
Gene Editing of the Follicle-Stimulating Hormone Gene to Sterilize Channel Catfish, Ictalurus punctatus, Using a Modified Transcription Activator-like Effector Nuclease Technology with Electroporation

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. TALEN Plasmid Construction and Preparation
2.2. Broodstock and Gamete Preparation
2.3. Mutant Detection amd Plasmid Integration Inspection for P1 Fish
2.4. Fish Culture and Mutant Detection for F1 Fish
2.5. Reproductive Evaluation for P1 fish
2.6. Hormone Therapy for P1 fish
2.7. Pleiotropic Effects Evaluation
2.8. Statistical Analysis
3. Results
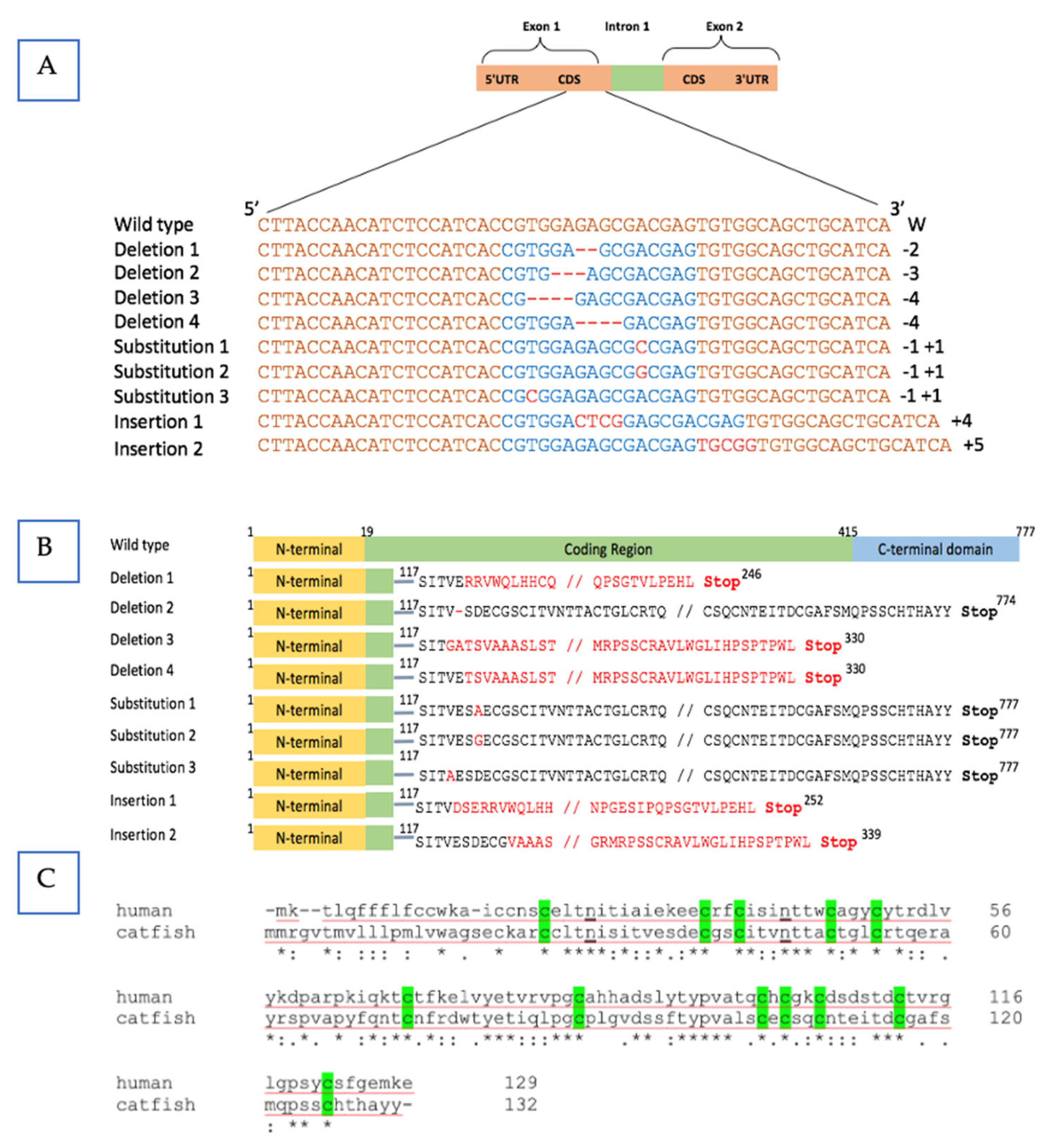
3.1. Mutation Evaluation and Plasmid Integration in the P1 Fish
3.2. Fertility Evaluation and Hormone Treatment and Intervention
3.3. Mutation Evaluation for the F1 Fish
3.4. Pleiotropic Effects
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Dunham, R.A.; Ramboux, A.C.; Duncan, P.L.; Hayat, M.; Chen, T.T.; Lin, C.M.; Kight, K.; Gonzalez-Villasenor, I.; Powers, D.A. Transfer, Expression, and Inheritance of Salmonid Growth Hormone Genes in Channel Catfish, Ictalurus punctatus, and Effects on Performance Traits. Mol. Mar. Biol. Biotechnol. 1992, 1, 380–389. [Google Scholar]
- Dunham, R.A.; Chitmanat, C.; Nichols, A.; Argue, B.; Powers, D.A.; Chen, T.T. Predator Avoidance of Transgenic Channel Catfish Containing Salmonid Growth Hormone Genes. Mar. Biotechnol. 1999, 1, 545–551. [Google Scholar] [CrossRef] [PubMed]
- Dunham, R.A.; Warr, G.W.; Nichols, A.; Duncan, P.L.; Argue, B.; Middleton, D.; Kucuktas, H. Enhanced Bacterial Disease Resistance of Transgenic Channel Catfish Ictalurus punctatus Possessing Cecropin Genes. Mar. Biotechnol. 2002, 4, 338–344. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, P.W. Invasion of Transgenes from Salmon or Other Genetically Modified Organisms into Natural Populations. Can. J. Fish. Aquat. Sci. 2001, 58, 2317. [Google Scholar] [CrossRef]
- Patiño, R.; Thomas, P. Effects of Gonadotropin on Ovarian Intrafollicular Processes during the Development of Oocyte Maturational Competence in a Teleost, the Atlantic Croaker: Evidence for Two Distinct Stages of Gonadotropic Control of Final Oocyte Maturation. Biol. Reprod. 1990, 43, 818–827. [Google Scholar] [CrossRef]
- Prat, F.; Sumpter, J.P.; Tyler, C.R. Validation of Radioimmunoassays for Two Salmon Gonadotropins (GTH I and GTH II) and Their Plasma Concentrations throughout the Reproductive Cycle in Male and Female Rainbow Trout (Oncorhynchus mykiss). Biol. Reprod. 1996, 54, 1375–1382. [Google Scholar] [CrossRef] [Green Version]
- Swanson, P. Salmon Gonadotropins: Reconciling Old and New Ideas. Reprod. Physiol. Fish 1991, 2–7. [Google Scholar]
- Swanson, P.; Dickey, J.T.; Campbell, B. Biochemistry and Physiology of Fish Gonadotropins. Fish Physiol. Biochem. 2003, 28, 53–59. [Google Scholar] [CrossRef]
- Kwok, H.F.; So, W.K.; Wang, Y.; Ge, W. Zebrafish Gonadotropins and Their Receptors: I. Cloning and Characterization of Zebrafish Follicle Stimulating Hormone and Luteinizing Hormone Receptors-Evidence for Their Distinct Functions in Follicle Development. Biol. Reprod. 2005, 72, 1370–1381. [Google Scholar] [CrossRef] [PubMed]
- Hsu, S.Y.; Nakabayashi, K.; Bhalla, A. Evolution of Glycoprotein Hormone Subunit Genes in Bilateral Metazoa: Identification of Two Novel Human Glycoprotein Hormone Subunit Family Genes, GPA2 and GPB5. Mol. Endocrinol. 2002, 16, 1538–1551. [Google Scholar] [CrossRef]
- Liu, Z.; Kim, S.; Karsi, A. Channel Catfish Follicle-Stimulating Hormone and Luteinizing Hormone: Complementary DNA Cloning and Expression during Ovulation. Mar. Biotechnol. 2001, 3, 590–599. [Google Scholar] [CrossRef]
- Dolomatov, S.I.; Zukow, W.; Novikov, N.Y.; Muszkieta, R.; Bulatowicz, I.; Dzierzanowski, M.; Kazmierczak, U.; Strojek, K. The Regulation of Osmotic and Ionic Balance in Fish Reproduction and in the Early Stages of Ontogeny. Russ. J. Mar. Biol. 2012, 38, 365–374. [Google Scholar] [CrossRef]
- Schulz, R.W.; Goos, H.J.T. Puberty in Male Fish: Concepts and Recent Developments with Special Reference to the African Catfish (Clarias gariepinus). Aquaculture 1999, 177, 5–12. [Google Scholar] [CrossRef]
- Ko, H.; Park, W.; Kim, D.J.; Kobayashi, M.; Sohn, Y.C. Biological Activities of Recombinant Manchurian Trout FSH and LH: Their Receptor Specificity, Steroidogenic and Vitellogenic Potencies. J. Mol. Endocrinol. 2007, 38, 99–111. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Campbell, B.; Dickey, J.; Beckman, B.; Young, G.; Pierce, A.; Fukada, H.; Swanson, P. Previtellogenic Oocyte Growth in Salmon: Relationships among Body Growth, Plasma Insulin-like Growth Factor-1, Estradiol-17beta, Follicle-Stimulating Hormone and Expression of Ovarian Genes for Insulin-like Growth Factors, Steroidogenic-Acute Regulatory Protein and Receptors for Gonadotropins, Growth Hormone, and Somatolactin. Biol. Reprod. 2006, 75, 34–44. [Google Scholar] [CrossRef]
- Qin, G.; Qin, Z.; Lu, C.; Ye, Z.; Elaswad, A.; Bangs, M.; Li, H.; Zhang, Y.; Huang, Y.; Shi, H.; et al. Gene Editing of the Catfish Gonadotropin-Releasing Hormone Gene and Hormone Therapy to Control the Reproduction in Channel Catfish, Ictalurus punctatus. Biology 2022, 11, 649. [Google Scholar] [CrossRef]
- Su, B.; Peatman, E.; Shang, M.; Thresher, R.; Grewe, P.; Patil, J.; Pinkert, C.A.; Irwin, M.H.; Li, C.; Perera, D.A.; et al. Expression and Knockdown of Primordial Germ Cell Genes, Vasa, Nanos and Dead End in Common Carp (Cyprinus carpio) Embryos for Transgenic Sterilization and Reduced Sexual Maturity. Aquaculture 2014, 420, S72–S84. [Google Scholar] [CrossRef]
- Su, B.; Shang, M.; Li, C.; Perera, D.A.; Pinkert, C.A.; Irwin, M.H.; Peatman, E.; Grewe, P.; Patil, J.G.; Dunham, R.A. Effects of Transgenic Sterilization Constructs and Their Repressor Compounds on Hatch, Developmental Rate and Early Survival of Electroporated Channel Catfish Embryos and Fry. Transgenic Res. 2015, 24, 333–352. [Google Scholar] [CrossRef]
- Su, B.; Shang, M.; Grewe, P.M.; Patil, J.G.; Peatman, E.; Perera, D.A.; Cheng, Q.; Li, C.; Weng, C.-C.; Li, P.; et al. Suppression and Restoration of Primordial Germ Cell Marker Gene Expression in Channel Catfish, Ictalurus punctatus, Using Knockdown Constructs Regulated by Copper Transport Protein Gene Promoters: Potential for Reversible Transgenic Sterilization. Theriogenology 2015, 84, 1499–1512. [Google Scholar] [CrossRef]
- Cheng, Q.; Su, B.; Qin, Z.; Weng, C.-C.; Yin, F.; Zhou, Y.; Fobes, M.; Perera, D.A.; Shang, M.; Soller, F.; et al. Interaction of Diet and the Masou Salmon Δ5-Desaturase Transgene on Δ6-Desaturase and Stearoyl-CoA Desaturase Gene Expression and N-3 Fatty Acid Level in Common Carp (Cyprinus carpio). Transgenic Res. 2014, 23, 729–742. [Google Scholar] [CrossRef]
- Qin, Z. Gene Editing of Luteinizing Hormone, Follicle-Stimulating Hormone and Gonadotropin-Releasing Hormone Genes to Sterilize Channel Catfish, Ictalurus Punctatus, Using Zinc Finger Nuclease, Transcription Activator-Like Effector Nuclease and Clustered Regularly Interspaced Short Palindromic Repeats/Cas9 Technologies. Ph.D. Thesis, Auburn University, Auburn, AL, USA, 2015. [Google Scholar]
- Qiu, P.; Shandilya, H.; D’Alessio, J.M.; O’Connor, K.; Durocher, J.; Gerard, G.F. Mutation Detection Using Surveyor Nuclease. Biotechniques 2004, 36, 702–707. [Google Scholar] [CrossRef]
- Pimkin, M.; Caretti, E.; Canutescu, A.; Yeung, J.B.; Cohn, H.; Chen, Y.; Oleykowski, C.; Bellacosa, A.; Yeung, A.T. Recombinant Nucleases CEL I from Celery and SP I from Spinach for Mutation Detection. BMC Biotechnol. 2007, 7, 29. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tsuji, T.; Niida, Y. Development of a Simple and Highly Sensitive Mutation Screening System by Enzyme Mismatch Cleavage with Optimized Conditions for Standard Laboratories. Electrophoresis 2008, 29, 1473–1483. [Google Scholar] [CrossRef] [Green Version]
- Steeby, J.; Avery, J. Channel Catfish Broodfish and Hatchery Management; Southern Regional Aquaculture Center: Stoneville, MS, USA, 2005. [Google Scholar]
- Small, B.C.; Chatakondi, N. Efficacy of Formalin as an Egg Disinfectant for Improving Hybrid Catfish (Channel Catfish × Blue Catfish) Hatching Success. N. Am. J. Aquac. 2006, 68, 9–13. [Google Scholar] [CrossRef]
- Liu, J.; Gaj, T.; Patterson, J.T.; Sirk, S.J.; Barbas, C.F., 3rd. Cell-Penetrating Peptide-Mediated Delivery of TALEN Proteins via Bioconjugation for Genome Engineering. PLoS ONE 2014, 9, e85755. [Google Scholar] [CrossRef] [Green Version]
- Mussolino, C.; Alzubi, J.; Fine, E.J.; Morbitzer, R.; Cradick, T.J.; Lahaye, T.; Bao, G.; Cathomen, T. TALENs Facilitate Targeted Genome Editing in Human Cells with High Specificity and Low Cytotoxicity. Nucleic Acids Res. 2014, 42, 6762–6773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreyer, A.K.; Hoffmann, D.; Lachmann, N.; Ackermann, M.; Steinemann, D.; Timm, B.; Siler, U.; Reichenbach, J.; Grez, M.; Moritz, T.; et al. TALEN-Mediated Functional Correction of X-Linked Chronic Granulomatous Disease in Patient-Derived Induced Pluripotent Stem Cells. Biomaterials 2015, 69, 191–200. [Google Scholar] [CrossRef]
- Veres, A.; Gosis, B.S.; Ding, Q.; Collins, R.; Ragavendran, A.; Brand, H.; Erdin, S.; Cowan, C.A.; Talkowski, M.E.; Musunuru, K. Low Incidence of Off-Target Mutations in Individual CRISPR-Cas9 and TALEN Targeted Human Stem Cell Clones Detected by Whole-Genome Sequencing. Cell Stem Cell 2014, 15, 254. [Google Scholar] [CrossRef] [Green Version]
- Saville, K.J.; Belote, J.M. Identification of an Essential Gene, l(3)73Ai, with a Dominant Temperature-Sensitive Lethal Allele, Encoding a Drosophila Proteasome Subunit. Proc. Natl. Acad. Sci. USA 1993, 90, 8842–8846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweisguth, F. Dominant-Negative Mutation in the β2 and β6 Proteasome Subunit Genes Affect Alternative Cell Fate Decisions in the Drosophila Sense Organ Lineage. Proc. Natl. Acad. Sci. USA 1999, 96, 11382–11386. [Google Scholar] [CrossRef] [Green Version]
- Kanamori, A.; Kagawa, H.; Nagahama, Y. Gonadotropin Receptors in the Postovulatory Ovary of Amago Salmon (Oncorhynchus rhodurus). Gen. Comp. Endocrinol. 1987, 66, 210–217. [Google Scholar] [CrossRef] [PubMed]
- Jamal Uddin, M.; Bhattacharya, S. In-Vitro Binding of Gonadotrophin to Fish Ovary. J. Endocrinol. 1986, 111, 407–413. [Google Scholar] [CrossRef] [PubMed]
- Breton, B.; Govoroun, M.; Mikolajczyk, T. GTH I and GTH II Secretion Profiles during the Reproductive Cycle in Female Rainbow Trout: Relationship with Pituitary Responsiveness to GnRH-A Stimulation. Gen. Comp. Endocrinol. 1998, 111, 38–50. [Google Scholar] [CrossRef] [PubMed]
- Weil, C.; Bougoussa-Houadec, M.; Gallais, C.; Itoh, S.; Sekine, S.; Valotaire, Y. Preliminary Evidence Suggesting Variations of GtH1 and GtH2 MRNA Levels at Different Stages of Gonadal Development in Rainbow Trout, Oncorhynchus mykiss. Gen. Comp. Endocrinol. 1995, 100, 327–333. [Google Scholar] [CrossRef] [PubMed]
- Senthilkumaran, B.; Kar, S. Advances in Reproductive Endocrinology and Neuroendocrine Research Using Catfish Models. Cells 2021, 10, 2807. [Google Scholar] [CrossRef]
- Fontaine, R.; Royan, M.R.; von Krogh, K.; Weltzien, F.-A.; Baker, D.M. Direct and Indirect Effects of Sex Steroids on Gonadotrope Cell Plasticity in the Teleost Fish Pituitary. Front. Endocrinol. 2020, 11, 605068. [Google Scholar] [CrossRef]
- Ulloa-Aguirre, A.; Timossi, C. Structure-Function Relationship of Follicle-Stimulating Hormone and Its Receptor. Hum. Reprod. Update 1998, 4, 260–283. [Google Scholar] [CrossRef] [Green Version]
- Zhang, Z.; Zhu, B.; Ge, W. Genetic Analysis of Zebrafish Gonadotropin (FSH and LH) Functions by TALEN-Mediated Gene Disruption. Mol. Endocrinol. 2015, 29, 76–98. [Google Scholar] [CrossRef] [Green Version]
- Dunham, R.A. Aquaculture and Fisheries Biotechnology: Genetic Approaches; CABI Publishing: Wallingford, UK, 2023. [Google Scholar]
- Yan, L.; Swanson, P.; Dickhoff, W.W. A Two-Receptor Model for Salmon Gonadotropins (GTH I and GTH II). Biol. Reprod. 1992, 47, 418–427. [Google Scholar] [CrossRef]
- Vischer, H.F.; Granneman, J.C.M.; Linskens, M.H.K.; Schulz, R.W.; Bogerd, J. Both Recombinant African Catfish LH and FSH Are Able to Activate the African Catfish FSH Receptor. J. Mol. Endocrinol. 2003, 31, 133–140. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Basu, D.; Bhattacharya, S. Purification of Two Types of Gonadotropin Receptors from Carp Ovarian Follicles: Over-Lapping Recognition by Two Different Ligands. Gen. Comp. Endocrinol. 2002, 129, 152–162. [Google Scholar] [CrossRef] [PubMed]
- Ascoli, M.; Fanelli, F.; Segaloff, D.L. The Lutropin/Choriogonadotropin Receptor, a 2002 Perspective. Endocr. Rev. 2002, 23, 141–174. [Google Scholar] [CrossRef]
- Troppmann, B.; Kleinau, G.; Krause, G.; Gromoll, J. Structural and Functional Plasticity of the Luteinizing Hormone/Choriogonadotrophin Receptor. Hum. Reprod. Update 2013, 19, 583–602. [Google Scholar] [CrossRef] [Green Version]
- De Rensis, F.; López-Gatius, F. Use of Equine Chorionic Gonadotropin to Control Reproduction of the Dairy Cow: A Review. Reprod. Domest. Anim. 2014, 49, 177–182. [Google Scholar] [CrossRef]
- Min, K.-S.; Park, J.-J.; Byambaragchaa, M.; Kang, M.-H. Characterization of Tethered Equine Chorionic Gonadotropin and Its Deglycosylated Mutants by Ovulation Stimulation in Mice. BMC Biotechnol. 2019, 19, 60. [Google Scholar] [CrossRef] [Green Version]
- Zohar, Y.; Mylonas, C.C. Endocrine Manipulations of Spawning in Cultured Fish: From Hormones to Genes. Aquaculture 2001, 197, 99–136. [Google Scholar] [CrossRef]
- Prentice, J.A.; Thomas, P. Successful Spawning of Orangemouth Corvina Following Injection with Des-Gly10,[D-Ala6]-Luteinizing Hormone-Releasing Hormone (1-9) Ethylamide and Pimozide. Prog. Fish-Cult. 1987, 49, 66–69. [Google Scholar] [CrossRef]
- Rottmann, R.W.; Shireman, J.V.; Chapman, F.A. Hormonal Control of Reproduction in Fish for Induced Spawning; Southern Regional Aquaculture Center: Stoneville, MS, USA, 1991. [Google Scholar]
- Goudie, C.A.; Simco, B.A.; Davis, K.B.; Parker, N.C. Reproductive Performance of Pigmented and Albino Female Channel Catfish Induced to Spawn with HCG or Ovaprim. J. World Aquac. Soc. 1992, 23, 138–145. [Google Scholar] [CrossRef]
- Shehadeh, Z.H.; Madden, W.D.; Dohl, T.P. The Effect of Exogenous Hormone Treatment on Spermiation and Vitellogenesis in the Grey Mullet, Mugil cephalus L. J. Fish Biol. 1973, 5, 479–487. [Google Scholar] [CrossRef]
- Kucharczyk, D.; Kujawa, R.; Murmurz, A.; Skrzypczak, A.; Wyszomirska, E. Induced Spawning in Perch, Perca fluviatilis L. Using Carp Pituitary Extract and HCG. Aquac. Res. 1996, 27, 847–852. [Google Scholar] [CrossRef]
- Kido, A.; Mori, M.; Adachi, Y.; Yukaya, H.; Ishida, T.; Sugimachi, K. Immunohistochemical Ex-Pression of β-Human Chorionic Gonadotropin in Colorectal Carcinoma. Surg. Today 1996, 26, 966–970. [Google Scholar] [CrossRef] [PubMed]
- Filicori, M.; Fazleabas, A.T.; Huhtaniemi, I.; Licht, P.; Rao, C.V.; Tesarik, J.; Zygmunt, M. Novel Concepts of Human Chorionic Gonadotropin: Reproductive System Interactions and Potential in the Management of Infertility. Fertil. Steril. 2005, 84, 275–284. [Google Scholar] [CrossRef] [PubMed]
- Shupnik, M.A.; Gharib, S.D.; Chin, W.W. Divergent Effects of Estradiol on Gonadotropin Gene Transcription in Pituitary Fragments. Mol. Endocrinol. 1989, 3, 474–480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mercer, J.E.; Clements, J.A.; Funder, J.W.; Clarke, I.J. Regulation of Follicle-Stimulating Hormone β and Common α-Subunit Messenger Ribonucleic Acid by Gonadotropin-Releasing Hormone and Estrogen in the Sheep Pituitary. Neuroendocrinology 1989, 50, 321–326. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Spawning in 2016 | Spawning in 2017 | ||||||||
|---|---|---|---|---|---|---|---|---|---|
| Fish N | Spawned Fish N | Spawning Rate (%) a | Mean Hatch Rate (%) | Fish N | Spawned Fish N | Spawning Rate b (%) | Mean Hatch Rate (%) c | ||
| fsh | F | 3 | 1 | 33.3 | 1.0 * | 10 | 8 | 80.0 | 35.0 ± 0.14 |
| M | 5 | 2 | 40.0 | 0.5 ± 0.05 * | 9 | 8 | 88.9 | 35.0 ± 0.14 | |
| iCTRL | M and F | / | / | / | / | 3 | 2 | 66.7 | 56.5 ± 0.34 |
| nCTRL | M and F | 5 | 3 | 60.0 | 74.3 ± 0.02 * | 7 | 5 | 71.4 | 32.3 ± 0.20 |
| Family Names of F1 Offspring | Survival Evaluation | Mutation Evaluation | Mean Body Weight (g) Mutant Fish a | Body Weight (g) of Non-Mutant Fish a | ||||
|---|---|---|---|---|---|---|---|---|
| N Fish | N Fish Survived | Survival Rate (%) | N Fish Sampled | N Mutant Fish | Mutation Rate (%) | |||
| 2016 FSH | 250 | 222 | 88.9 | 30 | 19 | 63.3 | 36.4 ± 2.13 | 35.6 ± 2.21 |
| 2017 FSH-1 | 22 | 20 | 91.0 | 20 | 11 | 55.0 | 30.2 ± 4.76 | 28.6 ± 5.26 |
| 2017 FSH-2 | 300 | 279 | 93.0 | 30 | 15 | 50.0 | 9.5 ± 0.55 | 10.9 ± 0.81 |
| 2017 FSH-3 | 24 | 20 | 83.3 | 25 | 16 | 64.0 | 15.2 ± 1.09 * | 22.8 ± 2.19 * |
| 2017 FSH-4 | 300 | 266 | 88.7 | 21 | 10 | 47.6 | 24.6 ± 3.22 | 26.6 ± 1.89 |
| 2017 FSH-5 | 300 | 168 | 56.0 | 30 | 18 | 60.0 b | 12.8 ± 0.82 | 11.9 ± 0.62 |
| 2017 FSH-5 dead fish | / | / | / | 30 | 18 | 60.0 b | 16.67 ± 0.811 | 17.67 ± 1.509 |
| 2017 FSH-6 | 300 | 259 | 86.3 | 28 | 13 | 46.4 | 18.3 ± 1.48 | 16.2 ± 0.78 |
| 2017 FSH-7 | 300 | 269 | 90.0 | 22 | 10 | 45.5 | 8.4 ± 0.70 | 9.3 ± 1.41 |
| Treatment | P1 Embryos and 6-Month-Old Fingerlings | Genotype | 4-Year-Old P1 Fish in 2017 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Recirculating System | Pond | ||||||||
| N Eggs | N Hatched | Hatch Rate (%) a | Survival Rate (%) b | N Fish | Survival Rate (%) c | N Fish | Survival Rate (%) * | ||
| Electroporated with plasmids | 200 | 80 | 44.0 | 71.3 | fsh Mutants | 42 | 54.8 | 37 | 54.1 |
| Electroporated without plasmids | 200 | 84 | 44.0 | 75.0 | fsh Non-mutants | 2 | 50.0 | 6 | 0 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, G.; Qin, Z.; Lu, C.; Ye, Z.; Elaswad, A.; Jin, Y.; Khan, M.G.Q.; Su, B.; Dunham, R.A. Gene Editing of the Follicle-Stimulating Hormone Gene to Sterilize Channel Catfish, Ictalurus punctatus, Using a Modified Transcription Activator-like Effector Nuclease Technology with Electroporation. Biology 2023, 12, 392. https://doi.org/10.3390/biology12030392
Qin G, Qin Z, Lu C, Ye Z, Elaswad A, Jin Y, Khan MGQ, Su B, Dunham RA. Gene Editing of the Follicle-Stimulating Hormone Gene to Sterilize Channel Catfish, Ictalurus punctatus, Using a Modified Transcription Activator-like Effector Nuclease Technology with Electroporation. Biology. 2023; 12(3):392. https://doi.org/10.3390/biology12030392
Chicago/Turabian StyleQin, Guyu, Zhenkui Qin, Cuiyu Lu, Zhi Ye, Ahmed Elaswad, Yulin Jin, Mohd Golam Quader Khan, Baofeng Su, and Rex A. Dunham. 2023. "Gene Editing of the Follicle-Stimulating Hormone Gene to Sterilize Channel Catfish, Ictalurus punctatus, Using a Modified Transcription Activator-like Effector Nuclease Technology with Electroporation" Biology 12, no. 3: 392. https://doi.org/10.3390/biology12030392