
Current Genetic Structure Analysis of Leopard Cats Reveals a Weak Disparity Trend in Subpopulations in Beijing, China
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
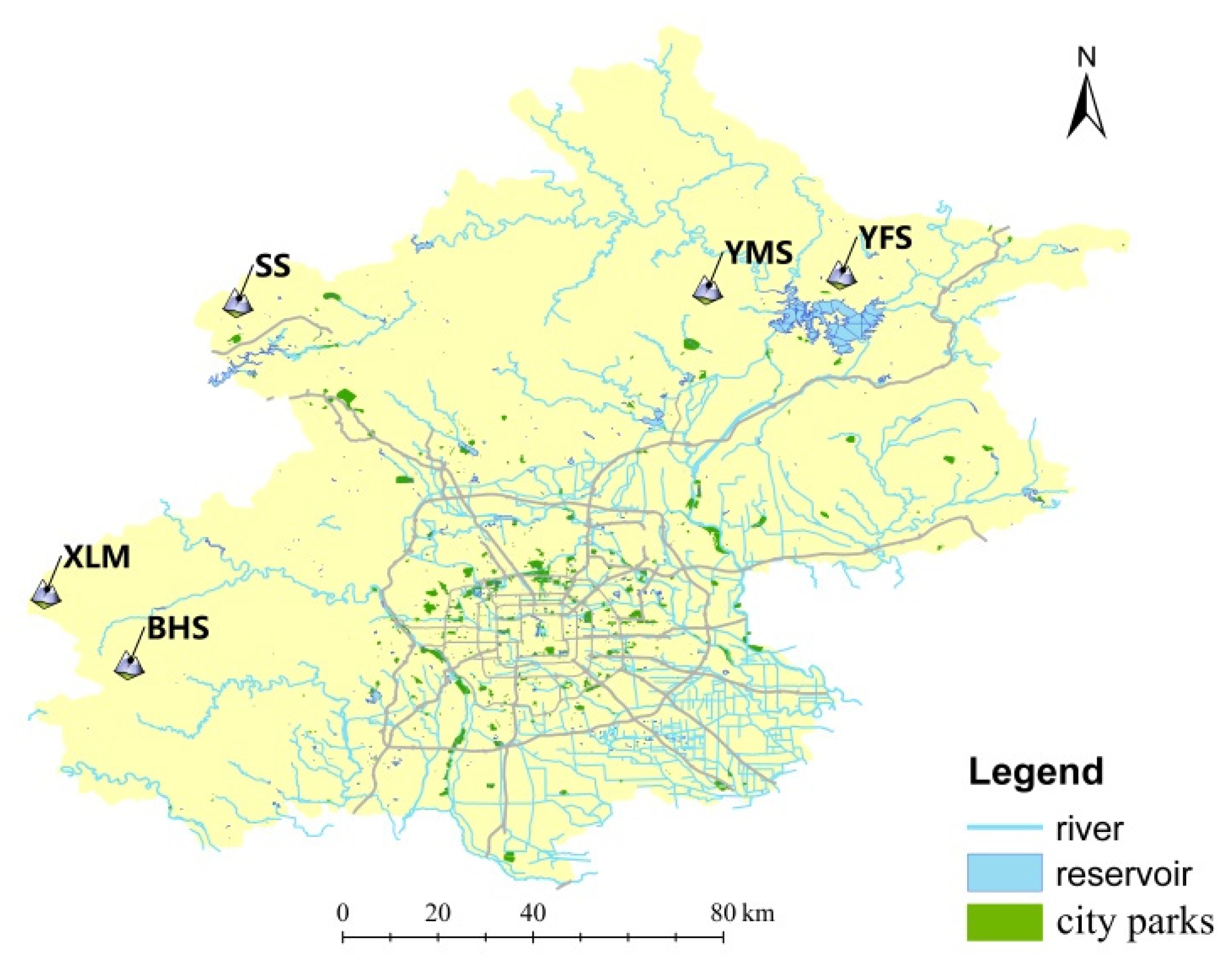
2.1. Sampling Procedure and DNA Extraction
2.2. Species and Sex Identification
2.3. Microsatellite Loci Selection and Amplification
2.4. Data Analysis
3. Results
3.1. Species and Individual Identification
3.2. Microsatellite Analysis of Genetic Diversity
4. Discussion
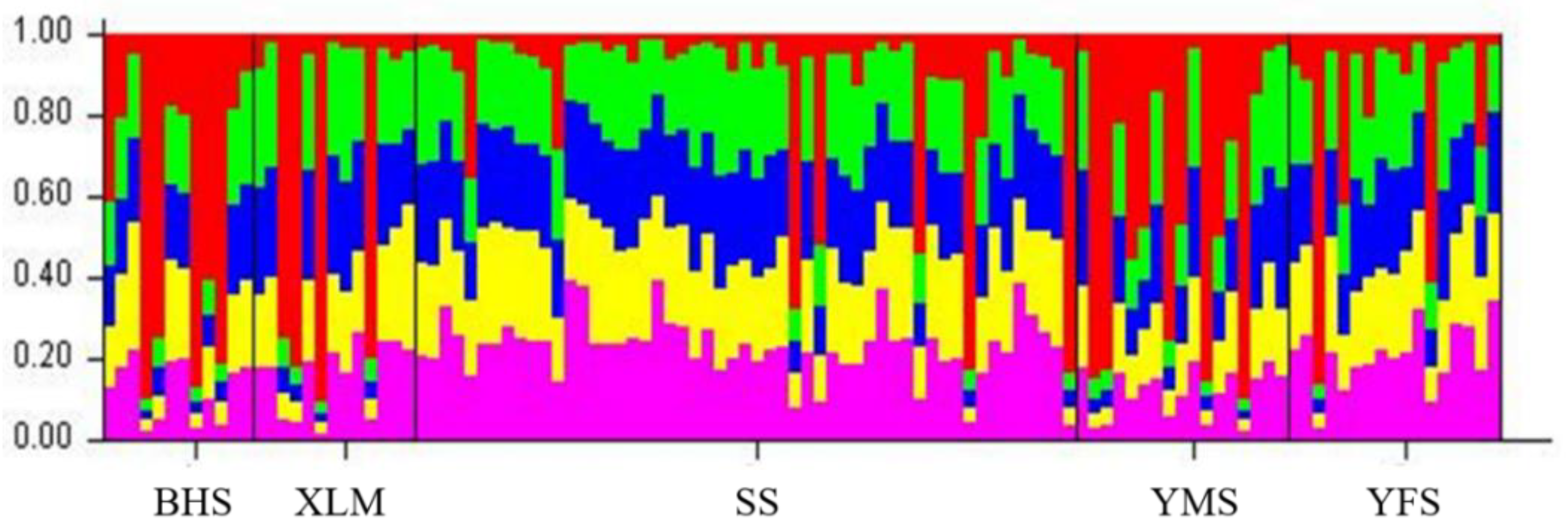
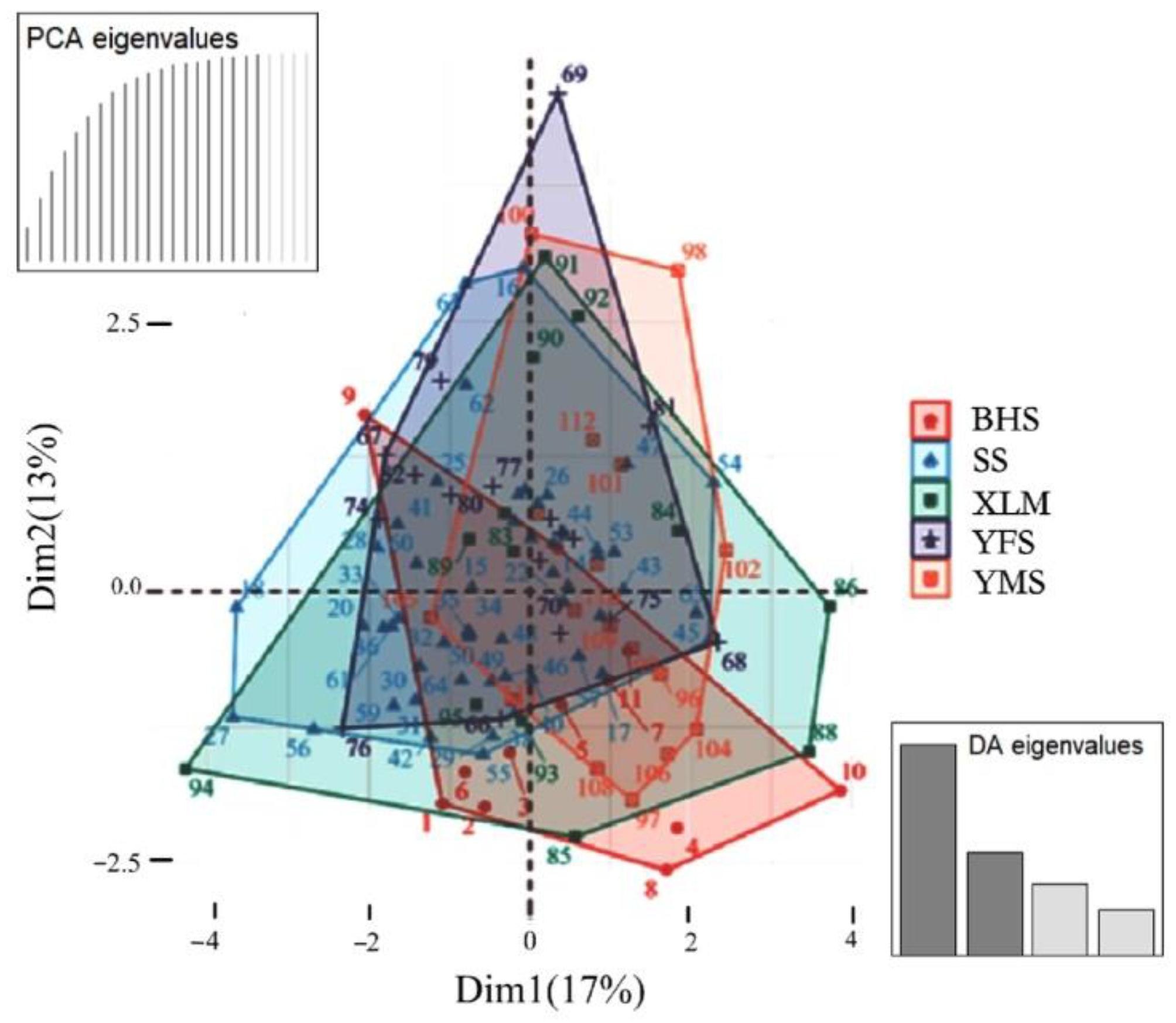
4.1. Genetic Structure Based on Microsatellite Diversity
4.2. Population Genetic Differentiation
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Hogbin, P.M.; Peakall, R. Evaluation of the contribution of genetic research to the management of the endangered plant Zieria prostrata. Conserv. Biol. 2010, 13, 514–522. [Google Scholar] [CrossRef]
- Romiguier, J.; Gayral, P.; Ballenghien, M.; Bernard, A.; Cahais, V.; Chenuil, A.; Chiari, A.; Dernat, Y.; Duret, L.; Faivre, N.; et al. Comparative population genomics in animals uncovers the determinants of genetic diversity. Nature 2014, 515, 261–263. [Google Scholar] [CrossRef]
- Sharma, R.; Stuckas, H.; Bhaskar, R.; Khan, I.; Goyal, S.P.; Tiedemann, R. Genetically distinct population of Bengal tiger (Panthera tigris tigris) in Terai Arc landscape (TAL) of India. Mamm. Biol. 2011, 76, 484–490. [Google Scholar] [CrossRef]
- Lino, A.; Fonseca, C.; Rojas, D.; Fischer, E.; Pereira, M.J.R. A meta-analysis of the effects of habitat loss and fragmentation on genetic diversity in mammals. Mamm. Biol. 2018, 94, 69–76. [Google Scholar] [CrossRef]
- Mukherjee, S.; Krishnan, A.; Tamma, K.; Home, C.; Navya, R.; Joseph, S.; Das, A.; Ramakrishnan, U. Ecology Driving Genetic Variation: A Comparative Phylogeography of Jungle Cat (Felis chaus) and Leopard Cat (Prionailurus bengalensis) in India. PLoS ONE 2010, 5, e13724. [Google Scholar] [CrossRef] [PubMed]
- Fernández, N.; Delibes, M.; Palomares, F. Landscape evaluation in conservation: Molecular sampling and habitat modeling for the Iberian lynx. Ecol. Appl. 2006, 16, 1037–1049. [Google Scholar] [CrossRef] [Green Version]
- Sunquist, M.; Sunquist, F. Leopard Cat (Prionailurus bengalensis) Kerr, 1792, Wild Cats of the World; University of Chicago Press: Chicago, IL, USA, 2002; pp. 1–14. [Google Scholar]
- Farrell, L.E.; Roman, J.; Sunquist, M.E. Dietary separation of sympatric carnivores identified by molecular analysis of scats. Mol. Ecol. 2000, 9, 1583–1590. [Google Scholar] [CrossRef] [PubMed]
- Ripple, W.J.; Estes, J.A.; Beschta, R.L.; Wilmers, C.C.; Ritchie, E.G.; Hebblewhite, M.; Berger, J.; Elmhagen, B.; Letnic, M.; Nelson, M.P.; et al. Status and ecological effects of the world’s largest carnivores. Science 2014, 343, 1241484. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Rodgers, T.W.; Janečka, J.E. Applications and techniques for non-invasive faecal genetics research in felid conservation. Eur. J. Wildl. Res. 2012, 59, 1–16. [Google Scholar] [CrossRef]
- Macdonald, D.; Loveridge, A. The Biology and Conservation of Wild Felids; Oxford University Press: London, UK, 2010; pp. 11–23. [Google Scholar]
- Caragiulo, A.; Kang, Y.; Rabinowitz, S.; Freedman, I.D.; Simone, L.; Zhou, X.W.; Bao, W.D.; Amato, G. Presence of the endangered Amur tiger Panthera tigris altaica in Jilin Province, China, detected using non-invasive genetic techniques. Oryx 2015, 49, 632–635. [Google Scholar] [CrossRef]
- Kurose, N.; Masuda, R.; Tatara, M. Fecal DNA analysis for identifying species and sex of sympatric carnivores: A noninvasive method for conservation on the Tsushima islands, Japan. J. Hered. 2005, 96, 688–697. [Google Scholar] [CrossRef] [Green Version]
- Janecka, J.E.; Jackson, R.; Munkhtsog, B.; Murphy, W.J. Characterization of 9 microsatellites and primers in snow leopards and a species-specific PCR assay for identifying noninvasive samples. Conserv. Genet. Resour. 2014, 6, 369–373. [Google Scholar] [CrossRef]
- Miotto, R.A.; Cervini, M.; Begotti, R.A.; Galetti, P.M., Jr. Monitoring a puma (Puma concolor) population in a fragmented landscape in southeast Brazil. Biotropica 2012, 44, 98–104. [Google Scholar] [CrossRef]
- Sicuro, F.L.; Oliveira, L.F.B. Variations in leopard cat (Prionailurus bengalensis) skull morphology and body size: Sexual and geograhic influences. PeerJ 2015, 3, e1309. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, A.T.; Xie, Y. A Guide to the Mammals of China; Princeton University Press: Princeton, NJ, USA, 2008; pp. 5–13. [Google Scholar]
- Ross, J.; Brodie, J.; Cheyne, S. Prionailurus bengalensis. The IUCN Red List of Threatened Species. 2015. Available online: www.iucnredlist.org (accessed on 15 January 2021).
- Wang, Y.X.; Jiang, X.L.; Feng, Q.; Chen, Z.P. Abundance, sustainable utilization and conservation of leopard cat in Yunnan. Acta Theriol. Sin. 1997, 1, 32–43. [Google Scholar]
- Seto, K.C.; Güneralp, B.; Hutyra, L.R. Global forecasts of urban expansion to 2030 and direct impacts on biodiversity and carbon pools. Proc. Nat. Acad. Sci. USA 2012, 109, 16083–16088. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lorica, R. Prionailurus bengalensis ssp. rabori. The IUCN Red List of Threatened Species. 2008. Available online: www.iucnredlist.org (accessed on 15 January 2021).
- Izawa, M.; Doi, T. Prionailurus bengalensis ssp. Iriomotensis (Errata Version Published in 2016). The IUCN Red List of Threatened Species 2015. Available online: www.iucnredlist.org (accessed on 15 January 2021).
- Jiang, Z.G.; Jiang, J.P.; Wang, Y.Z.; Zhang, E.; Zhang, Y.Y.; Li, L.L.; Xie, F.; Cai, B.; Cao, L.; Zheng, G.M.; et al. Red list of China’s vertebrates. Biodiv. Sci. 2016, 24, 500–551. [Google Scholar]
- Izawa, M.; Nakanishi, N. Prionailurus bengalensis euptilurus (Elliot, 1871). In The Wild Mammals of Japan; Ohdachi, S.D., Ishibashi, Y., Iwasa, M.A., Saitoh, T., Eds.; Soukadoh Book Sellers and the Mammalogical Society of Japan: Kyoto, Japan, 2009; pp. 226–227. [Google Scholar]
- Nakanishi, N.; Izawa, M. Prionailurus bengalensis iriomotensis (Imaizumi, 1967). In The Wild Mammals of Japan; Ohdachi, S.D., Ishibashi, Y., Iwasa, M.A., Saitoh, T., Eds.; Soukadoh Book Sellers and the Mammalogical Society of Japan: Kyoto, Japan, 2009; pp. 228–229. [Google Scholar]
- Saka, T.; Nishita, Y.; Masuda, R. Low genetic variation in the MHC class II DRB gene and MHC-linked microsatellites in endangered island populations of the leopard cat (Prionailurus bengalensis) in Japan. Immunogenetics 2018, 70, 115–124. [Google Scholar] [CrossRef]
- Tamada, T.; Siriaroonrat, B.; Subramaniam, V. Molecular diversity and phylogeography of the Asian leopard cat, Felis bengalensis, inferred from mitochondrial and Y-chromosomal DNA sequences. Zool. Sci. 2008, 25, 154–163. [Google Scholar] [CrossRef] [PubMed]
- Eo, S.H.; Ko, B.J.; Lee, B.J.; Seomun, H.; Kim, S.; Kim, M.J.; Kim, J.H.; An, J. A set of microsatellite markers for population genetics of leopard cat (Prionailurus bengalensis) and cross-species amplification in other felids. Biochem. Syst. Ecol. 2016, 66, 196–200. [Google Scholar] [CrossRef]
- Ko, B.J.; An, J.; Seomun, H.; Lee, M.-Y.; Eo, S.H. Microsatellite DNA analysis reveals lower than expected genetic diversity in the threatened leopard cat (Prionailurus bengalensis) in South Korea. Genes Genom. 2018, 40, 521–530. [Google Scholar] [CrossRef]
- Bai, S.Y. Genetic Diversity and Phylogenetic Relationships among Populations of the Leopard Cat Felis bengalensis; Northeast Forestry University: Harbin, China, 2001; pp. 45–49. (In Chinese) [Google Scholar]
- Wei, D.Q. Study on the Genetic Structure and Geographical Differentiation of Leopard Cat Population; Northeast Forestry University: Harbin, China, 2006; pp. 33–41. (In Chinese) [Google Scholar]
- Fan, Y.Q.; Yang, J.; Zhang, H.L.; Jiang, J.; Jiang, W.J.; Tang, S.P.; Bao, W.D. Food composition of medium sized carnivores in Beijing Songshan nature reserve. J. Biol. 2020, 37, 59–62. [Google Scholar]
- Bao, W.D.; Li, X.J.; Shi, Y. Comparative analysis of food habits in carnivores from three areas of Beijing. Zool. Res. 2005, 26, 118–122. [Google Scholar]
- Tang, X.M.; Zhang, D.H.; Ma, Z.H.; Wu, T.L.; Zhang, Y.S.; Bao, W.D. Camera trapping survey on ground-dwelling birds and mammals of spring and winter in Beijing Wulingshan nature reserve. Chinese J. Zool. 2019, 51, 751–760. [Google Scholar]
- Zhang, Y.S.; Jaing, W.J.; Jiang, J.; Dan, W.; Wu, J.; Liu, F.; Bao, W.D. Biodiversity monitoring of understory birds and mammals in Beijing Songshan national nature reserve. Chin. J. Wildl. 2017, 38, 367–375. [Google Scholar]
- Lan, H.; Jin, K. Infrared-triggered camera technology application in the investigation of mammals in Beijing Wulingshan national nature reserve. Acta Theriol. Sin. 2016, 36, 322–329. [Google Scholar]
- Liu, F.; Li, D.Q.; Wu, J.G. Using infra-red cameras to survey wildlife in Beijing Songshan national nature reserve. Acta Ecol. Sin. 2012, 32, 730–739. [Google Scholar]
- Yang, J.; Zhu, G.F.; Jiang, J.; Xiang, C.L.; Gao, F.L.; Bao, W.D. Non-invasive genetic analysis indicates low population connectivity in the vulnerable Chinese gorals: Concerns for segregated population management. Zool. Res. 2019, 40, 39–448. [Google Scholar] [CrossRef] [PubMed]
- Chaves, P.B.; Graeff, V.G.; Lion, M.B.; Oliveira, L.R.; Eizirik, E. DNA barcoding meets molecular scatology: Short mtDNA sequences for standardized species assignment of carnivore noninvasive samples. Mol. Ecol. Resour. 2012, 12, 18–35. [Google Scholar] [CrossRef]
- Pilgrim, K.L.; McKelvey, K.S.; Riddle, A.E.; Schwartz, M.K. Felid sex identification based on noninvasive genetic samples. Mol. Ecol. Notes 2005, 5, 60–61. [Google Scholar] [CrossRef]
- Mondol, S.; Karanth, K.U.; Kumar, N.S.; Andheria, A.; Ramakrishnan, U. Evaluation of non-invasive genetic sampling methods for estimating tiger population size. Biol. Conserv. 2009, 142, 2350–2360. [Google Scholar] [CrossRef]
- Bellemain, E.V.A.; Swenson, J.E.; Tallmon, D.; Brunberg, S.; Taberlet, P. Estimating population size of elusive animals with DNA from hunter-collected feces: Four methods for brown bears. Conserv. Biol. 2005, 19, 150–161. [Google Scholar] [CrossRef]
- Excoffier, L.; Lischer, H.E.L. Arlequin suite ver 3.5: A new series of programs to perform population genetics analyses under Linux and Windows. Mol. Ecol. Resour. 2010, 10, 564–567. [Google Scholar] [CrossRef] [PubMed]
- Rousset, F. GENEPOP’007: A complete re-implementation of the GENEPOP software for Windows and Linux. Mol. Ecol. Resour. 2008, 8, 103–106. [Google Scholar] [CrossRef]
- Peakall, R.; Smouse, P.E. GenAlEx 6.5: Genetic analysis in Excel. Population genetic software for teaching and research—An update. Bioinformatics 2012, 28, 2537–2539. [Google Scholar] [CrossRef] [Green Version]
- Evanno, G.; Regnaut, S.; Goudet, J. Detecting the number of clusters of individuals using the software STRUCTURE: A simulation study. Mol. Ecol. 2005, 14, 2611–2620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jombart, T.; Devillard, S.; Balloux, F. Discriminant analysis of principal components: A new method for the analysis of genetically structured populations. BMC Genet. 2010, 11, 2156–2171. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miller, C.R.; Joyce, P.; Waits, L.P. A new method for estimating the size of small populations from genetic mark–recapture data. Mol. Ecol. 2005, 14, 1991–2005. [Google Scholar] [CrossRef] [PubMed]
- Du, L.H.; Wang, X.P.; Chen, J.Q. A Comprehensive Scientific Investigation Report of Beijing Songshan National Nature Reserve; China Forestry Publishing House: Beijing, China, 2012; pp. 56–59. [Google Scholar]
- Zinn, H.C.; Pierce, C.L. Values, gender, and concern about potentially dangerous wildlife. Envi. Behav. 2002, 34, 239–256. [Google Scholar] [CrossRef]
- Sharma, S.; Dutta, T.; Maldonado, J.E.; Wood, T.C.; Panwar, H.S.; Seidensticker, J. A highly informative microsatellite panel for individual identification and sex determination of jungle cats (Felis chaus). Conserv. Genet. Resour. 2013, 5, 863–866. [Google Scholar] [CrossRef]
- Sharma, K.; Bayrakcismith, R.; Tumursukh, L. Vigorous dynamics underlie a stable population of the endangered snow leopard (Panthera uncia) in Tost Mountains, South Gobi, Mongolia. PLoS ONE 2014, 9, e101319. [Google Scholar] [CrossRef] [Green Version]
- Björklund, M. The risk of inbreeding due to habitat loss in the lion (Panthera leo). Conserv. Genet. 2003, 4, 515–523. [Google Scholar] [CrossRef]
- Barthold, J.A.; Loveridge, A.J.; Macdonald, D.W.; Packer, C.; Colchero, F. Bayesian estimates of male and female African lion mortality for future use in population management. J. Appl. Ecol. 2016, 53, 295–304. [Google Scholar] [CrossRef] [Green Version]
- Aziz, M.A.; Tollington, S.; Barlow, A.; Greenwood, C.; Goodrich, J.M.; Smith, O.; Shamsuddoha, M.; Islam, M.A.; Groombridge, J.J. Using non-invasively collected genetic data to estimate density and population size of tigers in the Bangladesh Sundarbans. Glob. Ecol. Conserv. 2017, 12, 272–282. [Google Scholar] [CrossRef]
- Reed, D.H.; Frankham, R. Correlation between fitness and genetic diversity. Conserv. Biol. 2003, 17, 230–237. [Google Scholar] [CrossRef]
- Chen, W.; Gao, W.; Fu, B.Q. Mammals of Beijing; Beijing Publishing House: Beijing, China, 2002; pp. 5–14. [Google Scholar]
- Waits, L.P.; Luikart, G.; Taberlet, P. Estimating the probability of identity among genotypes in natural populations: Cautions and guidelines. Mol. Ecol. 2001, 10, 249–256. [Google Scholar] [CrossRef] [PubMed]
- Botstein, D.; White, R.L.; Skolnick, M.H.; Davis, R. Construction of a genetic linkage map in man using restriction fragment length polymorphisms. Am. J. Hum. Genet. 1980, 32, 314–331. [Google Scholar]
- Waits, L.; Taberlet, P.; Swenson, J.E.; Sandegren, F.; Franzen, R. Nuclear DNA microsatellite analysis of genetic diversity and gene flow in the Scandinavian brown bear (Ursus arctos). Mol. Ecol. 2000, 9, 421–431. [Google Scholar] [CrossRef]
- Pavlova, A.; Amos, J.N.; Joseph, L.; Loynes, K.; Austin, J.J.; Keogh, J.S.; Stone, G.N.; Nicholls, J.A.; Sunnucks, P. Perched at the mito-nuclear crossroads: Divergent mitochondrial lineages correlate with environment in the face of ongoing nuclear gene flow in an Australian bird. Evolution 2013, 67, 3412–3428. [Google Scholar] [CrossRef]




{kind=link}
{kind=link}
{kind=link}
| Locus | Primer Sequence (5′-3′) | Ta (°C) | Size (bp) |
|---|---|---|---|
| Zn-finger | F: AAGTTTACACAACCACCTGG | 56 | X162/Y165 |
| R: CACAGAATTTACACTTGTGCA | |||
| Pbe03 | F: M13FCTGCCTTTGACTGCTCCAC | 58 | 131–163 |
| R: TGCTTACCATGTGACCTCC | |||
| Pbe05 | F: M13FTCACCTCTGGGCTCTTG | 60 | 181–193 |
| R: AGGGACACGGAAAGGCATC | |||
| Pbe13 | F: M13FTGCGGATGTTGGGAAAGAAC | 60 | 210–218 |
| R: AGGCCGAGACCAGTTAAGG | |||
| Pbe28 | F: M13FGGGAGACCTTGCCTCATTTC | 56 | 233–241 |
| R: TGCTTCCCTAACAGGCATC | |||
| Pbe32 | F: M13FAGCACTAGGCCAGAACACC | 64 | 174–178 |
| R: CCAGACCCTCTTTGCCTTG | |||
| Pbe33 | F:M13FAGAGGCACTTGGAGTTAGGG | 58 | 248–252 |
| R: GAG TCGGCA AACCTGGAAC |
| Microsatellites Loci | Allele | Allele Frequency | ||||
|---|---|---|---|---|---|---|
| SS | YMS | YFS | XLM | BHS | ||
| Pbe03 | 131 | 0.142 | 0.059 | 0.088 | 0.115 | 0.042 |
| 135 | 0.094 | 0.147 | 0.000 | 0.038 | 0.167 | |
| 139 | 0.057 | 0.000 | 0.059 | 0.000 | 0.292 | |
| 143 | 0.330 | 0.559 | 0.353 | 0.500 | 0.417 | |
| 147 | 0.019 | 0.000 | 0.059 | 0.000 | 0.000 | |
| 151 | 0.113 | 0.000 | 0.059 | 0.000 | 0.000 | |
| 155 | 0.038 | 0.000 | 0.059 | 0.000 | 0.000 | |
| 159 | 0.179 | 0.235 | 0.324 | 0.346 | 0.083 | |
| 163 | 0.028 | 0.000 | 0.000 | 0.000 | 0.000 | |
| 131 | 0.142 | 0.000 | 0.000 | 0.000 | 0.000 | |
| Pbe05 | 177 | 0.000 | 0.000 | 0.000 | 0.154 | 0.000 |
| 181 | 0.274 | 0.500 | 0.118 | 0.192 | 0.250 | |
| 185 | 0.698 | 0.500 | 0.882 | 0.654 | 0.708 | |
| 189 | 0.000 | 0.000 | 0.000 | 0.000 | 0.042 | |
| 193 | 0.028 | 0.000 | 0.000 | 0.000 | 0.000 | |
| Pbe13 | 202 | 0.000 | 0.000 | 0.000 | 0.115 | 0.083 |
| 206 | 0.038 | 0.000 | 0.029 | 0.000 | 0.000 | |
| 210 | 0.368 | 0.000 | 0.176 | 0.154 | 0.083 | |
| 214 | 0.377 | 0.588 | 0.471 | 0.500 | 0.625 | |
| 218 | 0.208 | 0.412 | 0.324 | 0.077 | 0.125 | |
| 222 | 0.009 | 0.000 | 0.000 | 0.154 | 0.083 | |
| Pbe28 | 233 | 0.028 | 0.235 | 0.088 | 0.231 | 0.542 |
| 237 | 0.623 | 0.441 | 0.500 | 0.538 | 0.208 | |
| 241 | 0.349 | 0.324 | 0.412 | 0.231 | 0.250 | |
| Pbe32 | 170 | 0.019 | 0.029 | 0.000 | 0.000 | 0.000 |
| 174 | 0.292 | 0.176 | 0.324 | 0.269 | 0.250 | |
| 178 | 0.679 | 0.794 | 0.676 | 0.731 | 0.750 | |
| 182 | 0.009 | 0.000 | 0.000 | 0.000 | 0.000 | |
| Pbe33 | 240 | 0.000 | 0.000 | 0.059 | 0.000 | 0.000 |
| 248 | 0.217 | 0.294 | 0.294 | 0.269 | 0.125 | |
| 252 | 0.783 | 0.706 | 0.647 | 0.731 | 0.875 | |
| Microsatellite Loci | Microsatellite Index | SS (N = 53) | YMS (N = 17) | YFS (N = 17) | XLM (N = 13) | BHS (N = 12) |
|---|---|---|---|---|---|---|
| Pbe03 | Na | 9.000 | 4.000 | 7.000 | 4.000 | 5.000 |
| Ne | 5.300 | 2.546 | 3.986 | 2.600 | 3.388 | |
| Ho | 0.698 | 0.588 | 0.588 | 0.462 | 0.583 | |
| He | 0.811 | 0.607 | 0.749 | 0.615 | 0.705 | |
| PIC | 0.819 | 0.554 | 0.713 | 0.544 | 0.655 | |
| PID | 0.0576 | 0.2074 | 0.0993 | 0.2188 | 0.1360 | |
| PID-sibs | 0.3587 | 0.4982 | 0.4003 | 0.4970 | 0.4316 | |
| Pbe05 | Na | 3.000 | 2.000 | 2.000 | 3.000 | 3.000 |
| Ne | 1.776 | 2.000 | 1.262 | 2.048 | 1.767 | |
| Ho | 0.415 | 0.294 | 0.235 | 0.385 | 0.583 | |
| He | 0.437 | 0.500 | 0.208 | 0.512 | 0.434 | |
| PIC | 0.441 | 0.375 | 0.186 | 0.458 | 0.369 | |
| PID | 0.03908 | 0.3750 | 0.6794 | 0.2919 | 0.3850 | |
| PID-sibs | 0.6292 | 0.5938 | 0.8086 | 0.5671 | 0.6292 | |
| Pbe13 | Na | 5.000 | 2.000 | 4.000 | 5.000 | 5.000 |
| Ne | 3.102 | 1.940 | 2.792 | 3.159 | 2.341 | |
| Ho | 0.642 | 0.353 | 0.529 | 0.462 | 0.250 | |
| He | 0.678 | 0.484 | 0.642 | 0.683 | 0.573 | |
| PIC | 0.684 | 0.367 | 0.575 | 0.647 | 0.544 | |
| PID | 0.1674 | 0.3831 | 0.1955 | 0.1366 | 0.2118 | |
| PID-sibs | 0.4530 | 0.6036 | 0.4780 | 0.4424 | 0.5165 | |
| Pbe28 | Na | 3.000 | 3.000 | 3.000 | 3.000 | 3.000 |
| Ne | 1.960 | 2.820 | 2.340 | 2.522 | 2.504 | |
| Ho | 0.566 | 0.471 | 0.412 | 0.154 | 0.667 | |
| He | 0.490 | 0.645 | 0.573 | 0.604 | 0.601 | |
| PIC | 0.494 | 0.571 | 0.481 | 0.536 | 0.533 | |
| PID | 0.3557 | 0.1997 | 0.2739 | 0.2246 | 0.2270 | |
| PID-sibs | 0.5941 | 0.4773 | 0.5321 | 0.5044 | 0.5064 | |
| Pbe32 | Na | 4.000 | 3.000 | 2.000 | 2.000 | 2.000 |
| Ne | 1.827 | 1.509 | 1.778 | 1.649 | 1.600 | |
| Ho | 0.547 | 0.176 | 0.294 | 0.231 | 0.167 | |
| He | 0.453 | 0.337 | 0.438 | 0.393 | 0.375 | |
| PIC | 0.457 | 0.297 | 0.342 | 0.316 | 0.305 | |
| PID | 0.3790 | 0.4795 | 0.4120 | 0.4453 | 0.4609 | |
| PID-sibs | 0.6184 | 0.7012 | 0.6341 | 0.6646 | 0.6777 | |
| Pbe33 | Na | 2.000 | 2.000 | 3.000 | 2.000 | 2.000 |
| Ne | 1.515 | 1.710 | 1.966 | 1.649 | 1.280 | |
| Ho | 0.321 | 0.235 | 0.294 | 0.077 | 0.083 | |
| He | 0.340 | 0.415 | 0.491 | 0.393 | 0.219 | |
| PIC | 0.343 | 0.329 | 0.415 | 0.316 | 0.195 | |
| PID | 0.4936 | 0.4282 | 0.3347 | 0.4453 | 0.6343 | |
| PID-sibs | 0.7035 | 0.6494 | 0.5880 | 0.6646 | 0.7992 | |
| Average | Na | 4.333 | 2.667 | 3.500 | 3.167 | 3.333 |
| Ne | 2.580 | 2.087 | 2.354 | 2.271 | 2.147 | |
| Ho | 0.531 | 0.353 | 0.392 | 0.295 | 0.389 | |
| He | 0.535 | 0.498 | 0.517 | 0.534 | 0.484 | |
| PIC | 0.470 | 0.418 | 0.452 | 0.470 | 0.434 | |
| combined PID | 2.5 × 10−4 | 1.2 × 10−3 | 4.7 × 10−4 | 3.9 × 10−4 | 7.3 × 10−4 | |
| combined PID-sibs | 2.6 × 10−2 | 3.8 × 10−2 | 3.0 × 10−2 | 2.7 × 10−2 | 3.8 × 10−2 |
| Sampling Area | SS | YMS | YFS | XLM | BHS |
|---|---|---|---|---|---|
| SS | 4.136 | 18.981 | 11.655 | 2.799 | |
| YMS | 0.057 | 4.852 | 9.366 | 4.467 | |
| YFS | 0.013 | 0.049 | 22.478 | 3.222 | |
| XLM | 0.021 | 0.026 | 0.011 | 6.695 | |
| BHS | 0.082 | 0.053 | 0.072 | 0.036 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Teng, Y.; Yang, J.; Ju, L.-F.; Huang, W.-H.; Zhang, X.; Gao, F.-L.; Bao, W.-D. Current Genetic Structure Analysis of Leopard Cats Reveals a Weak Disparity Trend in Subpopulations in Beijing, China. Biology 2022, 11, 1478. https://doi.org/10.3390/biology11101478
Teng Y, Yang J, Ju L-F, Huang W-H, Zhang X, Gao F-L, Bao W-D. Current Genetic Structure Analysis of Leopard Cats Reveals a Weak Disparity Trend in Subpopulations in Beijing, China. Biology. 2022; 11(10):1478. https://doi.org/10.3390/biology11101478
Chicago/Turabian StyleTeng, Yang, Jing Yang, Long-Fei Ju, Wen-Hua Huang, Xin Zhang, Fu-Li Gao, and Wei-Dong Bao. 2022. "Current Genetic Structure Analysis of Leopard Cats Reveals a Weak Disparity Trend in Subpopulations in Beijing, China" Biology 11, no. 10: 1478. https://doi.org/10.3390/biology11101478