
Morphometric and Genetic Description of Trophic Adaptations in Cichlid Fishes

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Fishes and Pedigree
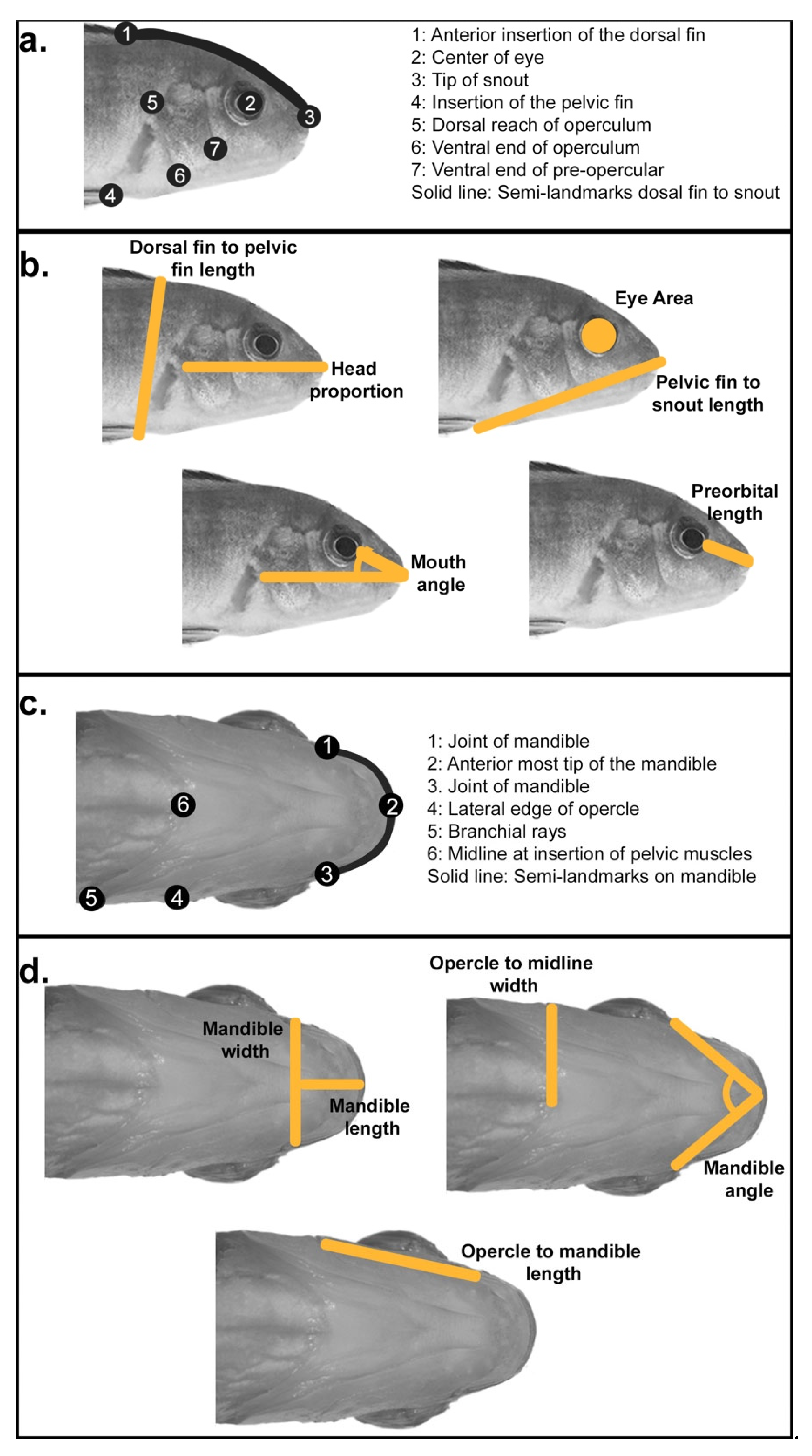
2.2. Linear Measures of Head Shape Variation
2.3. Geometric Morphometric Shape Analysis
2.4. Genotyping with ddRAD Sequencing
2.5. Generation of the Linkage Map
2.6. Quantitative Trait Loci (QTL) Mapping
2.7. Candidate Gene Annotation and Enrichment Analysis
3. Results and Discussion
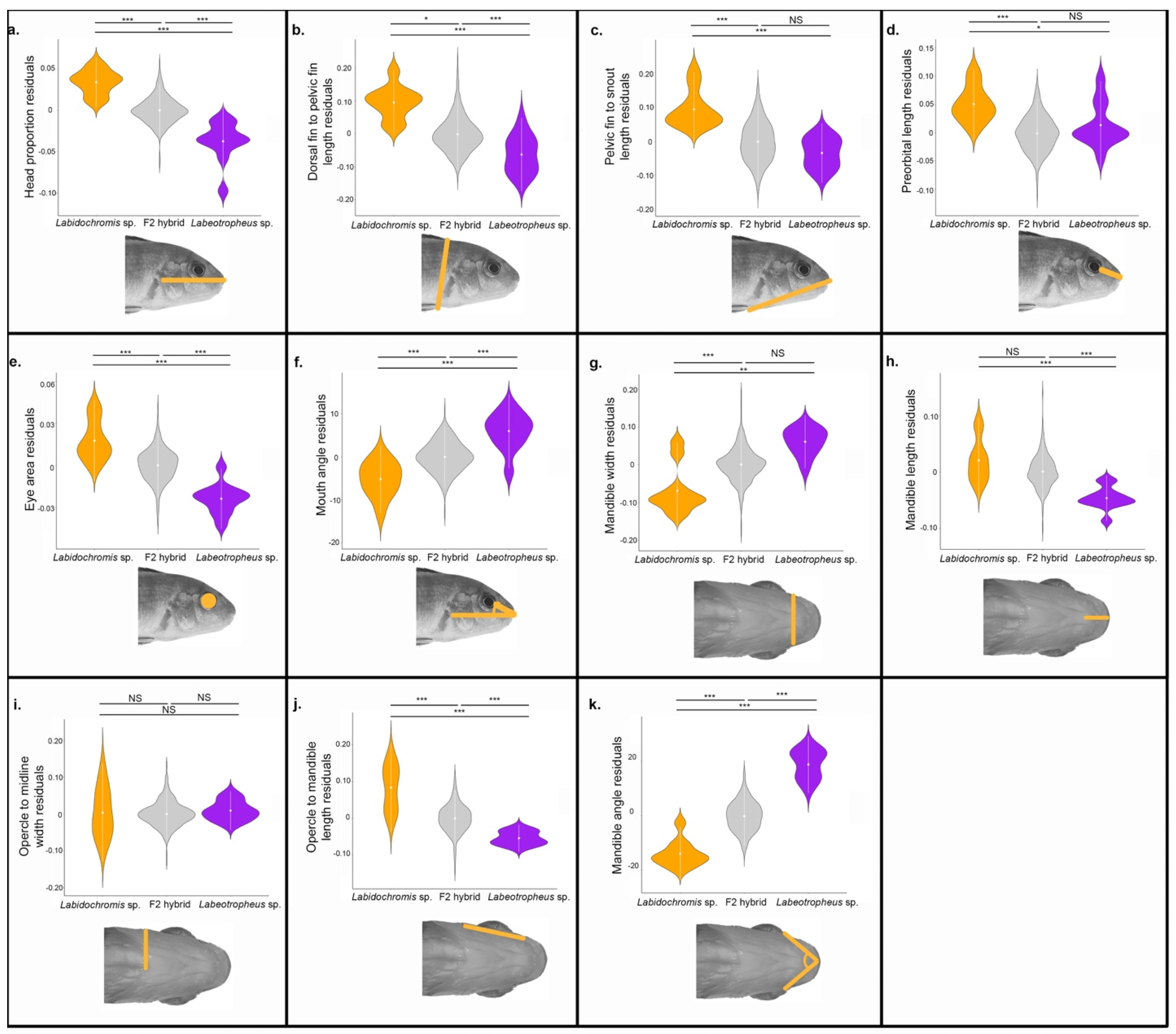
3.1. Lateral Head Shape Variation
3.2. Ventral Head Shape Variation
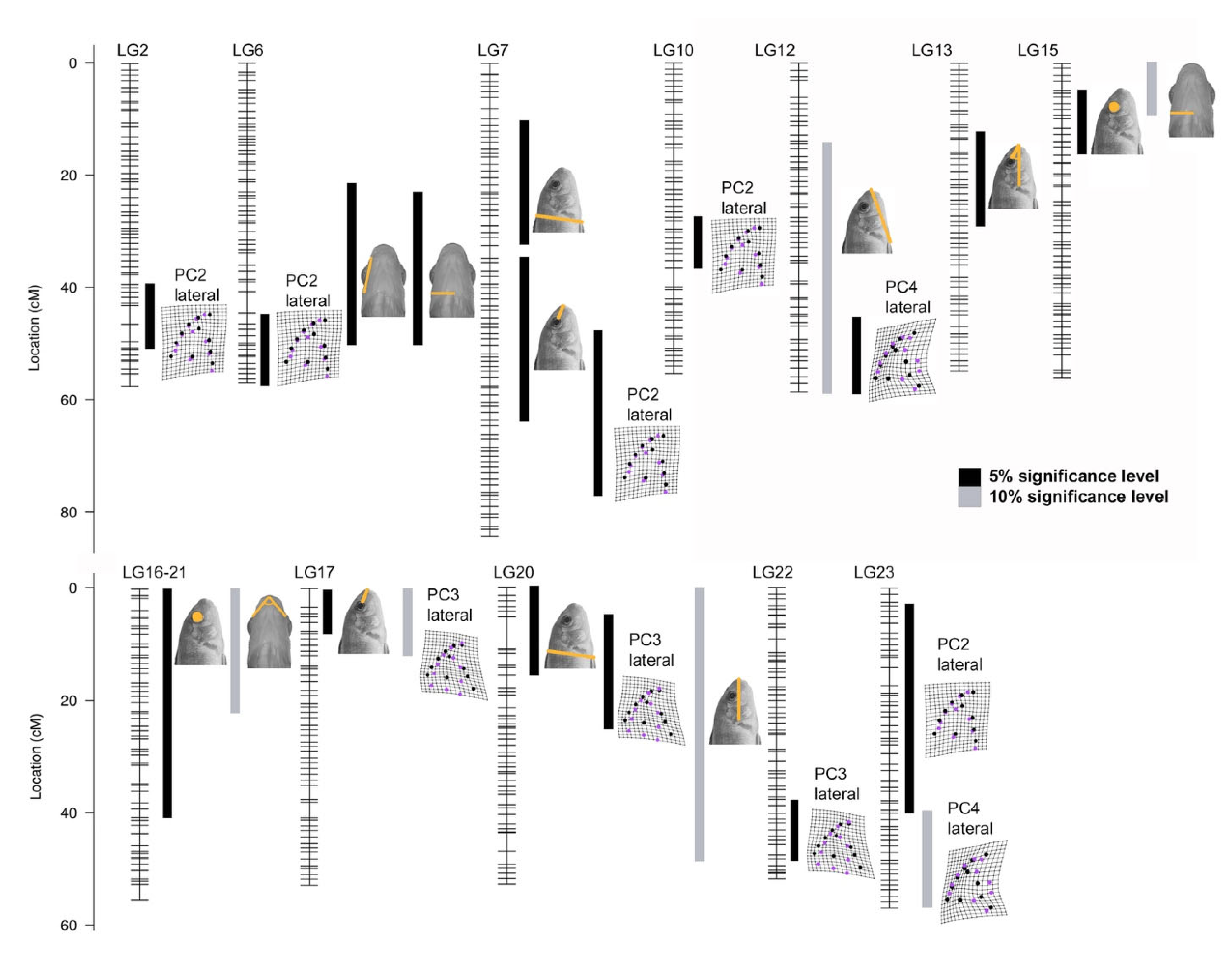
3.3. Genetic Basis of Head Shape
3.4. Patterns of Covariation and Modularity
3.5. Gene Ontology (GO) Analysis
3.6. Candidate Genes in QTL Intervals
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Evans, K.M.; Waltz, B.; Tagliacollo, V.; Chakrabarty, P.; Albert, J.S. Why the short face? Developmental disintegration of the neurocranium drives convergent evolution in neotropical electric fishes. Ecol. Evol. 2017, 7, 1783–1801. [Google Scholar] [CrossRef]
- Powder, K.E.; Albertson, R.C. Cichlid fishes as a model to understand normal and clinical craniofacial variation. Dev. Biol. 2016, 415, 338–346. [Google Scholar] [CrossRef]
- McGirr, J.; Martin, C. Few fixed variants between trophic specialist pupfish species reveal candidate cis-regulatory alleles underlying rapid craniofacial divergence. Mol. Biol. Evol. 2021, 38, 405–423. [Google Scholar] [CrossRef]
- Brugmann, S.; Powder, K.E.; Young, N.M.; Goodnough, L.H.; Hahn, S.M.; James, A.W.; Helms, J.A.; Lovett, M. Comparative gene expression analysis of avian embryonic facial structures reveals new candidates for human craniofacial disorders. Hum. Mol. Genet. 2010, 19, 920–930. [Google Scholar] [CrossRef] [Green Version]
- Grant, P.R.; Grant, B.R. Unpredictable evolution in a 30-year study of Darwin’s finches. Science 2002, 296, 707–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Navalón, G.; Marugán-Lobón, J.; Bright, J.A.; Cooney, C.R.; Rayfield, E.J. The consequences of craniofacial integration for the adaptive radiations of Darwin’s finches and Hawaiian honeycreepers. Nat. Ecol. Evol. 2020, 4, 270–278. [Google Scholar] [CrossRef] [PubMed]
- Stayton, C.T. Morphological evolution of the lizard skull: A geometric morphometrics survey. J. Morphol. 2005, 263, 47–59. [Google Scholar] [CrossRef] [PubMed]
- Sanger, T.J.; Mahler, D.L.; Abzhanov, A.; Losos, J.B. Roles for modularity and constraint in the evolution of cranial diversity among Anolis lizards. Evolution 2011, 66, 1525–1542. [Google Scholar] [CrossRef] [PubMed]
- Cardini, A.; Polly, P. Larger mammals have longer faces because of size-related constraints on skull form. Nat. Commun. 2013, 4, 2458. [Google Scholar] [CrossRef] [PubMed]
- Porto, A.; Shirai, L.T.; de Oliveira, F.B.; Marroig, G. Size variation, growth strategies, and the evolution of modularity in the mammalian skull. Evolution 2013, 67, 3305–3322. [Google Scholar] [CrossRef]
- Schoenebeck, J.J.; Ostrander, E.A. The genetics of canine skull shape variation. Genetics 2013, 193, 317–325. [Google Scholar] [CrossRef] [Green Version]
- Sears, K.E. Differences in growth generate the diverse palate shapes of New World leaf-nosed bats (order Chiroptera, family Phyllostomidae). Evol. Biol. 2014, 41, 12–21. [Google Scholar] [CrossRef]
- Arbour, J.H.; Curtis, A.A.; Santana, S.E. Signatures of echolocation and dietary ecology in the adaptive evolution of skull shape in bats. Nat. Commun. 2019, 10, 2036. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arnold, S. Morphology, performance and fitness. Amer. Zool. 1983, 23, 347–361. [Google Scholar] [CrossRef]
- Hallgrímsson, B.; Jamniczky, H.; Young, N.M.; Rolian, C.; Parsons, T.E.; Boughner, J.C.; Marcucio, R.S. Deciphering the Palimpsest: Studying the Relationship Between Morphological Integration and Phenotypic Covariation. Evol. Biol. 2009, 36, 355–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hallgrimsson, B.; Mio, W.; Marcucio, R.S.; Spritz, R. Let’s Face It-Complex Traits Are Just Not That Simple. PLoS Genet. 2014, 10, e1004724. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hochheiser, H.; Aronow, B.J.; Artinger, K.; Beaty, T.H.; Brinkley, J.F.; Chai, Y.; Clouthier, D.; Cunningham, M.L.; Dixon, M.; Donahue, L.R.; et al. The FaceBase Consortium: A comprehensive program to facilitate craniofacial research. Dev. Biol. 2011, 355, 175–182. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fish, J.; Sklar, R.S.; Woronowicz, K.C.; Schneider, R.A. Multiple developmental mechanisms regulate species-specific jaw size. Development 2014, 141, 674–684. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Glazier, A.M.; Nadeau, J.H.; Aitman, T.J. Finding genes that underlie complex traits. Science 2002, 298, 2345–2349. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hirschhorn, J.N.; Daly, M.J. Genome-wide association studies for common diseases and complex traits. Nat. Rev. Genet. 2005, 6, 95–108. [Google Scholar] [CrossRef] [PubMed]
- Turner, G.F.; Seehausen, O.; Knight, M.E.; Allender, C.J.; Robinson, R.L. How many species of cichlid fishes are there in African lakes? Mol Ecol. 2008, 10, 793–806. [Google Scholar] [CrossRef]
- Salzburger, W.; Meyer, A. The species flocks of East African cichlid fishes: Recent advances in molecular phylogenetics and population genetics. Naturwissenschaften 2004, 91, 277–290. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Turner, G.F. Adaptive radiation of cichlid fish. Curr. Biol. 2007, 17, R827–R831. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elmer, K.R.; Kusche, H.; Lehtonen, T.K.; Meyer, A. Local variation and parallel evolution: Morphological and genetic diversity across a species complex of neotropical crater lake cichlid fishes. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2010, 365, 1763–1782. [Google Scholar] [CrossRef] [Green Version]
- Kornfield, I.; Smith, P.F. African cichlid fishes: Model systems for evolutionary biology. Annu. Rev. Ecol. Syst. 2000, 31, 163–196. [Google Scholar] [CrossRef]
- Konings, A. Malawi Cichlids in Their Natural Habitat, 5th ed.; Cichlid Press: El Paso, TX, USA, 2016. [Google Scholar]
- Liem, K.J. Adaptive significance of intra- and interspecific differences in the feeding repertoires of cichlid fishes. Amer. Zool. 1980, 20, 295–314. [Google Scholar] [CrossRef]
- Cooper, W.J.; Parsons, K.; McIntyre, A.; Kern, B.; McGee-Moore, A.; Albertson, R.C. Bentho-pelagic divergence of cichlid feeding architecture was prodigious and consistent during multiple adaptive radiations within African rift-lakes. PLoS ONE 2010, 5, e9551. [Google Scholar] [CrossRef]
- Wainwright, P.; Carroll, A.M.; Collar, D.C.; Day, S.W.; Higham, T.E.; Holzman, R.A. Suction feeding mechanics, performance, and diversity in fishes. Integr. Comp. Biol. 2007, 47, 96–106. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wainwright, P.C.; Day, S.W. The forces exerted by aquatic suction feeders on their prey. J. R. Soc. Interface 2007, 4, 553–560. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Westneat, M.W. Skull biomechanics and suction feeding in fishes. In Fish Biomechanics. Fish Physiology; Shadwick, R., Lauder, G., Eds.; Academic: San Diego, CA, USA, 2005; pp. 29–75. [Google Scholar]
- Ferry-Graham, L.A.; Lauder, G.V. Aquatic prey capture in ray-finned fishes: A century of progress and new directions. J. Morphol. 2001, 248, 99–119. [Google Scholar] [CrossRef]
- Westneat, M.W. Evolution of levers and linkages in the feeding mechanisms of fishes. Integr. Comp. Biol. 2004, 44, 378–389. [Google Scholar] [CrossRef]
- Westneat, M.W. A biomechanical model for analysis of muscle force, power output and lower jaw motion in fishes. J. Theor. Biol. 2003, 223, 269–281. [Google Scholar] [CrossRef]
- Albertson, R.C.; Streelman, J.T.; Kocher, T.D.; Yelick, P.C. Integration and evolution of the cichlid mandible: The molecular basis of alternate feeding strategies. Proc. Natl. Acad. Sci. USA 2005, 102, 16287–16292. [Google Scholar] [CrossRef] [Green Version]
- Carroll, A.M.; Wainwright, P.C.; Huskey, S.H.; Collar, D.C.; Turingan, R.G. Morphology predicts suction feeding performance in centrarchid fishes. J. Exp. Biol. 2004, 207, 3873–3881. [Google Scholar] [CrossRef] [Green Version]
- Meer, H.; van Der, J.; Anker, G.C. Retinal resolving power and sensitivity of the photopic system in seven haplochromine species (Pisces, Teleostei). Neth. J. Zool. 1984, 34, 197–209. [Google Scholar] [CrossRef]
- Hulsey, C.D.; Mims, M.C.; Streelman, J.T. Do constructional constraints influence cichlid craniofacial diversification? Proc. R. Soc. B 2007, 274, 1867–1875. [Google Scholar] [CrossRef] [Green Version]
- Albertson, R.C.; Markert, J.A.; Danley, P.D.; Kocher, T.D. Phylogeny of a rapidly evolving clade: The cichlid fishes of Lake Malawi, East Africa. Proc. Natl. Acad. Sci. USA 1999, 96, 5107–5110. [Google Scholar] [CrossRef] [Green Version]
- Young, K.A.; Snoeks, J.; Seehausen, O. Morphological diversity and the roles of contingency, chance and determinism in african cichlid radiations. PLoS ONE 2009, 4, e4740. [Google Scholar] [CrossRef] [Green Version]
- Cooper, W.J.; Westneat, M.W. Form and function of damselfish skulls: Rapid and repeated evolution into a limited number of trophic niches. BMC Evol. Biol. 2009, 9, 24. [Google Scholar] [CrossRef] [Green Version]
- Westneat, M.W.; Alfaro, M.E.; Wainwright, P.C.; Bellwood, D.R.; Grubich, J.R.; Fessler, J.L.; Clements, K.D.; Smith, L.L. Local phylogenetic divergence and global evolutionary convergence of skull function in reef fishes of the family Labridae. Proc. Biol. Sci. 2005, 272, 993–1000. [Google Scholar] [CrossRef] [Green Version]
- Collar, D.C.; Wainwright, P.C. Ecomorphology of the Centrarchidae. In Centrarchid Fishes: Diversity, Biology and Conservation; Cook, S., Phillipp, D., Eds.; Blackwell Scientific: Cambridge, MA, USA, 2009; pp. 70–89. [Google Scholar]
- Arnold, S.J. Constraints on phenotypic evolution. Am. Nat. 1992, 140 (Suppl. S1), S85–S107. [Google Scholar] [CrossRef] [PubMed]
- Futuyma, D.J. Evolutionary constraint and ecological consequences. Evolution 2010, 64, 1865–1884. [Google Scholar] [CrossRef] [PubMed]
- Smith, J.M.; Burian, R.; Kauffman, S.; Alberch, P.; Campbell, J.; Goodwin, B.; Lande, R.; Raup, D.; Wolpert, L. Developmental Constraints and Evolution: A Perspective from the Mountain Lake Conference on Development and Evolution. Q. Rev. Biol. 1985, 60, 265–287. [Google Scholar] [CrossRef]
- Wagner, G.P.; Pavlicev, M.; Cheverud, J.M. The road to modularity. Nat. Rev. Genet. 2007, 8, 921–931. [Google Scholar] [CrossRef]
- Thompson, M.J.; Jiggins, C.D. Supergenes and their role in evolution. Heredity 2014, 113, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saenko, S.V.; Chouteau, M.; Piron-Prunier, F.; Blugeon, C.; Joron, M.; Llaurens, V. Unravelling the genes forming the wing pattern supergene in the polymorphic butterfly Heliconius numata. Evodevo 2019, 10, 16. [Google Scholar] [CrossRef] [Green Version]
- Reid, K.; Bell, M.A.; Veeramah, K.R. Threespine stickleback: A model system for evolutionary genomics. Annu. Rev. Genom. Hum. Genet. 2021, 22, 357–383. [Google Scholar] [CrossRef] [PubMed]
- Fabre, A.-C.; Dowling, C.; Miguez, R.P.; Fernandez, V.; Noirault, E.; Goswami, A. Functional constraints during development limit jaw shape evolution in marsupials. Proc. Biol. Sci. 2021, 288, 20210319. [Google Scholar] [CrossRef] [PubMed]
- Parsons, K.J.; Cooper, W.J.; Albertson, R.C. Albertson, Modularity of the oral jaws is linked to repeated changes in the craniofacial shape of african cichlids. Int. J. Evol. Biol. 2011, 2011, 641501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newton, A.H.; Weisbecker, V.; Pask, A.J.; Hipsley, C.A. Ontogenetic origins of cranial convergence between the extinct marsupial thylacine and placental gray wolf. Commun. Biol. 2021, 4, 51. [Google Scholar] [CrossRef] [PubMed]
- Goswami, A. Cranial modularity shifts during mammalian evolution. Am. Nat. 2006, 168, 270–280. [Google Scholar] [CrossRef]
- Felice, R.N.; Goswami, A. Developmental origins of mosaic evolution in the avian cranium. Proc. Natl. Acad. Sci. USA 2018, 115, 555–560. [Google Scholar] [CrossRef] [Green Version]
- Felice, R.N.; Watanabe, A.; Cuff, A.R.; Noirault, E.; Pol, D.; Witmer, L.M.; Norell, M.A.; O’Connor, P.M.; Goswami, A. Evolutionary integration and modularity in the Archosaur cranium. Integr. Comp. Biol. 2019, 59, 371–382. [Google Scholar] [CrossRef] [PubMed]
- Gammerdinger, W.J.; Kocher, T.D. Unusual diversity of sex chromosomes in African cichlid fishes. Genes 2018, 9, 480. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ser, J.R.; Roberts, R.B.; Kocher, T.D. Multiple interacting loci control sex determination in lake Malawi cichlid fish. Evolution 2010, 64, 486–501. [Google Scholar] [CrossRef] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- ImageJ Software. Available online: https://imagej.nih.gov/ij/ (accessed on 15 June 2022).
- R Core Team. R: A Language and Environment for Statistical Computing. 2021. Available online: https://www.R-project.org/ (accessed on 16 June 2022).
- TpsDig2. Available online: https://sbmorphometrics.org/ (accessed on 15 June 2022).
- Adams, D.C.; Otarola-Castillo, E. Geomorph: An r package for the collection and analysis of geometric morphometric shape data. Methods Ecol. Evol. 2013, 4, 393–399. [Google Scholar] [CrossRef]
- Burford Reiskind, M.O.; Coyle, K.; Daniels, H.V.; Labadie, P.; Reiskind, M.H.; Roberts, N.B.; Roberts, R.B.; Schaff, J.; Vargo, E.L.; Schaff, J. Development of a universal double-digest RAD sequencing approach for a group of non-model, ecologically and economically important insect and fish taxa. Mol. Ecol. Resour. 2016, 16, 1303–1314. [Google Scholar] [CrossRef]
- Broman, K.W. A Guide to QTL Mapping with R/qtl; Springer: New York, NY, USA, 2009. [Google Scholar]
- Arends, D.; Prins, P.; Jansen, R.C.; Broman, K.W. R/qtl: High-throughput multiple QTL mapping. Bioinformatics 2010, 26, 2990–2992. [Google Scholar] [CrossRef] [Green Version]
- Broman, K.W.; Wu, H.; Sen, Ś.; Churchill, G.A. R/qtl: QTL mapping in experimental crosses. Bioinformatics 2003, 19, 889–890. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jansen, R.C. Controlling the type I and type II errors in mapping quantitative trait loci. Genetics 1994, 138, 871–881. [Google Scholar] [CrossRef]
- Powder, K.E. Quantitative Trait Loci (QTL) Mapping. In eQTL Analysis; Methods in Molecular Biology; Shi, X.M., Ed.; Humana: New York, NY, USA, 2020; Volume 2082. [Google Scholar]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Systematic and integrative analysis of large gene lists using DAVID bioinformatics resources. Nat. Protoc. 2009, 4, 44–57. [Google Scholar] [CrossRef]
- Huang, D.W.; Sherman, B.T.; Lempicki, R.A. Bioinformatics enrichment tools: Paths toward the comprehensive functional analysis of large gene lists. Nucleic. Acids. Res. 2009, 37, 1–13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cooper, W.J.; Wernle, J.; Mann, K.; Albertson, R.C. Functional and genetic integration in the skulls of Lake Malawi cichlids. Evol. Biol. 2011, 38, 316–334. [Google Scholar] [CrossRef]
- Holzman, R.; Day, S.W.; Mehta, R.S.; Wainwright, P.C. Jaw protrusion enhances forces exerted on prey by suction feeding fishes. J. R. Soc. Interface 2008, 5, 1445–1457. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Wassenbergh, S.; Aerts, P. Aquatic suction feeding dynamics: Insights from computational modelling. J. R. Soc. Interface 2009, 6, 149–158. [Google Scholar] [CrossRef] [Green Version]
- Moran, P.; Kornfield, I. Retention of ancestral polymorphism in the Mbuna species flock of Lake Malawi. Mol. Biol. Evol. 1993, 10, 1015–1029. [Google Scholar]
- Smith, P.F.; Konings, A.; Kornfield, I. Hybrid origin of a cichlid population in Lake Malawi: Implications for genetic variation and species diversity. Mol. Ecol. 2003, 12, 2497–2504. [Google Scholar] [CrossRef] [PubMed]
- Nagl, S.; Tichy, H.; Mayer, W.E.; Takahata, N.; Klein, J. Persistence of neutral polymorphisms in Lake Victoria cichlid fish. Proc. Natl. Acad. Sci. USA 1998, 95, 14238–14243. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Loh, Y.H.; Katz, L.S.; Mims, M.C.; Kocher, T.D.; Yi, S.V.; Streelman, J.T. Comparative analysis reveals signatures of differentiation amid genomic polymorphism in Lake Malawi cichlids. Genome Biol. 2008, 9, R113. [Google Scholar] [CrossRef]
- Malinsky, M.; Svardal, H.; Tyers, A.M.; Miska, E.A.; Genner, M.J.; Turner, G.F.; Durbin, R. Whole-genome sequences of Malawi cichlids reveal multiple radiations interconnected by gene flow. Nat. Ecol. Evol. 2018, 2, 1940–1955. [Google Scholar] [CrossRef] [Green Version]
- Pigliucci, M.; Muller, G.B. Evolution, the Extended Synthesis; MIT Press: Cambridge, MA, USA, 2010. [Google Scholar]
- Melo, D.; Porto, A.; Cheverud, J.M.; Marroig, G. Modularity: Genes, development and evolution. Annu. Rev. Ecol. Evol. Syst. 2016, 47, 463–486. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hendrikse, J.L.; Parsons, T.E.; Hallgrímsson, B. Evolvability as the proper focus of evolutionary developmental biology. Evol. Dev. 2007, 9, 393–401. [Google Scholar] [CrossRef] [PubMed]
- Fish, J.L. Evolvability of the vertebrate craniofacial skeleton. Semin. Cell Dev. Biol. 2019, 91, 13–22. [Google Scholar] [CrossRef] [PubMed]
- Breuker, C.J.; Debat, V.; Klingenberg, C.P. Functional evo-devo. Trends Ecol. Evol. 2006, 21, 488–492. [Google Scholar] [CrossRef]
- Klingenberg, C.P. Evolution and development of shape: Integrating quantitative approaches. Nat. Rev. Genet. 2010, 11, 623–635. [Google Scholar] [CrossRef]
- Holzman, R.; Collar, D.C.; Price, S.A.; Hulsey, C.D.; Thomson, R.C.; Wainwright, P.C. Biomechanical trade-offs bias rates of evolution in the feeding apparatus of fishes. Proc. Biol. Sci. 2012, 279, 1287–1292. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Satoh, W.; Matsuyama, M.; Takemura, H.; Aizawa, S.; Shimono, A. Sfrp1, Sfrp2, and Sfrp5 regulate the Wnt/beta-catenin and the planar cell polarity pathways during early trunk formation in mouse. Genesis 2008, 46, 92–103. [Google Scholar] [CrossRef]
- Kawano, Y.; Kypta, R. Secreted antagonists of the Wnt signaling pathway. J. Cell Sci. 2003, 116, 2627–2634. [Google Scholar] [CrossRef] [Green Version]
- Thyssen, G.; Li, T.H.; Lehmann, L.; Zhuo, M.; Sharma, M.; Sun, Z. LZTS2 is a novel beta-catenin-interacting protein and regulates the nuclear export of beta-catenin. Mol. Cell Biol. 2006, 26, 8857–8867. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Miao, D.; Wang, Y.; Jia, Y.; Tong, J.; Jiang, S.; Liu, L. ZRANB1 enhances stem-cell-like features and accelerates tumor progression by regulating Sox9-mediated USP22/Wnt/beta-catenin pathway in colorectal cancer. Cell Signal 2022, 90, 110200. [Google Scholar] [CrossRef]
- Wei, M.; Li, L.; Zhang, Y.; Zhang, M.; Su, Z. Downregulated circular RNA zRANB1 mediates Wnt5a/beta-Catenin signaling to promote neuropathic pain via miR-24-3p/LPAR3 axis in CCI rat models. Gene 2020, 761, 145038. [Google Scholar] [CrossRef] [PubMed]
- Parsons, K.J.; Taylor, A.T.; Powder, K.E.; Albertson, R.C. Wnt signalling underlies the evolution of new phenotypes and craniofacial variability in Lake Malawi cichlids. Nat. Commun. 2014, 5, 3629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Powder, K.E.; Milch, K.; Asselin, G.; Albertson, R.C. Constraint and diversification of developmental trajectories in cichlid facial morphologies. Evodevo 2015, 6, 25. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, B.; Rooker, S.M.; Helms, J.A. Molecular control of facial morphology. Semin. Cell Dev. Biol. 2010, 21, 309–313. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brugmann, S.A.; Goodnough, L.H.; Gregorieff, A.; Leucht, P.; ten Berge, D.; Fuerer, C.; Clevers, H.; Nusse, R.; Helms, J.A. Wnt signaling mediates regional specification in the vertebrate face. Development 2007, 134, 3283–3295. [Google Scholar] [CrossRef] [Green Version]
- Alexander, C.; Piloto, S.; Le Pabic, P.; Schilling, T.F. Wnt signaling interacts with bmp and edn1 to regulate dorsal-ventral patterning and growth of the craniofacial skeleton. PLoS Genet. 2014, 10, e1004479. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Long, F. Building strong bones: Molecular regulation of the osteoblast lineage. Nat. Rev. Mol. Cell Biol. 2011, 13, 27–38. [Google Scholar] [CrossRef]
- Zhong, Z.; Ethen, N.J.; Williams, B.O. WNT signaling in bone development and homeostasis. Wiley Interdiscip. Rev. Dev. Biol. 2014, 3, 489–500. [Google Scholar] [CrossRef] [PubMed]
- Urrego, D.; Tomczak, A.P.; Zahed, F.; Stühmer, W.; Pardo, L.A. Potassium channels in cell cycle and cell proliferation. Philos. Trans. R. Soc. Lond. B Biol. Sci. 2014, 369, 20130094. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pini, J.; Giuliano, S.; Matonti, J.; Gannoun, L.; Simkin, D.; Rouleau, M.; Bendahhou, S. Osteogenic and chondrogenic master genes expression is dependent on the Kir2.1 potassium channel through the Bone Morphogenetic Protein pathway. J. Bone. Miner. Res. 2018, 33, 1826–1841. [Google Scholar] [CrossRef]
- Grössinger, E.M.; Kang, M.; Bouchareychas, L.; Sarin, R.; Haudenschild, D.R.; Borodinsky, L.N.; Adamopoulos, I.E. Ca(2+)-dependent regulation of NFATc1 via KCa3.1 in inflammatory osteoclastogenesis. J. Immunol. 2018, 200, 749–757. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, J.E.; Song, M.S.; Shen, Y.; Ryu, P.D.; Lee, S.Y. The role of KV7.3 in regulating osteoblast maturation and mineralization. Int. J. Mol. Sci. 2016, 17, 407. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- George, L.F.; Isner, T.; Bates, E.A. Ion channels in Bone Morphogenetic Protein signaling. Bioelectricity 2019, 1, 46–48. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tristani-Firouzi, M.; Etheridge, S.P. Kir 2.1 channelopathies: The Andersen-Tawil syndrome. Pflug. Arch. 2010, 460, 289–294. [Google Scholar] [CrossRef] [PubMed]
- Hamilton, M.J.; Suri, M. “Electrifying dysmorphology”: Potassium channelopathies causing dysmorphic syndromes. Adv. Genet. 2020, 105, 137–174. [Google Scholar] [PubMed]
- Adams, D.S.; Uzel, S.G.; Akagi, J.; Wlodkowic, D.; Andreeva, V.; Yelick, P.C.; Devitt-Lee, A.; Pare, J.-F.; Levin, M. Bioelectric signalling via potassium channels: A mechanism for craniofacial dysmorphogenesis in KCNJ2-associated Andersen-Tawil Syndrome. J. Physiol. 2016, 594, 3245–3270. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Graham, J.M., Jr.; Zadeh, N.; Kelley, M.; Tan, E.S.; Liew, W.; Tan, V.; Deardorff, M.A.; Wilson, G.N.; Sagi-Dain, L.; Shalev, S.A. KCNK9 imprinting syndrome-further delineation of a possible treatable disorder. Am. J. Med. Genet. A 2016, 170, 2632–2637. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Saint-Jeannet, J.-P.; Moody, S.A. Establishing the pre-placodal region and breaking it into placodes with distinct identities. Dev. Biol. 2014, 389, 13–27. [Google Scholar] [CrossRef] [Green Version]
- Lleras-Forero, L.; Streit, A. Development of the sensory nervous system in the vertebrate head: The importance of being on time. Curr. Opin. Genet. Dev. 2012, 22, 315–322. [Google Scholar] [CrossRef] [PubMed]
- Desban, L.; Prendergast, A.; Roussel, J.; Rosello, M.; Geny, D.; Wyart, C.; Bardet, P.-L. Regulation of the apical extension morphogenesis tunes the mechanosensory response of microvilliated neurons. PLoS Biol. 2019, 17, e3000235. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Atta, K.I. Morphological, anatomical and histological studies on the olfactory organs and eyes of teleost fish: Anguilla anguilla in relation to its feeding habits. J. Basic App. Zool. 2013, 66, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Kasumyan, A.O. The olfactory system in fish: Structure, function, and role in behavior. J. Ichthy 2004, 44, S180. [Google Scholar]
- Roberts, R.B.; Hu, Y.; Albertson, R.C.; Kocher, T.D. Craniofacial divergence and ongoing adaptation via the hedgehog pathway. Proc. Natl. Acad. Sci. USA 2011, 108, 13194–13199. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aoto, K.; Trainor, P.A. Co-ordinated brain and craniofacial development depend upon Patched1/XIAP regulation of cell survival. Hum. Mol. Genet. 2015, 24, 698–713. [Google Scholar] [CrossRef] [Green Version]
- Abrams, S.R.; Reiter, J.F. Ciliary Hedgehog signaling regulates cell survival to build the facial midline. Elife 2021, 10, e68558. [Google Scholar] [CrossRef] [PubMed]
- Andreu-Cervera, A.; Catala, M.; Schneider-Maunoury, S. Cilia, ciliopathies and hedgehog-related forebrain developmental disorders. Neurobiol. Dis. 2021, 150, 105236. [Google Scholar] [CrossRef] [PubMed]
- Yavropoulou, M.P.; Yovos, J.G. The molecular basis of bone mechanotransduction. J. Musculoskelet. Neuronal. Interact. 2016, 16, 221–236. [Google Scholar] [PubMed]
- Gilbert, M.C.; Tetrault, E.; Packard, M.; Navon, D.; Albertson, R.C. Ciliary Rootlet Coiled-Coil 2 (crocc2) Is Associated with Evolutionary Divergence and Plasticity of Cichlid Jaw Shape. Mol. Biol. Evol. 2021, 38, 3078–3092. [Google Scholar] [CrossRef] [PubMed]
- Santagati, F.; Rijli, F.M. Cranial neural crest and the building of the vertebrate head. Nat. Rev. Neurosci. 2003, 4, 806–818. [Google Scholar] [CrossRef]
- Schilling, T.F. Genetic analysis of craniofacial development in the vertebrate embryo. Bioessays 1997, 19, 459–468. [Google Scholar] [CrossRef] [PubMed]
- Berndt, J.D.; Halloran, M.C. Semaphorin 3d promotes cell proliferation and neural crest cell development downstream of TCF in the zebrafish hindbrain. Development 2006, 133, 3983–3992. [Google Scholar] [CrossRef] [Green Version]
- Yu, H.-H.; Moens, C.B. Semaphorin signaling guides cranial neural crest cell migration in zebrafish. Dev. Biol. 2005, 280, 373–385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, Z.Z.; Guo, J.; Lu, Y.; Liu, W.; Fu, X.; Yao, T.; Zhou, Y.; Xu, H.A. Sema3E is required for migration of cranial neural crest cells in zebrafish: Implications for the pathogenesis of CHARGE syndrome. Int. J. Exp. Pathol. 2019, 100, 234–243. [Google Scholar] [CrossRef] [PubMed]
- Theveneau, E.; Mayor, R. Neural crest migration: Interplay between chemorepellents, chemoattractants, contact inhibition, epithelial-mesenchymal transition, and collective cell migration. Wiley Interdiscip. Rev. Dev. Biol. 2012, 1, 435–445. [Google Scholar] [CrossRef] [PubMed]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| QTL Phenotype | LG | Number of Genes in Interval | Candidate Genes related to Craniofacial Development or Disease |
|---|---|---|---|
| PC2 lateral | 2 | 117 | fgf13, zic3 |
| Opercle-to-mandible Length | 6 | 573 | fgf8b, fgf20a, p300 (Rubinstein-Taybi syndrome) |
| Opercle-to-midline width | 6 | 510 | fgf8b, fgf20a, p300 (Rubinstein-Taybi syndrome) |
| PC2 lateral | 6 | 193 | axin2, smarce1 |
| Dorsal–pelvic length | 7 | 424 | alx4, hdac10, sema3a, sema3d, sema3e, smad3, smad6, wnt7bb |
| PC2 lateral | 7 | 635 | bmpr1bb, foxd1, lhx6, notch1a, pax8, smad4a, smad7, tbx3a |
| Preorbital length | 7 | 702 | apc, bmp1, bmp10-like, lhx6, nodal2, fgfr1a, smad2, smad4a, smad7, pax8, tbx3a, tbx5a, tcf4 |
| PC2 lateral | 10 | 149 | fgf1, fgf13b, spry4, tcf7 |
| Pelvic–snout length | 12 | 782 | bmp3, fgf10, fgf5, foxd4, lhx6b, ptch1, smarcad1, tbx5 |
| PC4 lateral | 12 | 246 | dlx5a, dlx6a, hoxa gene cluster, smarcc1b, sp8a, twist1a |
| Mouth angle | 13 | 300 | dkk1, grem2a, pax2a, sufu, wnt8b |
| Eye area | 15 | 158 | bmp2, dll4, med23 |
| Opercle-to-midline width | 15 | 10 | fzd3a |
| Eye area | 16 | 824 | acvr1, acvr1c, bbs5 (Bardet-Biedl ciliopathy), dlx1a, dlx2a, epha3, evx2, frzb, fzd5, hoxd gene cluster, pou3f3 (Williams–Beuren syndrome), satb2, shox, tbx15, tgfbr2l, zic2 (Holoprosencephaly), zic5 |
| Mandible angle | 16 | 504 | frzb, fzd5, pou3f3 (Williams–Beuren syndrome), satb2, shox, tbx15, tgfbr2l, zic2 (Holoprosencephaly), zic5 |
| PC3 lateral | 17 | 248 | dkk2 |
| Preorbital length | 17 | 3 | ephrin type-B receptor 1-B |
| Head proportion | 20 | 972 | alx3, hes4, hoxC gene cluster, irx7, wnt1, wnt10b, wnt2ba, wnt5a, wnt7a |
| Dorsal–pelvic length | 20 | 351 | alx3, hes4, hoxC gene cluster, wnt1, wnt10b, wnt2ba, wnt5a |
| PC3 lateral | 20 | 226 | hes4 |
| PC3 lateral | 22 | 246 | col1a2, dlx5, dlx6, hoxA gene cluster, smarcc1b |
| PC2 lateral | 23 | 556 | crocc2, fgf22, foxd2, lhx8, notch2, prdm5, tgfbr3 |
| PC4 lateral | 23 | 683 | acvr2a, bmpr2, fgf14, spry2, zeb2 (Mowat-Wilson neurocristopathy) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
DeLorenzo, L.; DeBrock, V.; Carmona Baez, A.; Ciccotto, P.J.; Peterson, E.N.; Stull, C.; Roberts, N.B.; Roberts, R.B.; Powder, K.E. Morphometric and Genetic Description of Trophic Adaptations in Cichlid Fishes. Biology 2022, 11, 1165. https://doi.org/10.3390/biology11081165
DeLorenzo L, DeBrock V, Carmona Baez A, Ciccotto PJ, Peterson EN, Stull C, Roberts NB, Roberts RB, Powder KE. Morphometric and Genetic Description of Trophic Adaptations in Cichlid Fishes. Biology. 2022; 11(8):1165. https://doi.org/10.3390/biology11081165
Chicago/Turabian StyleDeLorenzo, Leah, Victoria DeBrock, Aldo Carmona Baez, Patrick J. Ciccotto, Erin N. Peterson, Clare Stull, Natalie B. Roberts, Reade B. Roberts, and Kara E. Powder. 2022. "Morphometric and Genetic Description of Trophic Adaptations in Cichlid Fishes" Biology 11, no. 8: 1165. https://doi.org/10.3390/biology11081165