
Soil Geochemical Properties Influencing the Diversity of Bacteria and Archaea in Soils of the Kitezh Lake Area, Antarctica

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Study Sites and Sample Collection
2.2. Geochemical Properties of Soils
2.3. DNA Extraction, PCR Amplification, and Products Processing
2.4. Sequence Analysis
2.5. Statistical Analysis
3. Results
3.1. Geochemical Properties of Soil Samples
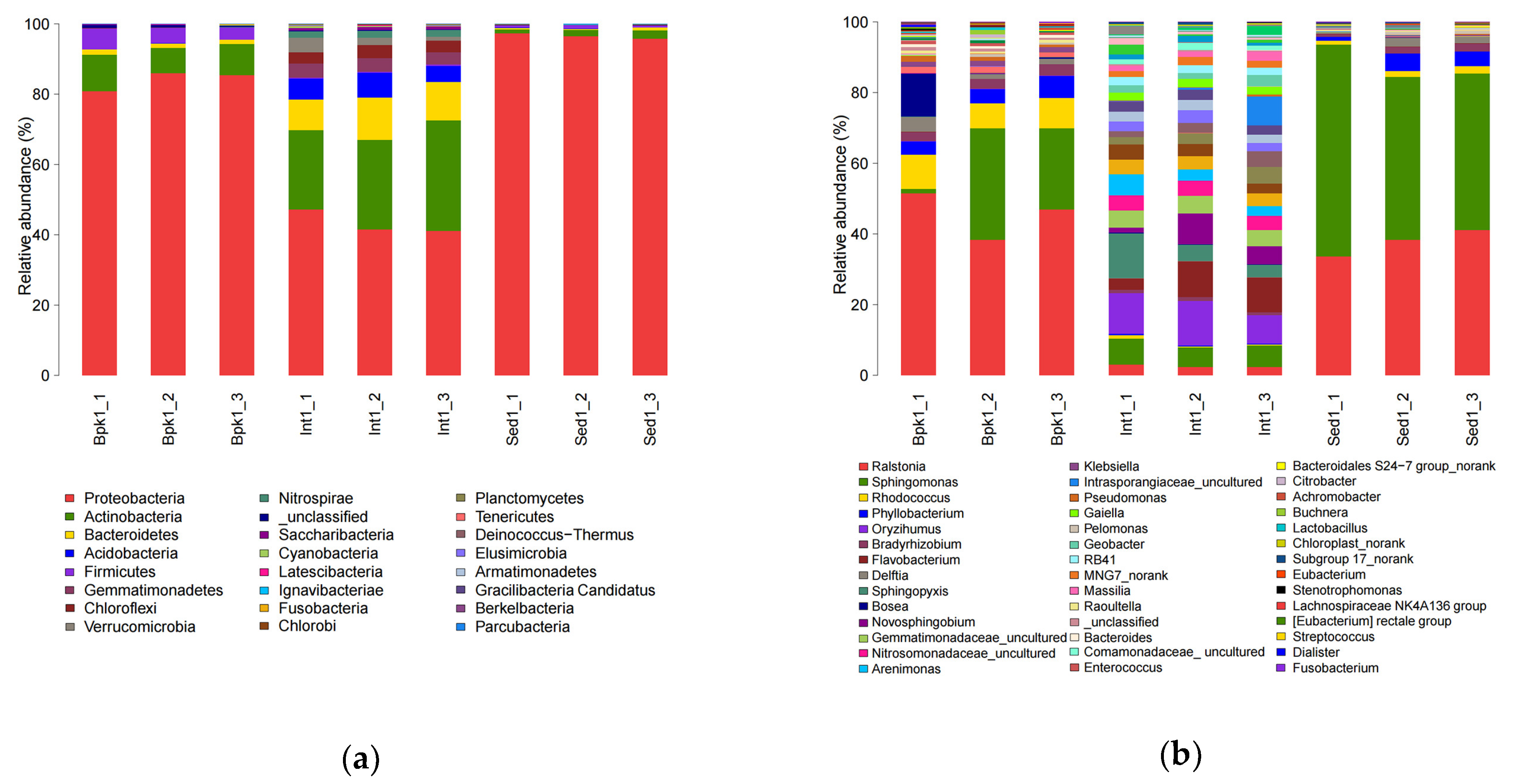
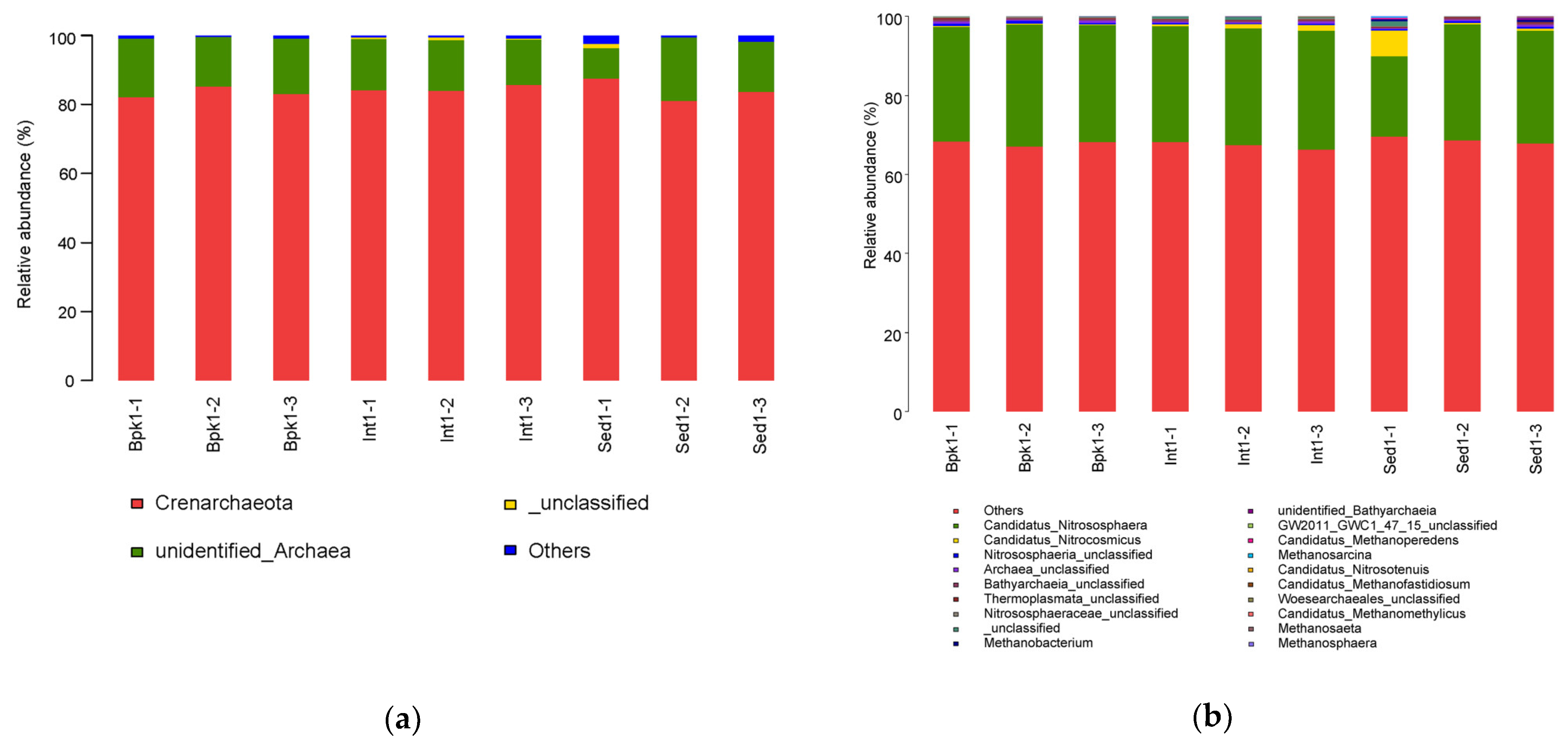
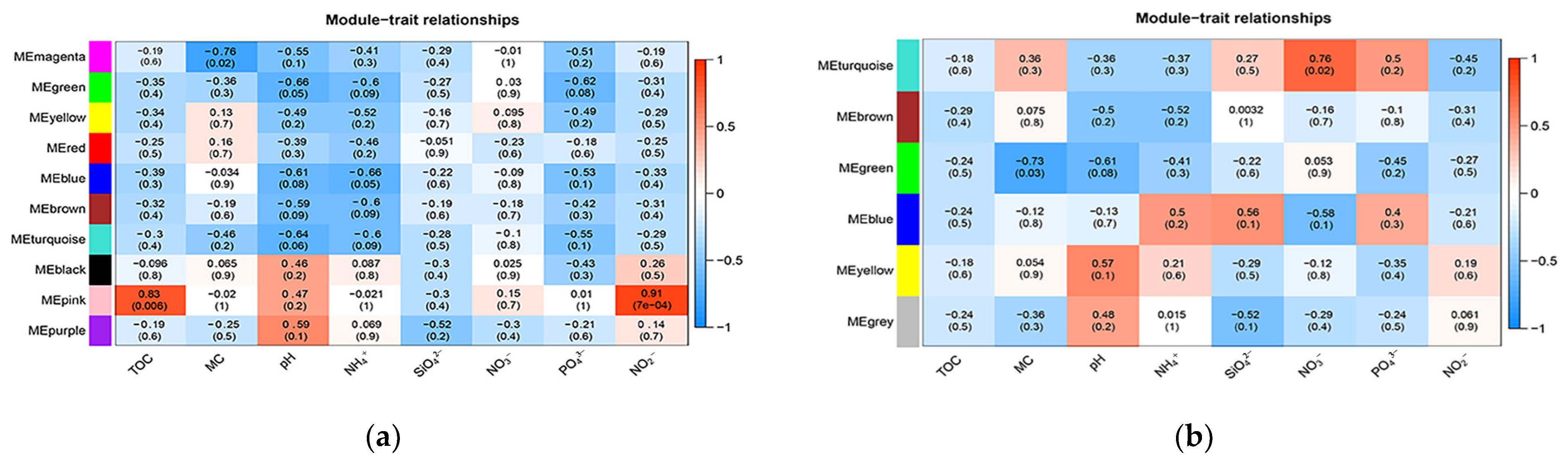
3.2. Diversity and Structure Analysis of Bacterial and Archaeal Communities
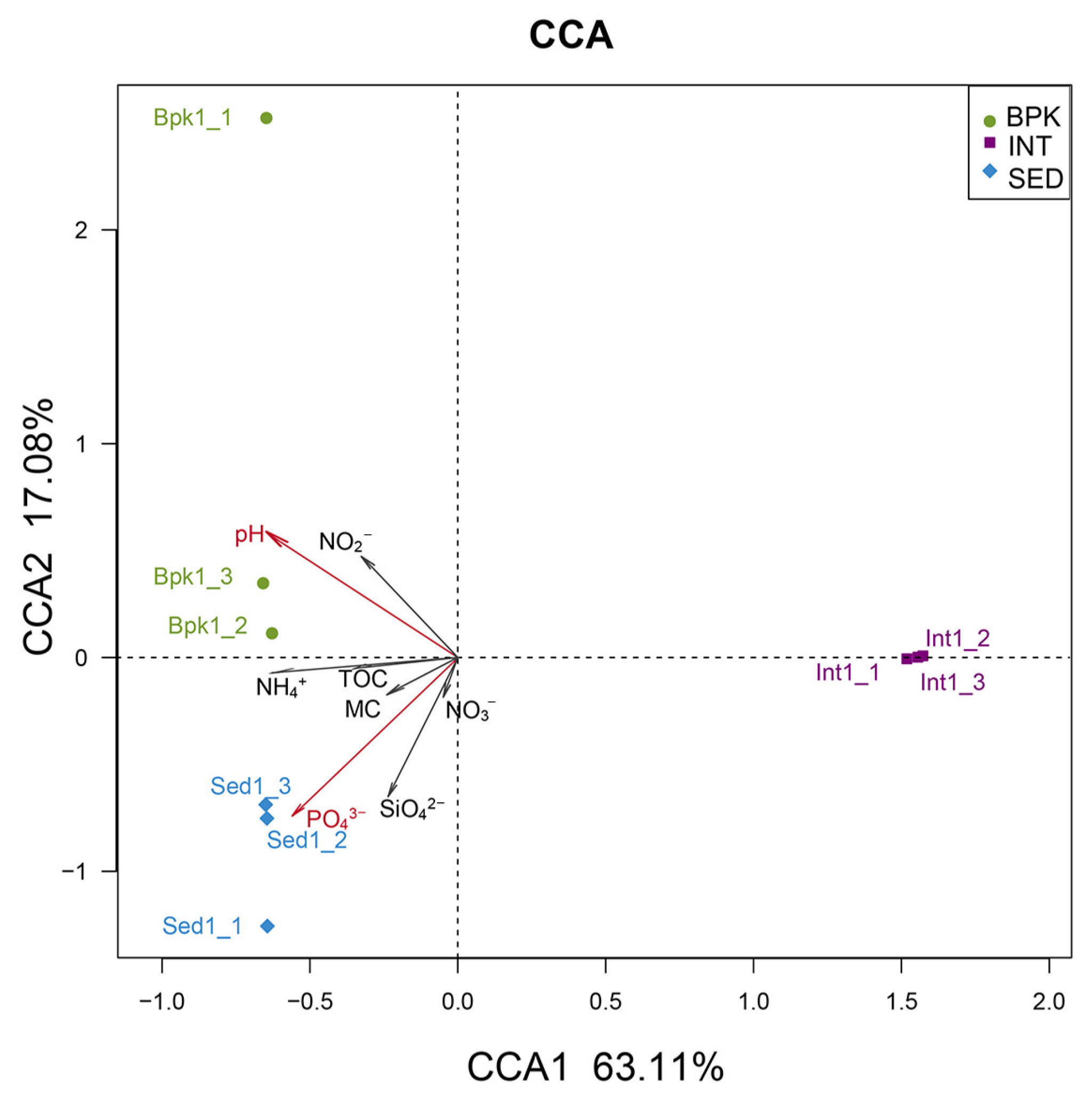
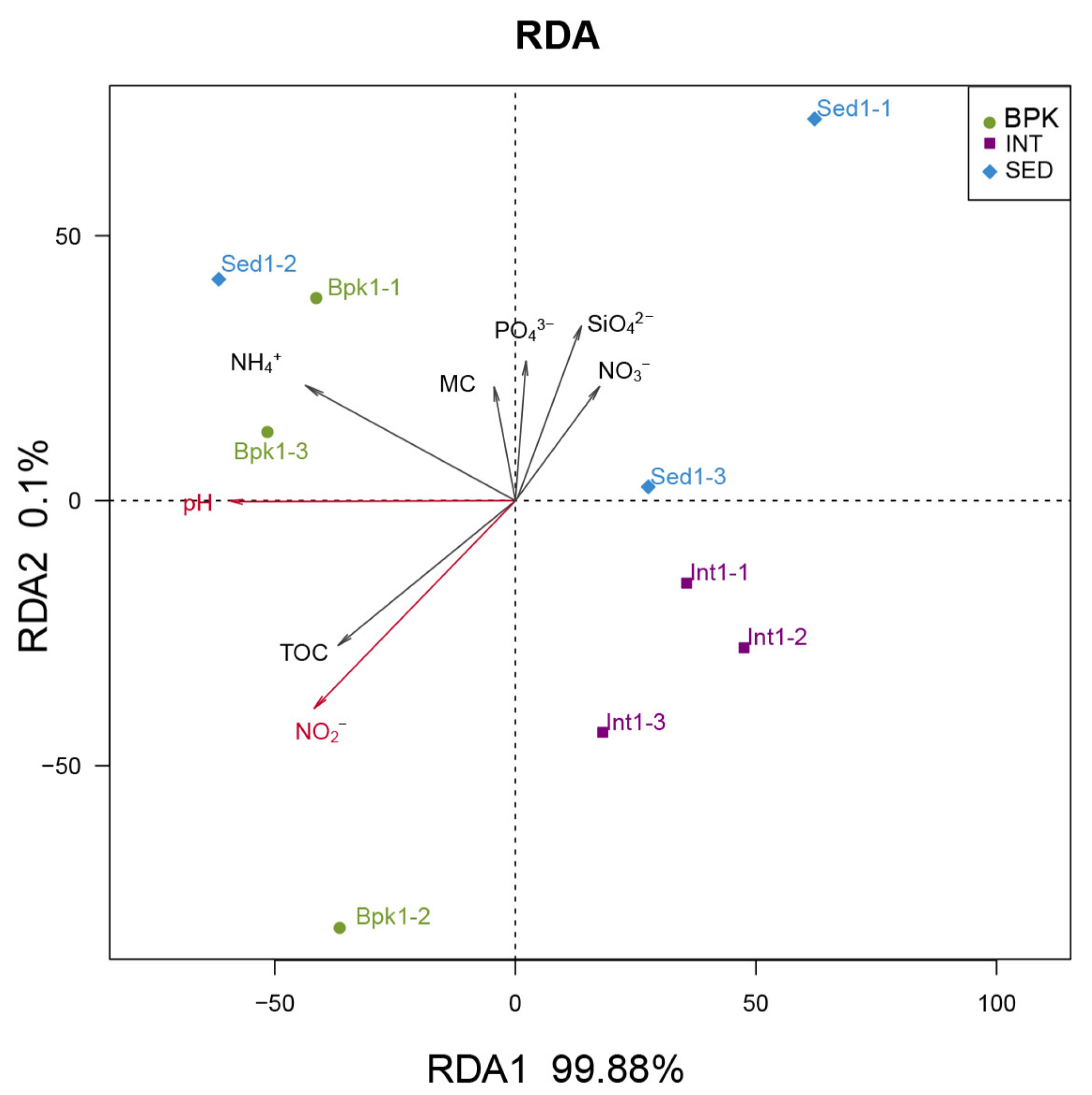
3.3. Correlation between Geochemical Properties and Bacterial and Archaeal Community
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Post, E.; Alley, R.B.; Christensen, T.R.; Macias-Fauria, M.; Forbes, B.C.; Gooseff, M.N.; Iler, A.; Kerby, J.T.; Laidre, K.L.; Mann, M.E.; et al. The polar regions in a 2°C warmer world. Sci. Adv. 2019, 5, eaaw9883. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jenkins, A.; Shoosmith, D.; Dutrieux, P.; Jacobs, S.; Kim, T.W.; Lee, S.H.; Ha, H.K.; Stammerjohn, S. West Antarctic Ice Sheet retreat in the Amundsen Sea driven by decadal oceanic variability. Nat. Geosci. 2018, 11, 733–738. [Google Scholar] [CrossRef]
- Kingslake, J.; Scherer, R.P.; Albrecht, T.; Coenen, J.; Powell, R.D.; Reese, R.; Stansell, N.D.; Tulaczyk, S.; Wearing, M.G.; Whitehouse, P.L. Extensive retreat and re-advance of the West Antarctic Ice Sheet during the Holocene. Nature 2018, 558, 430–434. [Google Scholar] [CrossRef] [Green Version]
- Edwards, T.L.; Brandon, M.A.; Durand, G.; Edwards, N.R.; Golledge, N.R.; Holden, P.B.; Nias, I.J.; Payne, A.J.; Ritz, C.; Wernecke, A. Revisiting Antarctic ice loss due to marine ice-cliff instability. Nature 2019, 566, 58–64. [Google Scholar] [CrossRef] [Green Version]
- Rignot, E.; Mouginot, J.; Scheuchl, B.; van den Broeke, M.; van Wessem, M.J.; Morlighem, M. Four decades of Antarctic Ice Sheet mass balance from 1979–2017. Proc. Natl. Acad. Sci. USA 2019, 116, 1095–1103. [Google Scholar] [CrossRef] [Green Version]
- Bronselaer, B.; Winton, M.; Griffies, S.M.; Hurlin, W.J.; Rodgers, K.B.; Sergienko, O.V.; Stouffer, R.J.; Russell, J.L. Change in future climate due to Antarctic meltwater. Nature 2018, 564, 53–58. [Google Scholar] [CrossRef] [PubMed]
- Bell, R.E.; Banwell, A.F.; Trusel, L.D.; Kingslake, J. Antarctic surface hydrology and impacts on ice-sheet mass balance. Nat. Clim. Chang. 2018, 8, 1044–1052. [Google Scholar] [CrossRef]
- Quayle, W.C.; Peck, L.S.; Peat, H.; Ellis-Evans, J.C.; Harrigan, P.R. Extreme responses to climate change in Antarctic lakes. Science 2002, 295, 645. [Google Scholar] [CrossRef]
- Vincent, W.F. Microbial ecosystem responses to rapid climate change in the Arctic. ISME J. 2010, 4, 1087–1090. [Google Scholar] [CrossRef] [Green Version]
- Rinnan, R.; Rousk, J.; Yergeau, E.; Kowalchuk, G.A.; Bååth, E. Temperature adaptation of soil bacterial communities along an Antarctic climate gradient: Predicting responses to climate warming. Glob. Chang. Biol. 2009, 15, 2615–2625. [Google Scholar] [CrossRef]
- Wang, N.; Guo, Y.; Li, G.; Xia, Y.; Ma, M.; Zang, J.; Ma, Y.; Yin, X.; Han, W.; Lv, J.; et al. Geochemical-Compositional-Functional Changes in Arctic Soil Microbiomes Post Land Submergence Revealed by Metagenomics. Microbes Environ. 2019, 34, 180–190. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, T.; Wang, N.F.; Liu, H.Y.; Zhang, Y.Q.; Yu, L.Y. Soil pH is a Key Determinant of Soil Fungal Community Composition in the Ny-Alesund Region, Svalbard (High Arctic). Front. Microbiol. 2016, 7, 227. [Google Scholar] [CrossRef] [PubMed]
- Han, W.; Wang, N.; Ma, Y.; Lv, J.; Wang, S.; Zhang, B.; Jiang, Z.; Cao, H. The Effect of Organic Carbon on Soil Bacterial Diversity in an Antarctic Lake Region. J. Ocean. Univ. China 2019, 18, 1402–1410. [Google Scholar] [CrossRef]
- Kanasugi, M.; Sarkodee-Addo, E.; Ansong Omari, R.; Mohammad Golam Dastogeer, K.; Fujii, Y.; Oppong Abebrese, S.; Bam, R.; Asuming-Brempong, S.; Okazaki, S. Exploring Rice Root Microbiome; The Variation, Specialization and Interaction of Bacteria and Fungi in Six Tropic Savanna Regions in Ghana. Sustainability 2020, 12, 5835. [Google Scholar] [CrossRef]
- Wang, H.; Tian, T.; Gong, Y.; Ma, S.; Altaf, M.M.; Wu, H.; Diao, X. Both environmental and spatial variables affect bacterial functional diversity in mangrove sediments at an island scale. Sci. Total Environ. 2021, 753, 142054. [Google Scholar] [CrossRef]
- Guo, Y.; Wang, N.; Li, G.; Rosas, G.; Zang, J.; Ma, Y.; Liu, J.; Han, W.; Cao, H. Direct and Indirect Effects of Penguin Feces on Microbiomes in Antarctic Ornithogenic Soils. Front. Microbiol. 2018, 9, 552. [Google Scholar] [CrossRef]
- Zhang, T.; Wang, N.F.; Zhang, Y.Q.; Liu, H.Y.; Yu, L.Y. Diversity and distribution of fungal communities in the marine sediments of Kongsfjorden, Svalbard (High Arctic). Sci. Rep. 2015, 5, 14524. [Google Scholar] [CrossRef] [Green Version]
- Chong, C.W.; Pearce, D.A.; Convey, P.; Yew, W.C.; Tan, I.K.P. Patterns in the distribution of soil bacterial 16S rRNA gene sequences from different regions of Antarctica. Geoderma 2012, 181–182, 45–55. [Google Scholar] [CrossRef]
- Tang, C.; Madigan, M.T.; Lanoil, B. Bacterial and archaeal diversity in sediments of west Lake Bonney, McMurdo Dry Valleys, Antarctica. Appl. Environ. Microbiol. 2013, 79, 1034–1038. [Google Scholar] [CrossRef] [Green Version]
- Cameron, K.A.; Hodson, A.J.; Osborn, A.M. Structure and diversity of bacterial, eukaryotic and archaeal communities in glacial cryoconite holes from the Arctic and the Antarctic. FEMS Microbiol. Ecol. 2012, 82, 254–267. [Google Scholar] [CrossRef]
- Roesch, L.F.W.; Fulthorpe, R.R.; Pereira, A.B.; Pereira, C.K.; Lemos, L.N.; Barbosa, A.D.; Suleiman, A.K.A.; Gerber, A.L.; Pereira, M.G.; Loss, A.; et al. Soil bacterial community abundance and diversity in ice-free areas of Keller Peninsula, Antarctica. Appl. Soil Ecol. 2012, 61, 7–15. [Google Scholar] [CrossRef]
- Zhang, F.; Zhang, J.-T.; Zhang, F. Pattern of forest vegetation and its environmental interpretation in Zhuweigou, Lishan Mountain Nature Reserve. Acta Ecol. Sin. 2003, 23, 421–427. [Google Scholar]
- Zhao, X. Revision Method of the Arch Effect of Quantification II and Its Application. Stat. Res. 2020, 37, 106–118. [Google Scholar]
- Miller, J.A.; Oldham, M.C.; Geschwind, D.H. A systems level analysis of transcriptional changes in Alzheimer’s disease and normal aging. J. Neurosci. 2008, 28, 1410–1420. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fuller, T.F.; Ghazalpour, A.; Aten, J.E.; Drake, T.A.; Lusis, A.J.; Horvath, S. Weighted gene coexpression network analysis strategies applied to mouse weight. Mamm. Genome 2007, 18, 463–472. [Google Scholar] [CrossRef] [Green Version]
- Li, H.; Su, Q.; Li, B.; Lan, L.; Wang, C.; Li, W.; Wang, G.; Chen, W.; He, Y.; Zhang, C. High expression of WTAP leads to poor prognosis of gastric cancer by influencing tumour-associated T lymphocyte infiltration. J. Cell Mol. Med. 2020, 24, 4452–4465. [Google Scholar] [CrossRef]
- Zuo, J.; Zu, M.; Liu, L.; Song, X.; Yuan, Y. Composition and diversity of bacterial communities in the rhizosphere of the Chinese medicinal herb Dendrobium. BMC Plant Biol. 2021, 21, 127. [Google Scholar] [CrossRef]
- Castillo, J.D.; Vivanco, J.M.; Manter, D.K. Bacterial Microbiome and Nematode Occurrence in Different Potato Agricultural Soils. Microb. Ecol. 2017, 74, 888–900. [Google Scholar] [CrossRef]
- Deng, Q.; Yu, T.; Zeng, Z.; Ashraf, U.; Shi, Q.; Huang, S.; Lian, T.; Chen, J.; Muzaffar, W.; Shen, W. Silicon Application Modulates the Growth, Rhizosphere Soil Characteristics, and Bacterial Community Structure in Sugarcane. Front. Plant Sci. 2021, 12, 710139. [Google Scholar] [CrossRef]
- Li, Y.; Kromer, B.; Schukraft, G.; Bubenzer, O.; Huang, M.R.; Wang, Z.M.; Bian, L.G.; Li, C.S. Growth rate of Usnea aurantiacoatra (Jacq.) Bory on Fildes Peninsula, Antarctica and its climatic background. PLoS ONE 2014, 9, e100735. [Google Scholar] [CrossRef]
- Niem, J.M.; Billones-Baaijens, R.; Stodart, B.; Savocchia, S. Diversity Profiling of Grapevine Microbial Endosphere and Antagonistic Potential of Endophytic Pseudomonas Against Grapevine Trunk Diseases. Front. Microbiol. 2020, 11, 477. [Google Scholar] [CrossRef] [PubMed]
- Wei, S.; Cui, H.; Zhang, Y.; Su, X.; Dong, H.; Chen, F.; Zhu, Y. Comparative evaluation of three archaeal primer pairs for exploring archaeal communities in deep-sea sediments and permafrost soils. Extremophiles 2019, 23, 747–757. [Google Scholar] [CrossRef]
- Ravi, R.K.; Walton, K.; Khosroheidari, M. MiSeq: A Next Generation Sequencing Platform for Genomic Analysis. Methods Mol. Biol. 2018, 1706, 223–232. [Google Scholar] [CrossRef] [PubMed]
- Prodan, A.; Tremaroli, V.; Brolin, H.; Zwinderman, A.H.; Nieuwdorp, M.; Levin, E. Comparing bioinformatic pipelines for microbial 16S rRNA amplicon sequencing. PLoS ONE 2020, 15, e0227434. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Quast, C.; Pruesse, E.; Yilmaz, P.; Gerken, J.; Schweer, T.; Yarza, P.; Peplies, J.; Glockner, F.O. The SILVA ribosomal RNA gene database project: Improved data processing and web-based tools. Nucleic Acids Res. 2013, 41, D590–D596. [Google Scholar] [CrossRef]
- Alekseev, I.; Zverev, A.; Abakumov, E. Microbial Communities in Permafrost Soils of Larsemann Hills, Eastern Antarctica: Environmental Controls and Effect of Human Impact. Microorganisms 2020, 8, 1202. [Google Scholar] [CrossRef] [PubMed]
- Langfelder, P.; Horvath, S. WGCNA: An R package for weighted correlation network analysis. BMC Bioinform. 2008, 9, 559. [Google Scholar] [CrossRef] [Green Version]
- Rochera, C.; Quesada, A.; Toro, M.; Rico, E.; Camacho, A. Plankton assembly in an ultra-oligotrophic Antarctic lake over the summer transition from the ice-cover to ice-free period: A size spectra approach. Polar Sci. 2017, 11, 72–82. [Google Scholar] [CrossRef]
- Yergeau, E.; Kowalchuk, G.A. Responses of Antarctic soil microbial communities and associated functions to temperature and freeze-thaw cycle frequency. Environ. Microbiol. 2008, 10, 2223–2235. [Google Scholar] [CrossRef]
- Li, W.; Morgan-Kiss, R.M. Influence of Environmental Drivers and Potential Interactions on the Distribution of Microbial Communities from Three Permanently Stratified Antarctic Lakes. Front. Microbiol. 2019, 10, 1067. [Google Scholar] [CrossRef] [Green Version]
- Hernández, E.A.; Lopez, J.L.; Piquet, A.M.T.; Mac Cormack, W.P.; Buma, A.G.J. Changes in salinity and temperature drive marine bacterial communities’ structure at Potter Cove, Antarctica. Polar Biol. 2019, 42, 2177–2191. [Google Scholar] [CrossRef]
- Papale, M.; Rizzo, C.; Villescusa, J.A.; Rochera, C.; Camacho, A.; Michaud, L.; Lo Giudice, A. Prokaryotic assemblages in the maritime Antarctic Lake Limnopolar (Byers Peninsula, South Shetland Islands). Extremophiles 2017, 21, 947–961. [Google Scholar] [CrossRef] [PubMed]
- Pershina, E.V.; Ivanova, E.A.; Abakumov, E.V.; Andronov, E.E. The impacts of deglaciation and human activity on the taxonomic structure of prokaryotic communities in Antarctic soils on King George Island. Antarct. Sci. 2018, 30, 278–288. [Google Scholar] [CrossRef]
- Aislabie, J.M.; Jordan, S.; Barker, G.M. Relation between soil classification and bacterial diversity in soils of the Ross Sea region, Antarctica. Geoderma 2008, 144, 9–20. [Google Scholar] [CrossRef]
- Aislabie, J.M.; Chhour, K.-L.; Saul, D.J.; Miyauchi, S.; Ayton, J.; Paetzold, R.F.; Balks, M.R. Dominant bacteria in soils of Marble Point and Wright Valley, Victoria Land, Antarctica. Soil Biol. Biochem. 2006, 38, 3041–3056. [Google Scholar] [CrossRef]
- Yang, C.; Tang, W.; Sun, J.; Guo, H.; Sun, S.; Miao, F.; Yang, G.; Zhao, Y.; Wang, Z.; Sun, J. Weeds in the Alfalfa Field Decrease Rhizosphere Microbial Diversity and Association Networks in the North China Plain. Front. Microbiol. 2022, 13, 840774. [Google Scholar] [CrossRef]
- Li, X.; Liu, L.; Zhu, Y.; Zhu, T.; Wu, X.; Yang, D. Microbial Community Structure and Its Driving Environmental Factors in Black Carp (Mylopharyngodon piceus) Aquaculture Pond. Water 2021, 13, 3089. [Google Scholar] [CrossRef]
- Zheng, Y.; Su, Z.; Dai, T.; Li, F.; Huang, B.; Mu, Q.; Feng, C.; Wen, D. Identifying human-induced influence on microbial community: A comparative study in the effluent-receiving areas in Hangzhou Bay. Front. Environ. Sci. Eng. 2019, 13, 90. [Google Scholar] [CrossRef]
- Kim, Y.-J.; Park, J.Y.; Balusamy, S.R.; Huo, Y.; Nong, L.K.; Thi Le, H.; Yang, D.C.; Kim, D. Comprehensive Genome Analysis on the Novel Species Sphingomonas panacis DCY99T Reveals Insights into Iron Tolerance of Ginseng. Int. J. Mol. Sci. 2020, 21, 2019. [Google Scholar] [CrossRef] [Green Version]
- Asaf, S.; Khan, A.L.; Khan, M.A.; Al-Harrasi, A.; Lee, I.-J. Complete genome sequencing and analysis of endophytic Sphingomonas sp. LK11 and its potential in plant growth. 3 Biotech 2018, 8, 389. [Google Scholar] [CrossRef] [Green Version]
- Ayton, J.; Aislabie, J.; Barker, G.M.; Saul, D.; Turner, S. Crenarchaeota affiliated with group 1.1b are prevalent in coastal mineral soils of the Ross Sea region of Antarctica. Environ. Microbiol. 2010, 12, 689–703. [Google Scholar] [CrossRef] [PubMed]
- Coleine, C.; Biagioli, F.; de Vera, J.P.; Onofri, S.; Selbmann, L. Endolithic microbial composition in Helliwell Hills, a newly investigated Mars-like area in Antarctica. Environ. Microbiol. 2021, 23, 4002–4016. [Google Scholar] [CrossRef] [PubMed]
- Magalhaes, C.M.; Machado, A.; Frank-Fahle, B.; Lee, C.K.; Cary, S.C. The ecological dichotomy of ammonia-oxidizing archaea and bacteria in the hyper-arid soils of the Antarctic Dry Valleys. Front. Microbiol. 2014, 5, 515. [Google Scholar] [CrossRef] [PubMed]
- Santos, A.; Bruna, P.; Martinez-Urtaza, J.; Solis, F.; Valenzuela, B.; Zamorano, P.; Barrientos, L. Two Archaeal Metagenome-Assembled Genomes from El Tatio Provide New Insights into the Crenarchaeota Phylum. Genes 2021, 12, 391. [Google Scholar] [CrossRef]
- Ni, J.; Yang, H.; Chen, L.; Xu, J.; Zheng, L.; Xie, G.; Shen, C.; Li, W.; Liu, Q. Metagenomic analysis of microbial community structure and function in a improved biofilter with odorous gases. Sci. Rep. 2022, 12, 1731. [Google Scholar] [CrossRef]
- Dong, Z.; Hu, L.; Li, J.; Kumwimba, M.N.; Tang, J.; Zhu, B. Nitrogen Retention in Mesocosm Sediments Received Rural Wastewater Associated with Microbial Community Response to Plant Species. Water 2020, 12, 3035. [Google Scholar] [CrossRef]
- Fan, X.-Y.; Gao, J.-F.; Pan, K.-L.; Li, D.-C.; Dai, H.-H.; Li, X. Temporal heterogeneity and temperature response of active ammonia-oxidizing microorganisms in winter in full-scale wastewater treatment plants. Chem. Eng. J. 2019, 360, 1542–1552. [Google Scholar] [CrossRef]
- Wu, F.; Zhang, Y.; He, D.; Gu, J.-D.; Guo, Q.; Liu, X.; Duan, Y.; Zhao, J.; Wang, W.; Feng, H. Community structures of bacteria and archaea associated with the biodeterioration of sandstone sculptures at the Beishiku Temple. Int. Biodeterior. Biodegrad. 2021, 164, 105290. [Google Scholar] [CrossRef]
- Archer, S.D.; McDonald, I.R.; Herbold, C.W.; Cary, S.C. Characterisation of bacterioplankton communities in the meltwater ponds of Bratina Island, Victoria Land, Antarctica. FEMS Microbiol. Ecol. 2014, 89, 451–464. [Google Scholar] [CrossRef] [Green Version]
- Valdespino-Castillo, P.M.; Cerqueda-Garcia, D.; Espinosa, A.C.; Batista, S.; Merino-Ibarra, M.; Tas, N.; Alcantara-Hernandez, R.J.; Falcon, L.I. Microbial distribution and turnover in Antarctic microbial mats highlight the relevance of heterotrophic bacteria in low-nutrient environments. FEMS Microbiol. Ecol. 2018, 94, fiy129. [Google Scholar] [CrossRef]








{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Site | MC (%) | TOC (%) | pH | NH4+-N (μg/g) | SiO42−-Si (μg/g) | NO3−-N (μg/g) | PO43−-P (μg/g) | NO2−-N (μg/g) | |
|---|---|---|---|---|---|---|---|---|---|
| BPK | Bpk1-1 | 22.55 | 0.024 | 7.69 | 0.1852 | 15.6632 | 0.5687 | 0.4938 | 0.0323 |
| Bpk1-2 | 22.01 | 0.138 | 7.62 | 0.1595 | 16.3551 | 0.6059 | 0.6860 | 0.0519 | |
| Bpk1-3 | 20.13 | 0.010 | 7.77 | 0.1774 | 13.3118 | 0.4997 | 0.5875 | 0.0269 | |
| Average | 21.56 ± 1.43 a | 0.057 ± 0.047 a | 7.69 ± 0.08 a | 0.1740 ± 0.0145 a | 15.1100 ± 1.7982 b | 0.5581 ± 0.0584 a | 0.5891 ± 0.0969 b | 0.037 1± 0.0148 a | |
| INT | Int1-1 | 24.80 | 0.004 | 6.91 | 0.0960 | 18.5088 | 0.5979 | 0.5165 | 0.0178 |
| Int1-2 | 14.95 | 0.022 | 6.74 | 0.1063 | 16.0028 | 0.5613 | 0.4836 | 0.0202 | |
| Int1-3 | 23.97 | 0.015 | 6.96 | 0.0966 | 19.3438 | 0.5076 | 0.6380 | 0.0185 | |
| Average | 21.24 ± 6.29 a | 0.014 ± 0.010 a | 6.87 ± 0.13 b | 0.0996 ± 0.0067 a | 17.9518 ± 1.9490 b | 0.5556 ± 0.048 a | 0.5460 ± 0.0920 b | 0.0188 ± 0.0014 b | |
| SED | Sed1-1 | 25.18 | 0.016 | 6.92 | 0.1037 | 22.8561 | 0.7280 | 0.8692 | 0.0113 |
| Sed1-2 | 23.90 | 0.085 | 7.20 | 0.3028 | 26.7257 | 0.5692 | 0.8407 | 0.0203 | |
| Sed1-3 | 20.91 | 0.012 | 7.09 | 0.2597 | 26.1310 | 0.4309 | 0.8116 | 0.0196 | |
| Average | 23.33 ± 2.42 a | 0.038 ± 0.047 a | 7.07 ± 0.15 b | 0.2221 ± 0.1184 a | 25.2376 ± 2.3815 a | 0.5761 ± 0.1519 a | 0.8405 ± 0.0289 a | 0.0171 ± 0.0058 b | |
| CCA1 | CCA2 | r2 | p | ||
|---|---|---|---|---|---|
| TOC | −0.98939 | −0.14529 | 0.1234 | 0.647 | - |
| MC | −0.81181 | −0.58392 | 0.0833 | 0.782 | - |
| pH | −0.74304 | 0.66925 | 0.7529 | 0.026 | * |
| NH4+ | −0.99353 | −0.11355 | 0.4000 | 0.207 | - |
| SiO42− | −0.34077 | −0.94015 | 0.4789 | 0.142 | - |
| NO3− | −0.25634 | −0.96659 | 0.0369 | 0.869 | - |
| PO43− | −0.60339 | −0.79744 | 0.8631 | 0.004 | ** |
| NO2− | −0.56632 | 0.82419 | 0.3120 | 0.284 | - |
| RDA1 | RDA2 | r2 | p | ||
|---|---|---|---|---|---|
| TOC | −0.80330 | −0.59557 | 0.4289 | 0.205 | - |
| MC | −0.20201 | 0.97938 | 0.0977 | 0.735 | - |
| pH | −0.99999 | −0.00319 | 0.6987 | 0.035 | * |
| NH4+ | −0.89564 | 0.44478 | 0.4669 | 0.160 | - |
| SiO42− | 0.42833 | 0.90362 | 0.1851 | 0.547 | - |
| NO3− | 0.63076 | 0.77598 | 0.1569 | 0.643 | - |
| PO43− | 0.08396 | 0.99647 | 0.1425 | 0.619 | - |
| NO2− | −0.72962 | −0.68386 | 0.6699 | 0.029 | * |
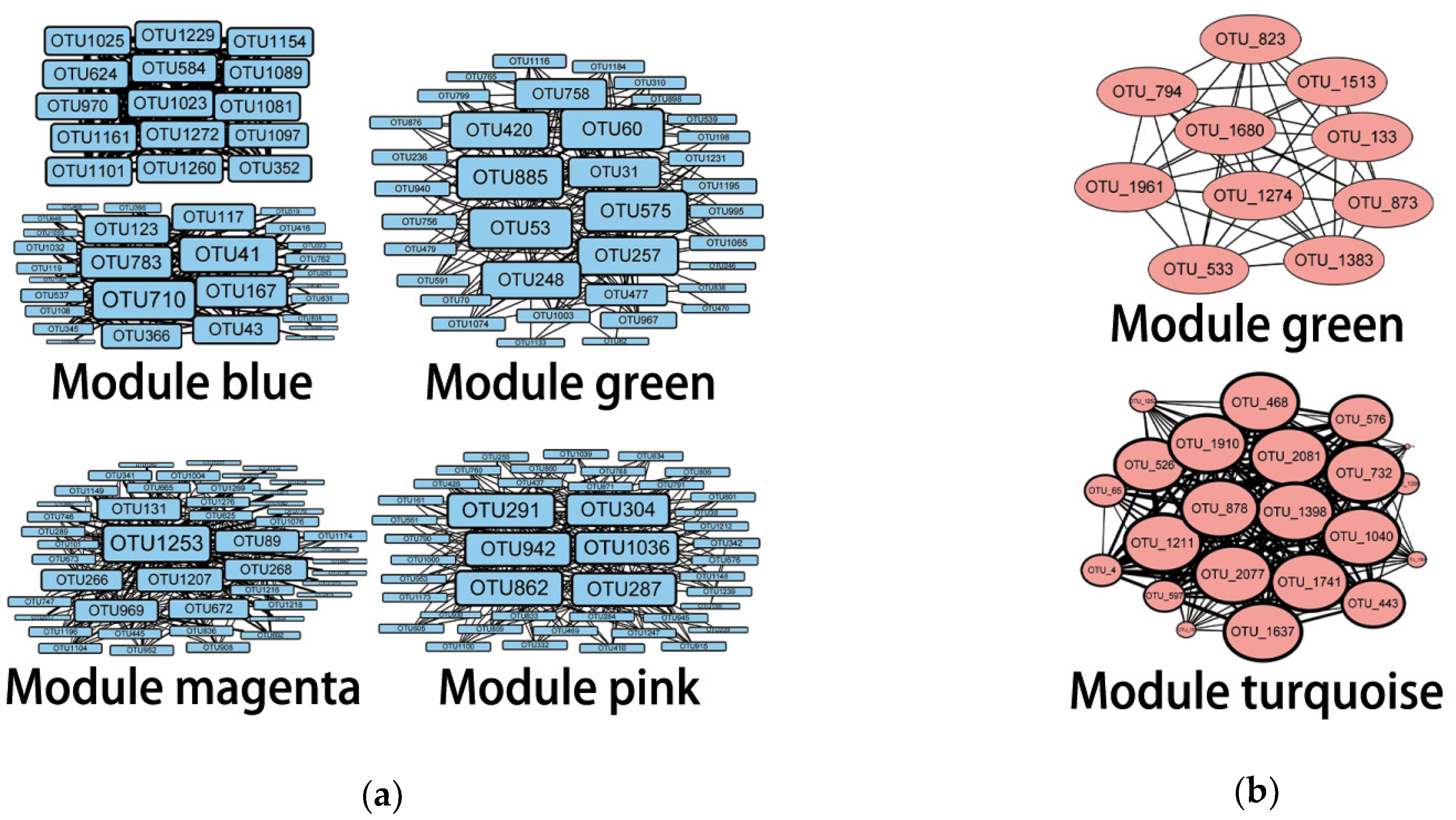
| Module | OTU Number | Assignment | Taxon |
|---|---|---|---|
| Bacterial module blue | OTU710 | Gaiellales | order |
| OTU41 | Actinobacteria MB-A2-108 | class | |
| OTU783 | Unclassified | - | |
| OTU167 | Sphingomonas | genus | |
| OTU123 | Betaproteobacteria SC-I-84 | order | |
| OTU43 | Blastocatellaceae RB41 | genus | |
| OTU1101 | Eisenbergiella | genus | |
| OTU1154 | Ruminococcaceae UCG-014 | genus | |
| OTU1260 | Brevundimonas | genus | |
| OTU584 | Unclassified | - | |
| OTU1272 | Roseburia | genus | |
| OTU1161 | Deinococcus | genus | |
| OTU1229 | Berkelbacteria | phylum | |
| OTU1081 | Ruminiclostridium 5 | genus | |
| OTU1025 | Fusobacterium | genus | |
| OTU1023 | Ruminococcaceae UCG-005 | genus | |
| OTU1089 | Ruminococcaceae UCG-014 | genus | |
| OTU624 | Lachnospiraceae NK4A136 group | genus | |
| Bacterial module green | OTU885 | Nitrospira | genus |
| OTU60 | Acidimicrobiaceae | family | |
| OTU53 | Ferruginibacter | genus | |
| OTU575 | Segetibacter | genus | |
| OTU257 | Verrucomicrobia OPB35 soil group | class | |
| Bacterial module magenta | OTU1253 | Desulfovibrio | genus |
| OTU1207 | Betaproteobacteria SC-I-84 | order | |
| OTU89 | Chthoniobacterales DA101 soil group | family | |
| OTU131 | Chloroflexi KD4-96 | class | |
| OTU969 | Verrucomicrobiaceae | family | |
| OTU268 | Hyphomicrobium | genus | |
| OTU266 | Holophagae Subgroup 7 | order | |
| OTU672 | Solibacteraceae | family | |
| Bacterial module pink | OTU291 | Xanthomonadales | order |
| OTU862 | Anaeromyxobacter | genus | |
| OTU942 | Unclassified | - | |
| OTU287 | Arsenophonus | genus | |
| OTU304 | Lachnospiraceae NK4A136 group | genus | |
| OTU1036 | Lachnospiraceae NK4A136 group | genus | |
| Archaeal module green | OTU_823 | Nitrososphaeria Group_1.1c | order |
| OTU_794 | Unknown | - | |
| OTU_1513 | Unknown | - | |
| OTU_1680 | Woesearchaeales | order | |
| OTU_133 | Nitrososphaeria | class | |
| OTU_1961 | Unknown | - | |
| OTU_1274 | Unknown | - | |
| OTU_873 | Unknown | - | |
| OTU_533 | Unknown | - | |
| OTU_1383 | Woesearchaeales | order | |
| Archaeal module turquoise | OTU_468 | Unknown | - |
| OTU_1910 | Unknown | - | |
| OTU_2081 | Methanomassiliicoccaceae | family | |
| OTU_732 | Archaea | kingdom | |
| OTU_878 | Micrarchaeales CG1-02-32-21 | family | |
| OTU_1398 | Woesearchaeales GW2011_GWC1_47_15 | family | |
| OTU_1040 | Woesearchaeales | order | |
| OTU_1211 | Woesearchaeales | order | |
| OTU_2077 | Woesearchaeales | order | |
| OTU_1741 | Woesearchaeales | order | |
| OTU_1637 | Methanosphaera | genus |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Li, Q.; Wang, N.; Han, W.; Zhang, B.; Zang, J.; Qin, Y.; Wang, L.; Liu, J.; Zhang, T. Soil Geochemical Properties Influencing the Diversity of Bacteria and Archaea in Soils of the Kitezh Lake Area, Antarctica. Biology 2022, 11, 1855. https://doi.org/10.3390/biology11121855
Li Q, Wang N, Han W, Zhang B, Zang J, Qin Y, Wang L, Liu J, Zhang T. Soil Geochemical Properties Influencing the Diversity of Bacteria and Archaea in Soils of the Kitezh Lake Area, Antarctica. Biology. 2022; 11(12):1855. https://doi.org/10.3390/biology11121855
Chicago/Turabian StyleLi, Qinxin, Nengfei Wang, Wenbing Han, Botao Zhang, Jiaye Zang, Yiling Qin, Long Wang, Jie Liu, and Tao Zhang. 2022. "Soil Geochemical Properties Influencing the Diversity of Bacteria and Archaea in Soils of the Kitezh Lake Area, Antarctica" Biology 11, no. 12: 1855. https://doi.org/10.3390/biology11121855