Purification and In Vitro Evaluation of an Anti-HER2 Affibody-Monomethyl Auristatin E Conjugate in HER2-Positive Cancer Cells

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
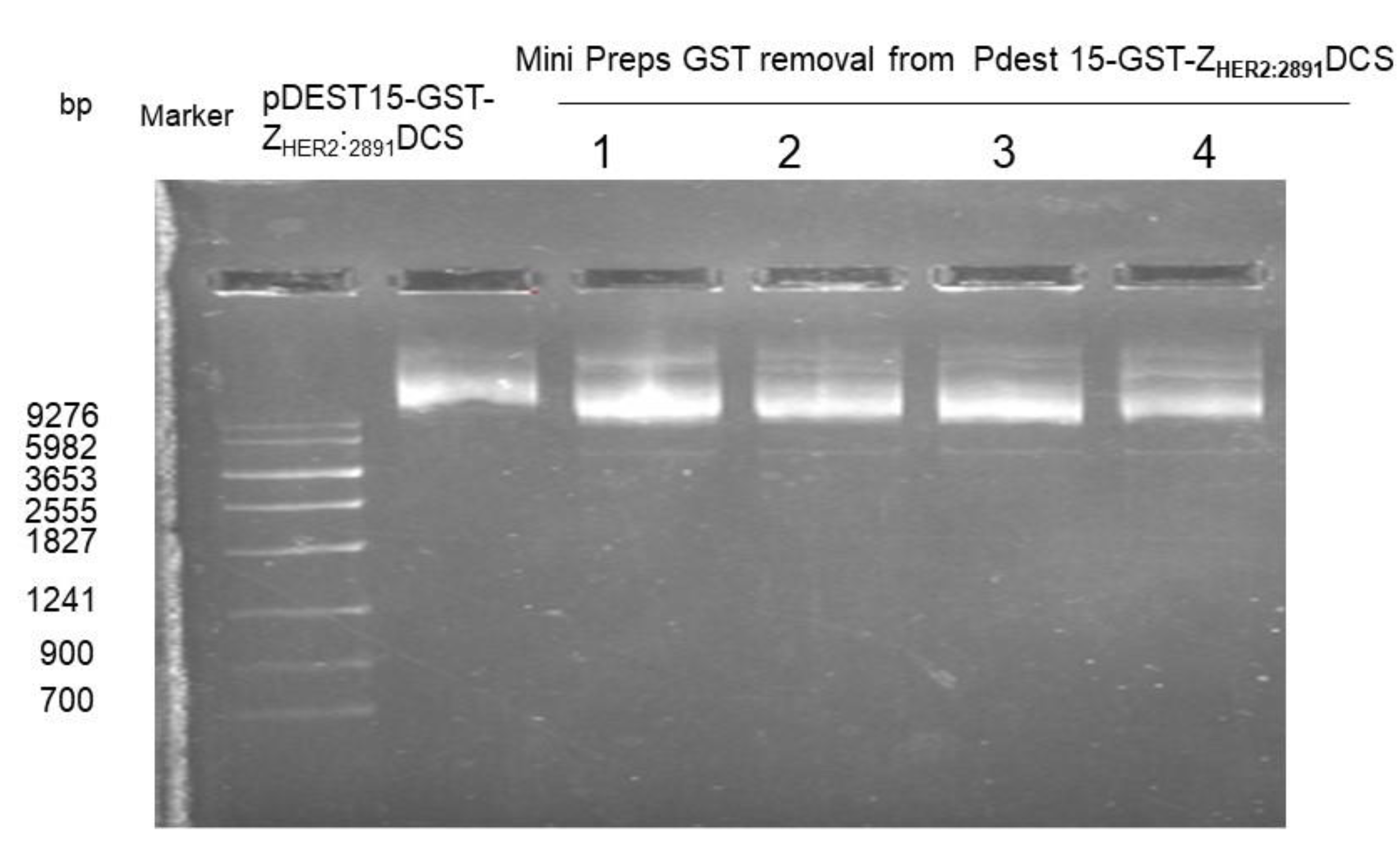
2.1. Generation of the pDEST15-GST Removed-ZHER2:2891-DCS
2.2. Affibody Expression
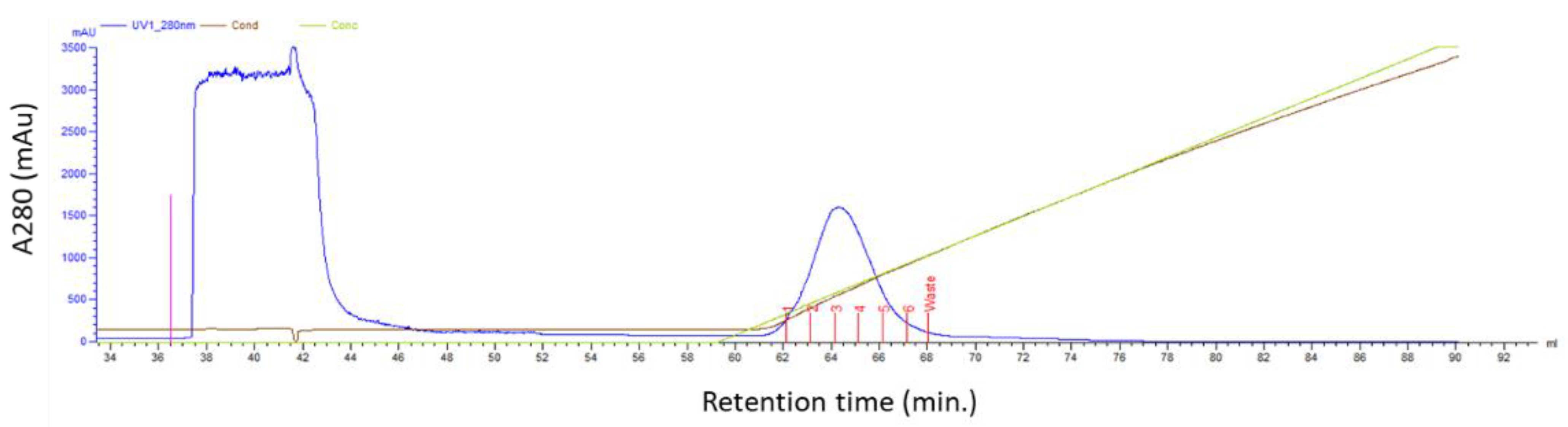
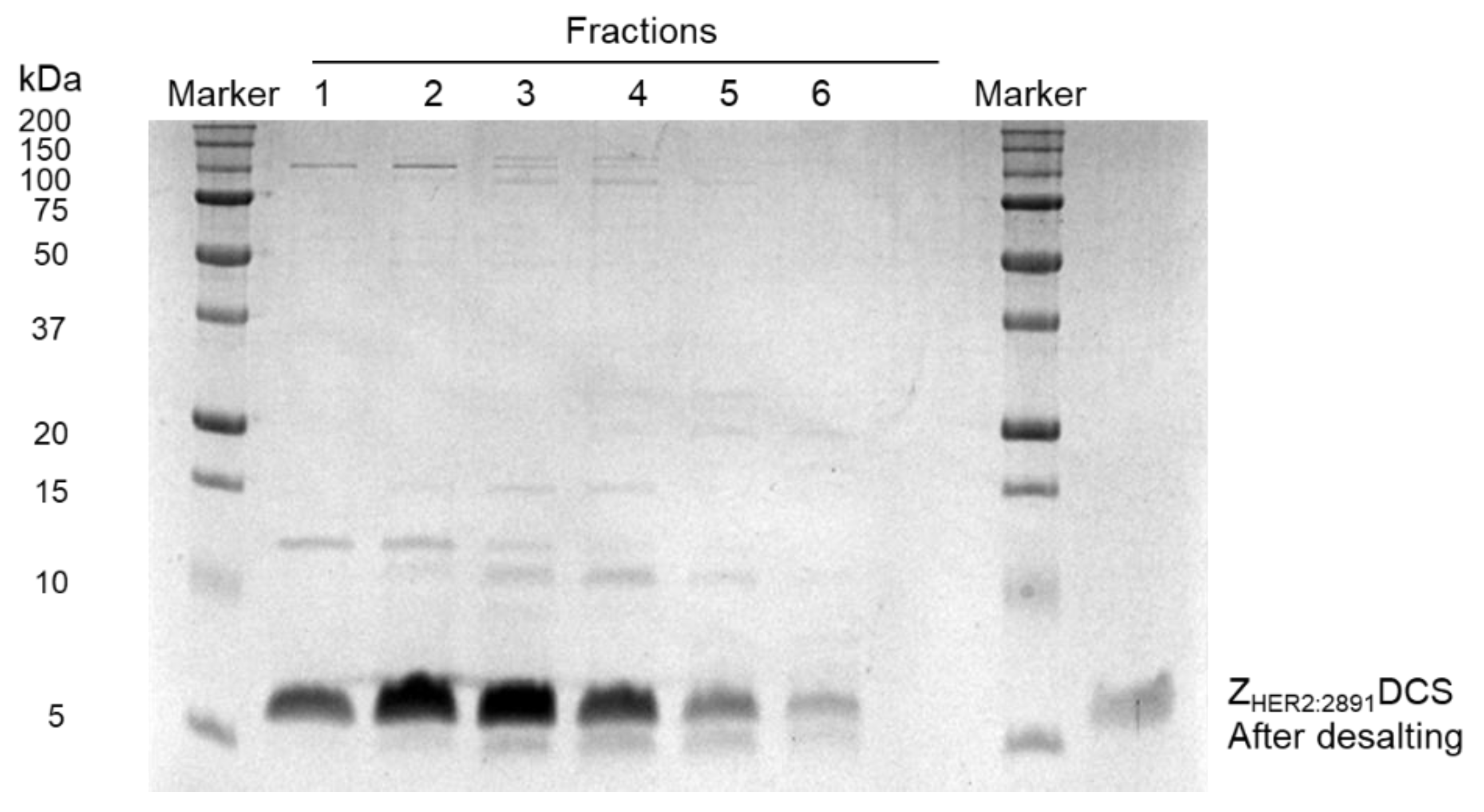
2.3. Affibody Purification
2.4. MMAE Conjugation Reaction and ZHER2:2891DCS-MMAE Purification
2.5. Reagents
2.6. Cell Lines
2.7. Cell Viability Assay
2.8. Cell Proliferation Assay
2.9. In Vitro Directional Migration (Wound Healing Assay)
2.10. Apoptosis Analysis
2.11. RNA Isolation and qRT-PCR
2.12. Western Blot Analysis
2.13. Statistical Analysis
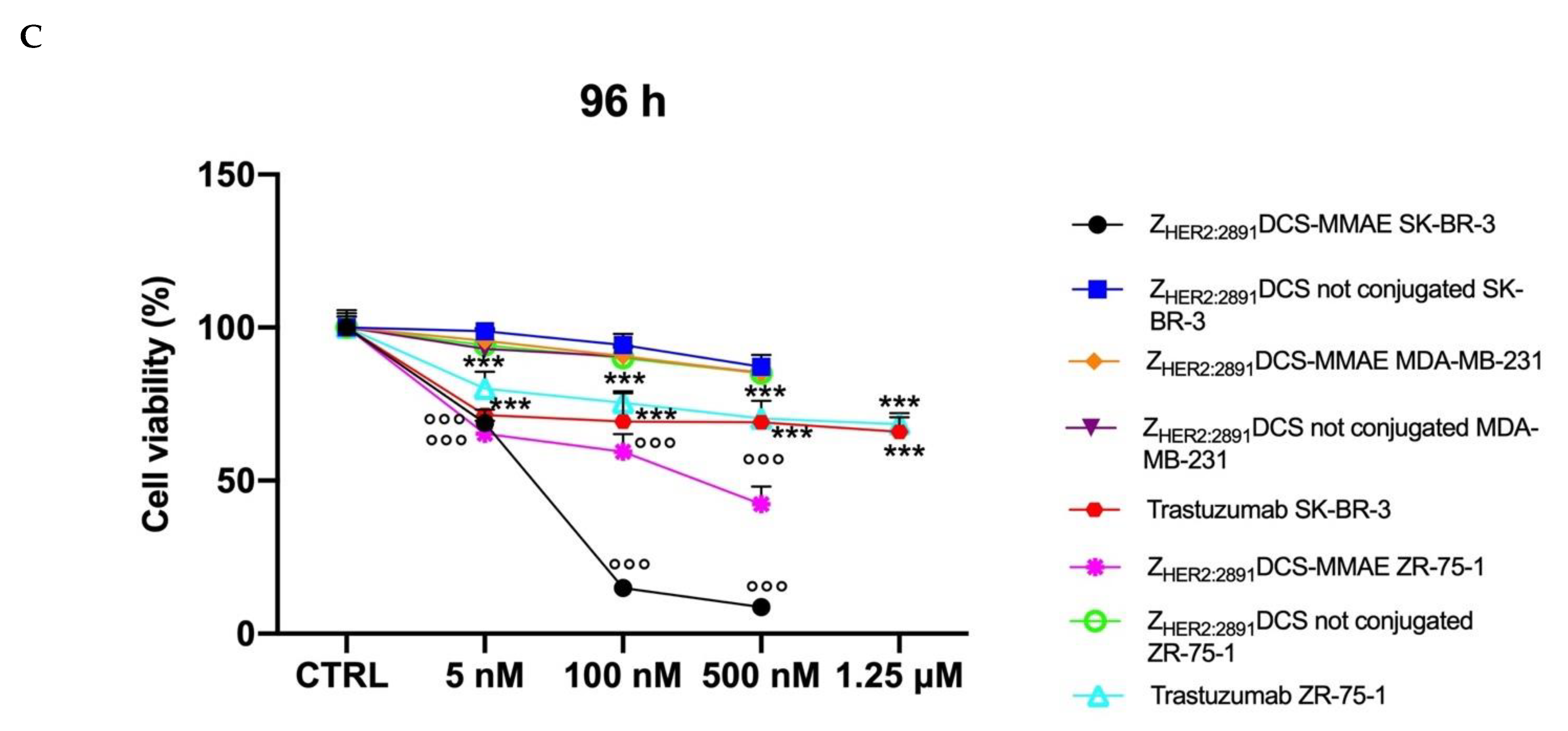
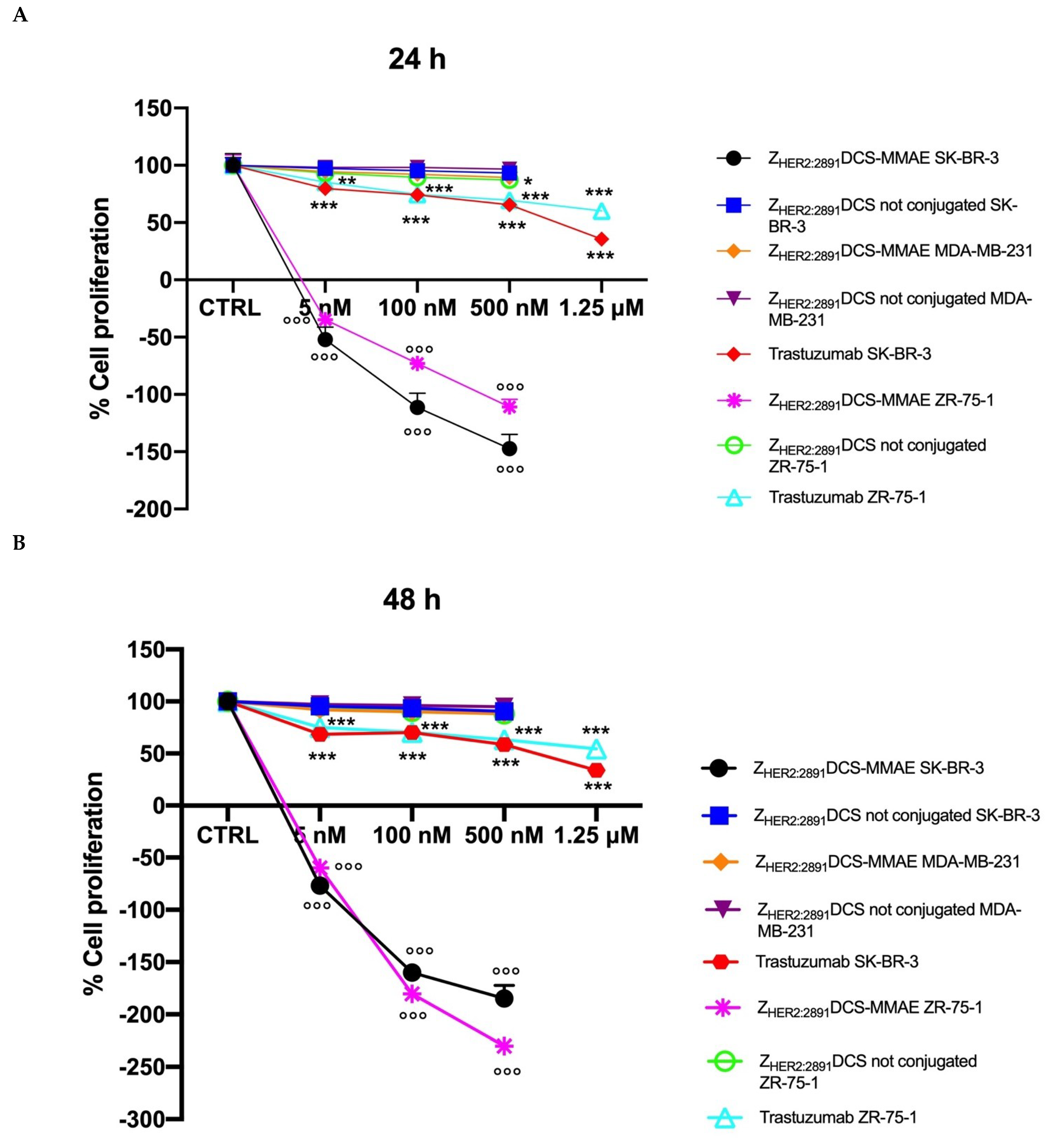
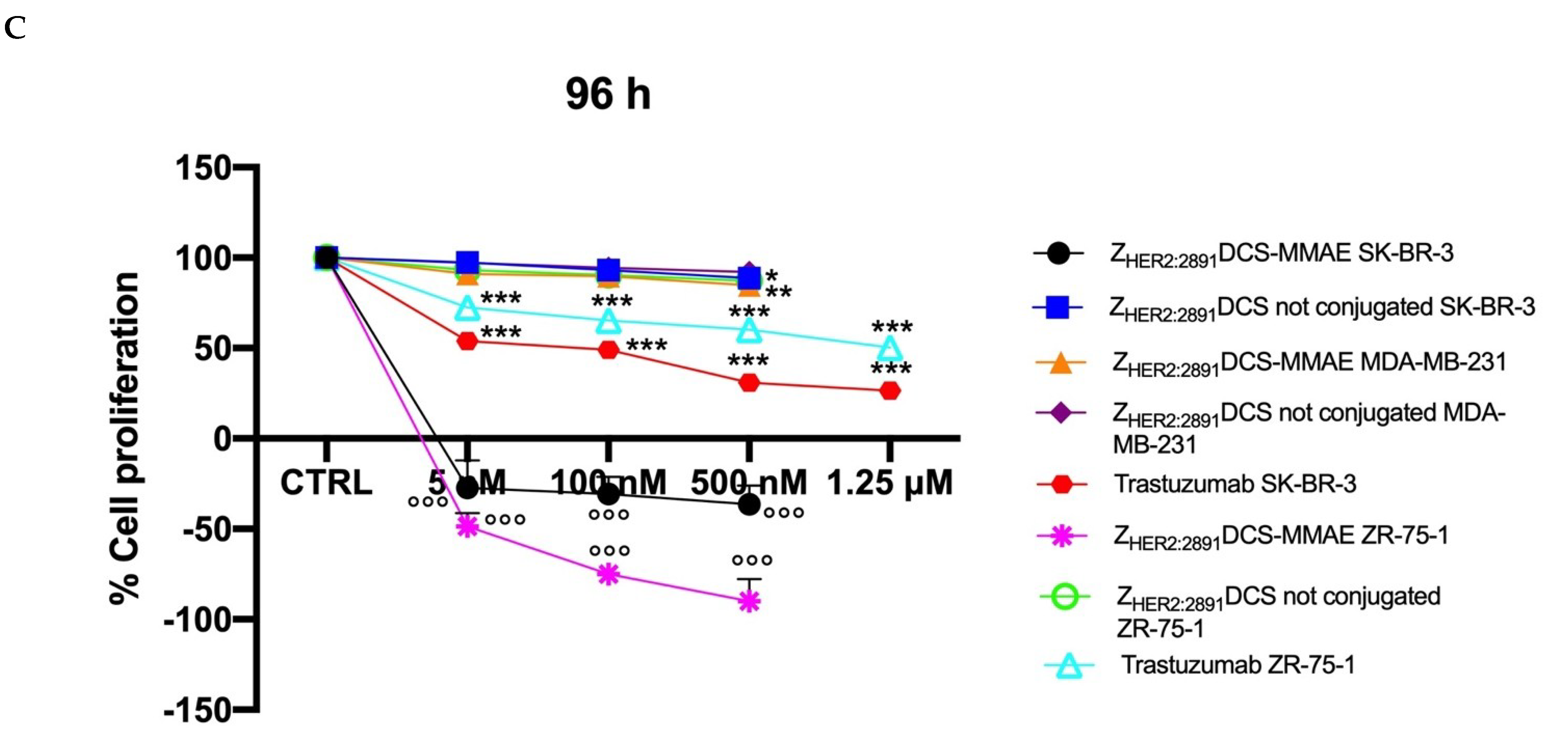
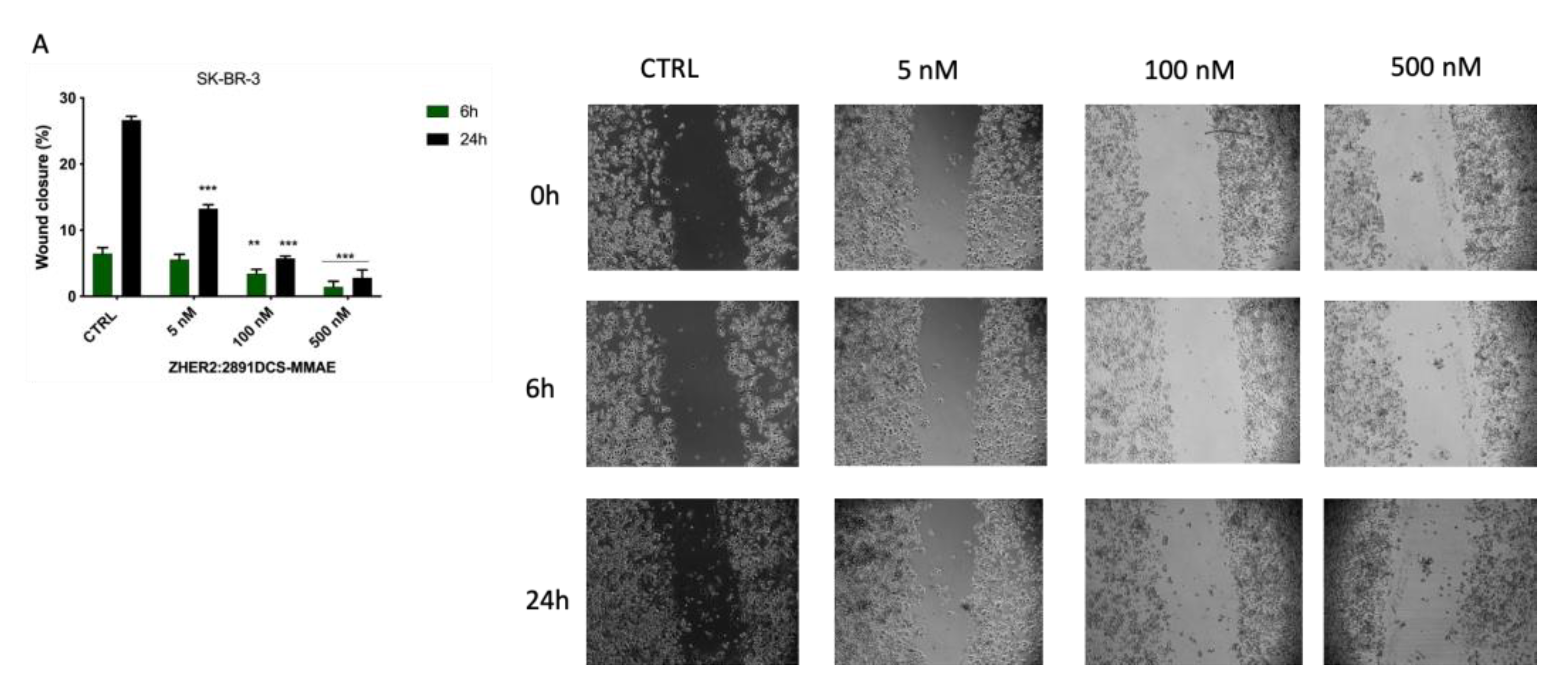
3. Results
3.1. Expression and Purification of the ZHER2:2891-DCS
3.2. Conjugation of MMAE to ZHER2:2891-DCS
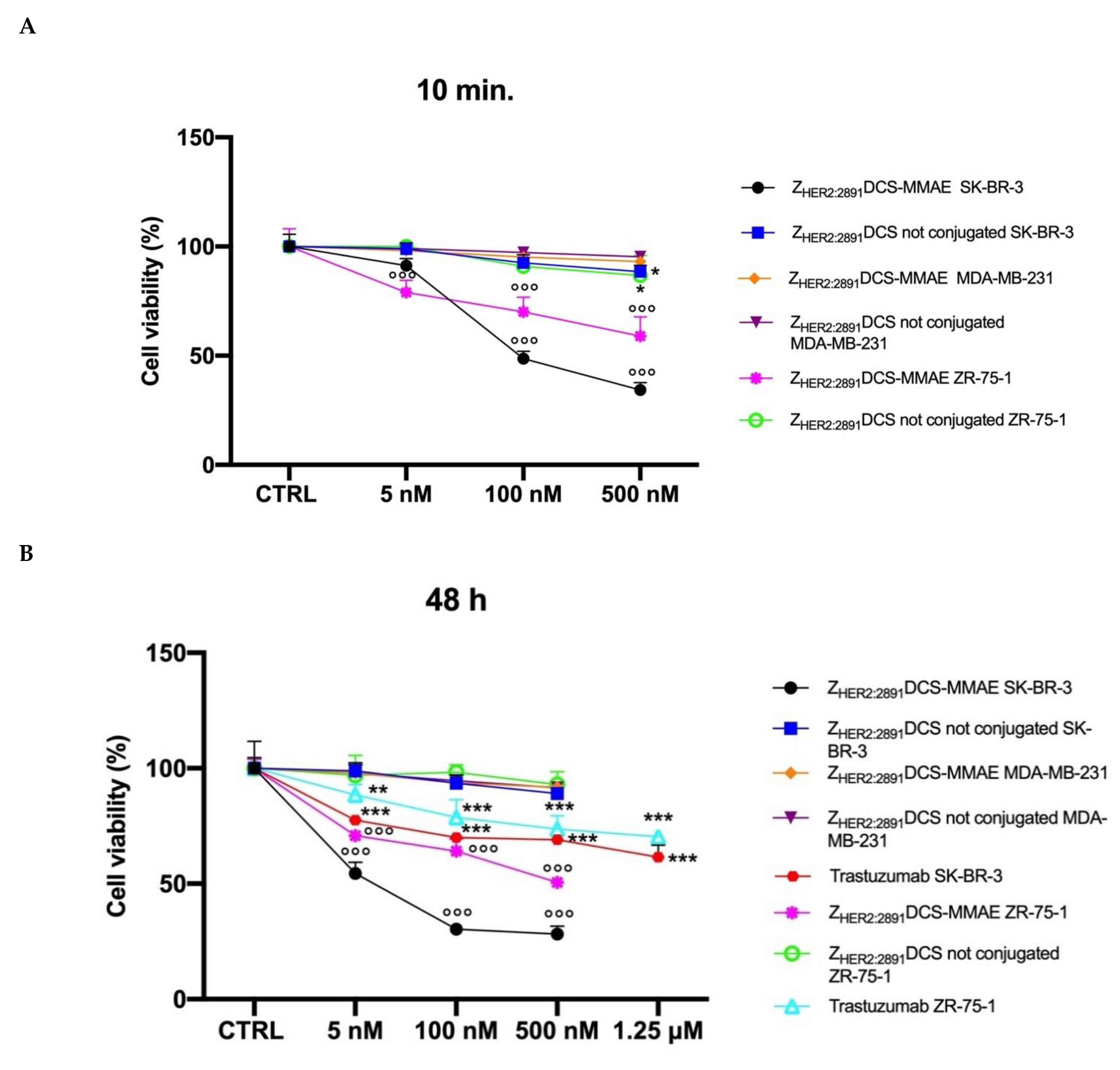
3.3. ZHER2:2891DCS-MMAE Affects HER2 Expressing Cell Line Viability In Vitro
3.4. ZHER2:2891DCS-MMAE Negatively Regulates HER2 Expressing Cell Line Proliferation
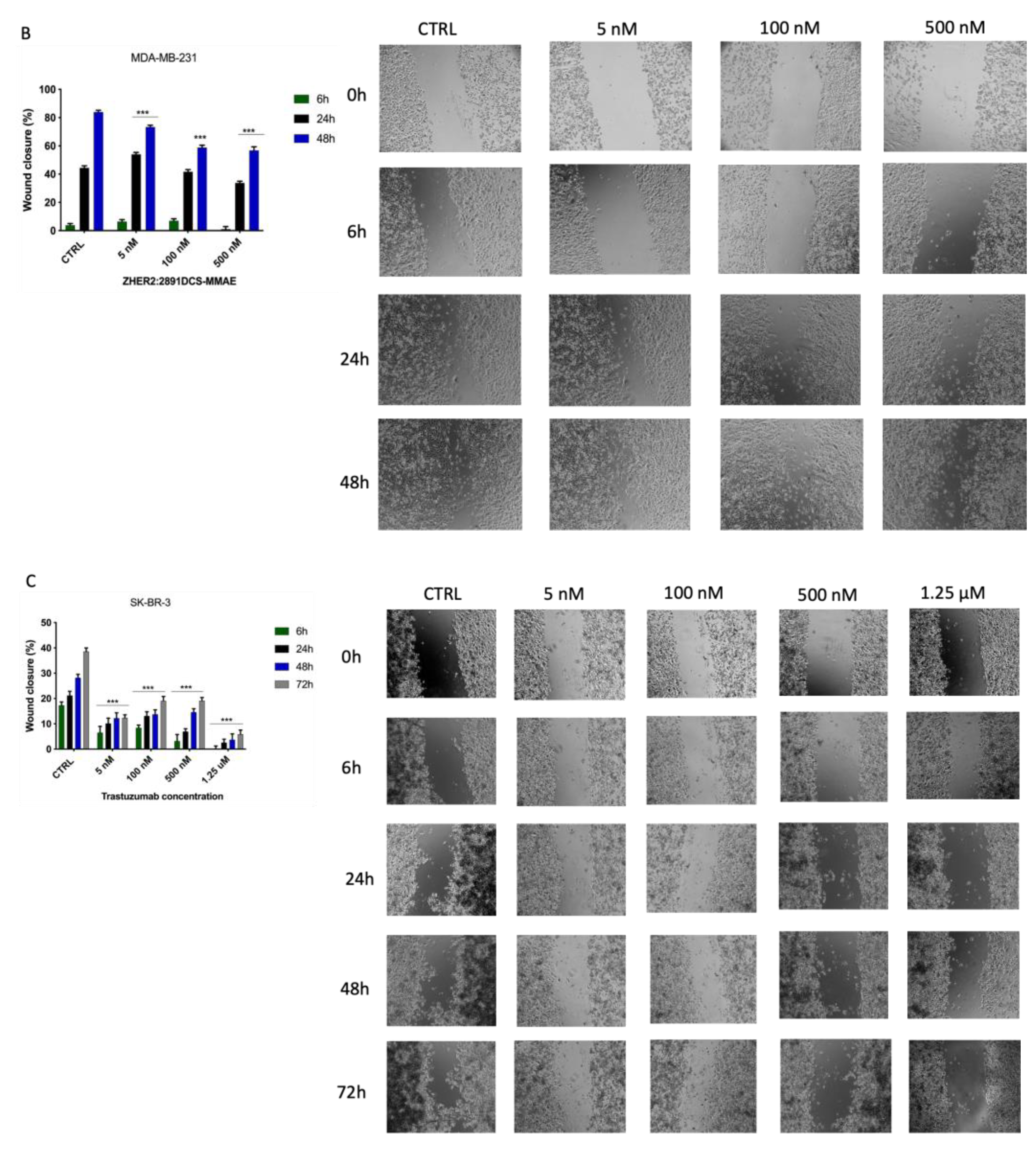
3.5. ZHER2:2891DCS-MMAE Inhibits SK-BR-3 Migration
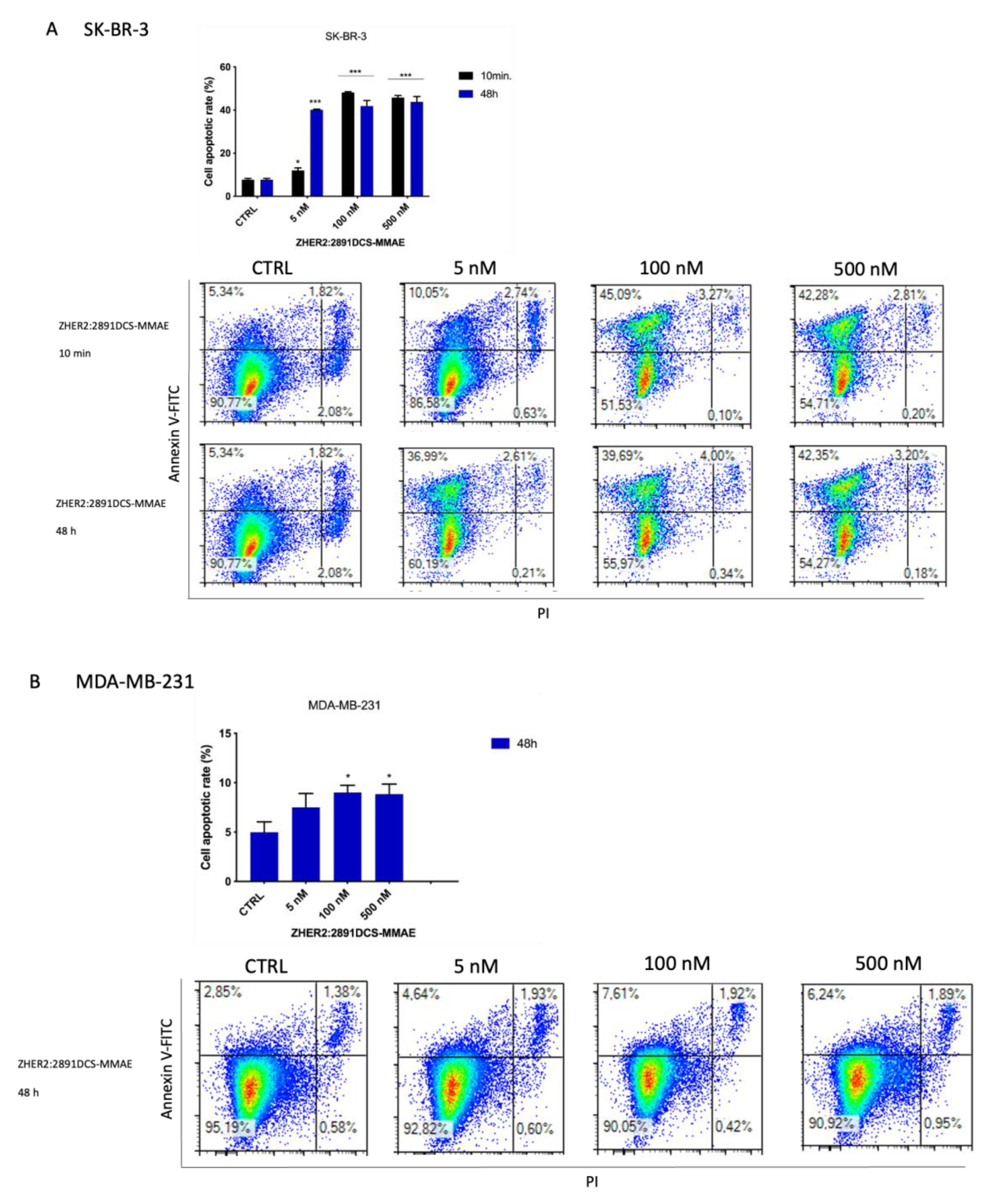
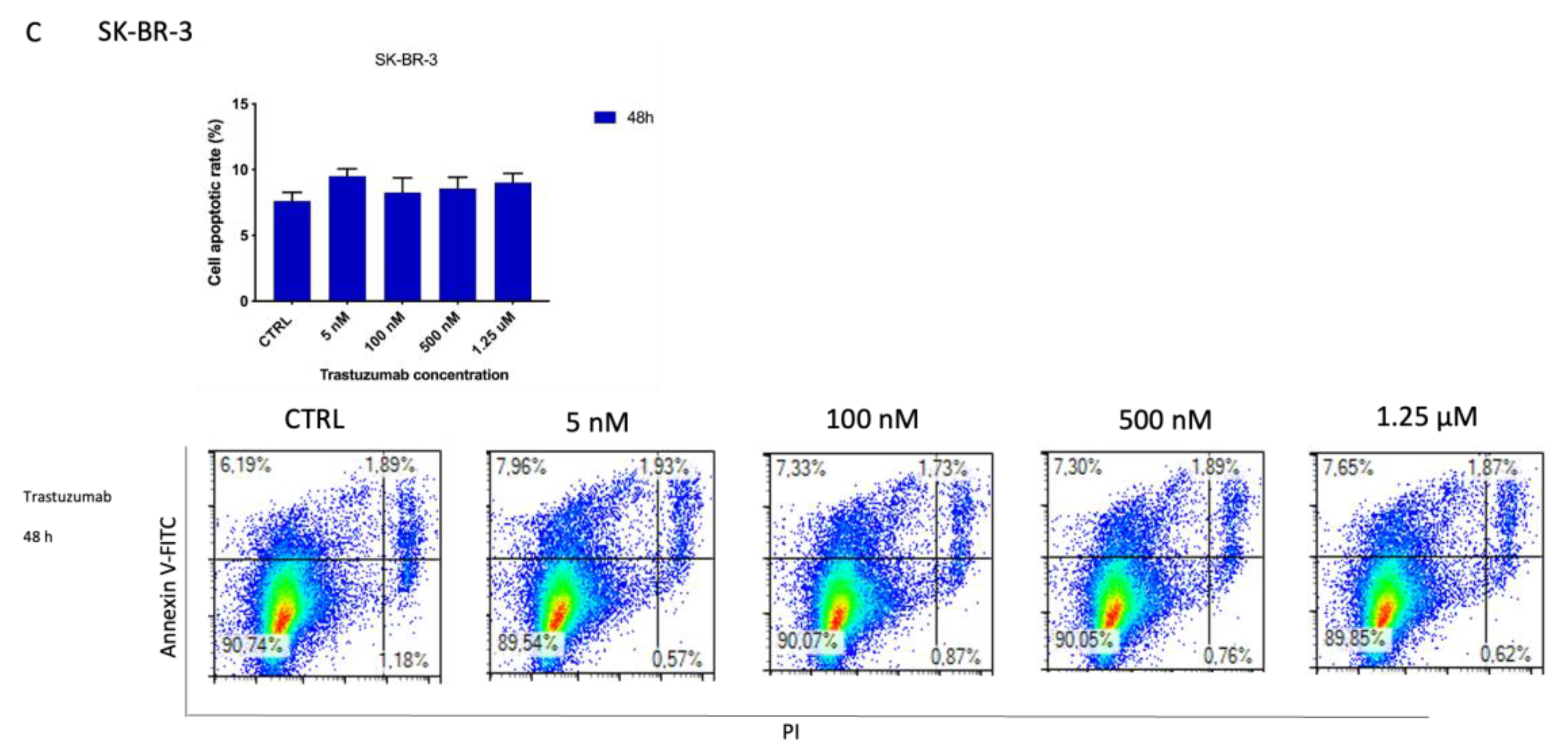
3.6. ZHER2:2891DCS-MMAE Induces Apoptosis of SK-BR-3 Cells
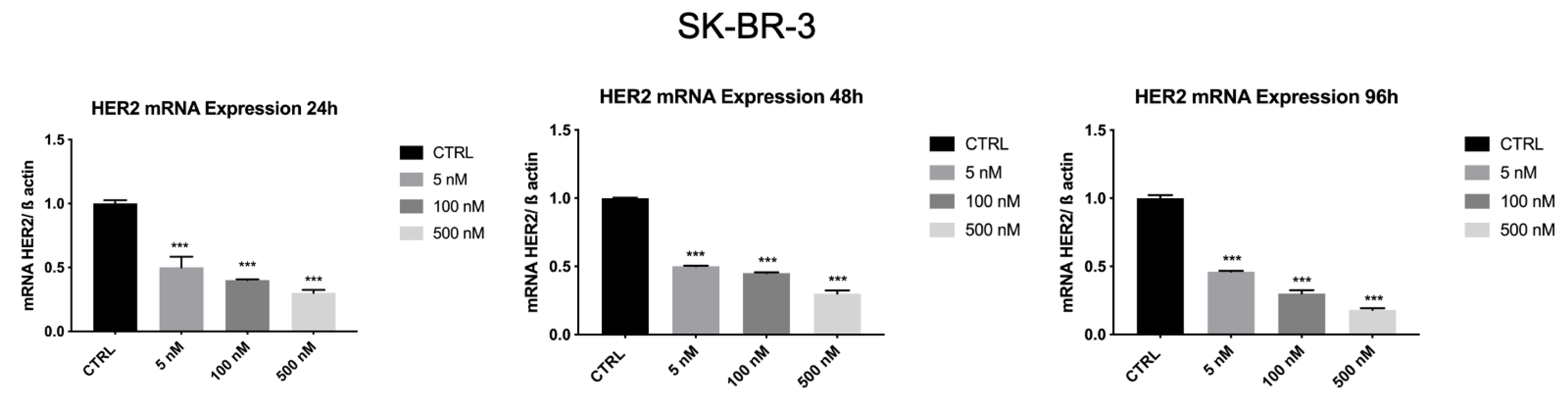
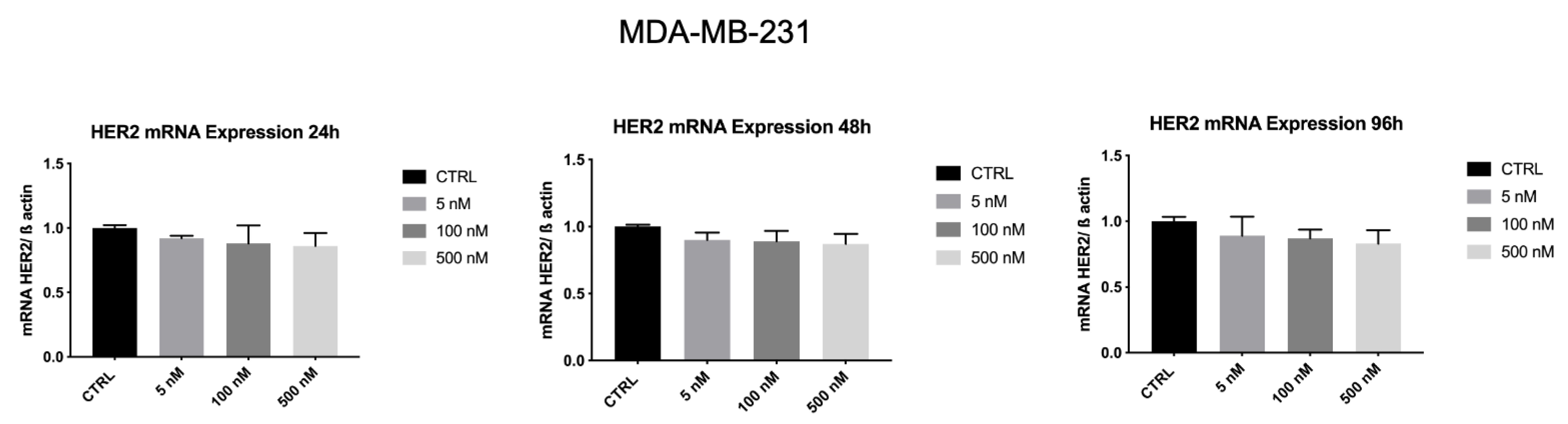
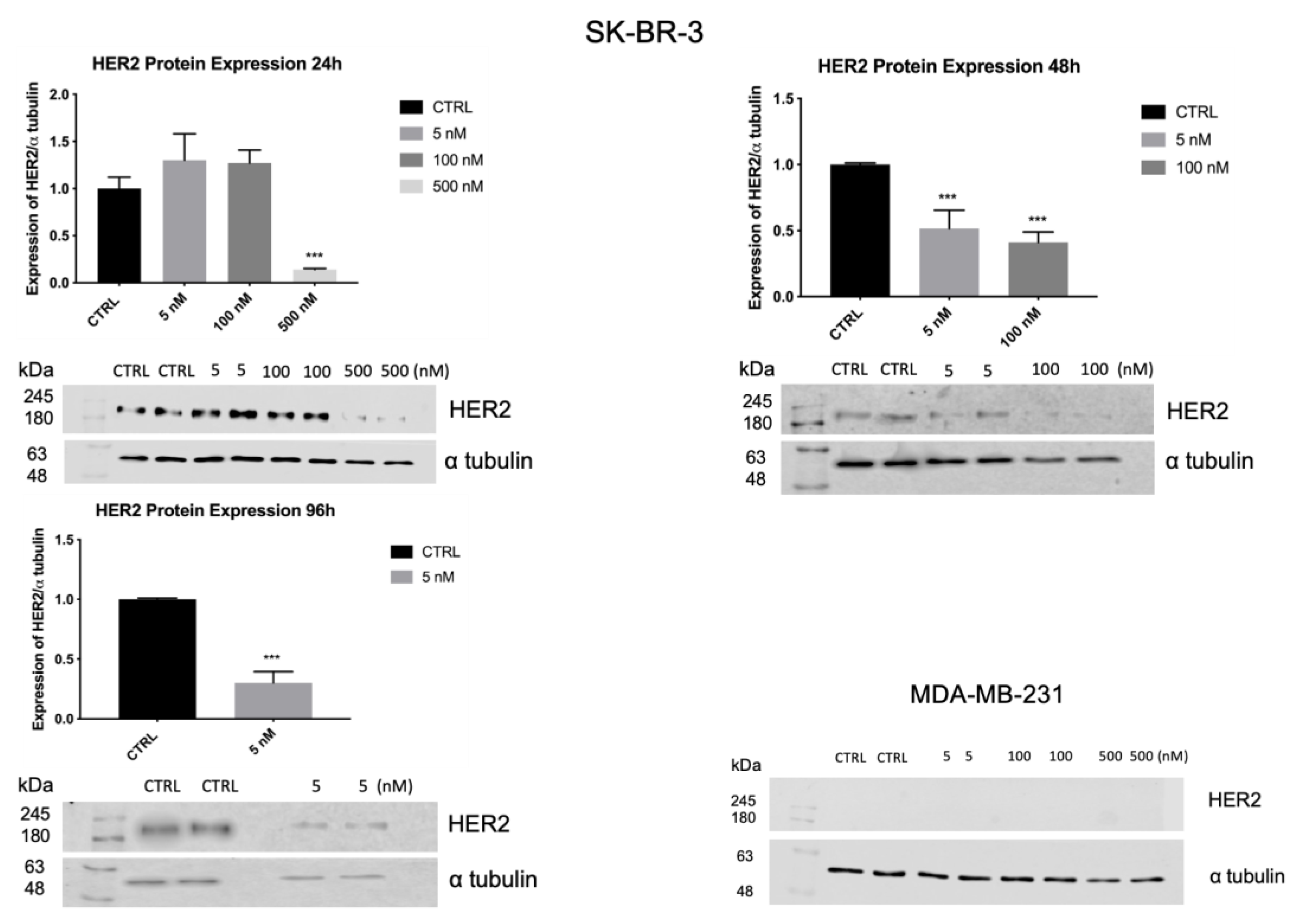
3.7. ZHER2:2891DCS-MMAE Reduces HER2 Expression by SK-BR-3 Cells
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Slamon, D.J.; Clark, G.M.; Wong, S.G.; Levin, W.J.; Ullrich, A.; McGuire, W.L. Human Breast Cancer: Correlation of Relapse and Survival with Amplification of the HER-2/neu Oncogene. Science 1987, 235, 177–182. [Google Scholar] [CrossRef] [Green Version]
- Soares, M.; Ribeiro, R.; Najmudin, S.; Gameiro, A.; Rodrigues, R.V.; Cardoso, F.; Ferreira, F. Serum HER2 Levels are Increased in Cats with Mammary Carcinomas and Predict Tissue HER2 Status. Oncotarget 2016, 7, 17314–17326. [Google Scholar] [CrossRef] [Green Version]
- Chen, J.-S.; Lan, K.; Hung, M.-C. Strategies to Target HER2/neu Overexpression for Cancer Therapy. Drug Resist. Updates 2003, 6, 129–136. [Google Scholar] [CrossRef]
- Marmor, M.D.; Skaria, K.B.; Yarden, Y. Signal Transduction and Oncogenesis by ErbB/HER Receptors. Int. J. Radiat. Oncol. Biol. 2004, 58, 903–913. [Google Scholar] [CrossRef]
- Okines, A.F.; Cunningham, D. Trastuzumab: A Novel Standard Option for Patients with HER-2-Positive Advanced Gastric or Gastro-Oesophageal Junction Cancer. Ther. Adv. Gastroenterol. 2012, 5, 301–318. [Google Scholar] [CrossRef] [Green Version]
- Collins, D.M.; O’Donovan, N.; McGowan, P.; O’Sullivan, F.; Duffy, M.J.; Crown, J. Trastuzumab Induces Antibody-Dependent Cell-Mediated Cytotoxicity (ADCC) in HER-2-Non-Amplified Breast Cancer Cell Lines. Ann. Oncol. 2012, 23, 1788–1795. [Google Scholar] [CrossRef]
- Junttila, T.T.; Akita, R.W.; Parsons, K.; Fields, C.; Phillips, G.D.L.; Friedman, L.S.; Sampath, D.; Sliwkowski, M.X. Ligand-Independent HER2/HER3/PI3K Complex Is Disrupted by Trastuzumab and Is Effectively Inhibited by the PI3K Inhibitor GDC-0941. Cancer Cell 2009, 15, 429–440. [Google Scholar] [CrossRef] [Green Version]
- Molina, M.A.; Codony-Servat, J.; Albanell, J.; Rojo, F.; Arribas, J.; Baselga, J. Trastuzumab (herceptin), a Humanized Anti-Her2 Receptor Monoclonal Antibody, Inhibits Basal and Activated Her2 Ectodomain Cleavage in Breast Cancer Cells. Cancer Res. 2001, 61, 4744–4749. [Google Scholar] [PubMed]
- Slamon, D.J.; Leyland-Jones, B.; Shak, S.; Fuchs, H.; Paton, V.; Bajamonde, A.; Fleming, T.; Eiermann, W.; Wolter, J.; Pegram, M.; et al. Use of Chemotherapy plus a Monoclonal Antibody against HER2 for Metastatic Breast Cancer that Overexpresses HER2. N. Engl. J. Med. 2001, 344, 783–792. [Google Scholar] [CrossRef]
- Gajria, D.; Chandarlapaty, S. HER2-Amplified breast Cancer: Mechanisms of Trastuzumab Resistance and Novel Targeted Therapies. Expert Rev. Anticancer Ther. 2011, 11, 263–275. [Google Scholar] [CrossRef] [Green Version]
- Panowski, S.; Bhakta, S.; Raab, H.; Polakis, P.; Junutula, J.R. Site-Specific Antibody Drug Conjugates for Cancer Therapy. mAbs 2014, 6, 34–45. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beck, A.; Reichert, J.M. Antibody-Drug Conjugates. mAbs 2013, 6, 15–17. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Senter, P.D.; Sievers, E. The Discovery and Development of Brentuximab Vedotin for Use in Relapsed Hodgkin Lymphoma and Systemic Anaplastic Large Cell Lymphoma. Nat. Biotechnol. 2012, 30, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Phillips, G.D.L.; Li, G.; Dugger, D.L.; Crocker, L.M.; Parsons, K.L.; Mai, E.; Blättler, W.A.; Lambert, J.M.; Chari, R.V.; Lutz, R.J.; et al. Targeting HER2-Positive Breast Cancer with Trastuzumab-DM1, an Antibody–Cytotoxic Drug Conjugate. Cancer Res. 2008, 68, 9280–9290. [Google Scholar] [CrossRef] [Green Version]
- Gameiro, A.; Nascimento, C.; Correia, J.; Ferreira, F. HER2-Targeted Immunotherapy and Combined Protocols Showed Promising Antiproliferative Effects in Feline Mammary Carcinoma Cell-Based Models. Cancers 2021, 13, 2007. [Google Scholar] [CrossRef]
- Chames, P.; Van Regenmortel, M.; Weiss, E.; Baty, D. Therapeutic Antibodies: Successes, Limitations and Hopes for the Future. Br. J. Pharmacol. 2009, 157, 220–233. [Google Scholar] [CrossRef]
- Löfblom, J.; Feldwisch, J.; Tolmachev, V.; Carlsson, J.; Ståhl, S.; Frejd, F.Y. Affibody molecules: Engineered Proteins for Therapeutic, Diagnostic and Biotechnological Applications. FEBS Lett. 2010, 584, 2670–2680. [Google Scholar] [CrossRef] [Green Version]
- Nilsson, B.; Moks, T.; Jansson, B.; Abrahmsén, L.; Elmblad, A.; Holmgren, E.; Henrichson, C.; Jones, T.A.; Uhlen, M. A Synthetic IgG-Binding Domain Based on Staphylococcal Protein A. Protein Eng. Des. Sel. 1987, 1, 107–113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wikman, M.; Steffen, A.-C.; Gunneriusson, E.; Tolmachev, V.; Adams, G.; Carlsson, J.; Stahl, S. Selection and characterization of HER2/neu-Binding Affibody Ligands. Protein Eng. Des. Sel. 2004, 17, 455–462. [Google Scholar] [CrossRef] [Green Version]
- Nygren, P.Å. Alternative Binding Proteins: Affibody Binding Proteins Developed from a Small Three-Helix Bundle Scaffold. FEBS J. 2008, 275, 2668–2676. [Google Scholar] [CrossRef]
- Sochaj-Gregorczyk, A.M.; Serwotka-Suszczak, A.M.; Otlewski, J. A Novel Affibody-Auristatin E Conjugate With a Potent and Selective Activity against HER2+ Cell Lines. J. Immunother. 2016, 39, 223–232. [Google Scholar] [CrossRef]
- Ma, D.; Hopf, C.E.; Malewicz, A.D.; Donovan, G.P.; Senter, P.D.; Goeckeler, W.F.; Maddon, P.J.; Olson, W.C. Potent Antitumor Activity of an Auristatin-Conjugated, Fully Human Monoclonal Antibody to Prostate-Specific Membrane Antigen. Clin. Cancer Res. 2006, 12, 2591–2596. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pettit, G.R.; Kamano, Y.; Herald, C.L.; Tuinman, A.A.; Boettner, F.E.; Kizu, H.; Schmidt, J.M.; Baczynskyj, L.; Tomer, K.B.; Bontems, R.J. The Isolation and Structure of a Remarkable Marine Animal Antineoplastic Constituent: Dolastatin 10. J. Am. Chem. Soc. 1987, 109, 6883–6885. [Google Scholar] [CrossRef]
- Doronina, S.O.; Toki, B.E.; Torgov, M.Y.; Mendelsohn, B.A.; Cerveny, C.G.; Chace, D.F.; Deblanc, R.L.; Gearing, R.P.; Bovee, T.D.; Siegall, C.B.; et al. Development of Potent Monoclonal Antibody Auristatin Conjugates for Cancer Therapy. Nat. Biotechnol. 2003, 21, 778–784, Erratum in 2003, 21, 941. [Google Scholar]
- Yip, V.; Lee, M.; Saad, O.; Ma, S.; Khojasteh, S.; Shen, B.-Q. Preclinical Characterization of the Distribution, Catabolism, and Elimination of a Polatuzumab Vedotin-Piiq (POLIVY®) Antibody–Drug Conjugate in Sprague Dawley Rats. J. Clin. Med. 2021, 10, 1323. [Google Scholar] [CrossRef]
- Cancer, H.; Lines, C.; Abdollahpour-alitappeh, M.; Razavi-vakhshourpour, S.; Lotfinia, M.; Jahandideh, S. Monomethyl Auristatin E Exhibits Potent Cytotoxic Activity against Human Cancer Cell Lines SKBR3 and HEK293. Nov. Biomed. 2017, 5, 145–151. [Google Scholar] [CrossRef]
- Yamaguchi, Y.; Hironaka, K.; Okawaki, M.; Okita, R.; Matsuura, K.; Ohshita, A.; Toge, T. HER2-Specific Cytotoxic Activity of Lymphokine-Activated Killer Cells in the Presence of Trastuzumab. Anticancer Res. 2005, 25, 827–832. [Google Scholar] [PubMed]
- Magnifico, A.; Albano, L.; Campaner, S.; Delia, D.; Castiglioni, F.; Gasparini, P.; Sozzi, G.; Fontanella, E.; Menard, S.; Tagliabue, E. Tumor-Initiating Cells of HER2-Positive Carcinoma Cell Lines Express the Highest Oncoprotein Levels and Are Sensitive to Trastuzumab. Clin. Cancer Res. 2009, 15, 2010–2021. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdollahpour-Alitappeh, M.; Lotfinia, M.; Gharibi, T.; Mardaneh, J.; Farhadihosseinabadi, B.; Larki, P.; Faghfourian, B.; Sepehr, K.S.; Abbaszadeh-Goudarzi, K.; Johari, B.; et al. Antibody-Drug Conjugates (ADCs) for Cancer Therapy: Strategies, Challenges, and Successes. J. Cell. Physiol. 2019, 234, 5628–5642. [Google Scholar] [CrossRef]
- Junutula, J.R.; Raab, H.; Clark, S.; Bhakta, S.; Leipold, D.D.; Weir, S.; Chen, Y.; Simpson, M.; Tsai, S.P.; Dennis, M.S.; et al. Site-specific Conjugation of a Cytotoxic Drug to an Antibody Improves the Therapeutic Index. Nat. Biotechnol. 2008, 26, 925–932. [Google Scholar] [CrossRef]
- Wagh, A.; Song, H.; Zeng, M.; Tao, L.; Das, T.K. Challenges and New Frontiers in Analytical Characterization of Antibody-Drug Conjugates. mAbs 2018, 10, 222–243. [Google Scholar] [CrossRef]
- Carlsson, J.; Nordberg, E.; Ekerljung, L.; Sahlberg, S.H.; Lennartsson, J.; Glimelius, B. Effects of an EGFR-Binding Affibody Molecule on Intracellular Signaling Pathways. Int. J. Oncol. 2010, 36, 967–972. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Lundberg, E.; Vernet, E.; Larsson, B.; Höidén-Guthenberg, I.; Gräslund, T. Selection of Affibody Molecules to the Ligand Binding Site of the Insulin-Like Growth Factor-1 Receptor. Biotechnol. Appl. Biochem. 2010, 55, 99–109. [Google Scholar] [CrossRef]
- Eigenbrot, C.; Ultsch, M.; Dubnovitsky, A.; Abrahmsén, L.; Härd, T. Structural Basis for High-Affinity HER2 Receptor Binding by an Engineered Protein. Proc. Natl. Acad. Sci. USA 2010, 107, 15039–15044. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Orlova, A.; Tran, T.; Widström, C.; Engfeldt, T.; Karlström, A.E.; Tolmachev, V. Pre-Clinical Evaluation of [111In]-Benzyl-DOTA-ZHER2:342, a Potential Agent for Imaging of HER2 Expression in Malignant Tumors. Int. J. Mol. Med. 2007, 20, 397–404. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zielinski, R.; Lyakhov, I.; Hassan, M.; Kuban, M.; Shafer-Weaver, K.; Gandjbakhche, A.; Capala, J. HER2-Affitoxin: A Potent Therapeutic Agent for the Treatment of HER2-Overexpressing Tumors. Clin. Cancer Res. 2011, 17, 5071–5081. [Google Scholar] [CrossRef] [Green Version]
- Xu, T.; Ding, H.; Vorobyeva, A.; Oroujeni, M.; Orlova, A.; Tolmachev, V.; Gräslund, T. Drug Conjugates Based on a Monovalent Affibody Targeting Vector Can Efficiently Eradicate HER2 Positive Human Tumors in an Experimental Mouse Model. Cancers 2020, 13, 85. [Google Scholar] [CrossRef]
- Lin, T.-Y.; Wang, P.-W.; Huang, C.-H.; Yang, P.-M.; Pan, T.-L. Characterizing the Relapse Potential in Different Luminal Subtypes of Breast Cancers with Functional Proteomics. Int. J. Mol. Sci. 2020, 21, 6077. [Google Scholar] [CrossRef]
- Subik, K.; Lee, J.-F.; Baxter, L.; Strzepek, T.; Costello, D.; Crowley, P.; Xing, L.; Hung, M.-C.; Bonfiglio, T.; Hicks, D.G.; et al. The Expression Patterns of ER, PR, HER2, CK5/6, EGFR, Ki-67 and AR by Immunohistochemical Analysis in Breast Cancer Cell Lines. Breast Cancer Basic Clin. Res. 2010, 4, 35–41. [Google Scholar] [CrossRef]
- Abdollahpour-Alitappeh, M.; Lotfinia, M.; Bagheri, N.; Sepehr, K.S.; Habibi-Anbouhi, M.; Kobarfard, F.; Balalaie, S.; Foroumadi, A.; Abbaszadeh-Goudarzi, K.; Abolhassani, M. Trastuzumab-Monomethyl Auristatin E Conjugate Exhibits Potent Cytotoxic Activity In Vitro against HER2-Positive Human Breast Cancer. J. Cell. Physiol. 2019, 234, 2693–2704. [Google Scholar] [CrossRef] [PubMed]
- Emlet, D.R.; Brown, K.A.; Kociban, D.L.; Pollice, A.A.; Smith, C.A.; Ong, B.B.L.; Shackney, S.E. Response to Trastuzumab, Erlotinib, and Bevacizumab, Alone and in Combination, is Correlated with the Level of Human Epidermal Growth Factor Receptor-2 Expression in Human Breast Cancer Cell Lines. Mol. Cancer Ther. 2007, 6, 2664–2674. [Google Scholar] [CrossRef] [PubMed] [Green Version]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Forward Primer | Reverse Primer |
|---|---|
| 5′ GCCGAAGCCAAATATGCA 3′ | 5′ CATATGTATATCTCCTTC 3′ |
| 3′ CGGCTTCGGTTTATACGT 5′ | 3′ GTATACATATAGAGGAAG 5′ |
| Target Gene | Forward Primer | Reverse Primer |
|---|---|---|
| βactin | AGTGTGACGTGGACATCCGCA | GCCAGGGCAGTGATCTCCTTCT |
| HER2 | CCAGGACCTGCTGAACTGGT | GTACGAGCCGCACATCC |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Damiani, I.; Castiglioni, S.; Sochaj-Gregorczyk, A.; Bonacina, F.; Colombo, I.; Rusconi, V.; Otlewski, J.; Corsini, A.; Bellosta, S. Purification and In Vitro Evaluation of an Anti-HER2 Affibody-Monomethyl Auristatin E Conjugate in HER2-Positive Cancer Cells. Biology 2021, 10, 758. https://doi.org/10.3390/biology10080758
Damiani I, Castiglioni S, Sochaj-Gregorczyk A, Bonacina F, Colombo I, Rusconi V, Otlewski J, Corsini A, Bellosta S. Purification and In Vitro Evaluation of an Anti-HER2 Affibody-Monomethyl Auristatin E Conjugate in HER2-Positive Cancer Cells. Biology. 2021; 10(8):758. https://doi.org/10.3390/biology10080758
Chicago/Turabian StyleDamiani, Isabella, Silvia Castiglioni, Alicja Sochaj-Gregorczyk, Fabrizia Bonacina, Irma Colombo, Valentina Rusconi, Jacek Otlewski, Alberto Corsini, and Stefano Bellosta. 2021. "Purification and In Vitro Evaluation of an Anti-HER2 Affibody-Monomethyl Auristatin E Conjugate in HER2-Positive Cancer Cells" Biology 10, no. 8: 758. https://doi.org/10.3390/biology10080758