
Gene Editing of the Catfish Gonadotropin-Releasing Hormone Gene and Hormone Therapy to Control the Reproduction in Channel Catfish, Ictalurus punctatus

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Construction of Plasmids
2.2. TALEN Plasmid Replication, Extraction and Dilution
2.3. Experimental Brood Stock and Gametes
2.4. Fertilization, Double Electroporation and Embryo Incubation
2.5. Sample Collection and Mutation Detection for P1 Generation
2.6. TA Clone and Sequencing
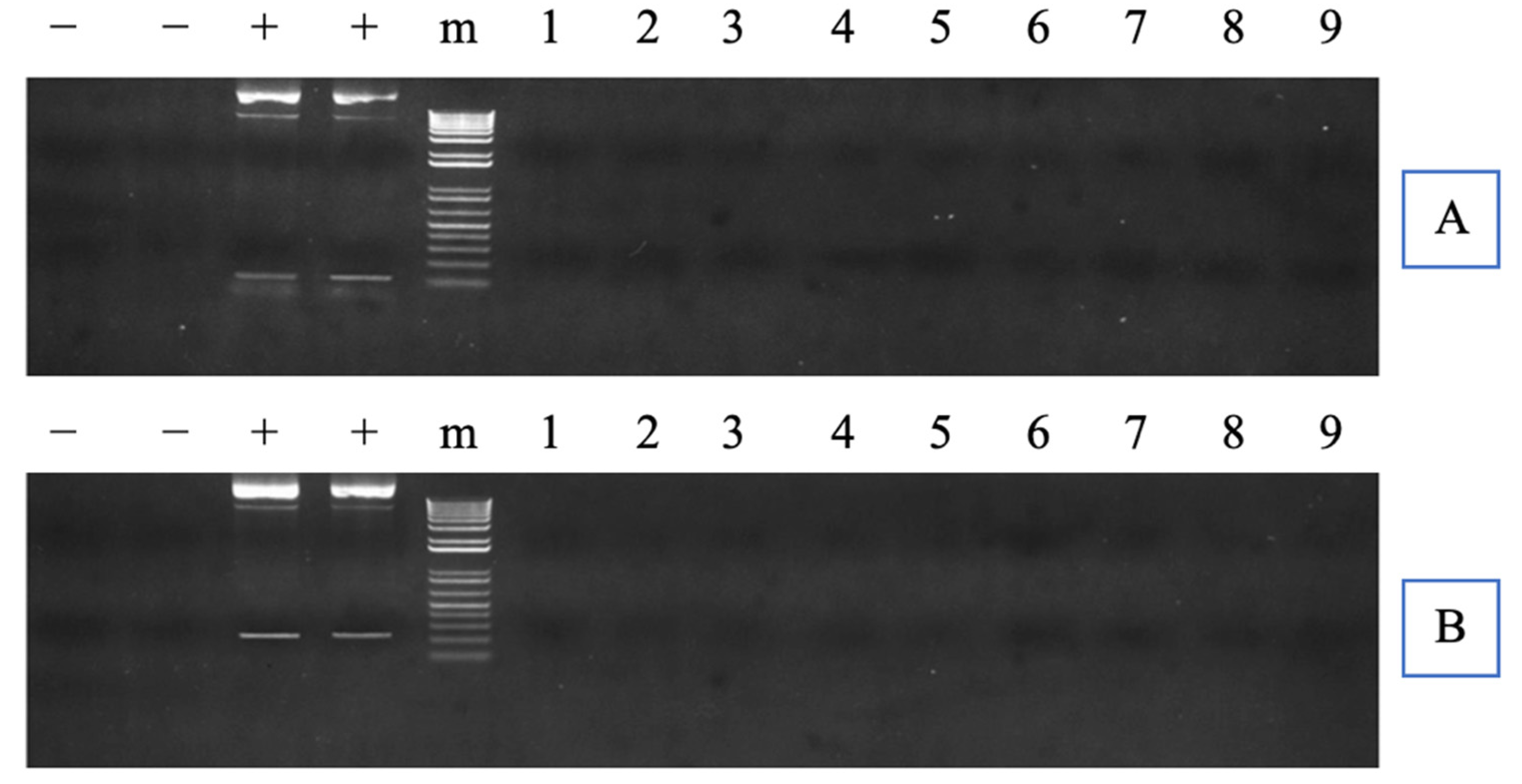
2.7. Plasmid Integration Inspection
2.8. Reproductive Evaluation
2.9. Hormone Therapy
2.10. F1 Fish Culture
2.11. Sample Collection and Genotype Analysis for F1 Generation
2.12. Pleiotropic Effects
2.13. Statistical Analysis
3. Results
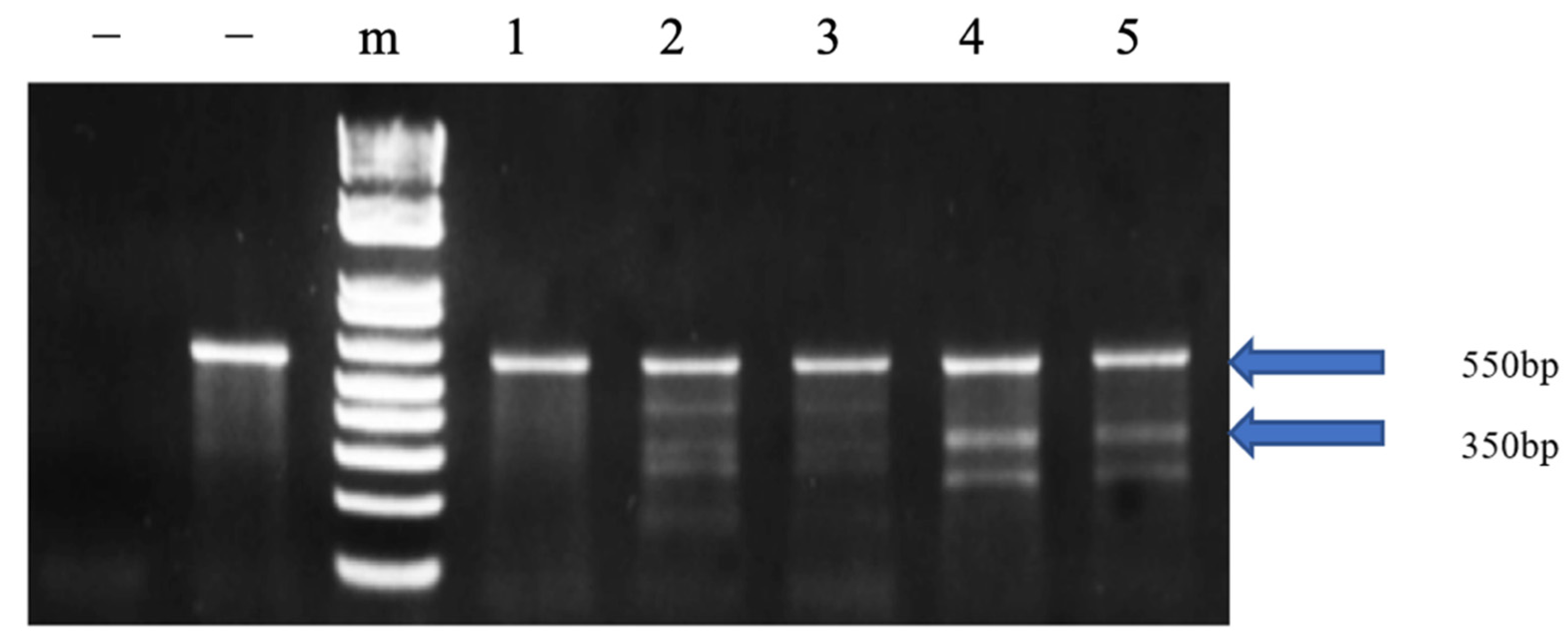
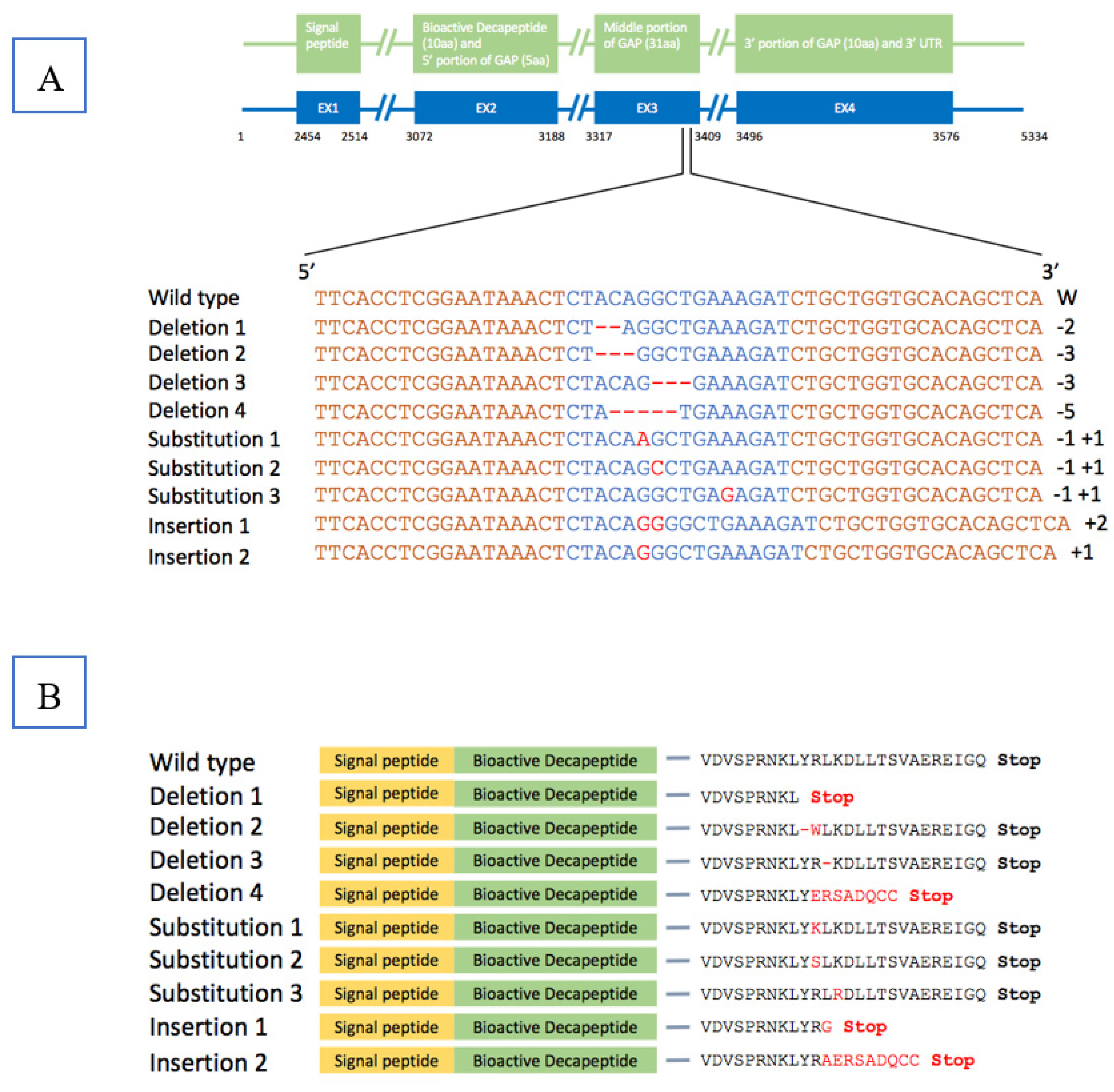
3.1. Analysis of Mutation Efficiency and Plasmid Integration in the P1 Generation
3.2. Spawning Experiments and Hormone Therapy
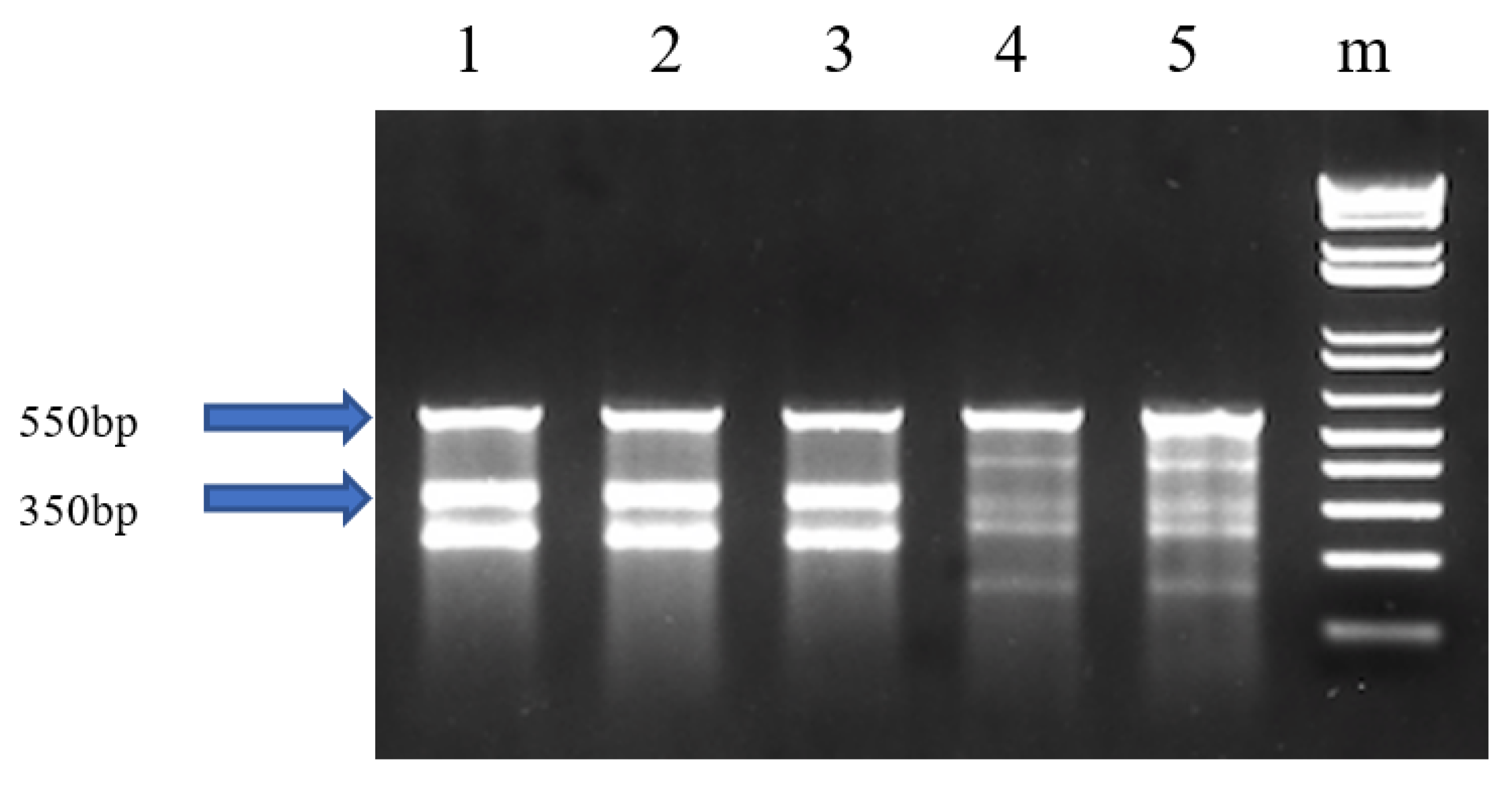
3.3. Analysis of Mutation Efficiency for the F1 Generation
3.4. Pleiotropic Effects
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Liu, Z.; Liu, S.; Yao, J.; Bao, L.; Zhang, J.; Li, Y.; Jiang, C.; Sun, L.; Wang, R.; Zhang, Y.; et al. The Channel Catfish Genome Sequence Provides Insights into the Evolution of Scale Formation in Teleosts. Nat. Commun. 2016, 7, 11757. [Google Scholar] [CrossRef] [PubMed]
- Dong, S.; Perera, D.A.; Vo, K.; Odin, R.; Alsaquifi, A.; Fobes, M.; Elaswad, A.; Ye, Z.; Qin, Z.; Li, H.; et al. Channel Catfish Ictalurus punctatus Strain Comparison for Induced Ovulation in the Early Spawning Season to Produce Channel Catfish ♀×blue Catfish, I. Furcatus ♂ Hybrid Catfish Embryos. Aquaculture 2017, 471, 185–189. [Google Scholar] [CrossRef]
- U.S. Catfish Database. 2014. Available online: https://www.agecon.msstate.edu/whatwedo/budgets/docs/catfish2014.pdf (accessed on 13 March 2022).
- Abass, N.Y.; Elwakil, H.E.; Hemeida, A.A.; Abdelsalam, N.R.; Ye, Z.; Su, B.; Alsaqufi, A.S.; Weng, C.-C.; Trudeau, V.L.; Dunham, R.A. Genotype–Environment Interactions for Survival at Low and Sub-Zero Temperatures at Varying Salinity for Channel Catfish, Hybrid Catfish and Transgenic Channel Catfish. Aquaculture 2016, 458, 140–148. [Google Scholar] [CrossRef]
- Abdelrahman, H.; The Aquaculture Genomics, Genetics and Breeding Workshop; ElHady, M.; Alcivar-Warren, A.; Allen, S.; Al-Tobasei, R.; Bao, L.; Beck, B.; Blackburn, H.; Bosworth, B.; et al. Aquaculture Genomics, Genetics and Breeding in the United States: Current Status, Challenges, and Priorities for Future Research. BMC Genom. 2017, 18, 191. [Google Scholar] [CrossRef] [Green Version]
- Flajšhans, M.; Gela, D.; Kocour, M.; Buchtová, H.; Rodina, M.; Pšenička, M.; Kašpar, V.; Piačková, V.; Sudová, E.; Linhart, O. A review on the potential of triploid tench for aquaculture. Rev. Fish. Biol. Fish. 2010, 20, 317–329. [Google Scholar] [CrossRef]
- Shi, H.; Zhou, T.; Wang, X.; Yang, Y.; Wu, C.; Liu, S.; Bao, L.; Li, N.; Yuan, Z.; Jin, Y.; et al. Genome-Wide Association Analysis of Intra-Specific QTL Associated with the Resistance for Enteric Septicemia of Catfish. Mol. Genet. Genom. 2018, 293, 1365–1378. [Google Scholar] [CrossRef] [PubMed]
- Hedrick, P.W. Erratum: Invasion of Transgenes from Salmon or Other Genetically Modified Organisms into Natural Populations. Can. J. Fish. Aquat. Sci. 2001, 58, 2317. [Google Scholar] [CrossRef]
- Dunham, R.A. Aquaculture and Fisheries Biotechnology: Genetic Approaches, 2nd ed.; Dunham, R.A., Ed.; CABI Publishing: Wallingford, UK, 2011. [Google Scholar]
- Li, H.; Su, B.; Qin, G.; Ye, Z.; Alsaqufi, A.; Perera, D.; Shang, M.; Odin, R.; Vo, K.; Drescher, D.; et al. Salt Sensitive Tet-off-like Systems to Knockdown Primordial Germ Cell Genes for Repressible Transgenic Sterilization in Channel Catfish, Ictalurus punctatus. Mar. Drugs 2017, 15, 155. [Google Scholar] [CrossRef]
- Li, H.; Su, B.; Qin, G.; Ye, Z.; Elaswad, A.; Alsaqufi, A.; Perera, D.A.; Qin, Z.; Odin, R.; Vo, K.; et al. Repressible Transgenic Sterilization in Channel Catfish, Ictalurus punctatus, by Knockdown of Primordial Germ Cell Genes with Copper-Sensitive Constructs. Mar. Biotechnol. 2018, 20, 324–342. [Google Scholar] [CrossRef]
- Thresher, R.; Grewe, P.; Patil, J.G.; Whyard, S.; Templeton, C.M.; Chaimongol, A.; Hardy, C.M.; Hinds, L.A.; Dunham, R. Development of Repressible Sterility to Prevent the Establishment of Feral Populations of Exotic and Genetically Modified Animals. Aquaculture 2009, 290, 104–109. [Google Scholar] [CrossRef]
- Qin, Z. Gene Editing of Luteinizing Hormone, Follicle-Stimulating Hormone and Gonadotropin-Releasing Hormone Genes to Sterilize Channel Catfish, Ictalurus punctatus, Using Zinc Finger Nuclease, Transcription Activator-like Effector Nuclease and Clustered Regularly Interspaced Short Palindromic Repeats/Cas9 Technologies. Ph.D. Thesis, Auburn University, Auburn, AL, USA, 2015. [Google Scholar]
- Qin, Z.; Li, Y.; Su, B.; Cheng, Q.; Ye, Z.; Perera, D.A.; Fobes, M.; Shang, M.; Dunham, R.A. Editing of the Luteinizing Hormone Gene to Sterilize Channel Catfish, Ictalurus punctatus, Using a Modified Zinc Finger Nuclease Technology with Electroporation. Mar. Biotechnol. 2016, 18, 255–263. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Zhu, B.; Ge, W. Genetic Analysis of Zebrafish Gonadotropin (FSH and LH) Functions by TALEN-Mediated Gene Disruption. Mol. Endocrinol. 2015, 29, 76–98. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mylonas, C.C.; Fostier, A.; Zanuy, S. Broodstock Management and Hormonal Manipulations of Fish Reproduction. Gen. Comp. Endocrinol. 2010, 165, 516–534. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Choi, Y.J.; Kim, N.N.; Habibi, H.R.; Choi, C.Y. Effects of Gonadotropin Inhibitory Hormone or Gonadotropin-Releasing Hormone on Reproduction-Related Genes in the Protandrous Cinnamon Clownfish, Amphiprion melanopus. Gen. Comp. Endocrinol. 2016, 235, 89–99. [Google Scholar] [CrossRef]
- Temple, J.L.; Millar, R.P.; Rissman, E.F. An Evolutionarily Conserved Form of Gonadotropin-Releasing Hormone Coordinates Energy and Reproductive Behavior. Endocrinology 2003, 144, 13–19. [Google Scholar] [CrossRef]
- Klausen, C.; Chang, J.P.; Habibi, H.R. The Effect of Gonadotropin-Releasing Hormone on Growth Hormone and Gonadotropin Subunit Gene Expression in the Pituitary of Goldfish, Carassius auratus. Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2001, 129, 511–516. [Google Scholar] [CrossRef]
- Weber, G.M.; Powell, J.F.; Park, M.; Fischer, W.H.; Craig, A.G.; Rivier, J.E.; Nanakorn, U.; Parhar, I.S.; Ngamvongchon, S.; Grau, E.G.; et al. Evidence That Gonadotropin-Releasing Hormone (GnRH) Functions as a Prolactin-Releasing Factor in a Teleost Fish (Oreochromis mossambicus) and Primary Structures for Three Native GnRH Molecules. J. Endocrinol. 1997, 155, 121–132. [Google Scholar] [CrossRef] [Green Version]
- Dubois, E.A.; Slob, S.; Zandbergen, M.A.; Peute, J.; Goos, H.J. Gonadal Steroids and the Maturation of the Species-Specific Gonadotropin-Releasing Hormone System in Brain and Pituitary of the Male African Catfish (Clarias gariepinus). Comp. Biochem. Physiol. B Biochem. Mol. Biol. 2001, 129, 381–387. [Google Scholar] [CrossRef]
- Kumar, K.V.S.S.; Swapna, I.; Gupta, A.D.; Majumdar, K.C.; Senthilkumaran, B. Effect of Thyroid Hormone Depletion on Monoamines and Expression Patterns of Catfish GnRH in the Air-Breathing Catfish, Clarias gariepinus. Fish. Physiol. Biochem. 2005, 31, 189–192. [Google Scholar] [CrossRef]
- Zmora, N.; González-Martínez, D.; Muñoz-Cueto, J.A.; Madigou, T.; Mañanos-Sanchez, E.; Doste, S.Z.; Zohar, Y.; Kah, O.; Elizur, A. The GnRH System in the European Sea Bass (Dicentrarchus labrax). J. Endocrinol. 2002, 172, 105–116. [Google Scholar] [CrossRef] [Green Version]
- Nikolics, K.; Mason, A.J.; Szönyi, E.; Ramachandran, J.; Seeburg, P.H. A Prolactin-Inhibiting Factor within the Precursor for Human Gonadotropin-Releasing Hormone. Nature 1985, 316, 511–517. [Google Scholar] [CrossRef]
- Millar, R.P.; Wormald, P.J.; Milton, R.C. Stimulation of Gonadotropin Release by a Non-GnRH Peptide Sequence of the GnRH Precursor. Science 1986, 232, 68–70. [Google Scholar] [CrossRef]
- Andersen, O.; Klungland, H. The Salmon GnRH Encoding Gene in Teleost Fish. Int. Rev. Cytol. 1993, 147, 165–191. [Google Scholar] [CrossRef] [PubMed]
- Wetsel, W.C.; Srinivasan, S. Pro-GnRH Processing. Prog. Brain Res. 2002, 141, 221–241. [Google Scholar] [CrossRef] [PubMed]
- Ogiwara, K.; Fujimori, C.; Rajapakse, S.; Takahashi, T. Characterization of Luteinizing Hormone and Luteinizing Hormone Receptor and Their Indispensable Role in the Ovulatory Process of the Medaka. PLoS ONE 2013, 8, e54482. [Google Scholar] [CrossRef] [PubMed]
- Kucharczyk, D.; Kucharczyk, D.J.; Nowosad, J.; Omirzhanova, N. Optimization of Artificial Insemination Outcomes of African Catfish (Clarias gariepinus) with Differing Hatchery Conditions. Anim. Reprod. Sci. 2019, 211, 106222. [Google Scholar] [CrossRef]
- Dunham, R.A.; Elaswad, A.; Qin, Z. Gene Editing in Channel Catfish via Double Electroporation of Zinc-Finger Nucleases. Methods Mol. Biol. 2018, 1867, 201–214. [Google Scholar] [CrossRef]
- Su, B.; Peatman, E.; Shang, M.; Thresher, R.; Grewe, P.; Patil, J.; Pinkert, C.A.; Irwin, M.H.; Li, C.; Perera, D.A.; et al. Expression and Knockdown of Primordial Germ Cell Genes, Vasa, Nanos and Dead End in Common Carp (Cyprinus carpio) Embryos for Transgenic Sterilization and Reduced Sexual Maturity. Aquaculture 2014, 420–421, S72–S84. [Google Scholar] [CrossRef]
- Cheng, Q.; Su, B.; Qin, Z.; Weng, C.-C.; Yin, F.; Zhou, Y.; Fobes, M.; Perera, D.A.; Shang, M.; Soller, F.; et al. Interaction of Diet and the Masou Salmon Δ5-Desaturase Transgene on Δ6-Desaturase and Stearoyl-CoA Desaturase Gene Expression and N-3 Fatty Acid Level in Common Carp (Cyprinus carpio). Transgenic Res. 2014, 23, 729–742. [Google Scholar] [CrossRef]
- Small, B.C.; Chatakondi, N. Efficacy of Formalin as an Egg Disinfectant for Improving Hybrid Catfish (Channel Catfish × Blue Catfish) Hatching Success. N. Am. J. Aquac. 2006, 68, 9–13. [Google Scholar] [CrossRef]
- Zuberi, A.; Gima, M.; Gima, A.; Hutson, A.; Chaimongkol, A.; Li, M.; Umali-Maceina, G.; Dunham, R. Diet Fed to Brood Stock Effects the Growth of Channel Catfish (Ictalurus punctatus L.) Fry: Brood Stock Diet Effects Growth of Catfish Fry. Aquac. Res. 2011, 42, 1899–1904. [Google Scholar] [CrossRef]
- Quintero, H.E.; Durland, E.; Allen Davis, D.; Dunham, R. Effect of Lipid Supplementation on Reproductive Performance of Female Channel Catfish, Ictalurus punctatus, Induced and Strip-Spawned for Hybridization: Effect of Lipid Supplementation on Reproductive Performance. Aquac. Nutr. 2011, 17, 117–129. [Google Scholar] [CrossRef]
- Kim, Y.; Kweon, J.; Kim, A.; Chon, J.K.; Yoo, J.Y.; Kim, H.J.; Kim, S.; Lee, C.; Jeong, E.; Chung, E.; et al. A Library of TAL Effector Nucleases Spanning the Human Genome. Nat. Biotechnol. 2013, 31, 251–258. [Google Scholar] [CrossRef] [PubMed]
- Reyon, D.; Tsai, S.Q.; Khayter, C.; Foden, J.A.; Sander, J.D.; Joung, J.K. FLASH Assembly of TALENs for High-Throughput Genome Editing. Nat. Biotechnol. 2012, 30, 460–465. [Google Scholar] [CrossRef]
- Elaswad, A.; Khalil, K.; Ye, Z.; Liu, Z.; Liu, S.; Peatman, E.; Odin, R.; Vo, K.; Drescher, D.; Gosh, K.; et al. Effects of CRISPR/Cas9 Dosage on TICAM1 and RBL Gene Mutation Rate, Embryonic Development, Hatchability and Fry Survival in Channel Catfish. Sci. Rep. 2018, 8, 16499. [Google Scholar] [CrossRef]
- Qin, G. Gene Editing of Hormone Therapy to Control Reproduction in Channel Catfish, Ictalurus punctatus. Ph.D. Thesis, Auburn University, Auburn, AL, USA, 2019. [Google Scholar]
- Le, Q.A.; Tanihara, F.; Wittayarat, M.; Namula, Z.; Sato, Y.; Lin, Q.; Takebayashi, K.; Hirata, M.; Otoi, T. Comparison of the Effects of Introducing the CRISPR/Cas9 System by Microinjection and Electroporation into Porcine Embryos at Different Stages. BMC Res. Notes 2021, 14, 7. [Google Scholar] [CrossRef]
- Takahashi, A.; Kanda, S.; Abe, T.; Oka, Y. Evolution of the Hypothalamic-Pituitary-Gonadal Axis Regulation in Vertebrates Revealed by Knockout Medaka. Endocrinology 2016, 157, 3994–4002. [Google Scholar] [CrossRef]
- Spicer, O.S.; Wong, T.-T.; Zmora, N.; Zohar, Y. Targeted Mutagenesis of the Hypophysiotropic Gnrh3 in Zebrafish (Danio rerio) Reveals No Effects on Reproductive Performance. PLoS ONE 2016, 11, e0158141. [Google Scholar] [CrossRef] [Green Version]
- Marvel, M.; Spicer, O.S.; Wong, T.-T.; Zmora, N.; Zohar, Y. Knockout of the Gnrh Genes in Zebrafish: Effects on Reproduction and Potential Compensation by Reproductive and Feeding-Related Neuropeptides. Biol. Reprod. 2018, 99, 565–577. [Google Scholar] [CrossRef]
- Drabkin, M.; Birk, O.S.; Birk, R. Heterozygous versus Homozygous Phenotype Caused by the Same MC4R Mutation: Novel Mutation Affecting a Large Consanguineous Kindred. BMC Med. Genet. 2018, 19, 135. [Google Scholar] [CrossRef] [Green Version]
- Saville, K.J.; Belote, J.M. Identification of an Essential Gene, l(3)73Ai, with a Dominant Temperature-Sensitive Lethal Allele, Encoding a Drosophila Proteasome Subunit. Proc. Natl. Acad. Sci. USA 1993, 90, 8842–8846. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schweisguth, F. Dominant-Negative Mutation in the Beta2 and Beta6 Proteasome Subunit Genes Affect Alternative Cell Fate Decisions in the Drosophila Sense Organ Lineage. Proc. Natl. Acad. Sci. USA 1999, 96, 11382–11386. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mussolino, C.; Alzubi, J.; Fine, E.J.; Morbitzer, R.; Cradick, T.J.; Lahaye, T.; Bao, G.; Cathomen, T. TALENs Facilitate Targeted Genome Editing in Human Cells with High Specificity and Low Cytotoxicity. Nucleic Acids Res. 2014, 42, 6762–6773. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dreyer, A.-K.; Hoffmann, D.; Lachmann, N.; Ackermann, M.; Steinemann, D.; Timm, B.; Siler, U.; Reichenbach, J.; Grez, M.; Moritz, T.; et al. TALEN-Mediated Functional Correction of X-Linked Chronic Granulomatous Disease in Patient-Derived Induced Pluripotent Stem Cells. Biomaterials 2015, 69, 191–200. [Google Scholar] [CrossRef] [PubMed]
- Veres, A.; Gosis, B.S.; Ding, Q.; Collins, R.; Ragavendran, A.; Brand, H.; Erdin, S.; Cowan, C.A.; Talkowski, M.E.; Musunuru, K. Low Incidence of Off-Target Mutations in Individual CRISPR-Cas9 and TALEN Targeted Human Stem Cell Clones Detected by Whole-Genome Sequencing. Cell Stem Cell 2014, 15, 254. [Google Scholar] [CrossRef] [Green Version]
- Cressey, D. Aquaculture: Future Fish. Nature 2009, 458, 398–400. [Google Scholar] [CrossRef] [Green Version]
- National Oceanic and Atmospheric Administration. 2011. Available online: https://www.noaa.gov/stories/what-is-aquaculture (accessed on 22 February 2022).
- Dunham, R.A.; Elaswad, A. Catfish Biology and Farming. Annu. Rev. Anim. Biosci. 2018, 6, 305–325. [Google Scholar] [CrossRef]
- Dunham, R.A.; Ramboux, A.C.; Duncan, P.L.; Hayat, M.; Chen, T.T.; Lin, C.M.; Kight, K.; Gonzalez-Villasenor, I.; Powers, D.A. Transfer, Expression, and Inheritance of Salmonid Growth Hormone Genes in Channel Catfish, Ictalurus punctatus, and Effects on Performance Traits. Mol. Mar. Biol. Biotechnol. 1992, 1, 380–389. [Google Scholar]
- Palmiter, R.D.; Brinster, R.L.; Hammer, R.E.; Trumbauer, M.E.; Rosenfeld, M.G.; Birnberg, N.C.; Evans, R.M. Dramatic Growth of Mice That Develop from Eggs Microinjected with Metallothionein-Growth Hormone Fusion Genes. 1982. Biotechnology 1992, 24, 429–433. [Google Scholar]
- Devlin, R.H.; Johnsson, J.I.; Smailus, D.E.; Biagi, C.A.; Jönsson, E.; Björnsson, B.T. Increased Ability to Compete for Food by Growth Hormone-Transgenic Coho Salmon Oncorhynchus kisutch (Walbaum): Competitive Food Intake of Transgenic SalmonR H Devlinet Al. Aquac. Res. 1999, 30, 479–482. [Google Scholar] [CrossRef]
- Du, S.J.; Gong, Z.; Fletcher, G.L.; Shears, M.A.; King, M.J.; Idler, D.R.; Hew, C.L. Growth Enhancement in Transgenic Atlantic Salmon by the Use of an “All Fish” Chimeric Growth Hormone Gene Construct. Nat. Biotechnol. 1992, 10, 176–181. [Google Scholar] [CrossRef] [PubMed]
- Agellon, L.B.; Emery, C.J.; Jones, J.M.; Davies, S.L.; Dingle, A.D.; Chen, T.T. Promotion of Rapid Growth of Rainbow Trout (Salmo gairdneri) by a Recombinant Fish Growth Hormone. Can. J. Fish. Aquat. Sci. 1988, 45, 146–151. [Google Scholar] [CrossRef]
- Chatakondi, N.; Lovell, R.T.; Duncan, P.L.; Hayat, M.; Chen, T.T.; Powers, D.A.; Weete, J.D.; Cummins, K.; Dunham, R.A. Body Composition of Transgenic Common Carp, Cyprinus carpio, Containing Rainbow Trout Growth Hormone Gene. Aquaculture 1995, 138, 99–109. [Google Scholar] [CrossRef]
- Bugg, W. The Effects of the Masou Salmon Delta5-Desaturase Transgene on n-3 Fatty Acid Production in F1 Transgenic Common Carp (Cyprinus carpio) and Channel Catfish (Ictalurus punctatus). Ph.D. Thesis, Auburn University, Auburn, AL, USA, 2017. [Google Scholar]
- Wang, Y.-D.; Peng, K.-C.; Wu, J.-L.; Chen, J.-Y. Transgenic Expression of Salmon Delta-5 and Delta-6 Desaturase in Zebrafish Muscle Inhibits the Growth of Vibrio Alginolyticus and Affects Fish Immunomodulatory Activity. Fish Shellfish Immunol. 2014, 39, 223–230. [Google Scholar] [CrossRef] [PubMed]
- Peng, K.-C.; Pan, C.-Y.; Chou, H.-N.; Chen, J.-Y. Using an Improved Tol2 Transposon System to Produce Transgenic Zebrafish with Epinecidin-1 Which Enhanced Resistance to Bacterial Infection. Fish Shellfish Immunol. 2010, 28, 905–917. [Google Scholar] [CrossRef]
- Srisai, D.; Gillum, M.P.; Panaro, B.L.; Zhang, X.-M.; Kotchabhakdi, N.; Shulman, G.I.; Ellacott, K.L.J.; Cone, R.D. Characterization of the Hyperphagic Response to Dietary Fat in the MC4R Knockout Mouse. Endocrinology 2011, 152, 890–902. [Google Scholar] [CrossRef] [Green Version]
- Dong, Z.; Ge, J.; Li, K.; Xu, Z.; Liang, D.; Li, J.; Li, J.; Jia, W.; Li, Y.; Dong, X.; et al. Heritable Targeted Inactivation of Myostatin Gene in Yellow Catfish (Pelteobagrus fulvidraco) Using Engineered Zinc Finger Nucleases. PLoS ONE 2011, 6, e28897. [Google Scholar] [CrossRef] [Green Version]
- Khalil, K.; Elayat, M.; Khalifa, E.; Daghash, S.; Elaswad, A.; Miller, M.; Abdelrahman, H.; Ye, Z.; Odin, R.; Drescher, D.; et al. Generation of Myostatin Gene-Edited Channel Catfish (Ictalurus punctatus) via Zygote Injection of CRISPR/Cas9 System. Sci. Rep. 2017, 7, 7301. [Google Scholar] [CrossRef] [Green Version]
- Nielsen, K.M. Transgenic Organisms—Time for Conceptual Diversification? Nat. Biotechnol. 2003, 21, 227–228. [Google Scholar] [CrossRef]
- Jhingan, E.; Devlin, R.H.; Iwama, G.K. Disease Resistance, Stress Response and Effects of Triploidy in Growth Hormone Transgenic Coho Salmon. J. Fish. Biol. 2003, 63, 806–823. [Google Scholar] [CrossRef]
- Razak, S.A.; Hwang, G.L.; Rahman, M.A.; Maclean, N. Growth Performance and Gonadal Development of Growth Enhanced Transgenic Tilapia Oreochromis niloticus (L.) Following Heat-Shock-Induced Triploidy. Mar. Biotechnol. 1999, 1, 533–544. [Google Scholar] [CrossRef] [PubMed]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Targeting Site | Sequence | Annealing Temperature | Product Length (bp) |
|---|---|---|---|
| cfGnRH Forward | 5′-ATGGATGCTGTCTTTGTTTTCC-3′ | 60 ° C | 550 |
| cfGnRH Reverse | 5′-CCACACGAAATAAAGGCAAAG-3′ | ||
| CMV promoter Forward | 5′-AACAACAACGGCGGTAAG-3′ | 60 ° C | 114 |
| CMV promoter Reverse | 5′-CCCATTATTGTTCGCGATTG-3′ | ||
| TAL Repeats Forward | 5′-GCATGACGGAGGGAAAC-3′ | 60 ° C | 215 |
| TAL Repeats Reverse | 5′-CCATTATTGTTCGCGATTGA-3′ |
| Before Hormone Therapy (2016) | After Hormone Therapy (2016) | After Hormone Therapy (2017) | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| Fish N | Spawned Fish N | Spawning Rate (%) a | Mean Hatch Rate ± SEM (%) | Fish N | Spawned Fish N b | Spawning Rate (%) | Mean Hatch Rate (%) | Fish N | Spawned Fish N | Spawning Rate c (%) | Mean Hatch Rate (%) d | ||
| cfGnRH | F | 5 | 1 | 20.0 | 66.0 * | 3 | 0 | 0 | / | 6 | 3 | 50.0 | 72.1 ± 0.07 |
| M | 8 | 4 | 50.0 | 2.0 ± 0.01 * | 3 | 0 | 0 | / | 5 | 3 | 60.0 | 72.1 ± 0.07 | |
| nCTRL | F and M | 5 | 3 | 60.0 | 74.3 ± 0.02 * | / | / | / | / | 7 | 5 | 71.4 | 32.3 ± 0.20 |
| iCTRL | F and M | / | / | / | / | / | / | / | / | 3 | 2 | 66.7 | 56.5 ± 0.34 |
| Treatment | P1 Embryo and 6-Month-Old Fingerlings | Genotype | 3-Year-Old P1 Fish in 2016 (Recirculating) | 4-Year-Old P1 Fish in 2017 | |||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Recirculating System | Pond | ||||||||||
| N Eggs | N Hatched | Hatch Rate (%) a | Survival Rate (%) b | N Fish | Survival Rate (%) c | N Fish | Survival Rate (%) c | N Fish | Survival Rate (%) c | ||
| Electroporated with TALEN plasmids | 200 | 76 | 38.0 | 67.1 | Mutants | 42 | 50.0 | 12 | 41.2 | 32 | 46.9 |
| Electroporated without plasmids | 200 | 84 | 42.0 | 75.0 | Non-mutants | 18 | 33.3 | 8 | 37.5 | 6 | 33.3 |
| Different Families of F1 Offspring | Survival Rate | Mutation Rate * | Mean Body Weight (g) ± SEM of Mutant Fish a | Mean Body Weight (g) ± SEM of Non-mutant Fish a | ||||
|---|---|---|---|---|---|---|---|---|
| N Fish | N Fish Survived | Survival Rate (%) | N Fish Sampled | N Mutant Fish | Mutation Rate (%) | |||
| 2016 GnRH-1 | 30 | 14 | 46.7 | 30 | 18 | 60.0 | 164.5 ± 5.4 | 166.6 ± 7.8 |
| 2016 GnRH-2 | 30 | 12 | 40.0 | 30 | 19 | 63.3 | 154.3 ± 6.3 | 161.9 ± 12.4 |
| 2016 GnRH-3 | 30 | 22 | 73.3 | 30 | 23 | 76.7 | 70.7 ± 3.7 | 68.3 ± 6.3 |
| 2016 GnRH-4 | 30 | 16 | 53.3 | 30 | 20 | 66.7 | 154.9 ± 8.5 | 170.5 ± 11.8 |
| 2017 GnRH-1 | 300 | 224 | 74.7 | 100 | 53 | 53.0 c | 6.1 ± 0.2 | 5.9 ± 0.2 |
| 2017 GnRH-1 fish died from columnaris | / | / | / | 56 | 36 | 64.3 c | 13.13 ± 0.5 b | 13.30 ± 0.8 b |
| 2017 GnRH-2 | 200 | 185 | 92.5 | 30 | 14 | 46.7 | 10.0 ± 0.7 | 9.4 ± 0.6 |
| 2017 GnRH-3 | 300 | 274 | 91.3 | 100 | 51 | 51.0 | 6.8 ± 0.2 | 6.7 ± 0.2 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Qin, G.; Qin, Z.; Lu, C.; Ye, Z.; Elaswad, A.; Bangs, M.; Li, H.; Zhang, Y.; Huang, Y.; Shi, H.; et al. Gene Editing of the Catfish Gonadotropin-Releasing Hormone Gene and Hormone Therapy to Control the Reproduction in Channel Catfish, Ictalurus punctatus. Biology 2022, 11, 649. https://doi.org/10.3390/biology11050649
Qin G, Qin Z, Lu C, Ye Z, Elaswad A, Bangs M, Li H, Zhang Y, Huang Y, Shi H, et al. Gene Editing of the Catfish Gonadotropin-Releasing Hormone Gene and Hormone Therapy to Control the Reproduction in Channel Catfish, Ictalurus punctatus. Biology. 2022; 11(5):649. https://doi.org/10.3390/biology11050649
Chicago/Turabian StyleQin, Guyu, Zhenkui Qin, Cuiyu Lu, Zhi Ye, Ahmed Elaswad, Max Bangs, Hanbo Li, Yiliu Zhang, Yingqi Huang, Huitong Shi, and et al. 2022. "Gene Editing of the Catfish Gonadotropin-Releasing Hormone Gene and Hormone Therapy to Control the Reproduction in Channel Catfish, Ictalurus punctatus" Biology 11, no. 5: 649. https://doi.org/10.3390/biology11050649