
DNA Methylation Levels of the TBX5 Gene Promoter Are Associated with Congenital Septal Defects in Mexican Paediatric Patients

 and
and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Individuals Studied
2.2. DNA Extraction and Bisulfite Treatment
2.3. DNA Methylation
2.4. In Silico Analysis
2.5. Statistical Analysis
3. Results
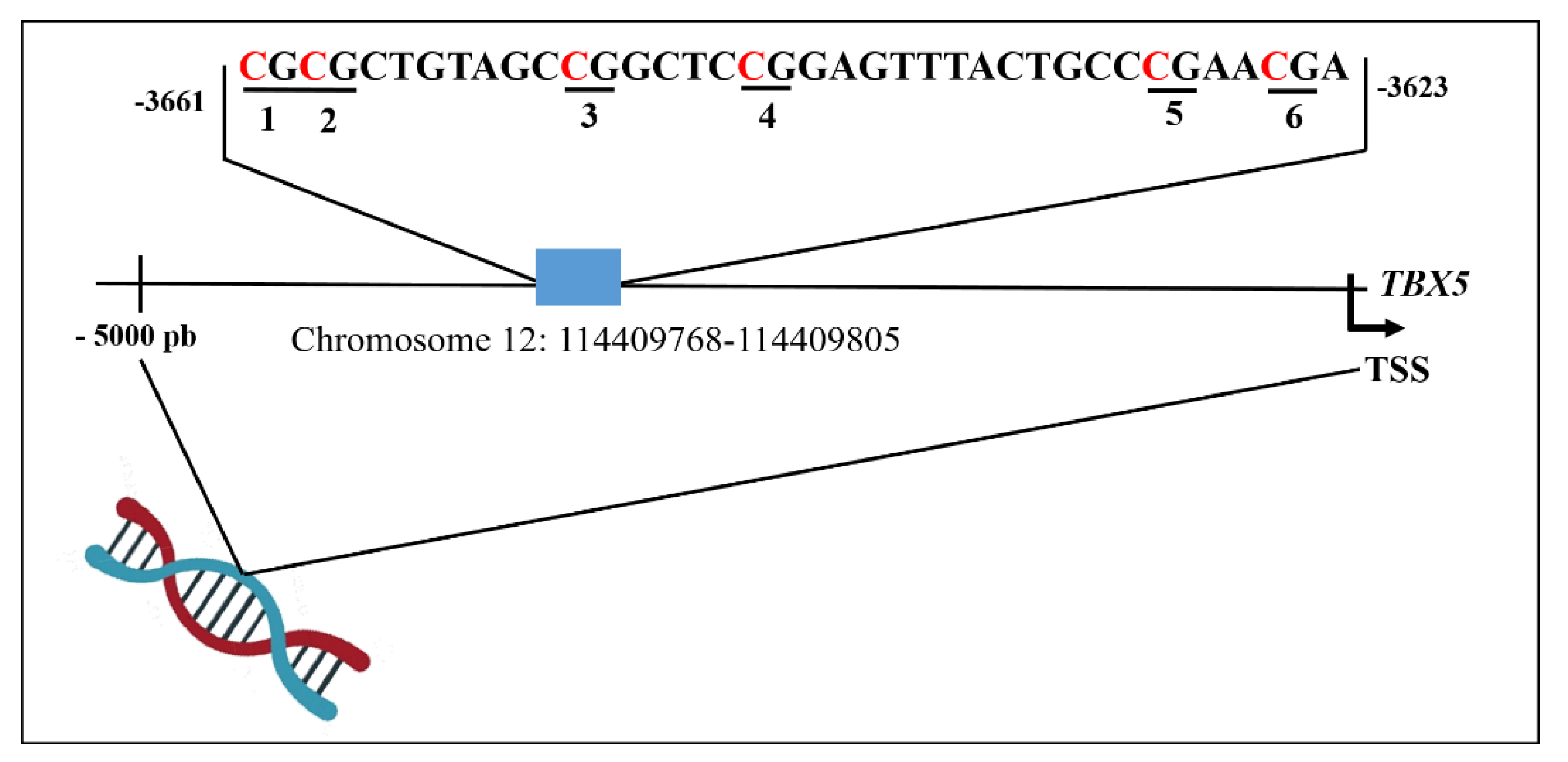
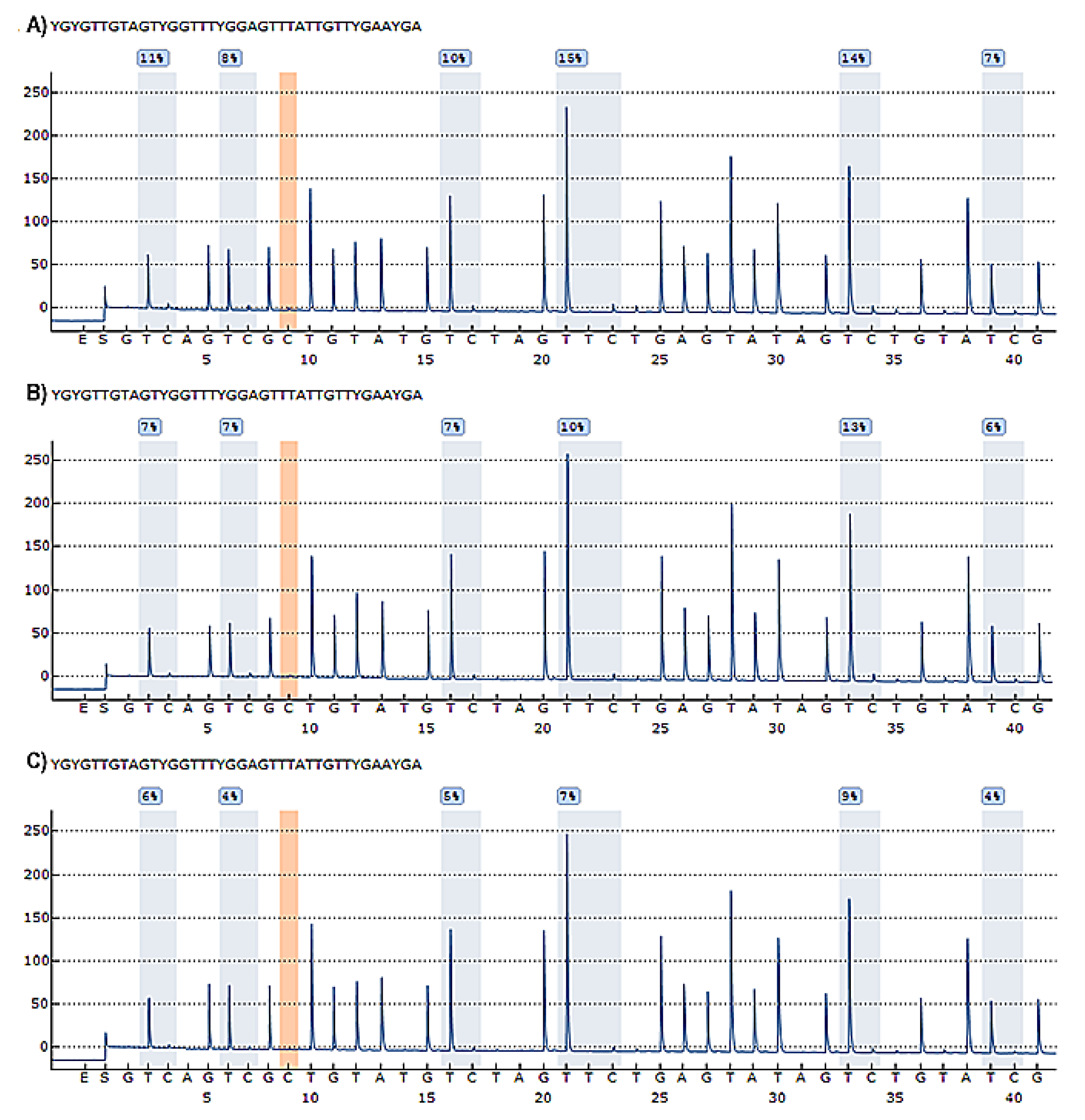
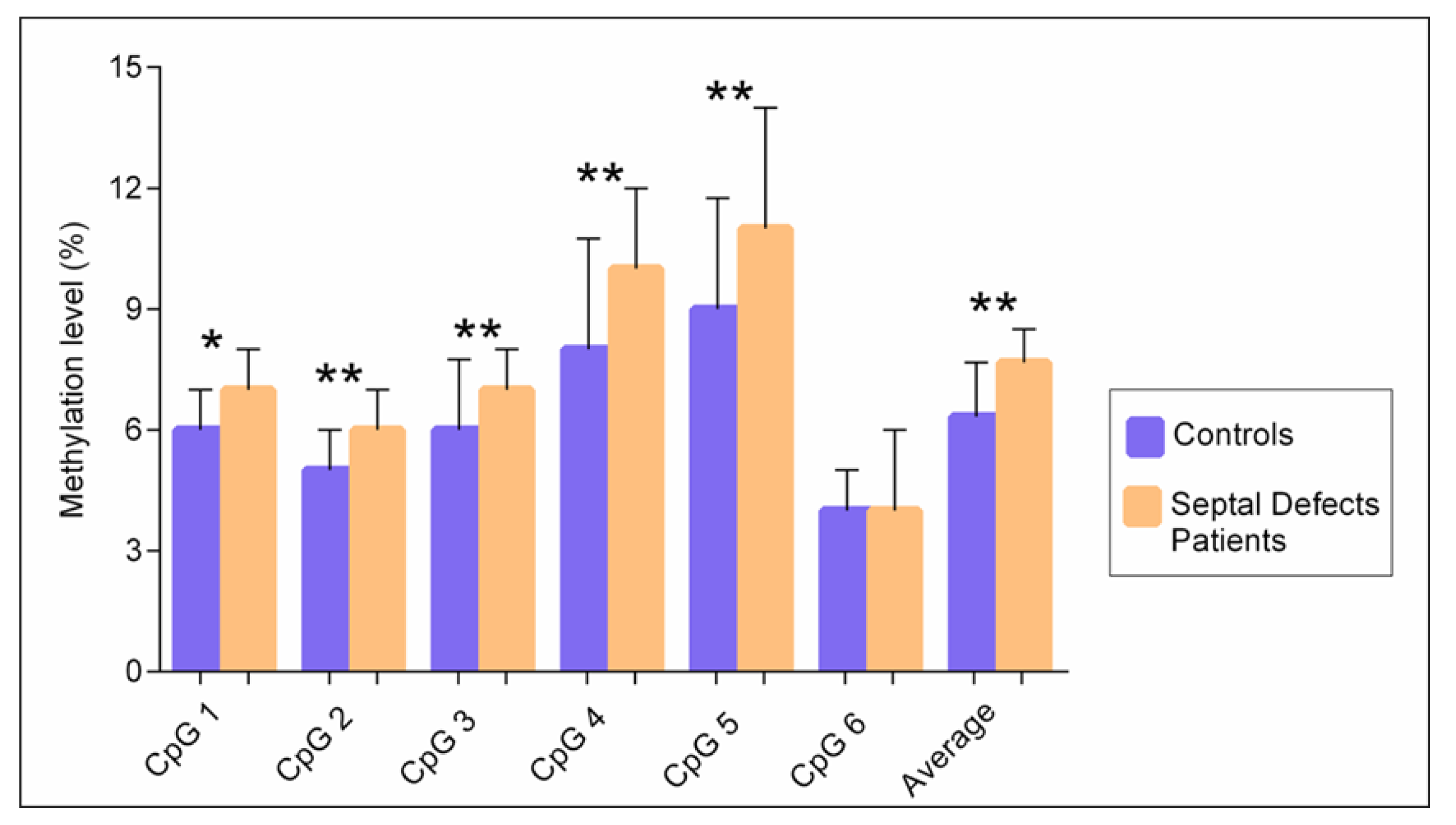
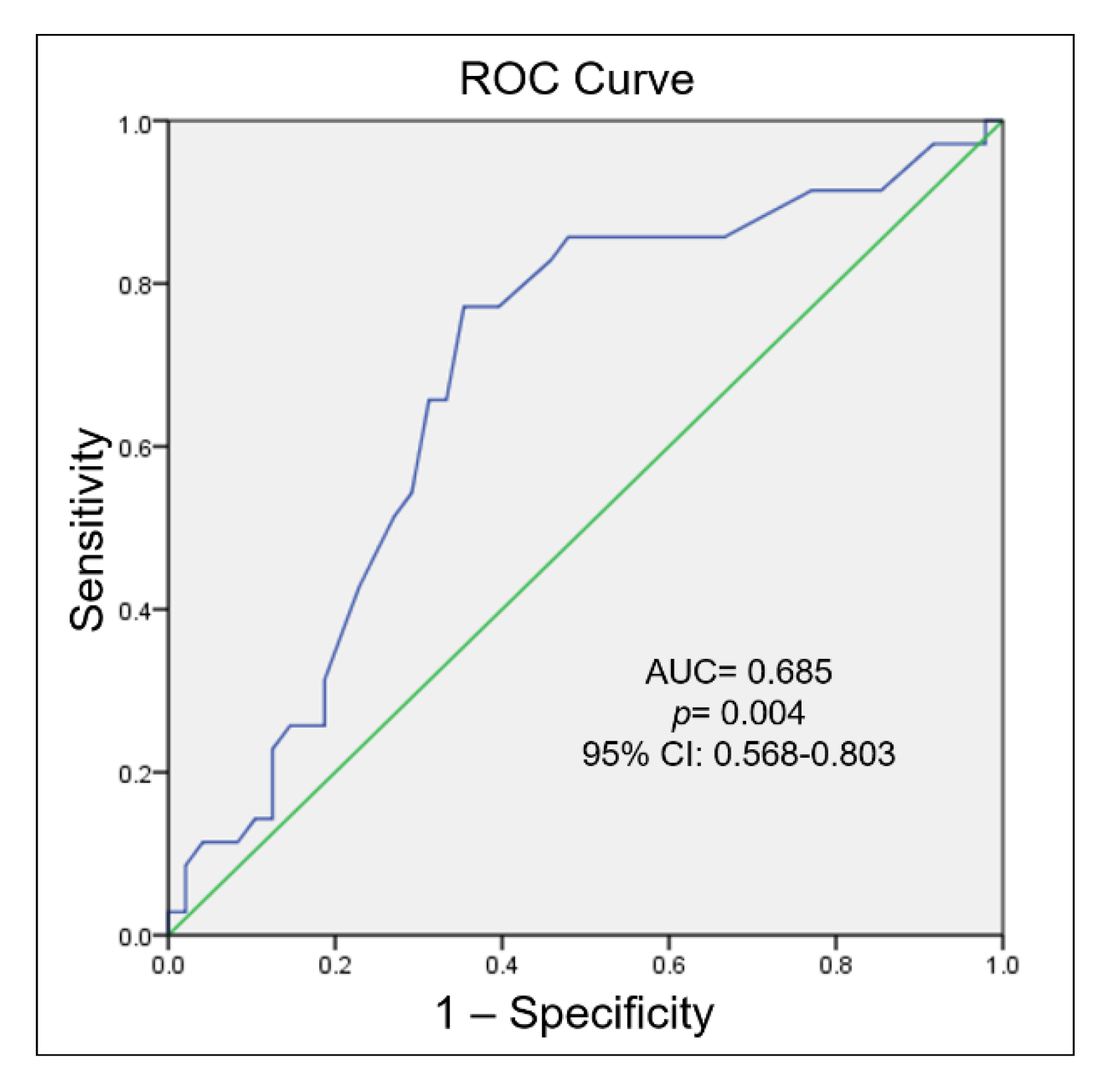
3.1. DNA Methylation of the TBX5 Gene Promoter and Its Association with Septal Defects
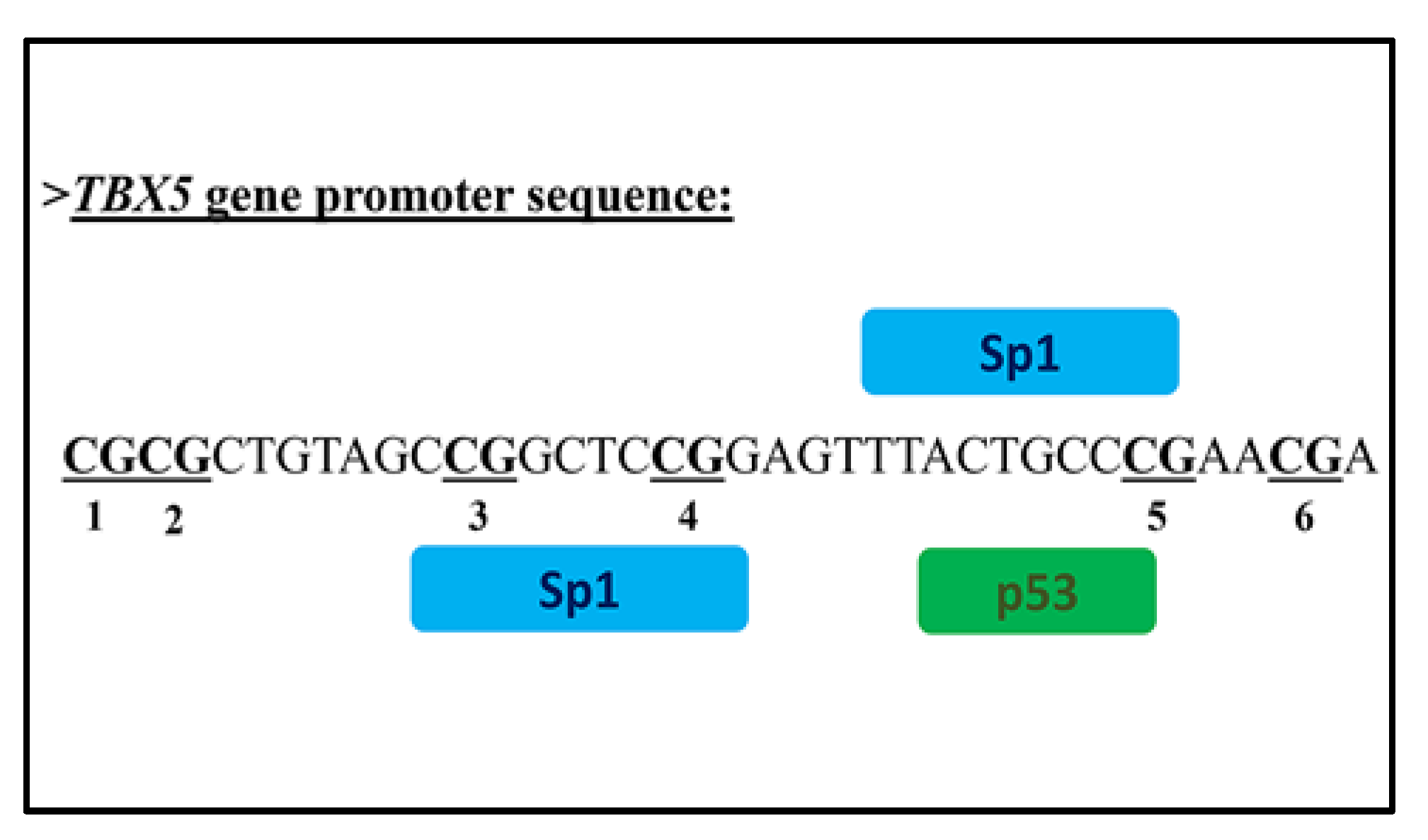
3.2. In Silico Analysis of Transcription Factor Binding Sites to TBX5 Promoter Region
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Yoshida, A.; Morisaki, H.; Nakaji, M.; Kitano, M.; Kim, K.S.; Sagawa, K.; Ishikawa, S.; Satokata, I.; Mitani, Y.; Kato, H.; et al. Genetic mutation analysis in Japanese patients with non-syndromic congenital heart disease. J. Hum. Genet. 2016, 61, 157–162. [Google Scholar] [CrossRef] [PubMed]
- Zhang, H.; Zhao, X.; Wang, C.; Du, R.; Wang, X.; Fu, J.; Sun, Q. A Preliminary Study of the Association between Apolipoprotein E Promoter Methylation and Atherosclerotic Cerebral Infarction. J. Stroke Cerebrovasc. Dis. Off. J. Natl. Stroke Assoc. 2019, 28, 1056–1061. [Google Scholar] [CrossRef]
- Thomford, N.E.; Dzobo, K.; Yao, N.A.; Chimusa, E.; Evans, J.; Okai, E.; Kruszka, P.; Muenke, M.; Awandare, G.; Wonkam, A.; et al. Genomics and Epigenomics of Congenital Heart Defects: Expert Review and Lessons Learned in Africa. Omics A J. Integr. Biol. 2018, 22, 301–321. [Google Scholar] [CrossRef]
- Xie, H.; Zhang, E.; Hong, N.; Fu, Q.; Li, F.; Chen, S.; Yu, Y.; Sun, K. Identification of TBX2 and TBX3 variants in patients with conotruncal heart defects by target sequencing. Hum. Genom. 2018, 12, 44. [Google Scholar] [CrossRef]
- Greulich, F.; Rudat, C.; Kispert, A. Mechanisms of T-box gene function in the developing heart. Cardiovasc. Res. 2011, 91, 212–222. [Google Scholar] [CrossRef] [Green Version]
- Papaioannou, V.E. The T-box gene family: Emerging roles in development, stem cells and cancer. Development 2014, 141, 3819–3833. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Showell, C.; Binder, O.; Conlon, F.L. T-box genes in early embryogenesis. Dev. Dyn. Off. Publ. Am. Assoc. Anat. 2004, 229, 201–218. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yang, X.; Kong, Q.; Li, Z.; Xu, M.; Cai, Z.; Zhao, C. Association between the promoter methylation of the TBX20 gene and tetralogy of fallot. Scand. Cardiovasc. J. SCJ 2018, 52, 287–291. [Google Scholar] [CrossRef]
- Yang, X.F.; Zhang, Y.F.; Zhao, C.F.; Liu, M.M.; Si, J.P.; Fang, Y.F.; Xing, W.W.; Wang, F.L. Relationship between TBX20 gene polymorphism and congenital heart disease. Genet. Mol. Res. GMR 2016, 1–9. [Google Scholar] [CrossRef]
- Boogerd, C.J.; Zhu, X.; Aneas, I.; Sakabe, N.; Zhang, L.; Sobreira, D.R.; Montefiori, L.; Bogomolovas, J.; Joslin, A.C.; Zhou, B.; et al. Tbx20 Is Required in Mid-Gestation Cardiomyocytes and Plays a Central Role in Atrial Development. Circ. Res. 2018, 123, 428–442. [Google Scholar] [CrossRef]
- Steimle, J.D.; Moskowitz, I.P. TBX5: A Key Regulator of Heart Development. Curr. Top. Dev. Biol. 2017, 122, 195–221. [Google Scholar]
- Zhang, K.K.; Xiang, M.; Zhou, L.; Liu, J.; Curry, N.; Heine Suñer, D.; Garcia-Pavia, P.; Zhang, X.; Wang, Q.; Xie, L. Gene network and familial analyses uncover a gene network involving Tbx5/Osr1/Pcsk6 interaction in the second heart field for atrial septation. Hum. Mol. Genet. 2016, 25, 1140–1151. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Waldron, L.; Steimle, J.D.; Greco, T.M.; Gomez, N.C.; Dorr, K.M.; Kweon, J.; Temple, B.; Yang, X.H.; Wilczewski, C.M.; Davis, I.J.; et al. The Cardiac TBX5 Interactome Reveals a Chromatin Remodeling Network Essential for Cardiac Septation. Dev. Cell 2016, 36, 262–275. [Google Scholar] [CrossRef] [Green Version]
- Ma, J.; Chen, S.; Hao, L.; Sheng, W.; Chen, W.; Ma, X.; Zhang, B.; Ma, D.; Huang, G. Hypermethylation-mediated down-regulation of lncRNA TBX5-AS1:2 in Tetralogy of Fallot inhibits cell proliferation by reducing TBX5 expression. J. Cell. Mol. Med. 2020, 24, 6472–6484. [Google Scholar] [CrossRef]
- Moore-Morris, T.; van Vliet, P.P.; Andelfinger, G.; Puceat, M. Role of Epigenetics in Cardiac Development and Congenital Diseases. Physiol. Rev. 2018, 98, 2453–2475. [Google Scholar] [CrossRef] [Green Version]
- Jarrell, D.K.; Lennon, M.L.; Jacot, J.G. Epigenetics and Mechanobiology in Heart Development and Congenital Heart Disease. Diseases 2019, 7, 52. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lewandowski, S.L.; Janardhan, H.P.; Smee, K.M.; Bachman, M.; Sun, Z.; Lazar, M.A.; Trivedi, C.M. Histone deacetylase 3 modulates Tbx5 activity to regulate early cardiogenesis. Hum. Mol. Genet. 2014, 23, 3801–3809. [Google Scholar] [CrossRef] [Green Version]
- Grunert, M.; Dorn, C.; Cui, H.; Dunkel, I.; Schulz, K.; Schoenhals, S.; Sun, W.; Berger, F.; Chen, W.; Sperling, S.R. Comparative DNA methylation and gene expression analysis identifies novel genes for structural congenital heart diseases. Cardiovasc. Res. 2016, 112, 464–477. [Google Scholar] [CrossRef] [PubMed]
- Mittal, A.; Sharma, R.; Prasad, R.; Bahl, A.; Khullar, M. Role of cardiac TBX20 in dilated cardiomyopathy. Mol. Cell. Biochem. 2016, 414, 129–136. [Google Scholar] [CrossRef] [PubMed]
- Yuan, Y.; Gao, Y.; Wang, H.; Ma, X.; Ma, D.; Huang, G. Promoter methylation and expression of the VANGL2 gene in the myocardium of pediatric patients with tetralogy of fallot. Birth Defects Res. Part A Clin. Mol. Teratol. 2014, 100, 973–984. [Google Scholar] [CrossRef]
- Dakkak, W.; Oliver, T.I. Ventricular Septal Defect. In StatPearls; Copyright © 2022; StatPearls Publishing LLC: Treasure Island, FL, USA, 2021. [Google Scholar]
- Menillo, A.M.; Lee, L.S.; Pearson-Shaver, A.L. Atrial Septal Defect. In StatPearls; Copyright © 2022; StatPearls Publishing LLC: Treasure Island, FL, USA, 2021. [Google Scholar]
- Backer, C.L.; Eltayeb, O.; Mongé, M.C.; Mazwi, M.L.; Costello, J.M. Shunt Lesions Part I: Patent Ductus Arteriosus, Atrial Septal Defect, Ventricular Septal Defect, and Atrioventricular Septal Defect. Pediatric Crit. Care Med. A J. Soc. Crit. Care Med. World Fed. Pediatric Intensive Crit. Care Soc. 2016, 17, S302–S309. [Google Scholar] [CrossRef]
- Farré, D.; Roset, R.; Huerta, M.; Adsuara, J.E.; Roselló, L.; Albà, M.M.; Messeguer, X. Identification of patterns in biological sequences at the ALGGEN server: PROMO and MALGEN. Nucleic Acids Res. 2003, 31, 3651–3653. [Google Scholar] [CrossRef] [Green Version]
- Messeguer, X.; Escudero, R.; Farré, D.; Núñez, O.; Martínez, J.; Albà, M.M. PROMO: Detection of known transcription regulatory elements using species-tailored searches. Bioinformatics 2002, 18, 333–334. [Google Scholar] [CrossRef]
- Tsunoda, T.; Takagi, T. Estimating transcription factor bindability on DNA. Bioinformatics 1999, 15, 622–630. [Google Scholar] [CrossRef] [PubMed]
- Sheng, W.; Qian, Y.; Zhang, P.; Wu, Y.; Wang, H.; Ma, X.; Chen, L.; Ma, D.; Huang, G. Association of promoter methylation statuses of congenital heart defect candidate genes with Tetralogy of Fallot. J. Transl. Med. 2014, 12, 31. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, J.; Ma, X.; Wang, H.; Ma, D.; Huang, G. Elevated methylation of the RXRA promoter region may be responsible for its downregulated expression in the myocardium of patients with TOF. Pediatric Res. 2014, 75, 588–594. [Google Scholar] [CrossRef] [Green Version]
- Song, X.; Cao, H.; Hong, L.; Zhang, L.; Li, M.; Shi, J.; Liu, J.; Ma, J.; Cui, L.; Zhang, Y.; et al. Ventricular Myocardial Deformation in Fetuses With Tetralogy of Fallot: A Necessary Field of Investigation. Front. Cardiovasc. Med. 2021, 8, 764676. [Google Scholar] [CrossRef] [PubMed]
- Diaz-Frias, J.; Guillaume, M. Tetralogy of Fallot. In StatPearls; Copyright © 2022; StatPearls Publishing LLC: Treasure Island, FL, USA, 2021. [Google Scholar]
- Green, B.B.; Marsit, C.J. Select Prenatal Environmental Exposures and Subsequent Alterations of Gene-Specific and Repetitive Element DNA Methylation in Fetal Tissues. Curr. Environ. Health Rep. 2015, 2, 126–136. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Nielsen, C.H.; Larsen, A.; Nielsen, A.L. DNA methylation alterations in response to prenatal exposure of maternal cigarette smoking: A persistent epigenetic impact on health from maternal lifestyle? Arch. Toxicol. 2016, 90, 231–245. [Google Scholar] [CrossRef] [PubMed]
- Alvarado-Cruz, I.; Alegría-Torres, J.A.; Montes-Castro, N.; Jiménez-Garza, O.; Quintanilla-Vega, B. Environmental Epigenetic Changes, as Risk Factors for the Development of Diseases in Children: A Systematic Review. Ann. Glob. Health 2018, 84, 212–224. [Google Scholar] [CrossRef] [Green Version]
- Rauschert, S.; Melton, P.E.; Burdge, G.; Craig, J.M.; Godfrey, K.M.; Holbrook, J.D.; Lillycrop, K.; Mori, T.A.; Beilin, L.J.; Oddy, W.H.; et al. Maternal Smoking During Pregnancy Induces Persistent Epigenetic Changes Into Adolescence, Independent of Postnatal Smoke Exposure and Is Associated With Cardiometabolic Risk. Front. Genet. 2019, 10, 770. [Google Scholar] [CrossRef] [PubMed]
- Cao, J.; Wu, Q.; Huang, Y.; Wang, L.; Su, Z.; Ye, H. The role of DNA methylation in syndromic and non-syndromic congenital heart disease. Clin. Epigenetics 2021, 13, 93. [Google Scholar] [CrossRef] [PubMed]
- Men, H.; Cai, H.; Cheng, Q.; Zhou, W.; Wang, X.; Huang, S.; Zheng, Y.; Cai, L. The regulatory roles of p53 in cardiovascular health and disease. Cell. Mol. Life Sci. CMLS 2021, 78, 2001–2018. [Google Scholar] [CrossRef]
- Zhang, M.; Zhang, Y.; Xu, E.; Mohibi, S.; de Anda, D.M.; Jiang, Y.; Zhang, J.; Chen, X. Rbm24, a target of p53, is necessary for proper expression of p53 and heart development. Cell Death Differ. 2018, 25, 1118–1130. [Google Scholar] [CrossRef] [Green Version]
- Lahalle, A.; Lacroix, M.; De Blasio, C.; Cissé, M.Y.; Linares, L.K.; Le Cam, L. The p53 Pathway and Metabolism: The Tree That Hides the Forest. Cancers 2021, 13, 133. [Google Scholar] [CrossRef]
- Van Nostrand, J.L.; Brady, C.A.; Jung, H.; Fuentes, D.R.; Kozak, M.M.; Johnson, T.M.; Lin, C.Y.; Lin, C.J.; Swiderski, D.L.; Vogel, H.; et al. Inappropriate p53 activation during development induces features of CHARGE syndrome. Nature 2014, 514, 228–232. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Gong, J.; Sheng, W.; Ma, D.; Huang, G.; Liu, F. DNA methylation status of TBX20 in patients with tetralogy of Fallot. BMC Med. Genom. 2019, 12, 75. [Google Scholar] [CrossRef] [Green Version]
- Beishline, K.; Azizkhan-Clifford, J. Sp1 and the ‘hallmarks of cancer’. FEBS J. 2015, 282, 224–258. [Google Scholar] [CrossRef]
- Li, J.; Cao, Y.; Wu, Y.; Chen, W.; Yuan, Y.; Ma, X.; Huang, G. The expression profile analysis of NKX2-5 knock-out embryonic mice to explore the pathogenesis of congenital heart disease. J. Cardiol. 2015, 66, 527–531. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Congenital Septal Defects Patients (n = 35) | Controls (n = 48) | p * | |
|---|---|---|---|
| Age (in years) a | 8 (4–13) | 3 (2–6) | 0.005 |
| Sex (%) b | 0.040 | ||
| Females | 17 (48.6) | 34 (70.8) | |
| Males | 18 (51.4) | 14 (29.2) |
| TBX5 Gene | Congenital Septal Defect Patients (n = 35) | Controls (n = 48) | OR (95% CI) | p * |
|---|---|---|---|---|
| Methylation Levels | n (%) | n (%) | ||
| Highest quartile (>75%) | 11 (31.4) | 9 (18.8) | 3.91 (1.02–14.8) | 0.045 |
| Medium quartile (25–75%) | 19 (54.3) | 23 (47.9) | 2.64 (0.8–8.5) | 0.105 |
| Lowest quartile (<25%) | 5 (14.3) | 16 (33.3) | 1.0 (Reference) |
| Environmental Risk Factors | OR (95% CI) | p * |
|---|---|---|
| Drug addiction | 0.75 (0.26–2.15) | 0.593 |
| Diseases during pregnancy | 1.77 (0.68–4.57) | 0.236 |
| Exposure to pollutants | 1.14 (0.47–2.77) | 0.765 |
| Maternal infections | 2.90 (1.01–8.33) | 0.048 |
| Medication consumption | 2.03 (0.84–4.93) | 0.115 |
| Consumption of vitamins | 0.22 (0.02–2.28) | 0.208 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
García-Flores, E.; Rodríguez-Pérez, J.M.; Borgonio-Cuadra, V.M.; Vargas-Alarcón, G.; Calderón-Colmenero, J.; Sandoval, J.P.; García-Montes, J.A.; Espinoza-Gutiérrez, V.M.; Reyes-García, J.G.; Cazarín-Santos, B.G.; et al. DNA Methylation Levels of the TBX5 Gene Promoter Are Associated with Congenital Septal Defects in Mexican Paediatric Patients. Biology 2022, 11, 96. https://doi.org/10.3390/biology11010096
García-Flores E, Rodríguez-Pérez JM, Borgonio-Cuadra VM, Vargas-Alarcón G, Calderón-Colmenero J, Sandoval JP, García-Montes JA, Espinoza-Gutiérrez VM, Reyes-García JG, Cazarín-Santos BG, et al. DNA Methylation Levels of the TBX5 Gene Promoter Are Associated with Congenital Septal Defects in Mexican Paediatric Patients. Biology. 2022; 11(1):96. https://doi.org/10.3390/biology11010096
Chicago/Turabian StyleGarcía-Flores, Esbeidy, José Manuel Rodríguez-Pérez, Verónica Marusa Borgonio-Cuadra, Gilberto Vargas-Alarcón, Juan Calderón-Colmenero, Juan Pablo Sandoval, José Antonio García-Montes, Víctor Manuel Espinoza-Gutiérrez, Juan Gerardo Reyes-García, Benny Giovanni Cazarín-Santos, and et al. 2022. "DNA Methylation Levels of the TBX5 Gene Promoter Are Associated with Congenital Septal Defects in Mexican Paediatric Patients" Biology 11, no. 1: 96. https://doi.org/10.3390/biology11010096