
Phyllanthus emblica (Amla) Fruit Powder as a Supplement to Improve Preweaning Dairy Calves’ Health: Effect on Antioxidant Capacity, Immune Response, and Gut Bacterial Diversity

Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Diet Preparation, Experimental Animals, and Treatments
2.2. Blood Sample Collection and Analysis
2.3. Rumen Fluid and Fecal Sample Collection
2.4. DNA Extraction, PCR Amplification, and 16S rRNA Sequencing
2.5. Sequencing Data Processing and Analysis
2.6. Statistical Analysis
3. Results
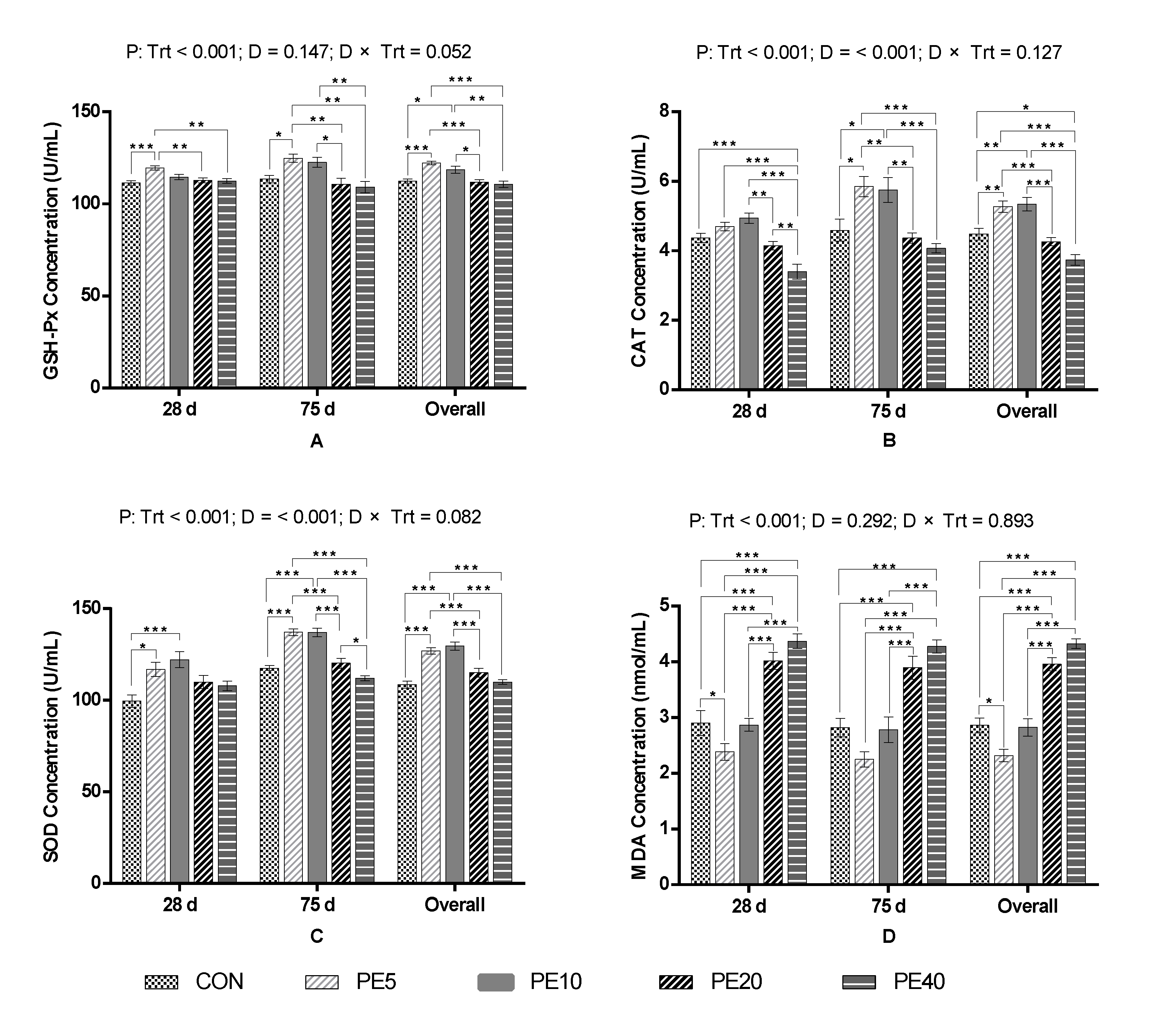
3.1. Antioxidant Capacity
3.2. Immune and Anti-Inflammatory Responses
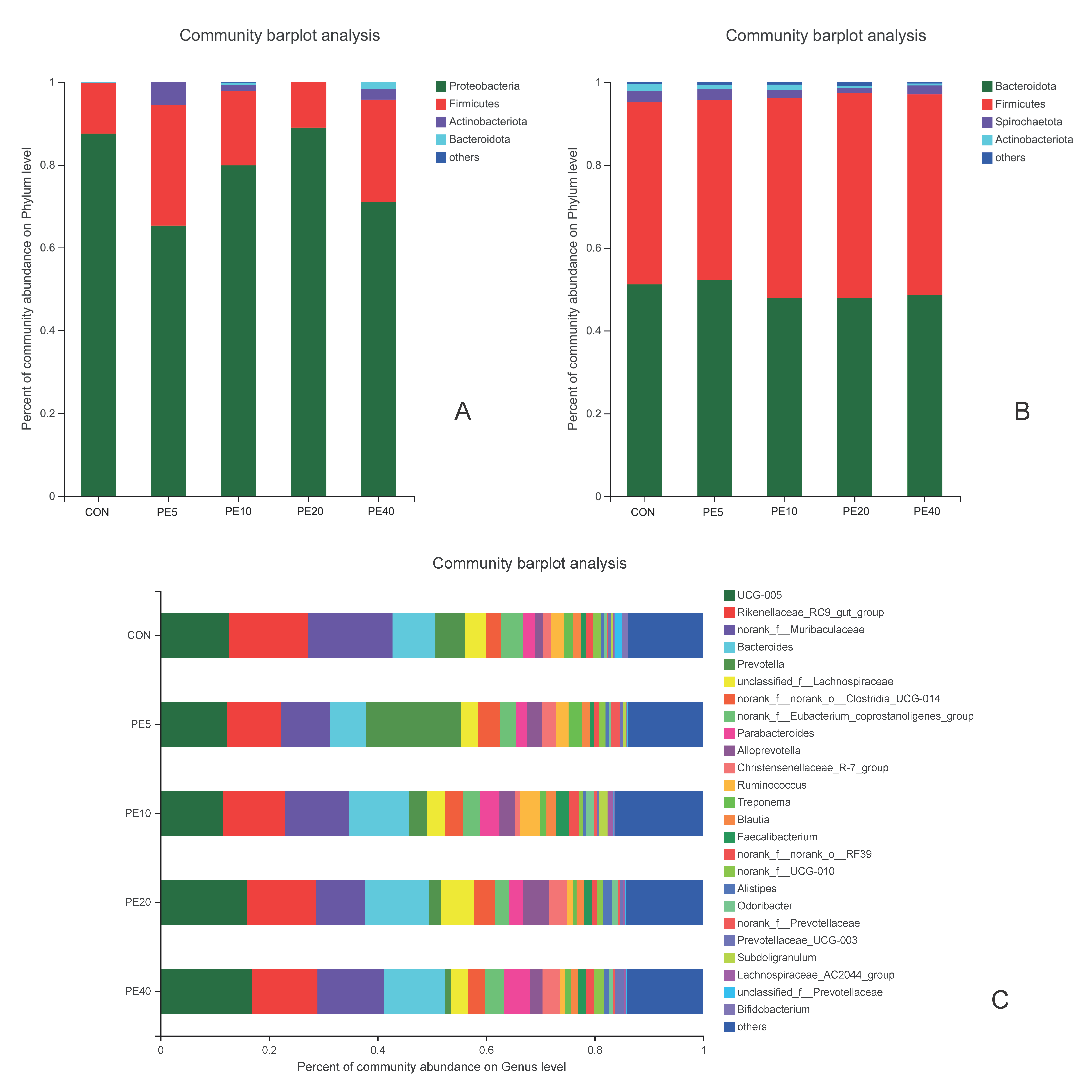
3.3. Ruminal and Fecal Bacterial Structure
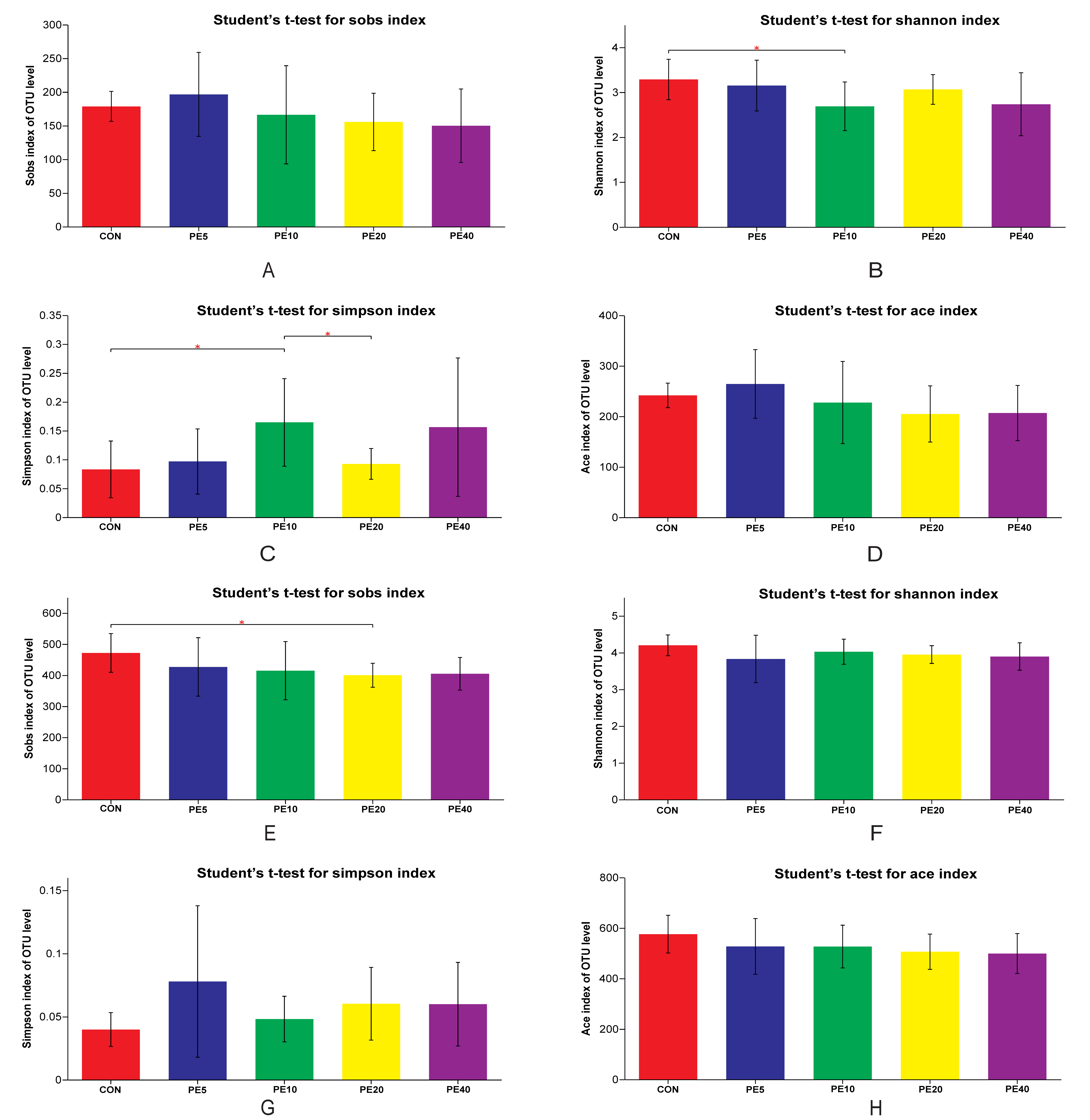
3.3.1. Diversity and Richness of Bacterial Flora
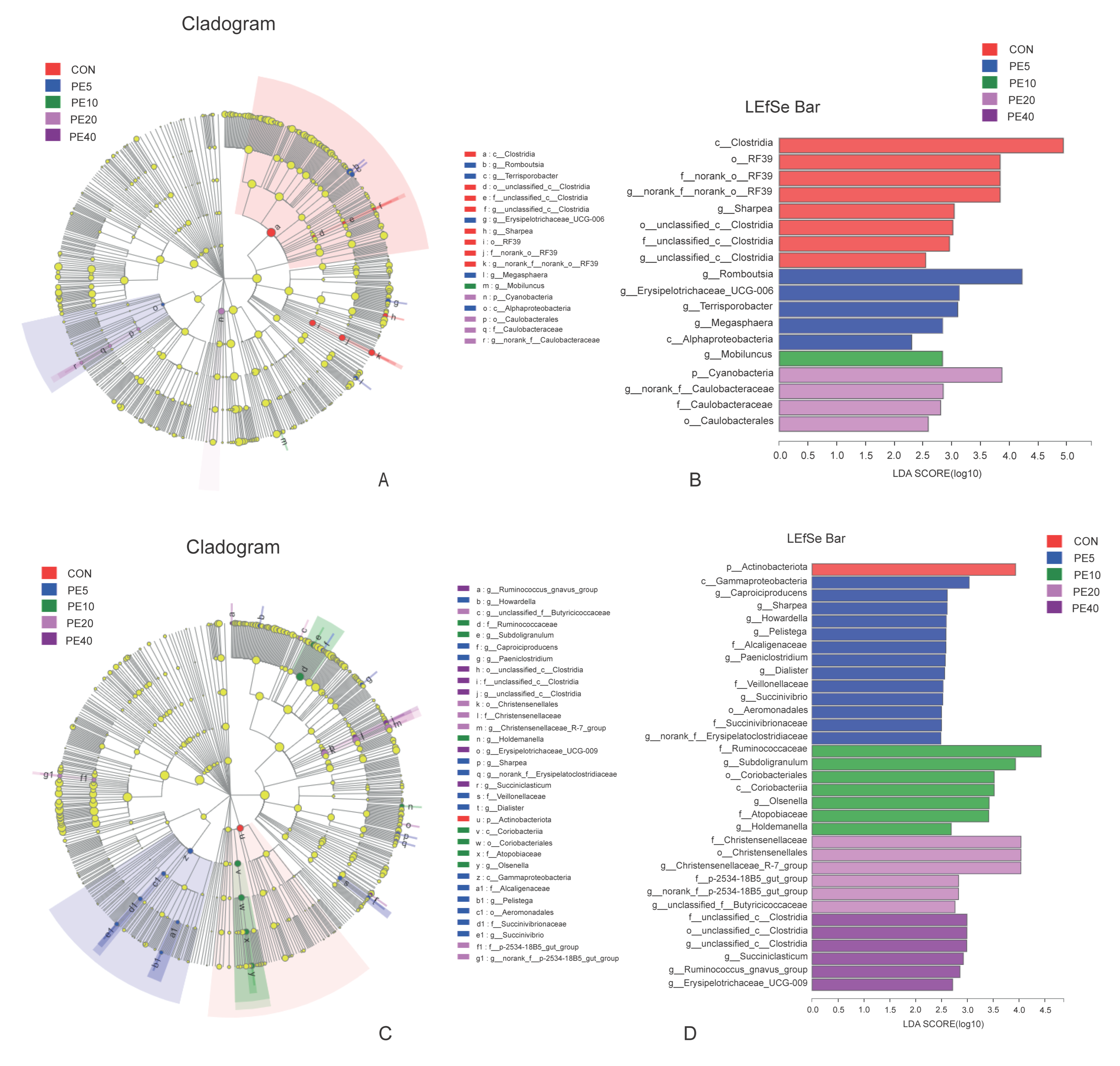
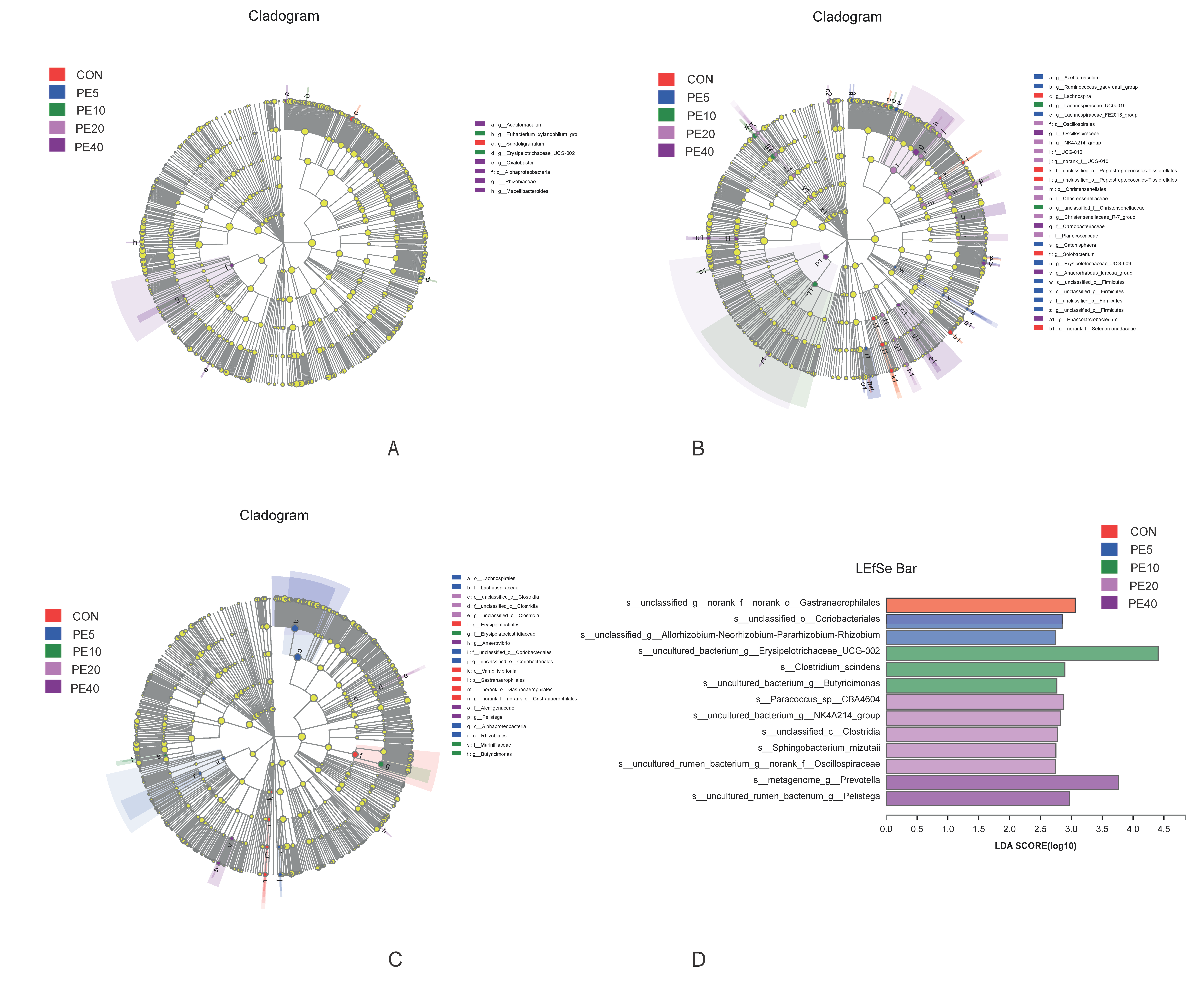
3.3.2. Difference in the Abundance of Bacterial Taxa among the Treatments
4. Discussion
4.1. Effect of Amla Fruit Powder on Antioxidant Capacity
4.2. Effect of Amla Fruit Powder on Immune and Anti-Inflammatory Responses
4.3. Effect of Amla Fruit Powder on Rumen and Fecal Bacterial Structure
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Izzo, M.M.; Kirkland, P.D.; Mohler, V.L.; Perkins, N.R.; Gunn, A.A.; House, J.K. Prevalence of Major Enteric Pathogens in Australian Dairy Calves with Diarrhoea. Aust. Vet. J. 2011, 89, 167–173. [Google Scholar] [CrossRef] [PubMed]
- Cho, Y.I.; Yoon, K.J. An Overview of Calf Diarrhea—Infectious Etiology, Diagnosis, and Intervention. J. Vet. Sci. 2014, 15, 1–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- USDA Dairy 2014: Health and Management Practices on, U.S. Dairy Operations, 2014. Report 3. 2018. Available online: https://www.aphis.usda.gov/animal_health/nahms/dairy/downloads/dairy14/Dairy14_dr_PartIII.pdf (accessed on 23 September 2022).
- Okello, E.; Williams, D.R.; ElAshmawy, W.R.; Adams, J.; Pereira, R.V.; Lehenbauer, T.W.; Aly, S.S. Survey on Antimicrobial Drug Use Practices in California Preweaned Dairy Calves. Front. Vet. Sci. 2021, 8, 636670. [Google Scholar] [CrossRef] [PubMed]
- Eibl, C.; Bexiga, R.; Viora, L.; Guyot, H.; Félix, J.; Wilms, J.; Tichy, A.; Hund, A. The Antibiotic Treatment of Calf Diarrhea in Four European Countries: A Survey. Antibiotics 2021, 10, 910. [Google Scholar] [CrossRef] [PubMed]
- Li, Z.; Hu, Y.; Yang, Y.; Lu, Z.; Wang, Y. Antimicrobial Resistance in Livestock: Antimicrobial Peptides Provide a New Solution for a Growing Challenge. Anim. Front. 2018, 8, 21–29. [Google Scholar] [CrossRef]
- Tedeschi, L.O.; Muir, J.P.; Naumann, H.D.; Norris, A.B.; Ramírez-Restrepo, C.A.; Mertens-Talcott, S.U. Nutritional Aspects of Ecologically Relevant Phytochemicals in Ruminant Production. Front. Vet. Sci. 2021, 8, 628445. [Google Scholar] [CrossRef]
- Peña-Torres, E.F.; González-Ríos, H.; Avendaño-Reyes, L.; Valenzuela-Grijalva, N.V.; Pinelli-Saavedra, A.; Muhlia-Almazán, A.; Peña-Ramos, E.A. Hydroxycinnamic Acids in Animal Production: Pharmacokinetics, Pharmacodynamics and Growth Promoting Effects. Review. Rev. Mex. Ciencias Pecu. 2019, 10, 391–415. [Google Scholar] [CrossRef]
- Gul, M.; Liu, Z.W.; Rabail, R.; Faheem, F.; Walayat, N.; Nawaz, A.; Shabbir, M.A.; Munekata, P.E.S.; Lorenzo, J.M.; Aadil, R.M. Functional and Nutraceutical Significance of Amla (Phyllanthus Emblica L.): A Review. Antioxidants 2022, 11, 816. [Google Scholar] [CrossRef]
- Krishnaveni, M.; Mirunalini, S. Therapeutic Potential of Phyllanthus Emblica (Amla): The Ayurvedic Wonder. J. Basic Clin. Physiol. Pharmacol. 2010, 21, 93–105. [Google Scholar] [CrossRef]
- Baliga, M.S.; Dsouza, J.J. Amla (Emblica Officinalis Gaertn), a Wonder Berry in the Treatment and Prevention of Cancer. Eur. J. Cancer Prev. 2011, 20, 225–239. [Google Scholar] [CrossRef]
- Chahal, A.K.; Chandan, G.; Kumar, R.; Chhillar, A.K.; Saini, A.K.; Saini, R.V. Bioactive Constituents of Emblica Officinalis Overcome Oxidative Stress in Mammalian Cells by Inhibiting Hyperoxidation of Peroxiredoxins. J. Food Biochem. 2020, 44, e13115. [Google Scholar] [CrossRef] [PubMed]
- Tilahun, M.; Zhao, L.; Sun, L.; Shen, Y.; Ma, L.; Callaway, T.R.; Xu, J.; Bu, D. Fresh Phyllanthus Emblica (Amla) Fruit Supplementation Enhances Milk Fatty Acid Profiles and the Antioxidant Capacities of Milk and Blood in Dairy Cows. Antioxidants 2022, 11, 485. [Google Scholar] [CrossRef] [PubMed]
- Sheoran, S.; Nidhi, P.; Kumar, V.; Singh, G.; Lal, U.R.; Sourirajan, A.; Dev, K. Altitudinal Variation in Gallic Acid Content in Fruits of Phyllanthus Emblica L. and Its Correlation with Antioxidant and Antimicrobial Activity. Vegetos 2019, 32, 387–396. [Google Scholar] [CrossRef]
- Saha, S.; Verma, R.J. Antioxidant Activity of Polyphenolic Extract of Phyllanthus Emblica against Lead Acetate Induced Oxidative Stress. Toxicol. Environ. Health Sci. 2015, 7, 82–90. [Google Scholar] [CrossRef]
- Nguse, M.; Yang, Y.; Fu, Z.; Xu, J.; Ma, L.; Bu, D. Amla (Phyllanthus Emblica) Fruit Powder Supplementation Decreased Frequency of Diarrhea and Fecal Score in Preweaning Dairy Calves. Anim. Feed Sci. Technol. 2022, Submitted. [Google Scholar]
- Dhivya Bharathi, M.; Justin-Thenmozhi, A.; Manivasagam, T.; Ahmad Rather, M.; Saravana Babu, C.; Mohamed Essa, M.; Guillemin, G.J. Amelioration of Aluminum Maltolate-Induced Inflammation and Endoplasmic Reticulum Stress-Mediated Apoptosis by Tannoid Principles of Emblica Officinalis in Neuronal Cellular Model. Neurotox. Res. 2019, 35, 318–330. [Google Scholar] [CrossRef]
- Li, W.; Zhang, X.; Chen, R.; Li, Y.; Miao, J.; Liu, G.; Lan, Y.; Chen, Y.; Cao, Y. HPLC Fingerprint Analysis of Phyllanthus Emblica Ethanol Extract and Their Antioxidant and Anti-Inflammatory Properties. J. Ethnopharmacol. 2020, 254, 112740. [Google Scholar] [CrossRef]
- Sai Ram, M.; Neetu, D.; Yogesh, B.; Anju, B.; Dipti, P.; Pauline, T.; Sharma, S.K.; Sarada, S.K.S.; Ilavazhagan, G.; Kumar, D.; et al. Cyto-Protective and Immunomodulating Properties of Amla (Emblica Officinalis) on Lymphocytes: An in-Vitro Study. J. Ethnopharmacol. 2002, 81, 5–10. [Google Scholar] [CrossRef]
- Singh, M.K.; Yadav, S.S.; Gupta, V.; Khattri, S. Immunomodulatory Role of Emblica Officinalis in Arsenic Induced Oxidative Damage and Apoptosis in Thymocytes of Mice. BMC Complement. Altern. Med. 2013, 13, 193. [Google Scholar] [CrossRef] [Green Version]
- Patel, P.; Singh, H.S.; Mishra, A.; Ansari, S.P.; Priyadershini, L.; Jain, A.K.; Ahirwar, M.K. Emblica Officinalis a Protective Herbal Supplementation to Prevent Buffalo Calf Mortality. Indian J. Dairy Sci. 2016, 69, 510–512. [Google Scholar]
- AOAC Official Methods of Analysis 17th Edition; The Association of Official Analytical Chemists: Gaithersburg, MD, USA, 2000.
- Sun, L.L.; Gao, S.T.; Wang, K.; Xu, J.C.; Sanz-Fernandez, M.V.; Baumgard, L.H.; Bu, D.P. Effects of Source on Bioavailability of Selenium, Antioxidant Status, and Performance in Lactating Dairy Cows during Oxidative Stress-Inducing Conditions. J. Dairy Sci. 2019, 102, 311–319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Soest, P.J.; Robertson, J.B.; Lewis, B.A. Methods for Dietary Fiber, Neutral Detergent Fiber, and Nonstarch Polysaccharides in Relation to Animal Nutrition. J. Dairy Sci. 1991, 74, 3583–3597. [Google Scholar] [CrossRef] [PubMed]
- Ren, Y.; Yu, G.; Shi, C.; Liu, L.; Guo, Q.; Han, C.; Zhang, D.; Zhang, L.; Liu, B.; Gao, H.; et al. Majorbio Cloud: A One-stop, Comprehensive Bioinformatic Platform for Multiomics Analyses. iMeta 2022, 1, e12. [Google Scholar] [CrossRef]
- Chen, S.; Zhou, Y.; Chen, Y.; Gu, J. Fastp: An Ultra-Fast All-in-One FASTQ Preprocessor. Bioinformatics 2018, 34, i884–i890. [Google Scholar] [CrossRef]
- Magoč, T.; Salzberg, S.L. FLASH: Fast Length Adjustment of Short Reads to Improve Genome Assemblies. Bioinformatics 2011, 27, 2957–2963. [Google Scholar] [CrossRef] [Green Version]
- Edgar, R.C. UPARSE: Highly Accurate OTU Sequences from Microbial Amplicon Reads. Nat. Methods 2013, 10, 996–998. [Google Scholar] [CrossRef]
- Stackebrandt, E.; Goebel, B.M. Taxonomic Note: A Place for DNA-DNA Reassociation and 16S rRNA Sequence Analysis in the Present Species Definition in Bacteriology. Int. J. Syst. Bacteriol. 1994, 44, 846–849. [Google Scholar] [CrossRef] [Green Version]
- Wang, Q.; Garrity, G.M.; Tiedje, J.M.; Cole, J.R. Naïve Bayesian Classifier for Rapid Assignment of rRNA Sequences into the New Bacterial Taxonomy. Appl. Environ. Microbiol. 2007, 73, 5261–5267. [Google Scholar] [CrossRef] [Green Version]
- Lobo, V.; Patil, A.; Phatak, A.; Chandra, N. Free Radicals, Antioxidants and Functional Foods: Impact on Human Health. Pharmacogn. Rev. 2010, 4, 118–126. [Google Scholar] [CrossRef] [Green Version]
- Pham-Huy, L.A.; He, H.; Pham-Huy, C. Free Radicals, Antioxidants in Disease and Health. Int. J. Biomed. Sci. 2008, 4, 89–96. [Google Scholar]
- Poljsak, B.; Šuput, D.; Milisav, I. Achieving the Balance between ROS and Antioxidants: When to Use the Synthetic Antioxidants. Oxid. Med. Cell. Longev. 2013, 2013, 956792. [Google Scholar] [CrossRef] [PubMed]
- Pamplona, R.; Costantini, D. Molecular and Structural Antioxidant Defenses against Oxidative Stress in Animals. Am. J. Physiol.-Regul. Integr. Comp. Physiol. 2011, 301, R843–R863. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beaulieu, M.; Costantini, D. Biomarkers of Oxidative Status: Missing Tools in Conservation Physiology. Conserv. Physiol. 2014, 2, cou014. [Google Scholar] [CrossRef] [PubMed]
- Katerji, M.; Filippova, M.; Duerksen-Hughes, P. Approaches and Methods to Measure Oxidative Stress in Clinical Samples: Research Applications in the Cancer Field. Oxid. Med. Cell. Longev. 2019, 2019, 1279250. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Patekar, D.; Kheur, S.; Bagul, N.; Kulkarni, M.; Mahalle, A.; Ingle, Y.; Dhas, V. Antioxidant Defence System. Oral Maxillofac. Pathol. J. 2013, 4, 309–315. [Google Scholar]
- Surai, P.F.; Kochish, I.I.; Fisinin, V.I.; Kidd, M.T. Antioxidant Defence Systems and Oxidative Stress in Poultry Biology: An Update. Antioxidants 2019, 8, 235. [Google Scholar] [CrossRef] [Green Version]
- Rajak, S.; Banerjee, S.K.; Sood, S.; Dinda, A.K.; Gupta, Y.K.; Gupta, S.K.; Maulik, S.K. Emblica Officinalis Causes Myocardial Adaptation and Protects against Oxidative Stress in Ischemic-Reperfusion Injury in Rats. Phyther. Res. 2004, 18, 54–60. [Google Scholar] [CrossRef]
- Shivananjappa, M.M.; Joshi, M.K. Influence of Emblica Officinalis Aqueous Extract on Growth and Antioxidant Defense System of Human Hepatoma Cell Line (HepG2). Pharm. Biol. 2012, 50, 497–505. [Google Scholar] [CrossRef]
- Charoenteeraboon, J.; Ngamkitidechakul, C.; Soonthornchareonnon, N.; Jaijoy, K.; Sireeratawong, S. Antioxidant Activities of the Standardized Water Extract from Fruit of Phyllanthus Emblica Linn. Songklanakarin J. Sci. Technol. 2010, 32, 599–604. [Google Scholar]
- Usharani, P.; Fatima, N.; Muralidhar, N. Effects of Phyllanthus Emblica Extract on Endothelial Dysfunction and Biomarkers of Oxidative Stress in Patients with Type 2 Diabetes Mellitus: A Randomized, Double-Blind, Controlled Study. Diabetes Metab. Syndr. Obes. Targets Ther. 2013, 6, 275–284. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, A.; Bandyopadhyay, S.K.; Chattopadhyay, S. Biphasic Effect of Phyllanthus Emblica L. Extract on NSAID-Induced Ulcer: An Antioxidative Trail Weaved with Immunomodulatory Effect. Evid.-Based Complement. Altern. Med. 2011, 2011, 146808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reddy, V.D.; Padmavathi, P.; Gopi, S.; Paramahamsa, M.; Varadacharyulu, N.C. Protective Effect of Emblica Officinalis against Alcohol-Induced Hepatic Injury by Ameliorating Oxidative Stress in Rats. Indian J. Clin. Biochem. 2010, 25, 419–424. [Google Scholar] [CrossRef] [Green Version]
- Karamać, M. Chelation of Cu(II), Zn(II), and Fe(II) by Tannin Constituents of Selected Edible Nuts. Int. J. Mol. Sci. 2009, 10, 5485–5497. [Google Scholar] [CrossRef] [PubMed]
- Sieniawska, E. Activities of Tannins-From in Vitro Studies to Clinical Trials. Nat. Prod. Commun. 2015, 10, 1877–1884. [Google Scholar] [CrossRef] [Green Version]
- Koleckar, V.; Kubikova, K.; Rehakova, Z.; Kuca, K.; Jun, D.; Jahodar, L.; Opletal, L. Condensed and Hydrolysable Tannins as Antioxidants Influencing the Health. Mini-Rev. Med. Chem. 2008, 8, 436–447. [Google Scholar] [CrossRef] [PubMed]
- Nambiar, S.S.; Paramesha, M.; Shetty, N.P. Comparative Analysis of Phytochemical Profile, Antioxidant Activities and Foam Prevention Abilities of Whole Fruit, Pulp and Seeds of Emblica Officinalis. J. Food Sci. Technol. 2015, 52, 7254–7262. [Google Scholar] [CrossRef]
- Sriwatcharakul, S. Evaluation of Bioactivities of Phyllanthus Emblica Seed. Energy Rep. 2020, 6, 442–447. [Google Scholar] [CrossRef]
- Middha, S.K.; Goyal, A.K.; Lokesh, P.; Yardi, V.; Mojamdar, L.; Keni, D.S.; Babu, D.; Usha, T. Toxicological Evaluation of Emblica Officinalis Fruit Extract and Its Anti-Inflammatory and Free Radical Scavenging Properties. Pharmacogn. Mag. 2015, 11, S427–S433. [Google Scholar] [CrossRef] [Green Version]
- Yang, B.; Liu, P. Composition and Biological Activities of Hydrolyzable Tannins of Fruits of Phyllanthus Emblica. J. Agric. Food Chem. 2014, 62, 529–541. [Google Scholar] [CrossRef]
- Ogra, P.L.; Karzon, D.T. The Role of Immunoglulins in the Mechanism of Mucosal Immunity to Virus Infection. Pediatr. Clin. N. Am. 1970, 17, 385–400. [Google Scholar] [CrossRef]
- Kaleem, Q.M.; Akhtar, M.; Awais, M.M.; Saleem, M.; Zafar, M.; Iqbal, Z.; Muhammad, F.; Anwar, M.I. Studies on Emblica Officinalis Derived Tannins for Their Immunostimulatory and Protective Activities against Coccidiosis in Industrial Broiler Chickens. Sci. World J. 2014, 2014, 378473. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Snoep, J.J.; Potters, J.B. Coccidiosis Causes Diarrhea in Calves in the Pasture. Pasture Coccidiosis Caused by Eimeria Alabamensis. Tijdschr. Diergeneeskd. 2004, 129, 158–160. [Google Scholar] [PubMed]
- Ramírez-Restrepo, C.A.; Pernthaner, A.; Barry, T.N.; López-Villalobos, N.; Shaw, R.J.; Pomroy, W.E.; Hein, W.R. Characterization of Immune Responses against Gastrointestinal Nematodes in Weaned Lambs Grazing Willow Fodder Blocks. Anim. Feed Sci. Technol. 2010, 155, 99–110. [Google Scholar] [CrossRef]
- Ramírez-Restrepo, C.A.; Barry, T.N.; López-Villalobos, N.; Kemp, P.D.; Harvey, T.G. Use of Lotus Corniculatus Containing Condensed Tannins to Increase Reproductive Efficiency in Ewes under Commercial Dryland Farming Conditions. Anim. Feed Sci. Technol. 2005, 121, 23–43. [Google Scholar] [CrossRef]
- Reed, J.D. Nutritional Toxicology of Tannins and Related Polyphenols in Forage Legumes. J. Anim. Sci. 1995, 73, 1516–1528. [Google Scholar] [CrossRef]
- Nicolis, E.; Lampronti, I.; Dechecchi, M.C.; Borgatti, M.; Tamanini, A.; Bianchi, N.; Bezzerri, V.; Mancini, I.; Grazia Giri, M.; Rizzotti, P.; et al. Pyrogallol, an Active Compound from the Medicinal Plant Emblica Officinalis, Regulates Expression of pro-Inflammatory Genes in Bronchial Epithelial Cells. Int. Immunopharmacol. 2008, 8, 1672–1680. [Google Scholar] [CrossRef]
- Gensollen, T.; Iyer, S.S.; Kasper, D.L.; Blumberg, R.S. How Colonization by Microbiota in Early Life Shapes the Immune system. Science 2016, 352, 539–544. [Google Scholar] [CrossRef] [Green Version]
- Fu, Y.; He, Y.; Xiang, K.; Zhao, C.; He, Z.; Qiu, M.; Hu, X. The Role of Rumen Microbiota and Its Metabolites in Subacute Ruminal Acidosis (SARA)-Induced Inflammatory Diseases of Ruminants. Microorganisms 2022, 10, 1495. [Google Scholar] [CrossRef]
- Gomez, D.E.; Galvão, K.N.; Rodriguez-Lecompte, J.C.; Costa, M.C. The Cattle Microbiota and the Immune System: An Evolving Field. Vet. Clin. North Am.-Food Anim. Pract. 2019, 35, 485–505. [Google Scholar] [CrossRef]
- Ott, S.J.; Musfeldt, M.; Wenderoth, D.F.; Hampe, J.; Brant, O.; Fölsch, U.R.; Timmis, K.N.; Schreiber, S. Reduction in Diversity of the Colonic Mucosa Associated Bacterial Microflora in Patients with Active Inflammatory Bowel Disease. Gut 2004, 53, 685–693. [Google Scholar] [CrossRef] [Green Version]
- Andersen, K.; Kesper, M.S.; Marschner, J.A.; Konrad, L.; Ryu, M.; Kumar, S.V.R.; Kulkarni, O.P.; Mulay, S.R.; Romoli, S.; Demleitner, J.; et al. Intestinal Dysbiosis, Barrier Dysfunction, and Bacterial Translocation Account for CKD-Related Systemic Inflammation. J. Am. Soc. Nephrol. 2017, 28, 76–83. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Garcia, M.; Bradford, B.J.; Nagaraja, T.G. INVITED REVIEW: Ruminal Microbes, Microbial Products, and Systemic Inflammation. Prof. Anim. Sci. 2017, 33, 635–650. [Google Scholar] [CrossRef]
- Kosiewicz, M.M.; Zirnheld, A.L.; Alard, P. Gut Microbiota, Immunity, and Disease: A Complex Relationship. Front. Microbiol. 2011, 2, 180. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheng, H.Y.; Ning, M.X.; Chen, D.K.; Ma, W.T. Interactions between the Gut Microbiota and the Host Innate Immune Response against Pathogens. Front. Immunol. 2019, 10, 607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ze, X.; Duncan, S.H.; Louis, P.; Flint, H.J. Ruminococcus Bromii Is a Keystone Species for the Degradation of Resistant Starch in the Human Colon. ISME J. 2012, 6, 1535–1543. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ottman, N.; Smidt, H.; de Vos, W.M.; Belzer, C. The Function of Our Microbiota: Who is out there and what do they do? Front. Cell. Infect. Microbiol. 2012, 2, 104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Frank, D.N.; St. Amand, A.L.; Feldman, R.A.; Boedeker, E.C.; Harpaz, N.; Pace, N.R. Molecular-Phylogenetic Characterization of Microbial Community Imbalances in Human Inflammatory Bowel Diseases. Proc. Natl. Acad. Sci. USA 2007, 104, 13780–13785. [Google Scholar] [CrossRef] [Green Version]
- Schogor, A.L.B.; Huws, S.A.; Santos, G.T.D.; Scollan, N.D.; Hauck, B.D.; Winters, A.L.; Kim, E.J.; Petit, H.V. Ruminal Prevotella spp. May Play an Important Role in the Conversion of Plant Lignans into Human Health Beneficial Antioxidants. PLoS ONE 2014, 9, e87949. [Google Scholar] [CrossRef] [Green Version]
- Khafipour, E.; Li, S.; Plaizier, J.C.; Krause, D.O. Rumen Microbiome Composition Determined Using Two Nutritional Models of Subacute Ruminal Acidosis. Appl. Environ. Microbiol. 2009, 75, 7115–7124. [Google Scholar] [CrossRef] [Green Version]
- Aguilar-Marin, S.B.; Betancur-Murillo, C.L.; Isaza, G.A.; Mesa, H.; Jovel, J. Lower Methane Emissions Were Associated with Higher Abundance of Ruminal Prevotella in a Cohort of Colombian Buffalos. BMC Microbiol. 2020, 20, 364. [Google Scholar] [CrossRef]
- Shen, X.; Miao, J.; Wan, Q.; Wang, S.; Li, M.; Pu, F.; Wang, G.; Qian, W.; Yu, Q.; Marotta, F.; et al. Possible Correlation between Gut Microbiota and Immunity among Healthy Middle-Aged and Elderly People in Southwest China. Gut Pathog. 2018, 10, 4. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Items | Calf Starter 1 | Whole Milk | Milk Replacer 1 | Amla (Phyllanthus emblica) 1 |
|---|---|---|---|---|
| Ingredients (% of DM): | - | - | - | |
| Steam-flaked corn | 40.50 | - | - | - |
| Corn gluten | 2.54 | - | - | - |
| Soybean meal | 20.51 | - | - | - |
| Extruded soybean | 6.10 | - | - | - |
| Wheat bran | 5.86 | - | - | - |
| Wheat shorts | 7.12 | - | - | - |
| Canola meal | 11.78 | - | - | - |
| Cane molasses | 1.66 | - | - | - |
| Calf starter premix 2 | 3.93 | - | - | - |
| Chemical composition: | - | - | - | |
| DM (%) | 92.90 | - | 95.79 | 93.53 |
| Ash (%) | 6.49 | - | 7.22 | 3.54 |
| CP (%) | 23.88 | - | 22.45 | 6.64 |
| EE (%) | 3.19 | - | 8.99 | 1.22 |
| NDF (%) | 16.89 | - | - | 36.25 |
| ADF (%) | 6.93 | - | - | 28.15 |
| Density (g/L) | - | 1030.78 | - | - |
| Milk protein (%) | - | 3.54 | - | - |
| Milk fat (%) | - | 3.76 | - | - |
| Total solid (%) | - | 12.90 | - | - |
| Lactose (%) | - | 4.70 | - | - |
| Items 1 | Treatments 2 | SEM | p-Value 3 | ||||||
|---|---|---|---|---|---|---|---|---|---|
| CON | PE5 | PE10 | PE20 | PE40 | Trt | D | Trt × D | ||
| IgA (µg/mL) | |||||||||
| Overall | 651.1 b | 723.1 a | 710.3 a | 616.8 bc | 601.7 c | 11.63 | <0.001 | 0.099 | 0.294 |
| D28 | 641.8 ab | 687.3 a | 688.2 a | 615.6 b | 610.9 b | 15.92 | 0.003 | ||
| D75 | 660.4 bc | 758.9 a | 732.5 ab | 617.9 c | 592.4 c | 22.22 | <0.001 | ||
| IgG (µg/mL) | |||||||||
| Overall | 6221.1 b | 6977.5 a | 6830.2 a | 5743.6 c | 5729.8 c | 112.14 | <0.001 | 0.263 | 0.379 |
| D28 | 6142.3 bc | 6757.0 a | 6715.9 ab | 5915.5 c | 5698.1 c | 143.93 | <0.001 | ||
| D75 | 6316.6 bc | 7197.9 a | 6944.4 ab | 5605.2 c | 5761.6 bc | 198.49 | <0.001 | ||
| IgM (µg/mL) | |||||||||
| Overall | 200.7 b | 227.9 a | 222.7 a | 192.6 b | 188.0 b | 4.29 | <0.001 | 0.017 | 0.191 |
| D28 | 190.4 bc | 216.8 a | 215.6 ab | 193.9 abc | 188.6 c | 6.14 | 0.004 | ||
| D75 | 211.1 bc | 138.9 a | 229.7 ab | 191.1 c | 187.5 c | 6.45 | <0.001 | ||
| TNF-α (µg/mL) | |||||||||
| Overall | 0.361 bc | 0.252 c | 0.280 c | 0.446 ab | 0.480 a | 0.03 | <0.001 | 0.123 | 0.974 |
| D28 | 0.333 ab | 0.236 b | 0.254 b | 0.427 ab | 0.475 a | 0.05 | 0.008 | ||
| D75 | 0.389 ab | 0.267 b | 0.306 b | 0.468 a | 0.485 a | 0.03 | <0.001 | ||
| IL-10 (pg/mL) | |||||||||
| Overall | 38.80 bc | 52.21 a | 46.78 ab | 39.42 bc | 35.57 c | 1.93 | <0.001 | 0.486 | 0.584 |
| D28 | 34.71 b | 51.45 a | 47.99 ab | 38.68 ab | 36.14 ab | 2.70 | 0.011 | ||
| D75 | 42.90 bc | 52.97 a | 45.57 ab | 40.15 bc | 34.99 c | 2.32 | <0.001 | ||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Nguse, M.; Yang, Y.; Fu, Z.; Xu, J.; Ma, L.; Bu, D. Phyllanthus emblica (Amla) Fruit Powder as a Supplement to Improve Preweaning Dairy Calves’ Health: Effect on Antioxidant Capacity, Immune Response, and Gut Bacterial Diversity. Biology 2022, 11, 1753. https://doi.org/10.3390/biology11121753
Nguse M, Yang Y, Fu Z, Xu J, Ma L, Bu D. Phyllanthus emblica (Amla) Fruit Powder as a Supplement to Improve Preweaning Dairy Calves’ Health: Effect on Antioxidant Capacity, Immune Response, and Gut Bacterial Diversity. Biology. 2022; 11(12):1753. https://doi.org/10.3390/biology11121753
Chicago/Turabian StyleNguse, Mebrahtom, Yi Yang, Zilin Fu, Jianchu Xu, Lu Ma, and Dengpan Bu. 2022. "Phyllanthus emblica (Amla) Fruit Powder as a Supplement to Improve Preweaning Dairy Calves’ Health: Effect on Antioxidant Capacity, Immune Response, and Gut Bacterial Diversity" Biology 11, no. 12: 1753. https://doi.org/10.3390/biology11121753