
Detection and Control of Fusarium oxysporum from Soft Rot in Dendrobium officinale by Loop-Mediated Isothermal Amplification Assays


1
Institute of Crop Protection, Guizhou University, Guiyang 550025, China
2
The Provincial Key Laboratory for Agricultural Pest Management in Mountainous Region, Guiyang 550025, China
3
College of Agriculture, Guizhou University, Guiyang 550025, China
*
Author to whom correspondence should be addressed.
Biology 2021, 10(11), 1136; https://doi.org/10.3390/biology10111136
Submission received: 8 October 2021
/
Revised: 2 November 2021
/
Accepted: 3 November 2021
/
Published: 5 November 2021
(This article belongs to the Section Biochemistry and Molecular Biology)
Abstract
:Simple Summary
In this study, we investigated the soft rot disease of Dendrobium officinale in Guizhou Province, and designed a rapid method to detect the pathogen is Fusarium oxysporum in Dendrobium officinale by using specific gene sequences and loop-mediated isothermal amplification technology. This method can accurately detect Fusarium oxysporum on soft rot plants of Dendrobium officinale. Therefore, the results of this study are crucial for the early diagnosis of soft rot on Dendrobium officinale.
Abstract
Soft rot causing Fusarium oxysporum is one of the most destructive diseases of Dendrobium officinale Kimura et Migo in China that reduces D. officinale yield and quality. A key challenge for an integrated management strategy for this disease is the rapid and accurate detection of F. oxysporum on D. officinale. Therefore, a new loop-mediated isothermal amplification (LAMP) assay was developed for this purpose. In this study, the primers were selected and designed using the translation elongation factor-1α (TEF-1α) gene region as the target DNA sequence in order to screen the best system of reaction of LAMP to detect F. oxysporum through optimizing different conditions of the LAMP reaction, including time, temperature, concentrations of MgSO4, and concentrations of inner and outer primers. The optimized system was able to efficiently amplify the target gene at 62 °C for 60 min with 1.2 μM internal primers, 0.4 μM external primers, 7 mM Mg2+, and 5 fg/µL minimum detection concentration of DNA for F. oxysporum. The amplified products could be detected with the naked eye after completion of the reaction with SYBR green I. We were better able to control the effect of soft rot in D. officinale using fungicides following a positive test result. Additionally, the control effect of synergism combinations against soft rot was higher than 75%. Thus, LAMP assays could detect F. oxysporum in infected tissues of D. officinale and soils in field, allowing for early diagnosis of the disease.
1. Introduction
Dendrobium officinale Kimura et Migo is a perennial herb belonging to the Orchidaceae Dendrobium Sw. that is commonly used in traditional Chinese medicine [1,2]. Most studies have shown that D. officinale contains abundant mineral elements, amino acids, and fatty acids, which explains its high medicinal value [3,4]. Modern pharmacological studies have proven that its components have a variety of effects, including antitumor, enhanced immunity, and reduced blood sugar, among others [5,6]. D. officinale is distributed in east Asia, southeast Asia, and Australia, along with a few other countries or regions. It is mainly distributed in the southern subtropical areas of China [7,8]. However, D. officinale requires a very specific growth environment and particular climactic conditions. It also grows slowly so the wild resources are very scarce [9,10]. With increasing market demand for D. officinale, people often choose to imitate the wild cultivation of D. officinale [11,12].
The imitated wild cultivation of D. officinale is often planted under forests and on the shady sides of stones [13,14]. High humidity, poor ventilation, and relative sultriness are conducive to the occurrence of diseases. Nowadays, soft rot caused by F. oxysporum is one of the most harmful diseases in D. officinale production, which can occur throughout the year. In particular, soft rot is one of the diseases that frequently occurs in D. officinale in Guizhou province, which is mostly caused by Fusarium oxysporum. The incidence of soft rot is from 30 to 50% after surveying seven regions of D. officinale in Guizhou, according to many surveys in our lab. The mode of transmission of F. oxysporum is horizontal transmission, which can infect the root or root neck of the plant. The mycelium living in the soil permeates the root through the seed and then infects the stem [15,16]. The pathogens invade the tissue through the large space between the outer skin cells of the plant. The mycelium and spores enter vessels and vascular tissues and expand into all parts of the plant [17,18]. When the temperature and humidity are ideal, F. oxysporum is infected rapidly. Roots, stems, and leaves begin to show symptoms of disease three days after infection [19,20]. Besides the damage inflicted on tubers, F. oxysporum also produces mycotoxins, which are harmful to humans and animals [21,22,23]. Therefore, it is necessary to develop a rapid method for the early diagnosis of soft rot in D. officinale for disease control.
Currently, the traditional pathogen isolation process includes separation, purification, microscope observation, and physiological biochemical determination. Conventional PCR identification requires special equipment and experienced technicians and takes several hours to conduct. Thus, it is not suitable for application beyond the specialist level [24,25]. A more specialized detection method, loop-mediated isothermal amplification (LAMP), is a new detection method that can efficiently amplify nucleic acid. It is widely used in the rapid detection of pathogens [26,27,28]. LAMP assays consist of two pairs of primers for six regionally identified target sequences, and substitute DNA polymerase chains under constant temperature without thermal cycler for active Bst [29,30,31]. The LAMP product can be used for real-time monitoring, adding SYBR Green I, hydroxynaphthol blue (HNB), calcein, or gel electrophoresis [32,33]. Therefore, there is no need for expensive specialist equipment, such as thermal circulators, and LAMP assays are suitable for the detection and identification of pathogens [34]. F. oxysporum is one of the pathogens that causes soft rot in D. officinale, but Fusarium. sp. are a complex group, which is reflected in their rich genetic diversity, high genome variability, and wide range of hosts [35,36]. Therefore, we need accurate and specific molecular markers of soft rot (F. oxysporum) in D. officinale. This study is based on using the TEF-1α gene of F. oxysporum to design a new set of LAMP-specific primers, following which the conditions of the LAMP assays were explored and optimized. We use mycelium and DNA extracted from fungi on pathogenic plant tissues to test the specificity, sensitivity, and stability of this method. This work provides an early and rapid diagnostic method for soft rot (F. oxysporum) in D. officinale for proper and timely disease control.
2. Materials and Methods
2.1. Strains
Fusarium oxysporum strains were isolated from samples of soft rot in D. officinale collected from different areas of Guizhou province, China. F. oxysporum strains were identified by their morphological characteristics, sequence analyses of the rDNA internal transcribed spacer (ITS), and translation elongation factor 1-alpha (TEF-1α) genes using primers ITS4/ITS5 [37] and EF1/EF2 [38]. All strains were preserved at Guizhou University (Table 1).
2.2. Isolation and Purification of Strain
The pathogens were isolated by a conventional tissue isolation method. Symptomatic tips of D. officinale stems and rots were collected from Guizhou province in China during 2019–2020. Samples of approximately 3–4 × 2–3 mm were taken from the margin of necrosis at the stem base of D. officinale, disinfected in 75% ethyl alcohol for 5 s, and then rinsed three times in sterile distilled water. Small pieces of tissue were soaked up in sterilized water with filter paper and plated on potato dextrose agar (PDA) culture at 25 °C for 7 d. The morphological characteristics of isolates, including colony texture and color, size, and conidiophores, were assessed. The purified strains were transferred to slope PDA for preservation.
2.3. DNA Extraction
Mycelia of F. oxysporum were grown at 25 °C for 3–5 d for DNA extraction. Genomic DNA was extracted from mycelia using a fungal gDNA isolation kit (Hangzhou Biomedical Technology Co., Ltd., Hangzhou, Zhejianng, China). The concentration of DNA was determined by spectrophotometry at 260 nm and the purity of the nucleic acid was evaluated by the ratio of absorbance at 260 to 280 nm. Genomic DNA was diluted to a concentration of 50 ng/μL and then stored at −20 °C until use.
2.4. Primers Design
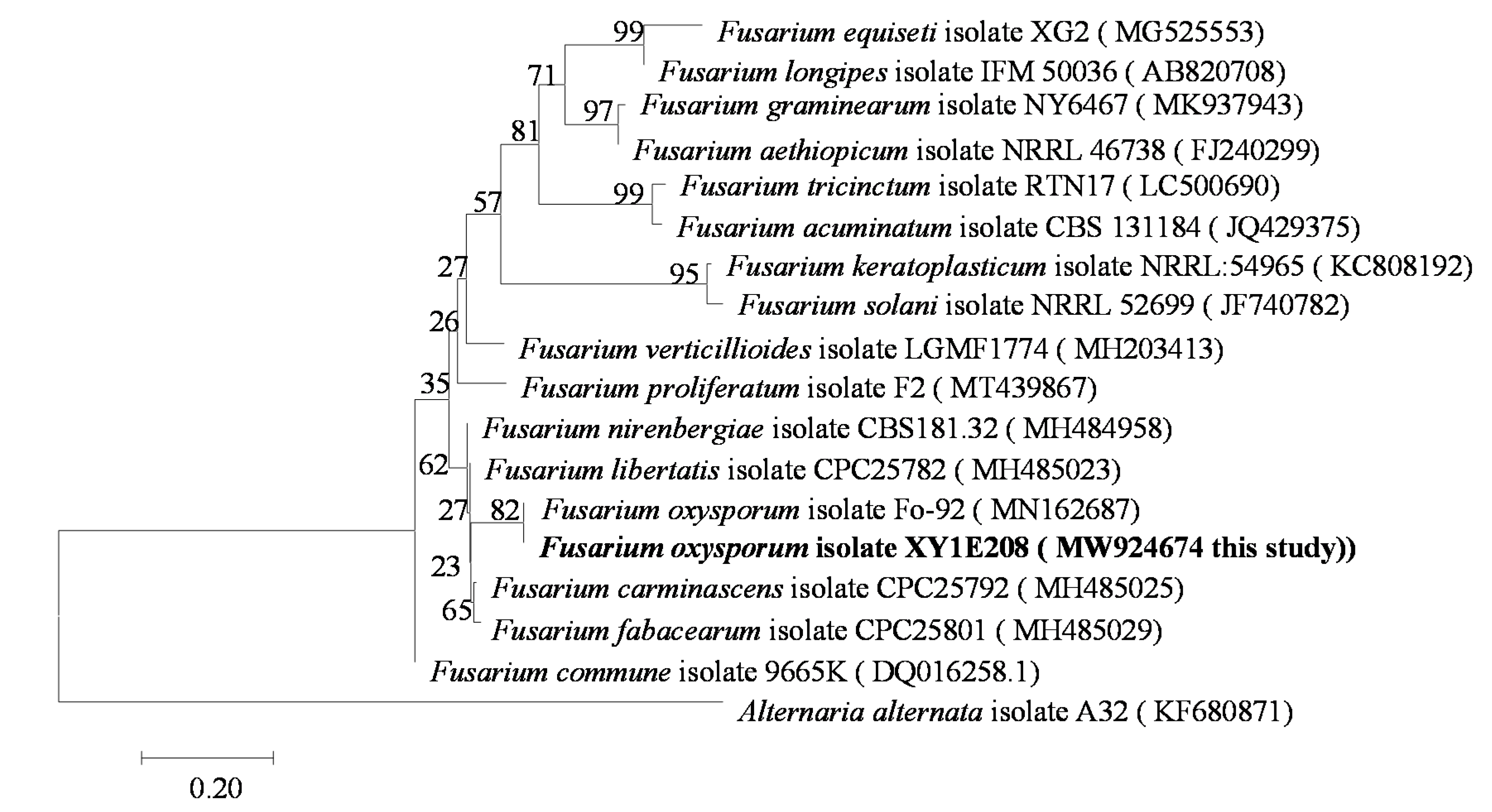
We used a molecular evolutionary genetics analysis (MEGA7.0) program to perform a phylogenetic analysis based on the TEF-1α gene sequence. The basic local alignment search tool (BLAST-N) software (http://blast.ncbi.nlm.nih.gov, 7 October 2021) was used to compare the sequences with other sequences of the genus Fusarium from the national biotechnology information center (NCBI). The alignment analysis was carried out using MEGA7.0 software. The phylogenetic tree was constructed using a data analysis model and the adjacency method (neighbor-joining). The bootstrap support value (bootstrap) was 1000. In the phylogenetic tree, the strain XY1E208 was in the same branch as F. oxysporum, being isolated from other Fusarium species (Figure 1). Therefore, the gene sequence of TEF-1α could be used to distinguish different species of F. oxysporum. We determined the base sequence of TEF-1a using a conserved region of F. oxysporum and used ClustalX software to compare and analyze the base sequence of the TEF-1α gene between different species of Fusarium and other pathogens.
LAMP primers were designed according to partial TEF-1α gene sequences as species-specific primers with primer explorer V5 software (http://primerexplorer.jp/e/, 13 May 2021). Six LAMP primers are shown in Figure 2 and listed in Table 2, including two external primers (F3 and B3), two internal primers (FIP and BIP), and two loop primers (F-loop and B-loop). The primers were synthesized by Sangon Biological Engineering Co., LTD (Shanghai, China) and then repackaged with ddH2O after being dissolved and stored at 4 °C.
2.5. Optimization of LAMP Reaction Conditions
LAMP reactions were conducted according to Notomi’s protocol [29]. The LAMP reactions were accomplished in 200 µL microtubes containing 2.5 µL of 10 × LAMP Master Mix (New England biolabs (Beijing) LTD., Beijing, China.), 6 mM of MgSO4, 1.4 mM of dNTP Mix, 0.2 µM of outer primers (F3 and B3), 1.6 µM of internal primers (FIP and BIP), 0.4 µM of loop primers, 320 U/mL Bst DNA polymerase, 1.0 µL DNA template, 3 µL of 1000 × diluted SYBR Green I (Sangon Biotech Co., Ltd., Shanghai, China), and autoclaved distilled water was used to adjust the volume to 25 µL in the Loopamp RealTime Turbidimeter LA-320C (Eiken Chemical Co., Ltd., Tokyo, Japan). The LAMP reaction conditions were optimized in terms of the concentration of Mg2+, outer primers, and internal primers, temperature, and time of Bst 2.0 DNA polymerase.
For the optimization of reagents, a range of reaction temperatures (55, 58, 60, 62, 64, 65, 66, 68, and 70 °C), a range of reaction times (15, 30, 45, 60, 75, and 90 min), a range of Mg2+ concentrations (2, 3, 4, 5, 6, 7, 8, 9, and 10 mM), a range of inner primer concentrations (0.4, 0.8, 1.2, 1.6, and 2 µM), and a range of outer primer concentrations (0.2, 0.4, 0.6, 0.8, and 1 µM) were evaluated under otherwise identical conditions. The LAMP reactions were performed in 200 µL microfuge tubes incubated in a water bath for 60 min at 65 °C. The reactions were halted by immersion at 80 °C for 5 min, at which point reaction products were detected by fluorescent LAMP and electrophoresis, following which LAMP reactions were completed. After the LAMP reactions took place, the naked eye color changes were directly observed after adding 3 μL 1000 × SYBR Green I dye and centrifuging at the end of the tube.
2.6. LAMP Assays Specificity
Eighteen fungus strains were used for the LAMP specificity test, including strains belonging to the Fusarium genus and other non-Fusarium species (Table 1). The DNA of the tested strains was extracted using a fungal genome DNA isolation kit (Hangzhou Bio-medical technology Co., Ltd. Hangzhou, Zhejiang, China). LAMP reactions were implemented with extracted DNA under optimized conditions. Each experiment was repeated three times.
2.7. Detection of F. axysporum by LAMP and Conventional PCR
To evaluate the sensitivity of the LAMP assays, the DNA of Fusarium wilt in D. officinale was extracted and used as a control for LAMP amplification for specific detection. The DNA of F. oxysporum was diluted at a range of concentrations (10–1, 10–2, 10–3, 10–4, 10–5, 10–6, 10–7, 10–8, and 10–9 times) as a template for the experiment. The conventional polymerase chain reaction (PCR) mixture contains 1 µL of DNA, 1 µL of outer primers (F3/B3), 10 µL of 2 × Taq Mix DNA polymerase, 7 µL of ddH2O, and 20 µL of distilled autoclaved water. The reaction mixtures were incubated in a BIO-RAD TP100 PCR machine. The program was 94 °C for 5 min; 35 cycles of 94 °C for 30 s, 56 °C for 30 s, 72 °C for 1 min; 72 °C for 10 min.
After a color reaction with fluorescent dye, the 5 μL amplification products were detected using electrophoresis with 1% agarose gel and the molecular weights of bands were determined using DNA molecular weight standards. The products were reacted for 25 min under conditions of 150V and 400A. The gels were photographed under ultraviolet light in a gel imaging system after electrophoresis. The existence of ladder bands under ultraviolet light showed that there was soft rot (F. oxysporum) in the D. officinale samples. However, no ladder bands indicated a negative result.
2.8. The Feasibility Detection of LAMP Assays
To evaluate the feasibility of diagnosis by LAMP assays in the field, healthy stems were inoculated with F. oxysporum to simulate field-infected stems and then DNA was extracted from the stems of D. officinale and cultivated soil using a fungal genome DNA isolation kit (Hangzhou Biomedical Technology Co., Ltd. Hangzhou, Zhejiang, China) with apparent disease symptoms. The LAMP assays were performed as described above, and non-symptomatic stems of D. officinale and sterilized soil were used as controls. In addition, suspected soft rot of D. officinale in six areas of Guizhou province were collected, and LAMP assays could be used to detect pathogens with observations of color changes after adding SYBR Green I.
2.9. Prevention and Control of F. oxysporum from Soft Rot in D. officinale by LAMP Assays
Control effects of fungicide combination against F. oxysporum were studied in a growth chamber. The pots for planting D. officinale were 6.5 × 8 cm (φ × h) and filled with nutrient soils, which were inoculated with F. oxysporum following an acupuncture method. The samples of D. officinale were detected by LAMP assays before controlling soft rot with a combination of fungicides.
The fungicide combinations were diluted with 0.5% Tween-20 water to spray onto D. officinale. Additionally, 10 mL of fungicide (pyraclostrobin, picoxystrobin, osthole, and physcion) liquid was sprayed onto D. officinale after positive detection by LAMP assay. A total of 10 mL of sterile water was set as the control group. The control effects of fungicide combinations were observed and recorded after spraying for 14 d. The control effect calculation formula is DI = (∑ number of diseased leaves (stalks) × disease grade index) / (the total number of leaves (stalks) × maximum disease level) × 100. An increased value of the disease index = the DI of leaves × 0.5 + the DI of stalks × 0.5; control effect (%) = (increased value of the disease index in the processing areas—increased value of the disease index in the control areas) / increased value of the disease index in the processing areas × 100.
3. Results
3.1. Identification of Pathogens
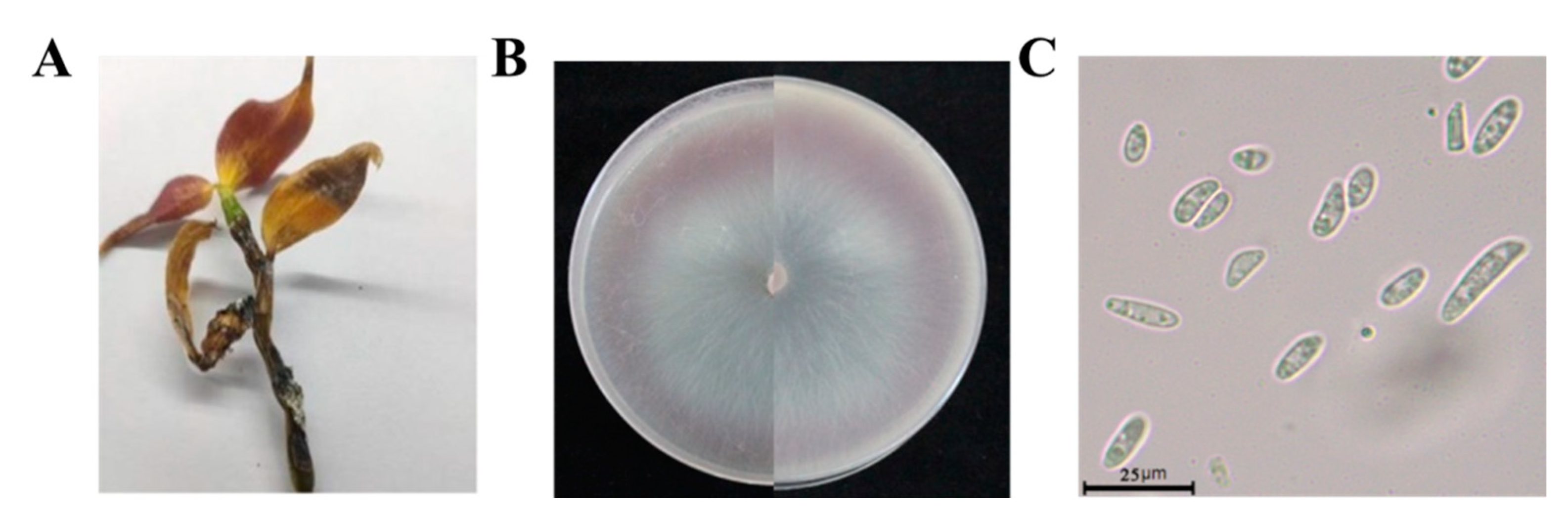
The disease mainly harmed stems of D. officinale. In the early stages of this disease, waterlogged disease spots appeared in plant stems. With the development of the disease, damaged areas expanded, leaves became yellow, stems gradually rotted from the bottom up, and finally whole plants wilted (Figure 3A). We collected samples of D. officinale from seven areas of the Guizhou province, isolating and purifying a total of 66 strains, which represented XY1E208 (23 strains of F. oxysporum) (Figure S1). The colonies of strain XY1E208 were circular on PDA medium with hyphae stripes. The aerial hyphae were white, flocculent, and villous. The early colonies were white, and then they changed to a pale purple pigment in the middle of the colonies (Figure 3B). The conidia of F. oxysporum were hyaline, ellipsoidal, unicellular, 1–2 intervals, and ranged from 4.97 to 25.58 × 1.36 to 4.33 µm in size (Figure 3C).
3.2. Optimization of LAMP Assays
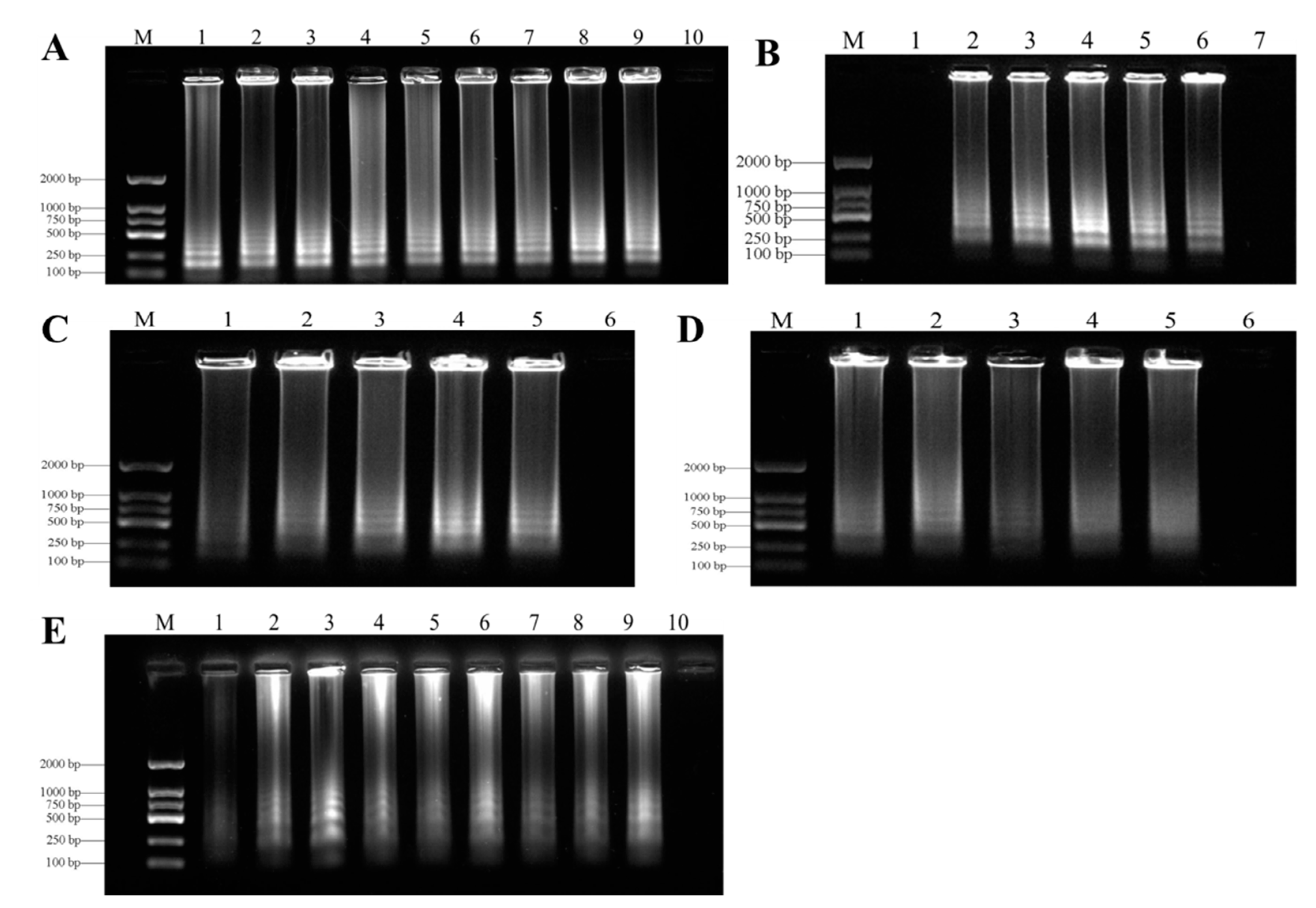
In order to find the best LAMP reaction conditions, we carried out many LAMP reactions under different temperatures (55–70 °C), times (15–90 min), concentrations of Mg2+ (2–10 mM), inner primers (04–2 µM), and outer primers (0.2–1 µM). The results of SYBR Green I staining showed that the best temperature for LAMP detection of genomic DNA in F. oxysporum was 62 °C (Figure 4A). After testing at several reaction times, 60 min was optimal for the LAMP assays for F. oxysporum (Figure 4B). The agarose gel of the ladder bands showed that the reaction had high efficiency with 1.2 µM of internal primers (Figure 4C), 0.4 µM of external primers (Figure 4D), and 7 mM Mg2+ (Figure 4E). All LAMP reactions were carried out in a Loopamp RealTime Turbidimeter LA-320C.
3.3. Specificity of LAMP Assays
LAMP specificity was determined with genome DNA from F. oxysporum and other pathogens through direct visual observation after adding SYBR Green I stain (Table 1). After the reaction, positive samples of color turned green but the color of negative samples remained orange. LAMP primers of F. oxysporum based on TEF-1α amplification yielded positive reactions when testing for DNA, while the DNA from other pathogens were used as negative controls (Figure 5 and Figure S2).
3.4. Sensitivities of LAMP and Conventional PCR Assays
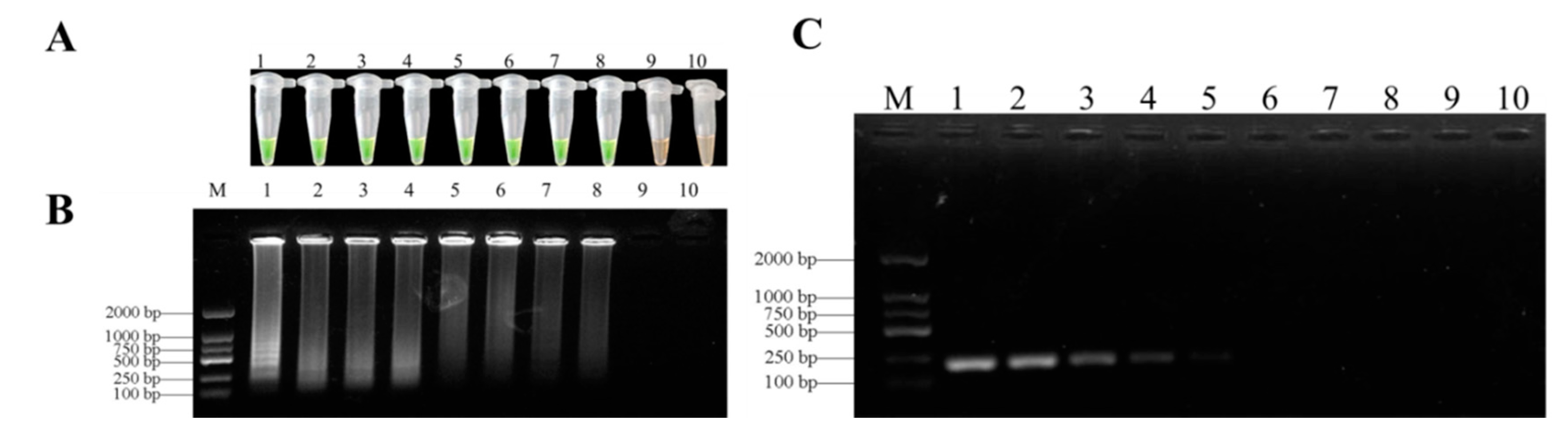
The limits of the LAMP assays were evaluated with a series of concentrations (10–1–10–9 ng/μL) of F. oxysporum DNA template based on partial TEF-1α gene sequences under optimized reaction conditions. The limit of LAMP assays was 5 fg/μL for the genomic DNA of F. oxysporum in D. officinale (Figure 6A,B). However, the sensitivity of conventional PCR was every 25 μL system of F. oxysporum DNA purified by 5 pg/µL (Figure 6C). Therefore, the LAMP assays were more sensitive than conventional PCR assays.
3.5. The Feasibility Detection of LAMP Assays
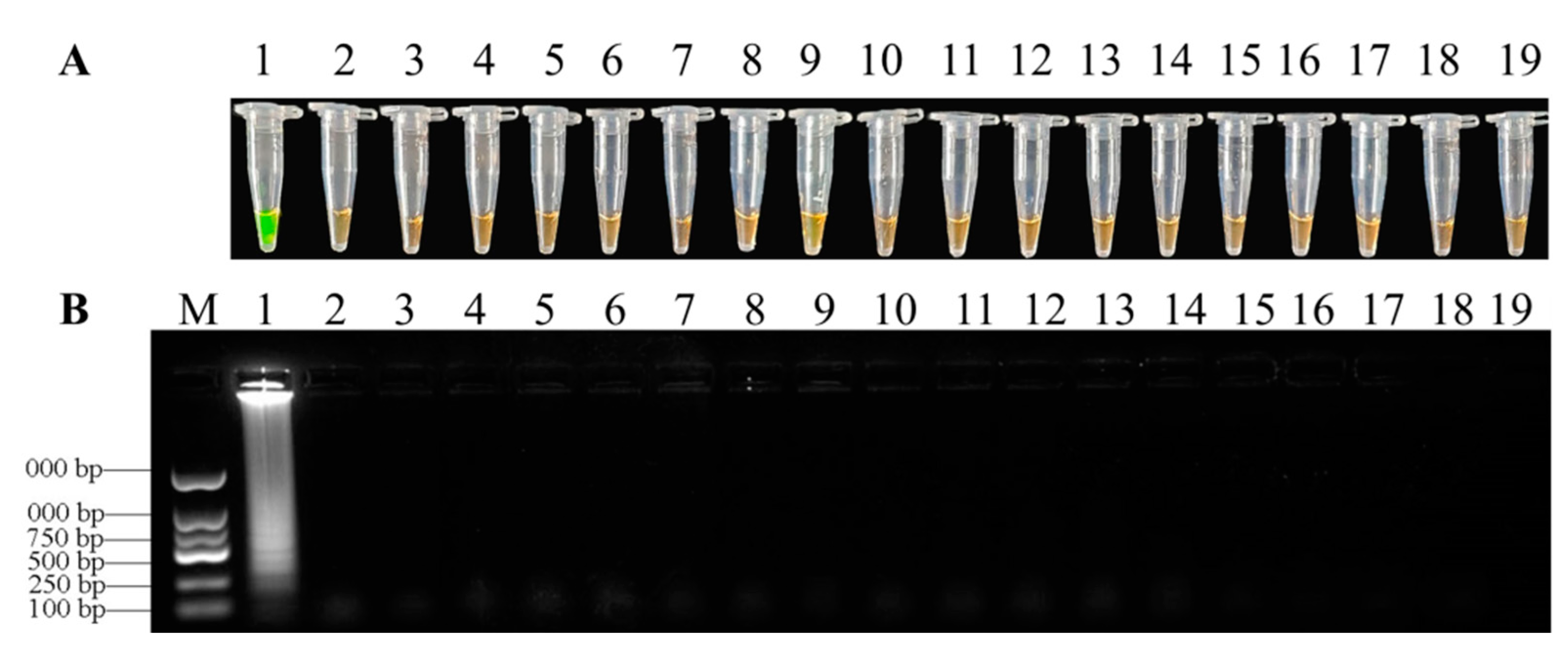
All disease samples and inoculated samples of D. officinale were detected by LAMP assays. Our search results showed that LAMP products of positive reactions turned green after mixing with SYBR Green I while products of samples without infection remained orange (Figure 7). After observing significant symptoms, eight samples were detected to be positive of nine samples from infected tissues by LAMP assays (Figure 7). Seven samples were also positive of nine soil samples (Figure 7B). In addition, six suspected samples of soft rot in D. officinale collected from the Guizhou province in the field were all positive when using LAMP assays (Figure 7C). As a result, the LAMP assays could be used for detecting directly diseased samples of F. oxysporum in D. officinale, and further used for rapid diagnosis of the disease in the field.
3.6. Prevention and Control of Soft Rot in D. officinale by LAMP Assays
We used 21 pots of D. officinale plants artificially planted with an average plant height of 15 cm in this pot experiment. After pathogenic back grafting with F. oxysporum, 12 samples were randomly selected for LAMP testing before disease symptoms appeared. Three samples showed positive results among 12 random samples (Figure 8A). However, 10 samples showed positive results among 12 random samples after disease symptoms appeared (Figure 8B). Then, we immediately sprayed our samples with fungicides in a synergistic combination to control the soft rot in D. officinale plants after most samples tested positive via LAMP assays. The control effects of fungicide synergism combinations (pyraclostrobin and picoxystrobin, osthole and physcion) against soft rot were higher than 75.00% after spraying for 14 days, with control effects being 82.39 and 76.74%, respectively (Table 3). Thus, the LAMP assays can be used for detecting directly diseased samples of F. oxysporum in D. officinale.
4. Discussion
The soft rot in D. officinale caused by F. oxysporum is one of the main diseases in the productive areas of China. This pathogen harms plants’ roots, causing vascular bundle disease and plant death, which can occur along the entire growth period of D. officinale, resulting in great losses to production [39,40,41,42]. The seriously damaged crops reported at home and abroad are eggplant, sugarcane, blackberry, cotton, etc. [43,44,45,46]. At present, chemical control is also the most common method to control plant diseases. However, the effect is often not ideal due to F. oxysporum being a soil-borne disease [47,48,49]. Thus, it is essential to develop an early and rapid diagnostic method for soft rot (F. oxysporum) in D. officinale for proper and timely disease control. In this study, when F. oxysporum primers showed positive reactions in LAMP detection assays, it could predict occurrences of the disease, which is sufficient evidence to guide further control of soft rot in D. officinale. TEF-1α is a highly conserved and ubiquitous protein, which has been widely used to study intra- and inter-species variation and phylogeny in the Fusarium genus [50,51]. Therefore, the TEF-1α region is suitable as a target for the design of LAMP primers. The results of this study also show that the designed primers were highly specific to F. oxysporum, thus LAMP detection could correctly distinguish F. oxysporum from a variety of Fusarium spp.
Our study showed that LAMP assays could correctly detect F. oxysporum. Compared with a conventional PCR assay, the LAMP assays were fast and simple because LAMP reactions are carried out under constant temperature conditions without a thermal circulator. The results of the LAMP assays can be visualized after adding SYBR Green I or Calcein without gel electrophoresis [52,53]. In our study, we used SYBR Green I dye, which showed a clear color change from orange to green to indicate a positive reaction. We also rapidly developed LAMP assays to successfully identify F. oxysporum in D. officinale with a detection limit of 5 fg/µL (Figure 5A), which was significantly higher than previous studies and more sensitive than conventional PCR tests [54,55]. The difference in detection limits by LAMP assays may be due to different sequences as targets in reactions [56]. Therefore, the optimization of a LAMP detection system is very important.
The LAMP method can detect plant pathogens based on the amplification of target DNA sequences [57,58]. We detected various specimens by extracting DNA from different types of samples, including hyphae, infected tissue of D. officinale and soil, and suspected samples in the field. Our search results indicated that LAMP assays could be used to directly detect F. oxysporum in diseased samples. These results were consistent with previous studies using LAMP assays [59,60]. The LAMP assays could detect the samples of F. oxysporum from plant tissues and soil, confirming that this method could be used to diagnose soft rot of F. oxysporum in D. officinale.
Previous studies have reported that external spores have been dispersed by wind, water, people, equipment, and the movement of soil particles as they contain the fungus [61]. Therefore, it is important to prevent and control Fusarium wilt, which is maintained in environmental conditions. In this study, we have established a method to rapidly detect F. oxysporum that also provides early diagnosis of various diseases caused by F. oxysporum. Combined with results of detection by LAMP assays, we may find more accurate and effective pesticides in order to rapidly bring infection under control.
5. Conclusion
In this study, a LAMP method for the detection of F. oxysporum was established. The LAMP assays of F. oxysporum were reacted for 60 min at 62 °C with 1.2 µM internal primers, 0.4 µM external primers, and 7 mM Mg2+, while the PCR method would usually have taken 2–6 h. Based on the TEF-1α gene sequence, designed primers were highly specific for F. oxysporum, with a detection limit of 5 fg/µL through visual inspection after SYBR Green I staining, which was significantly more sensitive than a conventional PCR test (5 pg/µL). Thus, LAMP assays can be used as an effective tool for the early diagnosis of soft rot (F. oxysporum) in D. officinale in the field for proper and timely disease control.
Supplementary Materials
The following are available online at https://www.mdpi.com/article/10.3390/biology10111136/s1: Figure S1: Phylogenetic tree for 23 strains of F. oxysporum based on internal transcribed spacer (ITS) gene sequences; Figure S2: Sensitivity of LAMP and conventional PCR for detection of genomic DNA in F. oxysporum. (A) Detection by LAMP with SYBR Green I staining; (B) LAMP products analyzed by agarose gel electrophoresis. 1, XY1E208; 2, XY1; 3, XY1A; 4, XY1C; 5, XY1K; 6, XY3; 7, SJ-3; 8, SJ-4; 9,SS-YP-1-2; 10, TG-3-1; 11, TG-3-2; 12, TG-4-1; 13, WSS-2; 14, WSS-7.;15, AL2-2; 16, AL2-3; 17, AL3-1; 18, AL3-3; 19, AL3-4; 20, AL5; 21, AL6-1; 22, AL6-2; 23, AL6-3; 24, double-distilled water as negative control.
Author Contributions
Conceptualization, R.L.; methodology and formal analysis, C.X.; funding acquisition, R.L.; writing—review and editing, C.X. and R.L.; visualization, R.L. All authors have read and agreed to the published version of the manuscript.
Funding
This work was supported by the National Natural Science Foundation of China (No. 32160657, 31701816), agricultural research projects of the science and technology department of the Guizhou province (No. [2019]3001), and the cultivation project of Guizhou university (No. [2019]43).
Institutional Review Board Statement
Not applicable.
Informed Consent Statement
Not applicable.
Data Availability Statement
Sequencing data are available from https://www.ncbi.nlm.nih.gov/nuccore/MW924674.
Acknowledgments
The authors thank Li, R.Y. for her help with the statistical analyses and the methodology.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Ge, D.; Li, X.; Ding, X.; Qian, L. Genetic diversity across natural populations of Dendrobium officinale, the endangered medicinal herb endemic to China, revealed by ISSR and RAPD markers. Russ. J. Genet. 2009, 45, 327–334. [Google Scholar] [CrossRef]
- Jin, Z.; Li, D.; Liu, T.; Yu, F.; Liu, Z. Cultural endophytic fungi associated with Dendrobium officinale: Identification, diversity estimation and their antimicrobial potential. Curr. Sci. 2017, 112, 1690–1697. [Google Scholar] [CrossRef]
- Luo, Q.L.; Tang, Z.H.; Zhang, X.F.; Zhong, Y.H.; Yao, S.Z.; Wang, L.S.; Lin, C.W.; Luo, X. Chemical properties and antioxidant activity of a water-soluble polysaccharide from Dendrobium officinale. Int. J. Biol. Macromol. 2016, 89, 219–227. [Google Scholar] [CrossRef] [PubMed]
- Chen, W.H.; Wu, J.J.; Li, X.F.; Lu, J.M.; Wu, W.; Sun, Y.Q.; Zhu, B.; Qin, L.P. Isolation, structural properties, bioactivities of polysaccharides from Dendrobium officinale Kimura et. Migo: A review. Int. J. Biol. Macromol. 2021, 184, 1000–1013. [Google Scholar] [CrossRef]
- Liu, X.F.; Zhu, J.; Ge, S.Y.; Xia, L.J.; Ren, F.Z. Orally Administered Dendrobium officinale and its Polysaccharides Enhance Immune Functions in BALB/c Mice. Nat. Prod. Commun. 2011, 6, 867–870. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, J.J.; Liu, Z.P.; Zhang, X.F.; Si, J.P. Effects of Various Processing Methods on the Metabolic Profile and Antioxidant Activity of Dendrobium catenatum Lindley Leaves. Metabolites 2021, 11, 351. [Google Scholar] [CrossRef]
- Li, G.; Li, R.Y.; Gao, W.W. Occurrence situation and control strategy of Dendrobium diseases in large-scale farming system. China J. Chin. Mater. Med. 2013, 38, 485. [Google Scholar]
- Guo, M.; Li, B.; Wang, R.; Liu, P.; Chen, Q. Occurrence of dieback disease caused by Fusarium equiseti on Dendrobium officinale in China. Crop. Prot. 2020, 137, 105209. [Google Scholar] [CrossRef]
- Vellupillai, S.; Jin, G. Histological and protein changes during early stages of seed germination in the orchid, Dendrobium crumenatum. J. Hortic. Sci. 1997, 72, 941–948. [Google Scholar] [CrossRef]
- Ng, T.B.; Liu, J.Y.; Wong, J.H.; Ye, X.J.; Wing Sze, S.C.; Tong, Y.; Zhang, K.Y. Review of research on Dendrobium, a prized folk medicine. Appl. Microbiol. Biotechnol. 2012, 93, 1795–1803. [Google Scholar] [CrossRef]
- Lin, X.; Zhou, Y.; Zhong, J.; Zhu, J.; Chen, X.; Lin, Y.; Huang, M.; Yang, L.; Zhang, Z. Imitated Ecological Cultivation Technology of Dendrobium offcinale: A Case Study of Longshitou Village, Longmen County. Plant. Dis. Pests 2021, 12, 24–26. [Google Scholar] [CrossRef]
- Chen, L.; Zhang, H.; Liang, D.; Peng, H.; Huang, C.; Zhou, Z.; Liao, Y.; Lin, D. Wild cultivation technology of Dendrobium officinale in natural secondary forest in northern Guangdong Province. South. China For. Sci. 2018, 46, 48–50. [Google Scholar] [CrossRef]
- Tang, L.M.; Chen, Y.F.; Dai, Q.; Li, D.Y.; Jiang, L.; Deng, Y.H. The Standardized Technology Demonstration of Cultivating Dendrobium officinale Kimura et Migo under Forest. Anhui Agric. Sci. Bull. 2015, 21, 45–53. [Google Scholar] [CrossRef]
- Guan, Y.J.; Li, L.R.; Liu, S.D. Effects of Meteorological Conditions on the Growth of Dendrobium candidum under Forest. Sci. Technol. Eng. 2018, 18, 200–203. [Google Scholar]
- Oren, L.; Ezrati, S.; Cohen, D.; Sharon, A. Early events in the Fusarium verticillioides-maize interaction characterized by using a green fluorescent protein-expressing transgenic isolate. Appl. Environ. Microbiol. 2003, 69, 1695–1701. [Google Scholar] [CrossRef] [Green Version]
- Olivain, C.; Humbert, C.; Nahalkova, J.; Fatehi, J.; L’Haridon, F.; Alabouvette, C. Colonization of tomato root by pathogenic and nonpathogenic Fusarium oxysporum strains inoculated together and separately into the soil. Appl. Environ. Microbiol. 2006, 72, 1523–1531. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagopodi Anastasia, L.; Ram Arthur, F.J.; Lamers, G.E.M.; Punt, P.J.; Van den Hondel, C.A.M.J.J. Novel aspects of tomato root colonization and infection by Fusarium oxysporum f. sp. radicis-lycopersici revealed by confocal laser scanning microscopic analysis using the green fluorescent protein as a marker. Mol. Plant.-Microbe Interact. 2002, 15, 172–179. [Google Scholar] [CrossRef] [Green Version]
- Eynck, C.; Koopmann, B.; Grunewaldt-Stoecker, G.; Karlovsky, P.; Tiedemann, A.V. Differential interactions of Verticillium longisporum and V. dahliae with Brassica napus detected with molecular and histological techniques. Eur. J. Plant. Pathol. 2007, 118, 259–274. [Google Scholar] [CrossRef]
- Zhou, L.; Hu, Q.; Johansson, A.; Dixelius, C. Verticillium longisporum and V. dahliae: Infection and disease in Brassica napus. Plant. Pathol. 2006, 55, 137–144. [Google Scholar] [CrossRef]
- Herman, R.; Zvirin, Z.; Kovalski, I.; Freeman, S.; Denisov, Y.; Zuri, G.; Katzir, N.; Perl-Treves, R.; Pitrat, M. Characterization of Fusarium race 1.2 resistance in melon and mapping of a major QTL for this trait near a fruit netting locus. In Proceedings of the IXth EUCARPIA Meeting on Genetics and Breeding of Cucurbitaceaee, Avignon, France, 21–24 May 2008; pp. 149–156. [Google Scholar] [CrossRef]
- Song, H.H.; Lee, H.S.; Jeong, J.H.; Park, H.S.; Chan, L. Diversity in Beauvericin and Enniatins H, I, and MK1688 by Fusarium oxysporum isolated from potato. Int. J. Food Microbiol. 2008, 122, 296–301. [Google Scholar] [CrossRef]
- Son, S.W.; Kim, H.Y.; Choi, G.J.; Lim, H.K.; Jang, K.S.; Lee, S.O.; Lee, S.; Sung, N.D.; Kim, C.J. Bikaverin and fusaric acid from Fusarium oxysporum show antioomycete activity against Phytophthora infestans. J. Appl. Microbiol. 2008, 104, 692–698. [Google Scholar] [CrossRef]
- Waskiewicz, A.; Golinski, P.; Karolewski, Z.; Irzykowska, L.; Weber, Z. Formation of fumonisins and other secondary metabolites by Fusarium oxysporum and F. proliferatum: A comparative study. Food Addit. Contam. Part A 2010, 27, 608–615. [Google Scholar] [CrossRef] [Green Version]
- Zaccardelli, M.; Spasiano, A.; Bazzi, C.; Merighi, M. Identificationand in planta detection of Pseudomonas syringae pv tomatousing PCR amplification of hrpZpst. Eur. J. Plant. Pathol. 2005, 111, 85–90. [Google Scholar] [CrossRef]
- George, J.R.; Vanneste, J.L.; Cornish, D.A.; Pushparajah, I.P.S.; Yu, J.; Templeton, M.D.; Everett, K.R. Detection of Pseudomonas syringae pv. actinidiae using polymerase chain reaction (PCR) primers based on the 16S–23S rDNA inter transcribed spacer region and comparison with PCR primers based on other gene regions. Plant. Pathol. 2010, 59, 453–464. [Google Scholar] [CrossRef]
- Niessen, L.; Luo, J.; Denschlag, C.; Vogel, R.F. The application of loop-mediated isothermal amplification (LAMP) in food testing for bacterial pathogens and fungal contaminants. Food Microbiol. 2013, 36, 191–206. [Google Scholar] [CrossRef] [PubMed]
- Lu, T.F.; Tao, L.Y.; Yu, H.B.; Zhang, H.; Wu, Y.J.; Wu, S.G.; Zhou, J. Development of a reverse transcription loop mediated isothermal amplification assays for the detection of Mouse reovirus type 3 in laboratory mice. Sci. Rep. 2021, 11, 3508. [Google Scholar] [CrossRef] [PubMed]
- Yao, J.A.; Huang, P.; Hou, X.Y.; Yu, D.Y. Rapid detection by a loop-mediated isothermal amplification assays based on EF-1α gene for stem rot on Cymbidium ensifolium. Eur. J. Hortic. Sci. 2021, 86, 212–218. [Google Scholar] [CrossRef]
- Notomi, T.; Okayama, H.; Masubuchi, H.; Yonekawa, T.; Watanabe, K.; Amino, N.; Hase, T. Loop-mediated isothermal amplification of DNA. Nucleic Acids Res. 2000, 28, 63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Iwamoto, T.; Sonobe, T.; Hayashi, K. Loop-Mediated Isothermal Amplification for Direct Detection of Mycobacterium tuberculosis Complex, M. avium, and M. intracellulare in Sputum Samples. J. Clin. Microbiol. 2003, 41, 2616–2622. [Google Scholar] [CrossRef] [Green Version]
- Duan, Y.B.; Yang, Y.; Wang, J.X.; Liu, C.C.; He, L.L.; Zhou, M.G. Development and application of loop-mediated isothermal amplification for detecting the highly benzimidazole-resistant isolates in Sclerotinia sclerotiorum. Sci. Rep. 2015, 5, 17278. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mori, Y.; Notomi, T. Loop-mediated isothermal amplification (LAMP): A rapid, accurate, and cost-effective diagnostic method for infectious diseases. J. Infect. Chemother. 2009, 15, 62–69. [Google Scholar] [CrossRef]
- Verma, S.; Avishek, K.; Sharma, V.; Negi, N.S.; Ramesh, V.; Salotra, P. Application of loop-mediated isothermal amplification assays for the sensitive and rapid diagnosis of visceral leishmaniasis and post-kala-azar dermal leishmaniasis. Diagn. Microbiol. Infect. Dis. 2013, 75, 390–395. [Google Scholar] [CrossRef] [PubMed]
- Hara, K.Y.; Yoshino, M.; Kojima, T.; Ikedo, M. Loop-mediated isothermal amplification for the rapid detection of Salmonella. Fems Microbiol. Lett. 2010, 253, 155–161. [Google Scholar] [CrossRef] [Green Version]
- Nirenberg, H.I.; O′Donnell, K. New Fusarium species and combinations within the Gibberella fujikuroi species complex. Mycologia 1998, 90, 434–458. [Google Scholar] [CrossRef]
- Taylor, J.W.; Jacobson, D.J.; Fisher, M.C. The evolution of asexual fungi: Reproduction, Speciation and Classification. Immunology 1999, 121, 383–391. [Google Scholar] [CrossRef]
- Amatulli, M.; Spadaro, D.; Gullino, M.L.; Garibaldi, A. Corrigendum: Molecular identification of Fusarium spp. associated with bakanae disease of rice in Italy and assessment of their pathogenicity. Plant. Pathol. 2012, 61, 820. [Google Scholar] [CrossRef]
- White, T.; Bruns, T.; Lee, S.; Taylor, F.; White, T.J.; Lee, S.H.; Taylor, L.; Shawe-taylor, J. Amplification and direct sequencing of fungal ribosomal RNA genes for phylogenetics. In PCR Protocols; Academic Press: San Diego, CA, USA, 1990; pp. 315–322. [Google Scholar] [CrossRef]
- Li, H.S.; Ye, W.; Wang, Y.; Chen, X.H. RNA sequencing-based exploration of the effects of far-red light on lncRNAs involved in the shade-avoidance response of D. officinale. PeerJ 2021, 9, e10769. [Google Scholar] [CrossRef]
- Zhang, M.Z.; Yu, Z.M.; Zeng, D.Q.; Si, C. Transcriptome and Metabolome Reveal Salt-Stress Responses of Leaf Tissues from Dendrobium officinale. Biomolecules 2021, 11, 736. [Google Scholar] [CrossRef]
- Almasi, M.A. Development of A Colorimetric Loop-Mediated Isothermal Amplification Assays for the Visual Detection of Fusarium oxysporum f.sp. melonis. Hortic. Plant. J. 2019, 5, 41–48. [Google Scholar] [CrossRef]
- Shamsher, A.; Hasmi, S.K.; Aftab, M.A.; Khan, R.U. Biological Management of Chrysanthemum Wilt (Fusarium oxysporum f.sp. chrysanthemi. Int. J. Curr. Microbiol. Appl. Sci. 2020, 9, 639–644. [Google Scholar] [CrossRef]
- Zhang, J.; Li, T.; Cai, Y.F.; Wang, Y.Z. Genetic and environmental effects on allometry of the medicinal plant Dendrobium officinale (Orchidaceae) from Yunnan, southwest China. Pak. J. Bot. 2021, 53, 1675–1682. [Google Scholar] [CrossRef]
- Bao, Y.X.; Sun, H.J.; Li, Y.F.; Duan, Z.Z.; McCord, P.H.; Cui, Y.P.; Zhang, M.Q. First Report of Fusarium oxysporum isolate gx3 Causing Sugarcane Pokkah Boeng in Guangxi of China. Plant. Dis. 2016, 100, 1785. [Google Scholar] [CrossRef]
- Pastrana, A.M.; Cline, W.; Wong, T.W.; Watson, D.C.; Mercier, J.; Ivors, K.; Broome, J.C.; Quesada-Ocampo, L.M.; Gordon, T.R. First report of Fusarium Wilt of Blackberry Caused by Fusarium oxysporum f. sp. mori in North Carolina. Plant. Dis. 2019, 104, 971. [Google Scholar] [CrossRef]
- Zhu, Y.; Lujan, P.; Wedegaertner, T.; Nichols, R.L.; Sanogo, S. First Report of Fusarium oxysporum f. sp. vasinfectum Race 4 Causing Fusarium Wilt of Cotton in New Mexico, U.S.A. Plant. Dis. 2020, 104, 588. [Google Scholar] [CrossRef]
- Bowers, J.H.; Locke, J.C. Effect of Botanical Extracts on the Population Density of Fusarium oxysporum in Soil and Control of Fusarium Wilt in the Greenhouse. Plant. Dis. 2000, 84, 300–305. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cal, A.D.; Pascual, S.; Larena, I.; Melgarejo, P. Biological control of Fusarium oxysporum f. sp. lycopersici. Plant. Pathol. 2010, 44, 909–917. [Google Scholar] [CrossRef]
- Hadian, S.; Rahnama, K.; Jamali, S.; Eskandari, A. Comparing neem extract with chemical control on Fusarium oxysporum and Meloidogyne incognita complex of tomato. Adv. Environ. Biol. 2011, 5, 2052–2057. [Google Scholar]
- Arif, M.; Chawla, S.; Zaidi, N.W.; Rayar, J.K.; Singh, U.S. Development of specific primers for genus Fusarium and F. solani using rDNA sub-unit and transcription elongation factor (TEF-1α) gene. Afr. J. Biotechnol. 2014, 11, 444–447. [Google Scholar] [CrossRef]
- Uribe-Cortés, T.B.; Silva-Rojas, H.V.; Mendoza-Onofre, L.E.; Velázquez-Cruz, C.; Rebollar-Alviter, N. Identificación de especies de Fusarium aisladas de semillas sintomáticas y asintomáticas de maíz con base en el gen TEF-1α. Rev. Fitotec. Mex. Publ. Por La Soc. Mex. De Fitogenética 2020, 43, 79. [Google Scholar] [CrossRef]
- Huang, S.; Xu, Y.; Yan, X.; Shang, Y.; Zhu, P.; Tian, W.; Xu, W. Development and application of a quantitative loop-mediated isothermal amplification method for detecting genetically modified maize MON863. J. Sci. Food Agric. 2015, 95, 253–259. [Google Scholar] [CrossRef]
- Singh, R.; Singh, D.P.; Savargaonkar, D.; Singh, O.P.; Bhatt, R.M.; Valecha, N. Evaluation of SYBR green I based visual loop-mediated isothermal amplification (LAMP) assays for genus and species-specific diagnosis of malaria in P. vivax and P. falciparum endemic regions. J. Vector Borne Dis. 2017, 54, 54–60. [Google Scholar]
- Zhang, X.; Zhang, H.; Pu, J.; Qi, Y.; Yu, Q.; Xie, Y.; Peng, J. Development of a Real-Time Fluorescence Loop-Mediated Isothermal Amplification Assays for Rapid and Quantitative Detection of Fusarium oxysporum f. sp. cubense Tropical Race 4 In Soil. PLoS ONE 2013, 349, 127–134. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ayukawa, Y.; Hanyuda, S.; Fujita, N.; Komatsu, K.; Arie, T. Novel loop-mediated isothermal amplification (LAMP) assays with a universal QProbe can detect SNPs determining races in plant pathogenic fungi. Sci. Rep. 2017, 7, 4253. [Google Scholar] [CrossRef] [PubMed]
- Ghosh, R.; Tarafdar, A.; Sharma, M. Rapid and sensitive diagnoses of dry root rot pathogen of chickpea (Rhizoctonia bataticola (Taub.) Butler) using loop-mediated isothermal amplification assays. Sci. Rep. 2017, 7, 42737. [Google Scholar] [CrossRef] [PubMed]
- Peng, J.; Zhang, H.; Chen, F.P.; Zhang, X.; Xie, Y.X.; Hou, X.W.; Li, G.Y.; Pu, J.J. Rapid and quantitative detection of Fusarium oxysporum f. sp. cubense race 4 in soil by real-time fluorescence loop-mediated isothermal amplification. J. Appl. Microbiol. 2015, 117, 1740–1749. [Google Scholar] [CrossRef]
- Cao, Y.Y.; Wang, L.; Duan, L.; Li, J.; Ma, J.; Xie, S.; Shi, L.; Li, H. Development of a real-time fluorescence loop-mediated isothermal amplification assays for rapid and quantitative detection of Ustilago maydis. Sci. Rep. 2017, 7, 13394. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, C.; Dai, T.; Zhang, H.F.; Wang, Y.C.; Zheng, X.B. Development of a Loop-Mediated Isothermal Amplification Assays to Detect Fusarium oxysporum. J. Phytopathol. 2015, 163, 63–66. [Google Scholar] [CrossRef]
- Lan, C.Z.; Ruan, H.C.; Yang, X.J.; Yao, J.A.; Jiang, J.X. Development of a loop-mediated isothermal amplification assays for sensitive and specific detection of Fusarium oxysporum f. sp. cucumerinum Owen. Phytoparasitica 2018, 46, 283–293. [Google Scholar] [CrossRef]
- Olivares, B.O.; Rey, J.C.; Lobo, D.; Navas-Cortés, J.A.; Gómez, J.A.; Landa, B.B. Fusarium Wilt of Bananas: A Review of Agro-Environmental Factors in the Venezuelan Production System Affecting Its Development. Agronomy 2021, 11, 986. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic tree for F. oxysporum based on translation elongation factor-1α (TEF-1α) gene sequences.
Figure 1.
Phylogenetic tree for F. oxysporum based on translation elongation factor-1α (TEF-1α) gene sequences.

Figure 2.
The nucleotide sequence alignment of the translation elongation factor-1α (TEF-1α) genes were used to design the LAMP primers.
Figure 2.
The nucleotide sequence alignment of the translation elongation factor-1α (TEF-1α) genes were used to design the LAMP primers.

Figure 3.
Isolation of pathogenic fungi from soft rot in Dendrobium officinale. (A) = The soft rot on Dendrobium officinale plant in the field; (B) = the morphology of strain XY1E208 after growing on PDA medium for 7 days; (C) = conidia morphology of strain XI1E208.
Figure 3.
Isolation of pathogenic fungi from soft rot in Dendrobium officinale. (A) = The soft rot on Dendrobium officinale plant in the field; (B) = the morphology of strain XY1E208 after growing on PDA medium for 7 days; (C) = conidia morphology of strain XI1E208.

Figure 4.
Optimization of LAMP assays. (A) = Optimization of LAMP reaction temperature for detection of F. oxysporum. Lane M, Ds 2000 DNA molecular weight marker ladder; Lane 1, 55 °C; Lane 2, 58 °C; Lane 3, 60 °C; Lane 4, 62 °C; Lane 5, 64 °C; Lane 6, 65 °C; Lane 7, 66 °C; Lane 8, 68 °C; Lane 9, 70 °C; Lane 10, negative control. (B) = Optimization of LAMP reaction time for detection of F. oxysporum. Lane M, Ds 2000 DNA molecular weight marker ladder; Lane 1, 15 min; Lane 2, 30 min; Lane 3, 45 min; Lane 4, 60 min; Lane 5, 75 min; Lane 6, 90 min; Lane 7, negative control. (C) = Optimization of LAMP reaction inner primer (FIP/BIP) concentration for detection of F. oxysporum. Lane M, Ds 2000 DNA molecular weight marker ladder; Lane 1, 0.4 μM; Lane 2, 0.8 μM; Lane 3, 1.2 μM; Lane 4, 1.6 μM; Lane 5, 2 μM; Lane 6, negative control. (D) = Optimization of LAMP reaction outer primer (F3/B3) concentration for detection of F. oxysporum. Lane M, Ds 2000 DNA molecular weight marker ladder; Lane 1, 0.2 μM; Lane 2, 0.4 μM; Lane 3, 0.6 μM; Lane 4, 0.8 μM; Lane 5, 1 μM; Lane 6, negative control. (E) = Optimization of LAMP reaction Mg2+ concentrations for detection of F. oxysporum. Lane M, Ds 2000 DNA molecular weight marker ladder; Lane 1, 2 mM; Lane 2, 3 mM; Lane 3, 4 mM; Lane 4, 5 mM; Lane 5, 6 mM; Lane 6, 7 mM; Lane 7, 8 mM; Lane 8, 9 mM; Lane 9, 10 mM; Lane 10, negative control.
Figure 4.
Optimization of LAMP assays. (A) = Optimization of LAMP reaction temperature for detection of F. oxysporum. Lane M, Ds 2000 DNA molecular weight marker ladder; Lane 1, 55 °C; Lane 2, 58 °C; Lane 3, 60 °C; Lane 4, 62 °C; Lane 5, 64 °C; Lane 6, 65 °C; Lane 7, 66 °C; Lane 8, 68 °C; Lane 9, 70 °C; Lane 10, negative control. (B) = Optimization of LAMP reaction time for detection of F. oxysporum. Lane M, Ds 2000 DNA molecular weight marker ladder; Lane 1, 15 min; Lane 2, 30 min; Lane 3, 45 min; Lane 4, 60 min; Lane 5, 75 min; Lane 6, 90 min; Lane 7, negative control. (C) = Optimization of LAMP reaction inner primer (FIP/BIP) concentration for detection of F. oxysporum. Lane M, Ds 2000 DNA molecular weight marker ladder; Lane 1, 0.4 μM; Lane 2, 0.8 μM; Lane 3, 1.2 μM; Lane 4, 1.6 μM; Lane 5, 2 μM; Lane 6, negative control. (D) = Optimization of LAMP reaction outer primer (F3/B3) concentration for detection of F. oxysporum. Lane M, Ds 2000 DNA molecular weight marker ladder; Lane 1, 0.2 μM; Lane 2, 0.4 μM; Lane 3, 0.6 μM; Lane 4, 0.8 μM; Lane 5, 1 μM; Lane 6, negative control. (E) = Optimization of LAMP reaction Mg2+ concentrations for detection of F. oxysporum. Lane M, Ds 2000 DNA molecular weight marker ladder; Lane 1, 2 mM; Lane 2, 3 mM; Lane 3, 4 mM; Lane 4, 5 mM; Lane 5, 6 mM; Lane 6, 7 mM; Lane 7, 8 mM; Lane 8, 9 mM; Lane 9, 10 mM; Lane 10, negative control.

Figure 5.
Specificity of LAMP detection of F. oxysporum. (A) = Assessment based on SYBR Green I visualization of color change; (B) = LAMP products analyzed by agarose gel electrophoresis. 1, Fusarium xysporum; 2, Fusarium proliferatum; 3, Fusarium equiseti; 4, Fusarium solani; 5, Fusarium chlamydosporum; 6, Fusarium fujikuroi; 7, Fusarium graminearum; 8, Colletotrichum fructicola; 9, Epicoccum sorghinum; 10, Neurospora sitophila; 11, Lasiodiplodia pseudotheobromae; 12, Trichoderma harzianum; 13, Botryosphaeria dothidea; 14, Phomopsis sp.; 15, Pythium ultimum; 16, Magnaporthe oryzae; 17, Rhizoctonia solani; 18, Botrytis cinerea; 19, double-distilled water as negative control.
Figure 5.
Specificity of LAMP detection of F. oxysporum. (A) = Assessment based on SYBR Green I visualization of color change; (B) = LAMP products analyzed by agarose gel electrophoresis. 1, Fusarium xysporum; 2, Fusarium proliferatum; 3, Fusarium equiseti; 4, Fusarium solani; 5, Fusarium chlamydosporum; 6, Fusarium fujikuroi; 7, Fusarium graminearum; 8, Colletotrichum fructicola; 9, Epicoccum sorghinum; 10, Neurospora sitophila; 11, Lasiodiplodia pseudotheobromae; 12, Trichoderma harzianum; 13, Botryosphaeria dothidea; 14, Phomopsis sp.; 15, Pythium ultimum; 16, Magnaporthe oryzae; 17, Rhizoctonia solani; 18, Botrytis cinerea; 19, double-distilled water as negative control.

Figure 6.
Sensitivity of LAMP and conventional PCR for detection F. oxysporum genomic DNA. (A) = Detection by LAMP with SYBR Green I staining; (B) = LAMP products analyzed by agarose gel electrophoresis; (C) = conventional PCR analyzed on gel electrophoresis. The sensitivities of these assays were evaluated using serially diluted genomic DNA as follows: 1, 50 ng/µL; 2, 5 ng/µL; 3, 0.5 ng/µL; 4, 50 pg/µL; 5, 5 pg/µL; 6, 0.5 pg/µL; 7, 50 fg/µL; 8, 5 fg/µL; 9, 0.5 fg/µL; 10, negative control. M, DL 2000 DNA marker.
Figure 6.
Sensitivity of LAMP and conventional PCR for detection F. oxysporum genomic DNA. (A) = Detection by LAMP with SYBR Green I staining; (B) = LAMP products analyzed by agarose gel electrophoresis; (C) = conventional PCR analyzed on gel electrophoresis. The sensitivities of these assays were evaluated using serially diluted genomic DNA as follows: 1, 50 ng/µL; 2, 5 ng/µL; 3, 0.5 ng/µL; 4, 50 pg/µL; 5, 5 pg/µL; 6, 0.5 pg/µL; 7, 50 fg/µL; 8, 5 fg/µL; 9, 0.5 fg/µL; 10, negative control. M, DL 2000 DNA marker.

Figure 7.
The feasibility detection of the LAMP assay. LAMP reactions were inspected by adding SYBR Green I dye. (A) = LAMP detection of F. oxysporum from tissues after infection: 1, purified genomic DNA of F. oxysporum (positive control); 2–10, DNA from D. officinale stems infected by F. oxysporum; 11, DNA from healthy D. officinale stems (negative control); (B) = LAMP detection of F. oxysporum from soil after infection: 1, purified genomic DNA of F. oxysporum (positive control); 2–10, DNA extracted from soil after infection; 11, DNA from sterilized soil (negative control); (C) = LAMP detection of field issues: 1, purified genomic DNA of F. oxysporum (positive control); 2–7, DNA extracted from field issues; 8, DNA from healthy D. officinale stems (negative control).
Figure 7.
The feasibility detection of the LAMP assay. LAMP reactions were inspected by adding SYBR Green I dye. (A) = LAMP detection of F. oxysporum from tissues after infection: 1, purified genomic DNA of F. oxysporum (positive control); 2–10, DNA from D. officinale stems infected by F. oxysporum; 11, DNA from healthy D. officinale stems (negative control); (B) = LAMP detection of F. oxysporum from soil after infection: 1, purified genomic DNA of F. oxysporum (positive control); 2–10, DNA extracted from soil after infection; 11, DNA from sterilized soil (negative control); (C) = LAMP detection of field issues: 1, purified genomic DNA of F. oxysporum (positive control); 2–7, DNA extracted from field issues; 8, DNA from healthy D. officinale stems (negative control).

Figure 8.
LAMP assay random sampling detection of F. oxysporum in potted plants of D. officinale before pesticide control. (A) = LAMP assay to detect F. oxysporum in potted plants of D. officinale from random samples before the symptoms of soft rot were exposed in pots: 1, purified genomic DNA of F. oxysporum (positive control); 2–13, DNA from D. officinale stems before the symptoms of soft rot were exposed in pots; 11, DNA from healthy D. officinale stems (negative control); (B) = LAMP assay to detect F. oxysporum in potted plants of D. officinale from random samples after the symptoms of soft rot were exposed in pots: 1, purified genomic DNA of F. oxysporum (positive control); 2–13, DNA from D. officinale stems after the symptoms of soft rot were exposed in pots; 11, DNA from healthy D. officinale stems (negative control).
Figure 8.
LAMP assay random sampling detection of F. oxysporum in potted plants of D. officinale before pesticide control. (A) = LAMP assay to detect F. oxysporum in potted plants of D. officinale from random samples before the symptoms of soft rot were exposed in pots: 1, purified genomic DNA of F. oxysporum (positive control); 2–13, DNA from D. officinale stems before the symptoms of soft rot were exposed in pots; 11, DNA from healthy D. officinale stems (negative control); (B) = LAMP assay to detect F. oxysporum in potted plants of D. officinale from random samples after the symptoms of soft rot were exposed in pots: 1, purified genomic DNA of F. oxysporum (positive control); 2–13, DNA from D. officinale stems after the symptoms of soft rot were exposed in pots; 11, DNA from healthy D. officinale stems (negative control).


{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Table 1.
Different strains of fungus used in this study to test the specificity of the LAMP assay.
| Species | Host | Geographical Location | Number of Strains | LAMP detection | |
|---|---|---|---|---|---|
| Agarose Gel | SYBR Green I | ||||
| Fusarium oxysporum | Dendrobium officinale | Xingyi, Guizhou | 1 | + | + |
| Fusarium proliferatum | Dendrobium officinale | Huaxi, Guiyang | 1 | − | − |
| Fusarium equiseti | Dendrobium officinale | Huaxi, Guiyang | 1 | − | − |
| Fusarium solani | Dendrobium officinale | Libo, Guiyang | 1 | − | − |
| Fusarium chlamydosporum | Dendrobium officinale | Anlong, Guiyang | 1 | − | − |
| Fusarium fujikuroi | Plum | Huishui, Guiyang | 1 | − | − |
| Fusarium graminearum | Kiwi Fruit | Xifeng, Guiyang | 1 | − | − |
| Colletotrichum fructicola | Dendrobium officinale | Sansui, Guizhou | 1 | − | − |
| Epicoccum sorghinum | Dendrobium officinale | Xingyi, Guizhou | 1 | − | − |
| Neurospora sitophila | Dendrobium officinale | Jinping, Guizhou | 1 | − | − |
| Lasiodiplodia pseudotheobromae | Dendrobium officinale | Jinping, Guizhou | 1 | − | − |
| Trichoderma harzianum | Soil | Jinping, Guizhou | 1 | − | − |
| Botryosphaeria dothidea | Dendrobium officinale | Jinping, Guizhou | 1 | − | − |
| Phomopsis sp. | Kiwi Fruit | Xifeng, Guizhou | 1 | − | − |
| Pythium ultimum | Dendrobium officinale | Huaxi, Guizhou | 1 | − | − |
| Magnaporthe grisea | Oryza sativa | Huaxi, Guizhou | 1 | − | − |
| Rhizoctonia solani | Oryza sativa | Huaxi, Guizhou | 1 | − | − |
| Botrytis cinerea | Kiwi Fruit | Xifeng, Guizhou | 1 | − | − |
Note: bold letter = the Fusarium oxysporum used for specific detection in this study.
Table 2.
Primers used for LAMP assays to detect F. oxysporum.
| Primer Name | Sequence (5′–3′) | Length |
|---|---|---|
| F3 | ACTGCTTGACACGTGACG | 18 |
| B3 | CACTTTCCCTTCGATCGCG | 19 |
| FIP | ACTTACCCCGCCACTTGAGCACGCACTCATTGAGGTTGTG | 40 |
| BIP | TTGGTCTCGAGCGGGGTAGCTCCTTTGCCCATCGATTTCC | 40 |
| LF | CGTTTGCCCTCTTAACCATTCT | 22 |
| LB | GGGCACATTTCGAGTCGTAGG | 21 |
Table 3.
Effect of fungicide synergism combinations against soft rot in D. officinale.
| Treatment | Concentration (g a.i./hm2) | Average Disease Index before Spraying Fungicide | Average Disease Index 14 Days after Spraying Fungicide | Control Efficacy (%) |
|---|---|---|---|---|
| CK | / | 32.08 | 87.41 | / |
| Pyraclostrobin(A) | 4.9635 | 35.83 | 44.44 | 84.25 a |
| Picoxystrobin(B) | 11.0865 | 43.47 | 55.65 | 78.07 c |
| A:B(1:3) | 4.128 | 47.78 | 57.5 | 82.39 ab |
| Osthole(C) | 16.839 | 41.59 | 53.06 | 79.29 bc |
| Physcion(D) | 192.3 | 35.65 | 54.21 | 66.47 d |
| C:D(7:1) | 18.195 | 38.33 | 51.11 | 76.74 c |
Note: Different lowercase letters in the same column show significant difference at 0.05 level by Duncan’s multiple range test.
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Xiao, C.; Li, R. Detection and Control of Fusarium oxysporum from Soft Rot in Dendrobium officinale by Loop-Mediated Isothermal Amplification Assays. Biology 2021, 10, 1136. https://doi.org/10.3390/biology10111136
AMA Style
Xiao C, Li R. Detection and Control of Fusarium oxysporum from Soft Rot in Dendrobium officinale by Loop-Mediated Isothermal Amplification Assays. Biology. 2021; 10(11):1136. https://doi.org/10.3390/biology10111136
Chicago/Turabian StyleXiao, Caiyun, and Rongyu Li. 2021. "Detection and Control of Fusarium oxysporum from Soft Rot in Dendrobium officinale by Loop-Mediated Isothermal Amplification Assays" Biology 10, no. 11: 1136. https://doi.org/10.3390/biology10111136
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.