enrichment and sequencing of recent endosymbiont-host lateral gene transfers. Sci. Rep. 2017, 7, 857. [Google Scholar] [CrossRef] [PubMed]
- Kent, B.N.; Bordenstein, S.R. Phage WO of Wolbachia: Lambda of the endosymbiont world. Trends Microbiol. 2010, 18, 173–181. [Google Scholar] [CrossRef] [PubMed]
- Wang, M.; Beck, C.R.; English, A.C.; Meng, Q.; Buhay, C.; Han, Y.; Doddapaneni, H.V.; Yu, F.; Boerwinkle, E.; Lupski, J.R.; et al. PacBio-LITS: A large-insert targeted sequencing method for characterization of human disease-associated chromosomal structural variations. BMC Genom. 2015, 16, 214. [Google Scholar] [CrossRef] [PubMed]
- McGraw, E.A.; Martin, W.; Esser, C.; Ahmadinejad, N.; Wu, M.; Sun, L.V.; Vamathevan, J.; Riegler, M.; Deboy, R.; Brownlie, J.C. Phylogenomics of the reproductive parasite Wolbachia pipientis wMel: A streamlined genome overrun by mobile genetic elements. PLoS Biol. 2004, 2, E69. [Google Scholar]
- Mavingui, P.; Valiente Moro, C.V.; Tran-Van, V.; Wisniewski-Dye, F.; Raquin, V.; Minard, G.; Tran, F.H.; Voronin, D.; Rouy, Z.; Bustos, P.; et al. Whole-genome sequence of Wolbachia strain wALbB, an endosymbiont of tiger mosquito vector Aedes albopictus. J. Bacteriol. 2012, 194, 18–40. [Google Scholar] [CrossRef] [PubMed]
- Klasson, L.; Westberg, J.; Sapountzis, P.; Naslund, K.; Lutnaes, Y.; Darby, E.A.C.; Veneti, Z.; Chen, L.; Braig, H.R.; Garrett, R.; et al. The mosaicgenome structure of the Wolbachia wRi strain infecting Drosophila simulans. Proc. Natl. Acad. Sci. USA 2009, 106, 5725–5730. [Google Scholar] [CrossRef] [PubMed]
- Foster, J.; Ganatra, M.; Kamal, I.; Ware, J.; Makarova, K.; Ivanova, N.; Bhattacharyya, A.; Kapatral, V.; Kumar, S.; Posfai, J.; et al. The Wolbachia genome of Brugia malayi: Endosymbiont evolution within a human pathogenic nematode. PLoS Biol. 2005, 3, e121. [Google Scholar] [CrossRef] [PubMed]
- Ishmael, N.; Hotopp, J.C.D.; Ioannidis, P.; Biber, S.; Sakamoto, J.; Siozios, S.; Nene, V.; Werren, J.; Bourtzis, K.; Bordenstein, S.R.; et al. Extensive genomic diversity of closely related Wolbachia strains. Microbiology 2009, 155, 2211–2222. [Google Scholar] [CrossRef] [PubMed]
- Perlmutter, J.I.; Bordenstein, S.R.; Unckless, R.L.; LePage, D.P.; Metcalf, J.A.; Hill, T.; Martinez, J.; Jiggins, F.M.; Bordenstein, S.R. The phage gene wmk is a candidate for male killing by a bacterial endosymbiont. PLoS Pathog. 2019, 15, e1007936. [Google Scholar] [CrossRef]
- Rice, D.W.; Sheehan, K.B.; Newton, I.L.G. Large-scale identification of Wolbachia pipientis effectors. Genome Biol. Evol. 2017, 9, 1925–1937. [Google Scholar] [CrossRef]
- Scholz, M.; Albanese, D.; Tuohy, K.; Donati, C.; Segata, N.; Rota-Stabelli, O. Large scale genome reconstructions illuminate Wolbachia evolution. Nat. Commun. 2020, 11, 5235. [Google Scholar] [CrossRef]
- Xi, Z.; Khoo, C.C.; Dobson, S.L. Wolbachia establishment and invasion in an Aedes aegypti laboratory population. Science 2005, 310, 326–328. [Google Scholar] [CrossRef] [PubMed]
- Walker, T.; Johnson, P.H.; Moreira, L.A.; Iturbe-Ormaetxe, I.; Frentiu, F.D.; McMeniman, C.J.; Leong, Y.S.; Dong, Y.; Axford, J.; Kriesner, P.; et al. The wMel Wolbachia strain blocks dengue and invades caged Aedes aegypti populations. Nature 2011, 476, 450–453. [Google Scholar] [CrossRef]
- Moreira, L.A.; Iturbe-Ormaetxe, I.; Jeffery, J.A.; Lu, G.; Pyke, A.T.; Hedges, L.M.; Rocha, B.C.; Hall-Mendelin, S.; Day, A.; Riegler, M.; et al. A Wolbachia symbiont in Aedes aegypti limits infection with dengue, Chikungunya, and Plasmodium. Cell 2009, 139, 1268–1278. [Google Scholar] [CrossRef] [PubMed]
- Marinotti, O.; de Brito, M.; Moreira, C.K. Apyrase and alpha-glucosidase in the salivary glands of Aedes albopictus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 1996, 113, 675–679. [Google Scholar] [CrossRef]
- Kremer, N.; Voronin, D.; Charif, D.; Mavingui, P.; Mollereau, B.; Vavre, F. Wolbachia interferes with ferritin expression and iron metabolism in insects. PLoS Pathog. 2009, 5, e1000630. [Google Scholar] [CrossRef] [PubMed]
- Ross, P.A.; Wiwatanaratanabutr, I.; Axford, J.K.; White, V.L.; EndersbyHarshman, N.M.; Hofmann, A.A. Wolbachia infections in Aedes aegypti difer markedly in their response to cyclical heat stress. PLoS Pathog. 2017, 13, e1006006. [Google Scholar] [CrossRef] [PubMed]
- Ye, Y.H.; Carrasco, A.M.; Dong, Y.; Sgró, C.M.; Mcgraw, E.A. The effect of temperature on Wolbachia-mediated dengue virus blocking in Aedes aegypti. Am. J. Trop. Med. Hyg. 2016, 94, 812. [Google Scholar] [CrossRef] [PubMed]
- Van Den Hurk, A.F.; Hall-Mendelin, S.; Pyke, A.T.; Frentiu, F.; Mcelroy, K.; Day, A.; Higgs, S.; O’neill, S.L. Impact of Wolbachia on infection with chikungunya and yellow fever viruses in the mosquito vector Aedes aegypti. PLOS Negl. Trop. Dis. 2012, 6, e1892. [Google Scholar] [CrossRef]
- Hussain, M.; Frentiu, F.D.; Moreira, L.A.; O’Neill, S.L.; Asgari, S. Wolbachia uses host MicroRNAs to manipulate host gene expression and facilitate colonization of the dengue vector Aedes aegypti. Proc. Natl. Acad. Sci. USA 2011, 108, 9250–9255. [Google Scholar] [CrossRef]
- Moreira, L.A.; Ye, H.Y.; Turner, K.; Eyles, D.W.; Mcgraw, E.A.; O’neill, S.L. The wMelPop strain of Wolbachia interferes with dopamine levels in Aedes aegypti. Parasit. Vectors 2011, 4, 28. [Google Scholar] [CrossRef]
- Caragata, E.P.; Dutra, H.L.C.; Moreira, L.A. Inhibition of Zika virus by Wolbachia in Aedes aegypti. Microbial Cell 2016, 3, 293–295. [Google Scholar] [CrossRef]
- Pietri, J.E.; Debruhl, H.; Sullivan, W. The rich somatic life of Wolbachia. Microbiol. Open. 2016, 5, 923. [Google Scholar] [CrossRef]
- Kamtchum-Tatuene, J.; Makepeace, B.L.; Benjamin, L.; Baylis, M.; Solomon, T. The potential role of Wolbachia in controlling the transmission of emerging human arboviral infections. Curr. Opin. Infect. Dis. 2017, 30, 108–116. [Google Scholar] [CrossRef]
- Caragata, E.P.; Pais, F.S.; Baton, L.A.; Silva, J.B.L.; Sorgine, M.H.F.; Moreira, L.A. The transcriptome of the mosquito Aedes fuviatilis (Diptera: Culicidae), and transcriptional changes associated with its native Wolbachia infection. BMC Genom. 2017, 18, 6. [Google Scholar] [CrossRef]
- Rancés, E.; Yh, Y.E.; Woolft, M.; Mcgraw, E.A.; O’neill, S.L. The relative importance of innate immune priming in Wolbachiamediated dengue. Interf. PLoS Pathog. 2012, 8, e1002548. [Google Scholar]
- Koh, C.; Islam, M.N.; Ye, Y.H.; Chotiwan, N.; Graham, B.; Belisle, J.T.; Kouremenos, K.A.; Dayalan, S.; Tull, D.L.; Klatt, S.; et al. Dengue virus dominates lipid metabolismo modulations in Wolbachia coinfected Aedes aegypti. Commun. Biol. 2020, 3, 518. [Google Scholar] [CrossRef] [PubMed]
- Martins, M.; Ramos, L.F.C.; Murillo, J.R.; Torres, A.; de Carvalho, S.S.; Domont, G.B.; de Oliveira, D.M.P.; Mesquita, R.D.; Nogueira, F.C.S.; Maciel-de-Freitas, R.; et al. Comprehensive quantitative proteome analysis of Aedes aegypti identifies proteins and pathways involved in wolbachia pipientis and zika virus interference phenomenon. Front. Physiol. 2021, 12, 642237. [Google Scholar] [CrossRef] [PubMed]
- Ogunlade, S.T.; Meehan, M.T.; Adekunle, A.I.; Rojas, D.P.; Adegboye, O.A.; McBryde, E.S. A review: Aedes-borne arboviral infections, controls and Wolbachia-based strategies. Vaccines 2021, 9, 32. [Google Scholar] [CrossRef] [PubMed]
- Ulrich, J.N.; Beier, J.C.; Devine, G.J.; Hugo, L.E. Heat Sensitivity of wMelWolbachia during Aedes aegypti development. PLOS Negl. Trop. Dis. 2016, 10, e0004873. [Google Scholar] [CrossRef] [PubMed]
- Lambrechts, L.; Ferguson, N.M.; Harris, E.; Holmes, E.C.; Mcgraw, E.A.; O’neill, S.L.; Ooi, O.O.; Ritchie, A.S.; Ryan, P.A.; Scott, T.W.; et al. Assessing the epidemiological impact of Wolbachia deployment for dengue control. Lancet Infect. Dis. 2015, 15, 862–866. [Google Scholar] [CrossRef] [PubMed]
- Chang, M.S.; Christophel, E.M.; Gopinath, D.; Abdur, R.M. Challenges and future perspective for dengue, vector control in the Western Pacifc Region. WPSAR 2011, 2, e1. [Google Scholar] [CrossRef] [PubMed]
- Bull, J.; Turelli, M. Wolbachia versus dengue evolutionary forecasts. Evol. Med. Public Health 2013, 2013, 197–207. [Google Scholar] [CrossRef]
- Roe, A.D.; Weller, S.J.; Baixeras, J.; Brown, J.W.; Cummings, M.P.; Davis, D.; Kawahara, A.; Parr, C.; Regier, J.; Rubinoff, D. Evolutionary framework for Lepidoptera model systems. In Genetics and Molecular Biology of Lepidoptera, Contemporary Topics in Entomology; Goldsmith, M., Marec, F., Eds.; CRC Press: Gainesville, FL, USA, 2009. [Google Scholar]
- Feener, D.H., Jr.; Brown, B.V. Diptera as parasitoids. Annu. Rev. Entomol. 1997, 42, 73–97. [Google Scholar] [CrossRef]
- Pennacchio, F.; Strand, M.R. Evolution of developmental strategies in parasitic hymenoptera. Annu. Rev. Entomol. 2006, 51, 233–258. [Google Scholar] [CrossRef]
- Narita, S.; Shimajiri, Y.; Nomura, M. Strong cytoplasmic incompatibility and high vertical transmission rate can explain the high frequencies of Wolbachia infection in Japanese populations of Colias erate poliographus (Lepidoptera: Pieridae). Bull. Entomol. Res. 2009, 99, 385–391. [Google Scholar] [CrossRef] [PubMed]
- Dyson, E.A.; Hurst, G.D.D. Persistence of an extreme sex-ratio bias in a natural population. Proc. Natl. Acad. Sci. USA 2004, 101, 6520–6523. [Google Scholar] [CrossRef]
- Sasaki, T.; Ishikawa, H. Wolbachia infections and cytoplasmic incompatibility in the almond moth and the mediterranean flour moth. Zoo. Sci. 1999, 16, 739–744. [Google Scholar] [CrossRef]
- Kageyama, D.; Nishimura, G.; Hoshizaki, S.; Ishikawa, Y. Feminizing Wolbachia in an insect, Ostrinia furnacalis (Lepidoptera: Crambidae). Heredity 2002, 88, 444–449. [Google Scholar] [CrossRef] [PubMed]
- Kaiser, W.; Huguet, E.; Casas, J.; Commin, C.; Giron, D. Plant green-island phenotype induced by leaf-miners is mediated by bacterial symbionts. Proc. R. Soc. B-Biol. Sci. 2010, 277, 2311–2319. [Google Scholar] [CrossRef]
- Graham, R.I.; Grzywacz, D.; Mushobozi, W.L.; Wilson, K. Wolbachia in a major African crop pest increases susceptibility to viral disease rather than protects. Ecol. Lett. 2012, 15, 9931000. [Google Scholar] [CrossRef]
- Werren, J.H.; Windsor, D.; Guo, L.R. Distribution of Wolbachia among Neotropical Arthropods. Proc. R. Soc. B-Biol. Sci. 1995, 1364, 197–204. [Google Scholar]
- Werren, J.H.; Windsor, D.M. Wolbachia infection frequencies in insects: Evidence of a global equilibrium? Proc. R. Soc. B Biol. Sci. 2000, 267, 1277–1285. [Google Scholar] [CrossRef]
- West, S.A.; Cook, J.M.; Werren, J.H.; Godfray, H.C.J. Wolbachia in two insect hostparasitoid communities. Mol. Ecol. 1998, 7, 1457–1465. [Google Scholar] [CrossRef]
- Tagami, Y.; Miura, K. Distribution and prevalence of Wolbachia in Japanese populations of Lepidoptera. Insect Mol. Biol. 2004, 13, 359–364. [Google Scholar] [CrossRef]
- Jiggins, F.M.; Hurst, G.D.D.; Schulenburg, J.; Majerus, M.E.N. Two male-killing Wolbachia strains coexist within a population of the butterfly Acraea encedon. Heredity 2001, 86, 161–166. [Google Scholar] [CrossRef] [PubMed]
- Salunke, B.K.; Salunkhe, R.C.; Dhotre, D.P.; Walujkar, S.A.; Khandagale, A.B.; Chaudhari, R.; Chandode, R.K.; Ghate, H.V.; Patole, M.S.; Werren, J.H.; et al. Determination of Wolbachia diversity in butterflies from western Ghats, India, by a multigene approach. Appl. Environ. Microbiol. 2012, 78, 4458–4467. [Google Scholar] [CrossRef]
- Duron, O.; Bouchon, D.; Boutin, S.; Bellamy, L.; Zhou, L.; Engelstaedter, J.; Hurst, G.D. The diversity of reproductive parasites among arthropods: Wolbachia do not walk alone. BMC Biology 2008, 6, 27. [Google Scholar] [CrossRef]
- Toju, H.; Fukatsu, T. Diversity and infection prevalence of endosymbionts in natural populations of the chestnut weevil: Relevance of local climate and host plants. Mol. Ecol. 2011, 20, 853–868. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Miao, H.; Hong, X.Y. Distribution of the endosymbiotic bacterium Cardinium in Chinese populations of the carmine spider mite Tetranychus cinnabarinus (Acari: Tetranychidae). J. Appl. Entomol. 2006, 130, 523–529. [Google Scholar] [CrossRef]
- Morag, N.; Klement, E.; Saroya, Y.; Lensky, I.; Gottlieb, Y. Prevalence of the symbiont Cardinium in Culicoides (Diptera: Ceratopogonidae) vector species is associated with land surface temperature. Am. Fasten. J. 2012, 26, 4025–4034. [Google Scholar] [CrossRef]
- Sumi, T.; Miura, K.; Miyatake, T. No seasonal trend in infection of the pale grass blue butterfly, Zizeeria maha (Lepidoptera: Lycaenidae), by Wolbachia. Appl. Entomol. Zool. 2013, 48, 35–38. [Google Scholar] [CrossRef]
- Van Opijnen, T.; Breeuwer, J.A.J. High temperatures eliminate Wolbachia, a cytoplasmic incompatibility inducing endosymbiont, from the two-spotted spider mite. Exp. Appl. Acarol. 1999, 23, 871–881. [Google Scholar] [CrossRef] [PubMed]
- Hurst, G.D.D.; Jiggins, F.M.; Robinson, S.J.W. What causes inefficient transmission of male-killing Wolbachia in Drosophila? Heredity 2001, 87, 220–226. [Google Scholar] [CrossRef] [PubMed]
- Jia, F.X.; Yang, M.S.; Yang, W.J.; Wang, J.J. Influence of Continuous High Temperature Conditions on Wolbachia Infection Frequency and the Fitness of Liposcelis tricolor (Psocoptera: Liposcelididae). Environ. Entomol. 2009, 38, 1365–1372. [Google Scholar] [CrossRef] [PubMed]
- Guruprasad, N.M.; Mouton, L.; Puttaraju, H.P. Effect of Wolbachia infection and temperature variations on the fecundity of the Uzifly Exorista sorbillans (Diptera: Tachinidae). Symbiosis 2011, 54, 151–158. [Google Scholar] [CrossRef]
- LePage, D.P.; Metcalf, J.A.; Bordenstein, S.R.; On, J.; Perlmutter, J.I.; Shropshire, J.D.; Layton, E.M.; Funkhouser-Jones, L.J.; Beckmann, J.F.; Bordenstein, S.R. Prophage WO genes recapitulate and enhance Wolbachia-induced cytoplasmic incompatibility. Nature 2017, 543, 243–247. [Google Scholar] [CrossRef] [PubMed]
- Beckmann, J.F.; Ronau, J.A.; Hochstrasser, M.A. Wolbachia deubiquitylating enzyme induces cytoplasmic incompatibility. Nat. Microbiol. 2017, 2, 17007. [Google Scholar] [CrossRef] [PubMed]
- Nikolouli, K.; Colinet, H.; Renault, D.; Enriquez, T.; Mouton, L.; Gibert, P.; Sassu, F.; Cáceres, C.; Stauffer, C.; Pereira, R.; et al. Sterile insect technique and Wolbachia symbiosis as potential tools for the control of the invasive species Drosophila suzukii. J. Pest. Sci. 2018, 91, 489–503. [Google Scholar] [CrossRef]
- Fukui, T.; Kawamoto, M.; Shoji, K.; Kiuchi, T.; Sugano, S.; Shimada, T.; Suzuki, Y.; Katsuma, S. The endosymbiotic bacterium Wolbachia selectively kills male hosts by targeting the masculinizing gene. PLoS Pathog. 2015, 11, e1005048. [Google Scholar] [CrossRef]
- Sugimoto, T.N.; Ishikawa, Y.A. male-killing Wolbachia carries a feminizing factor and is associated with degradation of the sexdetermining system of its host. Biol. Lett. 2012, 8, 412–415. [Google Scholar] [CrossRef]
- Charlat, S.; Davies, N.; Roderick, G.K.; Hurst, G.D.D. Disrupting the timing of Wolbachia-induced male-killing. Biol. Lett. 2007, 3, 154–156. [Google Scholar] [CrossRef] [PubMed]
- Arai, H.; Hirano, T.; Akizuki, N.; Abe, A.; Nakai, M.; Kunimi, Y.; Inoue, M.N. Multiple infection and reproductive manipulations of Wolbachia in Homona magnanima (Lepidoptera: Tortricidae). Micro. Ecol. 2019, 77, 257–266. [Google Scholar] [CrossRef]
- Katsuma, S.; Hirota, K.; Matsuda-Imai, N.; Fukui, T.; Muro, T.; Nishino, K.; Kosako, H.; Shoji, K.; Takanashi, H.; Fujii, T.; et al. A Wolbachia factor for male killing in lepidopteran Insects. Nat. Commun. 2022, 13, 6764. [Google Scholar] [CrossRef]
- García Morales, M.; Denno, B.D.; Miller, D.R.; Miller, G.L.; Ben-Dov, Y.; Hardy, N.B. ScaleNet: A literature-based model of scale insect biology and systematics. Database 2016, 2016, bav118. [Google Scholar] [CrossRef]
- Gullan, P.J.; Cook, L.G. Phylogeny and higher classification of the scale insects (Hemiptera: Sternorrhyncha: Coccoidea). Zootaxa 2007, 1668, 413–425. [Google Scholar] [CrossRef]
- Kondo, T.; Gullan, P.J.; Williams, D.J. Coccidology. The study of scale insects (Hemiptera: Sternorrhyncha: Coccoidea). Corpoica. Cienc. Y Tecnol. Agropecu. 2008, 9, 55–61. [Google Scholar] [CrossRef]
- Buckley, R.; Gullan, P. More aggressive ant species (Hymenoptera: Formicidae) provide better protection for soft scales and mealybugs (Homoptera: Coccidae, Pseudococcidae). Biotropica 1991, 23, 282–286. [Google Scholar] [CrossRef]
- Gullan, P.J.; Buckley, R.C.; Ward, P.S. Ant-tended scale insects (Hemiptera: Coccidae: Myzolecanium) within lowland rain Forest trees in Papua New Guinea. J. Trop. Ecol. 1993, 9, 81–91. [Google Scholar] [CrossRef]
- Johanowicz, D.L.; Hoy, M.A. Wolbachia in a predator–prey system: 16 S ribosomal Dna analysis of two Phytoseiids (Acari: Phytoseiidae) and their prey (Acari: Tetranychidae). Ann. Entomol. Soc. Am. 1996, 89, 435–441. [Google Scholar] [CrossRef]
- Kageyama, D.; Narita, S.; Imamura, T.; Miyanoshita, A. Detection and identification of Wolbachia endosymbionts from laboratory stocks of stored-product insect pests and their parasitoids. J. Stored Prod. Res. 2010, 46, 13–19. [Google Scholar] [CrossRef]
- Tzuri, N.; Caspi-Fluger, A.; Betelman, K.; Rohkin Shalom, S.; Chiel, E. Horizontal transmission of microbial symbionts within a Guild of fly Parasitoids. Microb. Ecol. 2020, 81, 818–827. [Google Scholar] [CrossRef] [PubMed]
- Ramalho, M.O.; Moreau, C.S. The evolution and biogeography of Wolbachia in ants (Hymenoptera: Formicidae). Diversity 2020, 12, 426. [Google Scholar] [CrossRef]
- Ivens, A.B.F.; Gadau, A.; Kiers, E.T.; Kronauer, D.J.C. Can social partnerships influence the microbiome? Insights from ant farmers and their trophobiont mutualists. Mol. Ecol. 2018, 27, 1898–1914. [Google Scholar] [CrossRef]
- Pringle, E.G.; Moreau, C.S. Community analysis of microbial sharing and specialization in a Costa Rican ant–plant–hemipteran symbiosis. Proc. R. Soc. B 2017, 284, 20162770. [Google Scholar] [CrossRef]
- Gruwell, M.E.; Wu, J.; Normark, B.B. Diversity and phylogeny of Cardinium (Bacteroidetes) in armored scale insects (Hemiptera: Diaspididae). Ann. Entomol. Soc. Am. 2009, 102, 1050–1061. [Google Scholar] [CrossRef]
- Sanaei, E.; Lin, Y.P.; Cook, L.G.; Engelstädter, J. Wolbachia in scale insects: A distinct pattern of infection frequencies and potential transfer routes via ant associates. Environ. Microbiol. 2021, 23, 457441. [Google Scholar] [CrossRef]
- Sanaei, E.; Albery, G.; Yeoh, Y.K.; Lin, Y.P.; Cook, L.; Engelstädter, J. Host phylogeny and ecological associations best explain Wolbachia host shifts in scale insects. Mol. Ecol. 2023, 32, 2351–2363. [Google Scholar] [CrossRef]
- Hibino, H. Biology and epidemiology of rice viruses. Annu. Rev. Phytopathol. 1996, 34, 249–274. [Google Scholar] [CrossRef]
- Zhang, H.; Zhang, K.J.; Hong, X.Y. Population dynamics of noncytoplasmic incompatibility-inducing Wolbachia in Nilaparvata lugens and its effects on host adult life span and female ftness. Environ. Entomol. 2010, 39, 1801–1809. [Google Scholar] [CrossRef] [PubMed]
- Zhou, X.F.; Li, Z.X. Establishment of the cytoplasmic incompatibility-inducing Wolbachia strain wMel in an important agricultural pest insect. Sci. Rep. 2016, 6, 39200. [Google Scholar] [CrossRef] [PubMed]
- Hosseinzadeh, S.; Shams-Bakhsh, M.; Mann, M.; Fattah-Hosseini, S.; Bagheri, A.; Mehrabadi, M.; Heck, M. Distribution and variation of bacterial endosymbiont and ‘Candidatus Liberibacter asiaticus’ titer in the Huanglongbing insect vector, Diaphorina citri Kuwayama. Microb. Ecol. 2019, 78, 206–222. [Google Scholar] [CrossRef] [PubMed]
- Ren, S.L.; Li, Y.H.; Ou, D.; Guo, Y.J.; Qureshi, J.A.; Stansly, P.A.; Qiu, B.L. Localization and dynamics of Wolbachia infection in Asian citrus psyllid Diaphorina citri, the insect vector of the causal pathogens of Huanglongbing. Microbio. Open. 2018, 7, e00561. [Google Scholar] [CrossRef]
- Guo, Y.; Hoffmann, A.A.; Xu, X.Q.; Mo, P.W.; Huang, H.J.; Gong, J.T.; Ju, J.F.; Hong, X.Y. Vertical transmission of Wolbachia is associated with host vitellogenin in Laodelphax striatellus. Front. Microbiol. 2018, 9, 2016. [Google Scholar] [CrossRef]
- Guo, Y.; Khan, J.; Zheng, X.Y.; Wu, Y. Wolbachia increase germ cell mitosis to enhance the fecundity of Laodelphax striatellus. Insect Biochem. Mol. Biol. 2020, 127, 103471. [Google Scholar] [CrossRef] [PubMed]
- Lu, Y.; Shiyuan, M.; Zhengyan, W.; Sibao, W. Prevalence of Wolbachia in 10 Tenebrionidae stored-product insects and spatiotemporal infection dynamics in Tribolium confusum (Jaquelin Du Val) (Coleoptera: Tenebrionidae). Grain Oil Sci. Tech. 2019, 2, 85–90. [Google Scholar] [CrossRef]
- Ju, J.F.; Bing, X.L.; Zhao, D.S.; Guo, Y.; Xi, Z.Y.; Hoffmann, A.A.; Zhang, K.-J.; Huang, H.-J.; Gong, J.-T.; Zhang, X.; et al. Wolbachia supplement biotin and riboflavin to enhance reproduction in planthoppers. ISME J. 2020, 14, 676–687. [Google Scholar] [CrossRef] [PubMed]



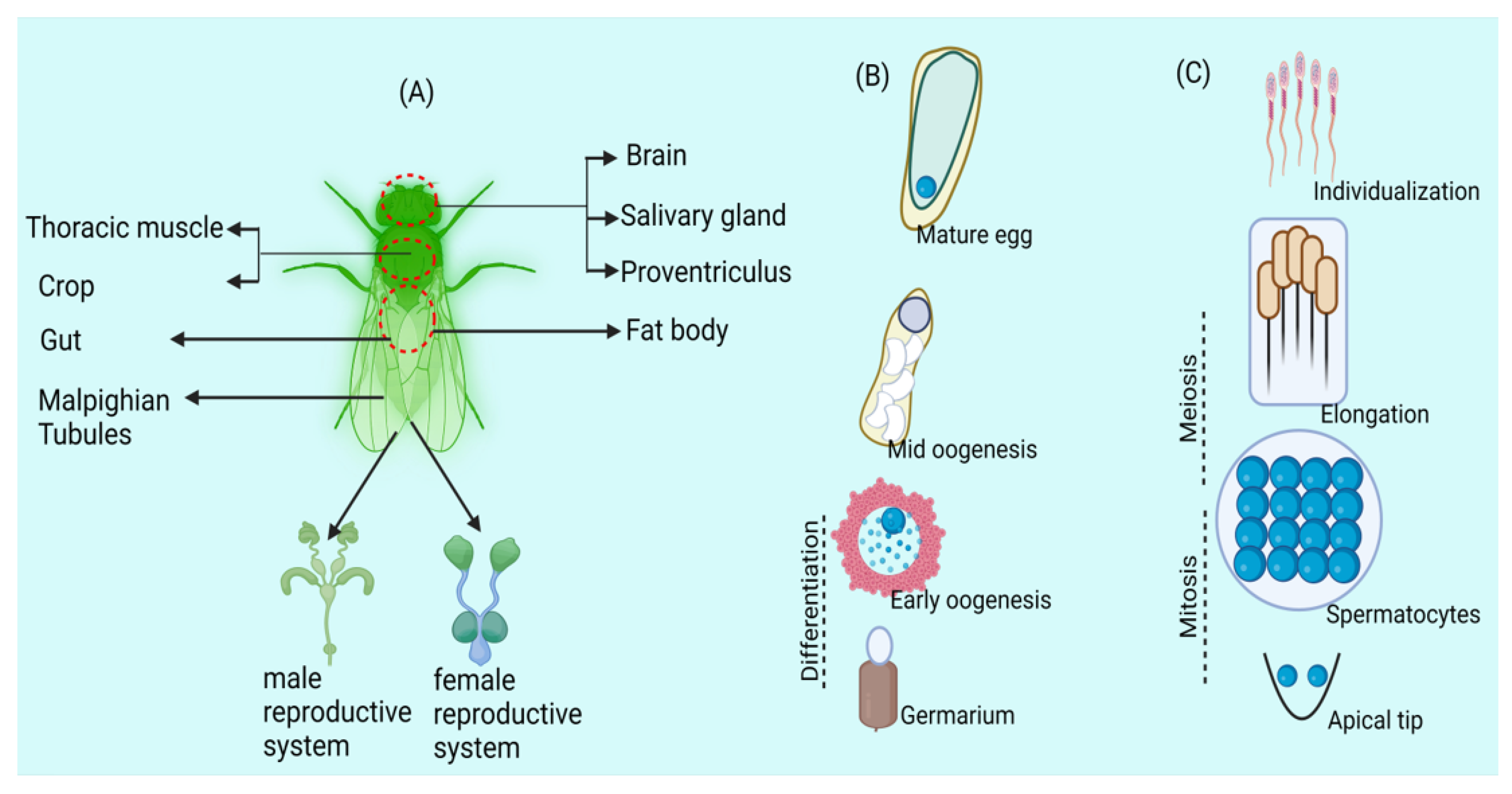{kind=link}
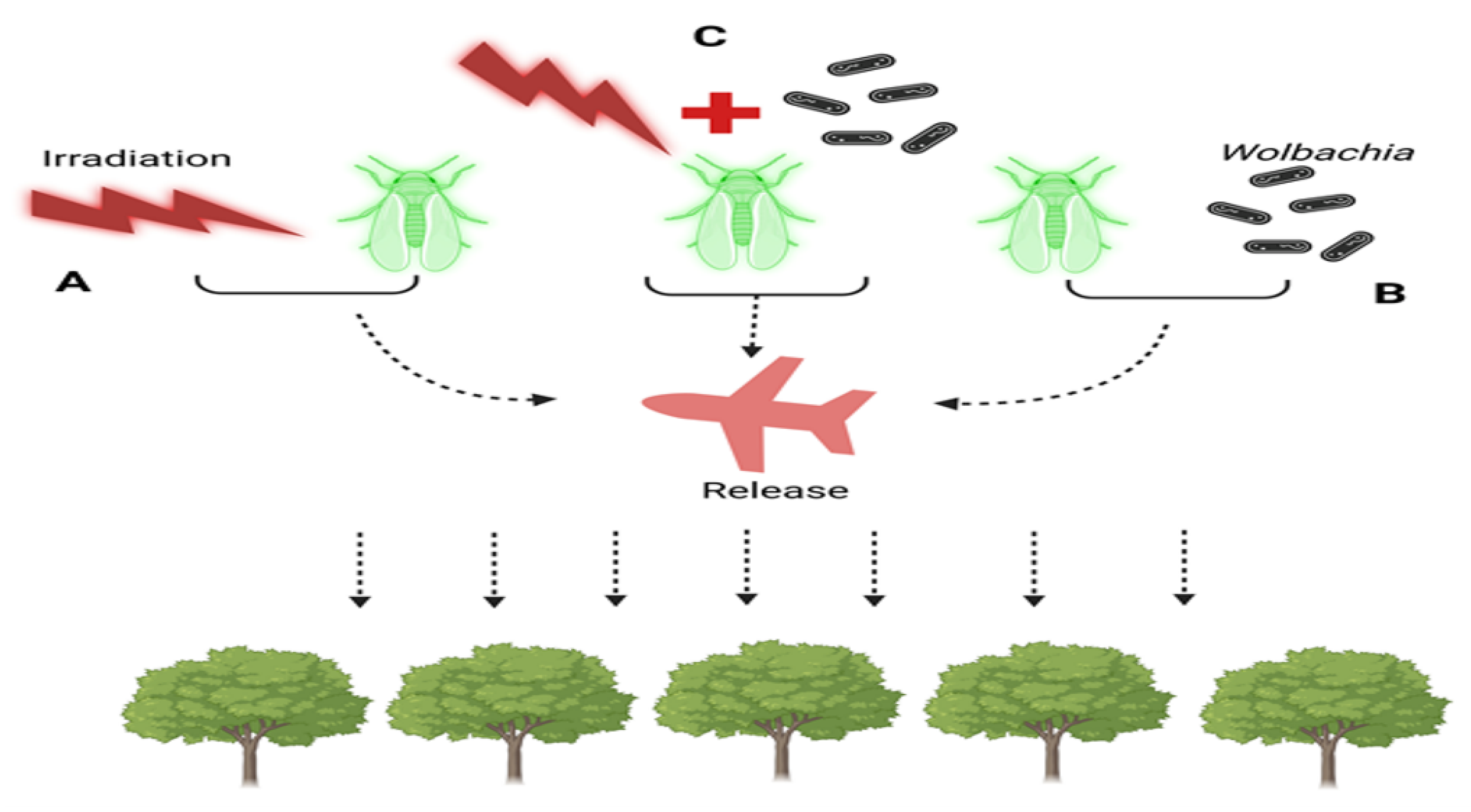{kind=link}
| Insect Name | Transmission Methods | Effect on Host | Citation |
|---|---|---|---|
| Rice Plant hoppers | Horizontal | The physiological functions of the host are impacted in observable ways by Wolbachia’s horizontal transmission, suggesting a complicated interaction between the bacterium and its host. | [73,74] |
| Wasps | Horizontal | Due to their dual function as possible immune modulators and vectors of horizontal transmission, parasitic mites present important questions in the context of predation. | [75,76] |
| Fruit Flies | Horizontal | Successful horizontal transmission and possible fitness impacts are revealed by hybridization with parasitic wasps, underscoring important dynamics in the complex host–parasite interaction. | [29,77] |
| Trypetids | Horizontal | The hybridization process with parasitoid wasps exhibits horizontal transmission and has the capacity to influence the reproductive characteristics of the host. | [78] |
| Psyllids | Horizontal | Feeding on common plants permits horizontal transmission and so offers the possibility of affecting the dynamics of host fitness. | [79] |
| Moths | Horizontal | Hybridization highlights the complex dynamics in the setting of interspecies interactions by revealing horizontal transmission and its possible implications on mating behaviour. | [24] |
| Ladybirds | Horizontal | Because predators make the spread of disease easier, possible consequences on host fitness in ecological dynamics should be taken into consideration. | [80] |
| Mites | Horizontal | Predation promotes horizontal transmission and may have an impact on the dynamics of mite populations in complex ecological interactions. | [81] |
| Various Diptera Insects | Vertical | Vertical transmission is demonstrated by transovarial transmission, which may have an impact on the kinetics of host reproduction. | [82] |
| Drosophila | Vertical | Actin-mediated transovarial transmission: Wolbachia affects host reproductive organs; comprehensive understanding of actin-mediated transmission mechanisms | [83,84] |
| Mechanism of Action | Impact on Host Physiology | Citations |
|---|---|---|
| Wolbachia exhibits obligatory reproductive parasitism | Wolbachia has a complex effect on the physiology of its host species, depending on its population density, which is highly regulated by dietary variables and ambient temperatures. | [108,109] |
| Temperature-induced changes affect cytoplasmic incompatibility (CI) capacity | Modifies the ability for cytoplasmic incompatibility (CI), resulting in a complex reaction where some males retain incompatibility while females lose this reproductive characteristic. | [110] |
| reduced life expectancy, decreased viral replication, and changed vector competency | Wolbachia has a complex effect that includes preventing virus replication, shortening mosquito lifespans, and impairing mosquito vector competence. | [56,111] |
| Reduction in saliva production affecting feeding capacity | Wolbachia reduces salivary flow, which limits mosquitoes’ ability to eat. | [112,113] |
| Provocation of changes linked to lead resistance, iron homeostasis, RNA and DNA processing, and digestive enzymes | Iron homeostasis, RNA and DNA functions, digestive enzymes, and lead resistance are all altered by Wolbachia. | [114,115,116,117,118] |
| Hypotheses on action: Competition or Modulation of lipid production | Two possible ways that Wolbachia may use to regulate lipid synthesis are competition and modulation, respectively. | [119,120] |
| Regulation of cellular autophagy and pre-activation of immune system | Wolbachia stimulates genes linked to the Toll pathway, pre-activates the immune system, and modifies cellular autophagy. | [121] |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Hyder, M.; Lodhi, A.M.; Wang, Z.; Bukero, A.; Gao, J.; Mao, R. Wolbachia Interactions with Diverse Insect Hosts: From Reproductive Modulations to Sustainable Pest Management Strategies. Biology 2024, 13, 151. https://doi.org/10.3390/biology13030151
Hyder M, Lodhi AM, Wang Z, Bukero A, Gao J, Mao R. Wolbachia Interactions with Diverse Insect Hosts: From Reproductive Modulations to Sustainable Pest Management Strategies. Biology. 2024; 13(3):151. https://doi.org/10.3390/biology13030151
Chicago/Turabian StyleHyder, Moazam, Abdul Mubeen Lodhi, Zhaohong Wang, Aslam Bukero, Jing Gao, and Runqian Mao. 2024. "Wolbachia Interactions with Diverse Insect Hosts: From Reproductive Modulations to Sustainable Pest Management Strategies" Biology 13, no. 3: 151. https://doi.org/10.3390/biology13030151