Body Size Variation in Italian Lesser Horseshoe Bats Rhinolophus hipposideros over 147 Years: Exploring the Effects of Climate Change, Urbanization and Geography

 ,
,  , and
, and 
Abstract
:Simple Summary
Abstract
1. Introduction
- (1)
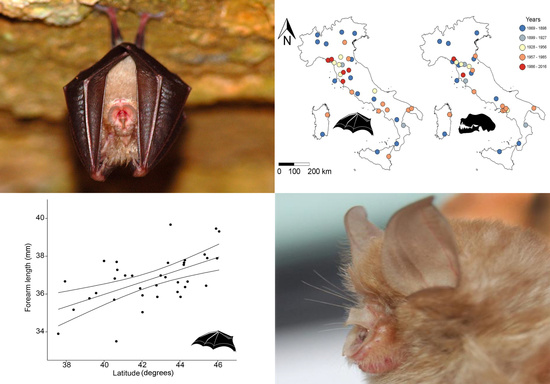
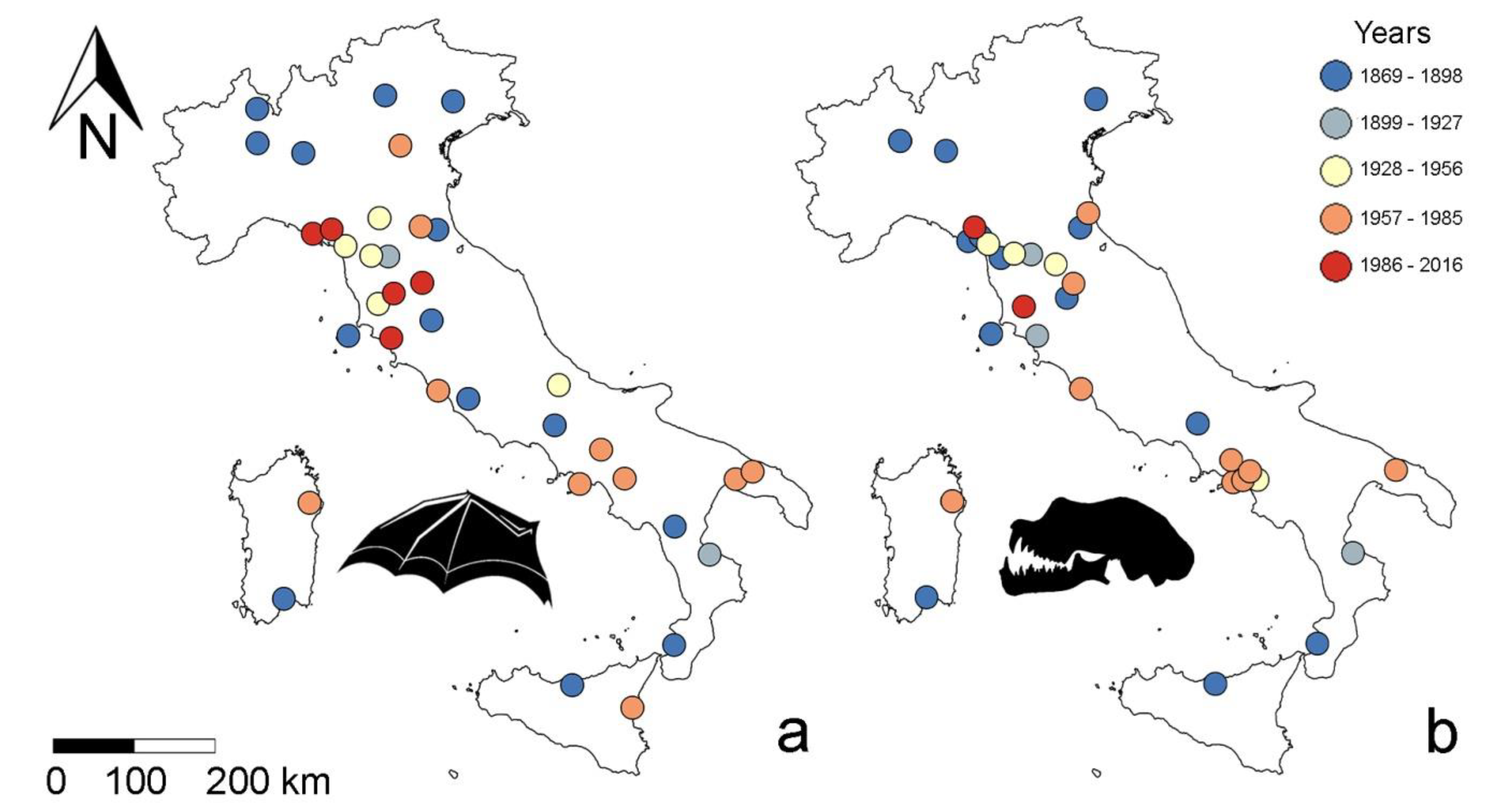
- Skull size and forearm length (hereafter FAL) of R. hipposideros will either decrease (to dissipate heat more effectively) or increase (to reduce the risk of dehydration) in response to a warming climate, so we predict an effect of the year of collection, which we adopt as a proxy for climate change as done in many other studies (see [27] for a review).
- (2)
- Rhinolophus hipposideros skull size will not increase selectively over time, that is, there will be no influence of artificial illumination on this light-averse species, in contrast to light-exploiting species [25]. This rhinolophid is, in fact, an ideal control taxon in this context because it shows strong adverse reactions to artificial illumination [28].
- (3)
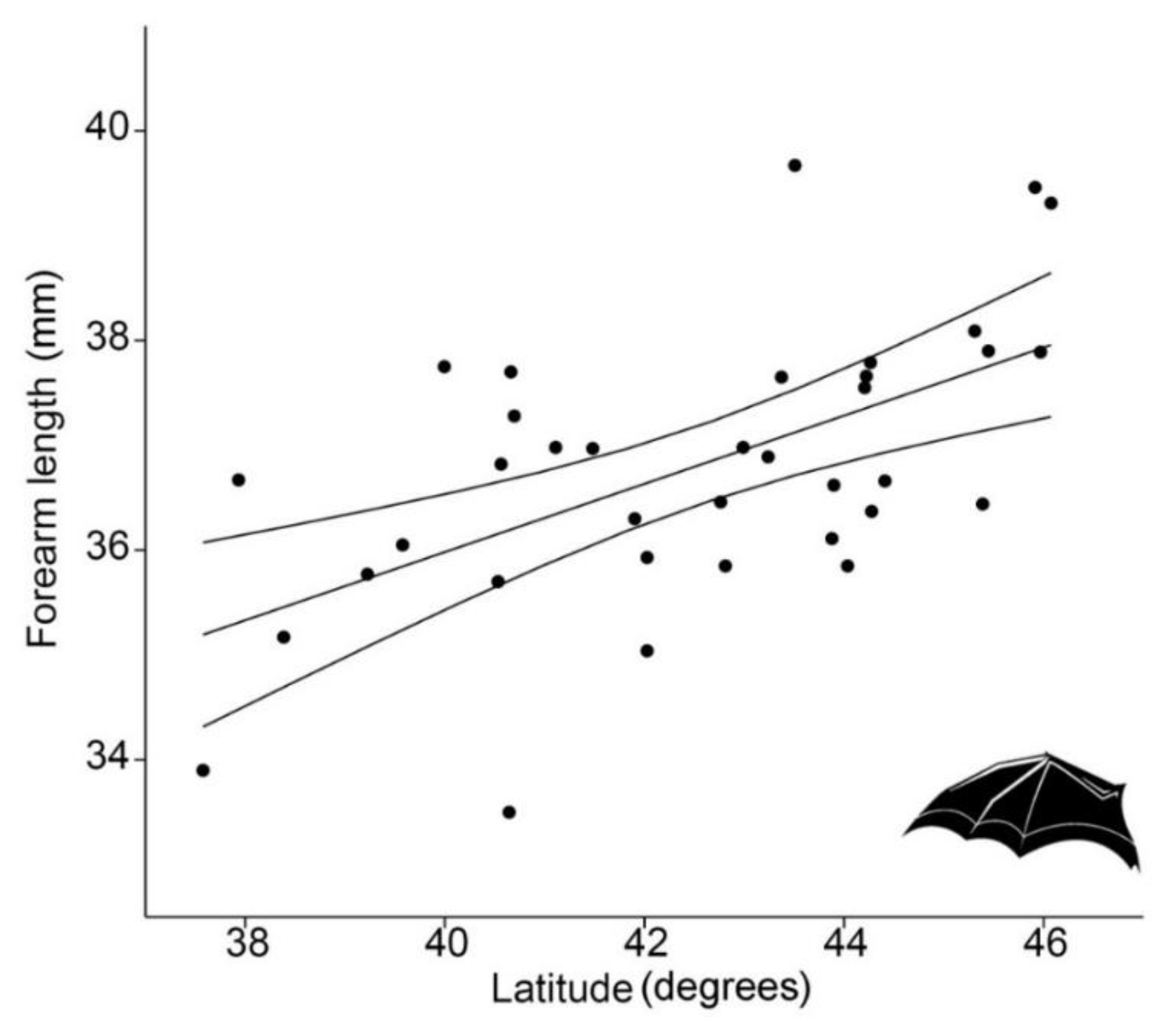
- Finally, we formulate a set of geographic-framed hypotheses, i.e., that body size may respond to latitude, longitude, elevation or insularity. Specific predictions arising from such hypotheses are that: (a) Body size will increase at higher latitudes, in agreement with Bergmann’s rule, as seen in other bats [29]; and (b) bats from islands (Sardinia and Sicily) will be smaller (dwarfism) or larger (giantism) than their conspecifics from the mainland (insularity syndrome, [30]).
2. Materials and Methods
2.1. Data Collection
2.2. Data Analysis
3. Results
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Walther, G.R. Community and ecosystem responses to recent climate change. Philos. Trans. R. Soc. B Biol. Sci. 2010, 365, 2019–2024. [Google Scholar] [CrossRef]
- Perry, A.L.; Low, P.J.; Ellis, J.R.; Reynolds, J.D. Climate change and distribution shifts in marine fishes. Science 2005, 308, 1912–1915. [Google Scholar] [CrossRef] [PubMed]
- Kullberg, C.; Fransson, T.; Hedlund, J.; Jonzén, N.; Langvall, O.; Nilsson, J.; Bolmgren, K. Change in spring arrival of migratory birds under an era of climate change, Swedish data from the last 140 years. Ambio 2015, 44, 69–77. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Visser, M.E.; Both, C.; Lambrechts, M.M. Global climate change leads to mistimed avian reproduction. Adv. Ecol. Res. 2004, 35, 89–110. [Google Scholar]
- Lane, J.E.; Kruuk, L.E.; Charmantier, A.; Murie, J.O.; Dobson, F.S. Delayed phenology and reduced fitness associated with climate change in a wild hibernator. Nature 2012, 489, 554–557. [Google Scholar] [CrossRef] [PubMed]
- Sheridan, J.A.; Bickford, D. Shrinking body size as an ecological response to climate change. Nat. Clim. Change 2011, 1, 401–406. [Google Scholar] [CrossRef]
- Meiri, S.; Dayan, T. On the validity of Bergmann’s rule. J. Biogeogr. 2003, 30, 331–351. [Google Scholar] [CrossRef]
- Gardner, J.L.; Peters, A.; Kearney, M.R.; Joseph, L.; Heinsohn, R. Declining body size: A third universal response to warming? Trends Ecol. Evol. 2011, 26, 285–291. [Google Scholar] [CrossRef]
- Stawski, C.; Willis, C.K.R.; Geiser, F. The importance of temporal heterothermy in bats. J. Zool. 2014, 292, 86–100. [Google Scholar] [CrossRef] [Green Version]
- Adams, R.A. Bat reproduction declines when conditions mimic climate change projections for western North America. Ecology 2010, 91, 2437–2445. [Google Scholar] [CrossRef]
- Arbuthnott, D.; Brigham, R.M. The influence of a local temperature inversion on the foraging behaviour of big brown bats, Eptesicus fuscus. Acta Chiropt. 2007, 9, 193–201. [Google Scholar] [CrossRef]
- Russo, D.; Bosso, L.; Ancillotto, L. Novel perspectives on bat insectivory highlight the value of this ecosystem service in farmland: Research frontiers and management implications. Agric. Ecosyst. Environ. 2018, 266, 31–38. [Google Scholar] [CrossRef]
- O’Shea, T.J.; Cryan, P.M.; Hayman, D.T.; Plowright, R.K.; Streicker, D.G. Multiple mortality events in bats: A global review. Mamm. Rev. 2016, 46, 175–190. [Google Scholar] [CrossRef]
- Wang, M.; Chen, K.; Guo, D.; Luo, B.; Wang, W.; Gao, H.; Liu, Y.; Feng, J. Ambient temperature correlates with geographic variation in body size of least horseshoe bats. Curr. Zool. 2020, 66, 459–465. [Google Scholar] [CrossRef] [Green Version]
- Yue, X.; Hughes, A.C.; Tomlinson, K.W.; Xia, S.; Li, S.; Chen, J. Body size and diet–related morphological variation of bats over the past 65 years in China. J. Mammal. 2020, 101, 61–79. [Google Scholar] [CrossRef]
- Wolf, T.; Menne, B.; World Health Organization. Environment and Health Risks from Climate Change and Variability in Italy (No. EUR/07/5046244); WHO Regional Office for Europe: Copenhagen, Denmark, 2007. [Google Scholar]
- Spano, D.; Mereu, V.; Bacciu, V.; Marras, S.; Trabucco, A.; Adinolf, M.; Barbato, G.; Bosello, F.; Breil, M.; Chiriacò, M.V.; et al. Analisi del rischio. I cambiamenti climatici in Italia. 2020. [Google Scholar] [CrossRef]
- McNab, B.K. Geographic and temporal correlations of mammalian size reconsidered: A resource rule. Oecologia 2010, 164, 13–23. [Google Scholar] [CrossRef]
- Yom-Tov, Y.; Yom-Tov, S.; Moller, H. Competition, coexistence, and adaptation amongst rodent invaders to Pacific and New Zealand islands. J. Biogeogr. 1999, 26, 947–958. [Google Scholar] [CrossRef] [Green Version]
- Phillips, B.L.; Shine, R. Adapting to an invasive species: Toxic cane toads induce morphological change in Australian snakes. Proc. Natl. Acad. Sci. USA 2004, 101, 17150–17155. [Google Scholar] [CrossRef] [Green Version]
- Ancillotto, L.; Santini, L.; Ranc, N.; Maiorano, L.; Russo, D. Extraordinary range expansion in a common bat: The potential roles of climate change and urbanisation. Sci. Nat. 2016, 103, 15. [Google Scholar] [CrossRef]
- Stone, E.L.; Harris, S.; Jones, G. Impacts of artificial lighting on bats: A review of challenges and solutions. Mamm. Biol. 2015, 80, 213–219. [Google Scholar] [CrossRef]
- Polak, T.; Korine, C.; Yair, S.; Holderied, M.W. Differential effects of artificial lighting on flight and foraging behaviour of two sympatric bat species in a desert. J. Zool. 2011, 285, 21–27. [Google Scholar] [CrossRef]
- Serangeli, M.T.; Cistrone, L.; Ancillotto, L.; Tomassini, A.; Russo, D. The post-release fate of hand-reared orphaned bats: Survival and habitat selection. Anim. Welf. 2012, 21, 9. [Google Scholar] [CrossRef] [Green Version]
- Tomassini, A.; Colangelo, P.; Agnelli, P.; Jones, G.; Russo, D. Cranial size has increased over 133 years in a common bat, Pipistrellus kuhlii: A response to changing climate or urbanization? J. Biogeogr. 2014, 41, 944–953. [Google Scholar] [CrossRef]
- Minnaar, C.; Boyles, J.G.; Minnaar, I.A.; Sole, C.L.; McKechnie, A.E. Stacking the odds: Light pollution may shift the balance in an ancient predator-prey arms race. J. Appl. Ecol. 2014, 52, 522–531. [Google Scholar] [CrossRef] [Green Version]
- Yom-Tov, Y.; Geffen, E. Recent spatial and temporal changes in body size of terrestrial vertebrates: Probable causes and pitfalls. Biol. Rev. 2011, 86, 531–541. [Google Scholar] [CrossRef]
- Stone, E.L.; Jones, G.; Harris, S. Street lighting disturbs commuting bats. Curr. Biol. 2009, 19, 1123–1127. [Google Scholar] [CrossRef] [Green Version]
- Ashton, K.G.; Tracy, M.C.; Queiroz, A.D. Is Bergmann’s rule valid for mammals? Am. Nat. 2000, 156, 390–415. [Google Scholar] [CrossRef]
- van Valen, L. Body size and numbers of plants and animals. Evolution 1973, 27, 27–35. [Google Scholar] [CrossRef] [Green Version]
- Lindenfors, P.; Gittleman, J.L.; Jones, K.E. Sexual Size Dimorphism in Mammals. In Sex, Size and Gender Roles: Evolutionary Studies of Sexual Size Dimorphism; Fairbairn, D.J., Blanckenhorn, W.U., Szekely, T., Eds.; Oxford University Press: New York, NY, USA, 2007; pp. 19–26. [Google Scholar]
- Russo, D.; Mucedda, M.; Bello, M.; Biscardi, S.; Pidinchedda, E.; Jones, G. Divergent echolocation call frequencies in insular rhinolophids (Chiroptera): A case of character displacement? J. Biogeogr. 2007, 34, 2129–2138. [Google Scholar] [CrossRef]
- Kunz, T.H. Reproduction, growth, and mortality of the vespertilionid bat, Eptesicus fuscus, in Kansas. J. Mammal. 1974, 55, 1–13. [Google Scholar] [CrossRef] [PubMed]
- Richards, L.R.; Taylor, P.J.; Schoeman, M.C.; Goodman, S.M.; Van Daele, P.A.A.G.; Lamb, J.M. Cranial size and shape variation in Afrotropical Otomops (Mammalia: Chiroptera: Molossidae): Testing species limits using a morphometric approach. Biol. J. Linn. Soc. 2012, 106, 910–925. [Google Scholar] [CrossRef]
- Azil, A.H.; Bruce, D.; Williams, C.R. Determining the spatial autocorrelation of dengue vector populations: Influences of mosquito sampling method, covariables, and vector control. J. Vector Ecol. 2014, 39, 153–163. [Google Scholar] [CrossRef] [PubMed]
- Smeraldo, S.; Bosso, L.; Fraissinet, M.; Bordignon, L.; Brunelli, M.; Ancillotto, L.; Russo, D. Modelling risks posed by wind turbines and power lines to soaring birds: The black stork (Ciconia nigra) in Italy as a case study. Biodivers. Conserv. 2020, 29, 1959–1976. [Google Scholar] [CrossRef]
- Ancillotto, L.; Bosso, L.; Smeraldo, S.; Mori, E.; Mazza, G.; Herkt, M.; Galimberti, A.; Ramazzotti, F.; Russo, D. An African bat in Europe, Plecotus gaisleri: Biogeographic and ecological insights from molecular taxonomy and Species Distribution Models. Ecol. Evol. 2020, 10, 5785–5800. [Google Scholar] [CrossRef] [PubMed]
- Dytham, C. Choosing and Using Statistics: A Biologist’s Guide; John Wiley & Sons: Hoboken, NJ, USA, 2011; p. 264. [Google Scholar]
- Yom-Tov, Y.; Yom-Tov, S.; Jarrell, G. Recent increase in body size of the American marten Martes americana in Alaska. Biol. J. Linn. Soc. 2008, 93, 701–707. [Google Scholar] [CrossRef] [Green Version]
- Aldasoro, M.; Garin, I.; Vallejo, N.; Baroja, U.; Arrizabalaga-Escudero, A.; Goiti, U.; Aihartza, J. Gaining ecological insight on dietary allocation among horseshoe bats through molecular primer combination. PLoS ONE 2019, 14, e0220081. [Google Scholar] [CrossRef] [Green Version]
- Arlettaz, R.; Godat, S.; Meyer, H. Competition for food by expanding pipistrelle bat populations (Pipistrellus pipistrellus) might contribute to the decline of lesser horseshoe bats (Rhinolophus hipposideros). Biol. Conserv. 2000, 93, 55–60. [Google Scholar] [CrossRef]
- Baroja, U.; Garin, I.; Aihartza, J.; Arrizabalaga-Escudero, A.; Vallejo, N.; Aldasoro, M.; Goiti, U. Pest consumption in a vineyard system by the lesser horseshoe bat (Rhinolophus hipposideros). PLoS ONE 2019, 14, e0219265. [Google Scholar] [CrossRef] [Green Version]
- Afonso, E.; Tournant, P.; Foltête, J.C.; Giraudoux, P.; Baurand, P.E.; Roué, S.; Canella, V.; Vey, D.; Scheifler, R. Is the lesser horseshoe bat (Rhinolophus hipposideros) exposed to causes that may have contributed to its decline? A non-invasive approach. Glob. Ecol. Conserv. 2016, 8, 123–137. [Google Scholar] [CrossRef]
- Van der Meij, T.; Van Strien, A.J.; Haysom, K.A.; Dekker, J.; Russ, J.; Biala, K.; Canella, V.; Vey, D.; Scheifler, R. Return of the bats? A prototype indicator of trends in European bat populations in underground hibernacula. Mamm. Biol. 2015, 80, 170–177. [Google Scholar] [CrossRef]
- Rondinini, C.; Battistoni, A.; Peronace, V.; Teofili, C. per il Volume: Lista Rossa IUCN dei Vertebrati Italiani; Comitato Italiano IUCN e Ministero dell’Ambiente e della Tutela del Territorio e del Mare: Roma, Italy, 2013. [Google Scholar]
- Russo, D.; Ancillotto, L. Sensitivity of bats to urbanization: A review. Mamm. Biol. 2015, 80, 205–212. [Google Scholar] [CrossRef] [PubMed]
- Snell-Rood, E.C.; Wick, N. Anthropogenic environments exert variable selection on cranial capacity in mammals. Proc. R. Soc. Lond. B Biol. Sci. 2013, 280, 20131384. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Santini, L.; González-Suárez, M.; Russo, D.; Gonzalez-Voyer, A.; von Hardenberg, A.; Ancillotto, L. One strategy does not fit all: Determinants of urban adaptation in mammals. Ecol. Lett. 2019, 22, 365–376. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reiter, G.; Pölzer, E.; Mixanig, H.; Bontadina, F.; Hüttmeir, U. Impact of landscape fragmentation on a specialised woodland bat, Rhinolophus hipposideros. Mamm. Biol. 2013, 78, 283–289. [Google Scholar] [CrossRef]
- Carrete, M.; Tella, J.L. Inter-individual variability in fear of humans and relative brain size of the species are related to contemporary urban invasion in birds. PLoS ONE 2011, 6, e18859. [Google Scholar] [CrossRef] [Green Version]
- Møller, A.P.; Erritzøe, J. Brain size and urbanization in birds. Avian Res. 2015, 6, 8. [Google Scholar] [CrossRef] [Green Version]
- Mayr, E. Animal Species and Evolution; Belknap Press: Cambridge, MA, USA, 1963; p. 797. [Google Scholar]
- Janis, C.M. Correlation of cranial and dental variables with body size in ungulates and macropodoids. In Body Size in Mammalian Paleobiology: Estimation and Biological Implications; Damuth, J.D., Damuth, J., MacFadden, B.J., John, D., Eds.; Cambridge University Press: Cambridge, UK, 1990; pp. 255–299. [Google Scholar]
- Arbour, J.H.; Curtis, A.A.; Santana, S.E. Signatures of echolocation and dietary ecology in the adaptive evolution of skull shape in bats. Nat. Commun. 2019, 10, 1–13. [Google Scholar] [CrossRef] [Green Version]
- Andreone, F.; Bartolozzi, L.; Boano, G.; Boero, F.; Bologna, M.A.; Bon, M.; Bressi, N.; Capula, M.; Casale, A.; Casiraghi, M.; et al. Italian natural history museums on the verge of collapse? ZooKeys 2014, 456, 139. [Google Scholar] [CrossRef]



{kind=link}
{kind=link}
{kind=link}
| Source | d.f. | F | p |
|---|---|---|---|
| Latitude (decimal degrees) | 1 | 4.27 | <0.05 |
| Longitude (decimal degrees) | 1 | 0.02 | n.s. |
| Altitude (m a.s.l.) | 1 | 0.98 | n.s. |
| Year of collection | 1 | 0.29 | n.s. |
| Sex | 1 | 5.95 | <0.05 |
| Mainland vs. Island | 1 | 0.08 | n.s. |
| Error | 29 | ||
| Total | 35 |
| Cranial Variable | PC1 | PC2 | PC3 | PC4 |
|---|---|---|---|---|
| Greatest length of skull (GSL) | −0.536 | −0.030 | 0.143 | −0.141 |
| Condylobasal length (CBL) | −0.490 | −0.070 | 0.178 | 0.174 |
| Mastoid breadth (MB) | −0.398 | −0.477 | 0.116 | −0.095 |
| Cranial depth (CRD) | −0.488 | 0.188 | −0.089 | 0.295 |
| Width of upper incisors (I2I2) | −0.227 | 0.168 | −0.917 | −0.094 |
| Length from the craniomandibular joint to the origin of the masseter muscle (A) | −0.150 | 0.494 | 0.200 | −0.793 |
| Length from the craniomandibular joint to the insertion of the masseter muscle (B) | 0.052 | −0.678 | −0.213 | −0.465 |
| Variable | Wilk’s Statistic | d.f. | F | p |
|---|---|---|---|---|
| Latitude (decimal degrees) | 0.646 | 4, 21 | 2.873 | <0.05 |
| Longitude (decimal degrees) | 0.778 | 4, 21 | 1.497 | n.s. |
| Altitude (m a.s.l.) | 0.823 | 4, 21 | 1.127 | n.s. |
| Year of collection | 0.785 | 4, 21 | 1.438 | n.s. |
| Sex | 0.888 | 8, 42 | 0.319 | n.s. |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations |
© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Salinas-Ramos, V.B.; Agnelli, P.; Bosso, L.; Ancillotto, L.; Sánchez-Cordero, V.; Russo, D. Body Size Variation in Italian Lesser Horseshoe Bats Rhinolophus hipposideros over 147 Years: Exploring the Effects of Climate Change, Urbanization and Geography. Biology 2021, 10, 16. https://doi.org/10.3390/biology10010016
Salinas-Ramos VB, Agnelli P, Bosso L, Ancillotto L, Sánchez-Cordero V, Russo D. Body Size Variation in Italian Lesser Horseshoe Bats Rhinolophus hipposideros over 147 Years: Exploring the Effects of Climate Change, Urbanization and Geography. Biology. 2021; 10(1):16. https://doi.org/10.3390/biology10010016
Chicago/Turabian StyleSalinas-Ramos, Valeria B., Paolo Agnelli, Luciano Bosso, Leonardo Ancillotto, Víctor Sánchez-Cordero, and Danilo Russo. 2021. "Body Size Variation in Italian Lesser Horseshoe Bats Rhinolophus hipposideros over 147 Years: Exploring the Effects of Climate Change, Urbanization and Geography" Biology 10, no. 1: 16. https://doi.org/10.3390/biology10010016