
An In Vivo Platform for Rebuilding Functional Neocortical Tissue
 ,
,
Abstract
:1. Introduction
2. Methods
2.1. Animals
2.2. Fetal Telencephalon Dissociation
2.3. Transplantation Procedures
2.4. Tissue Processing and Immunohistochemistry
2.5. Image Analysis
2.6. In Vivo Live Imaging
2.7. Perfusion of Blood Vessels
2.8. Headpost and Chronic Electrode Implantation Surgery
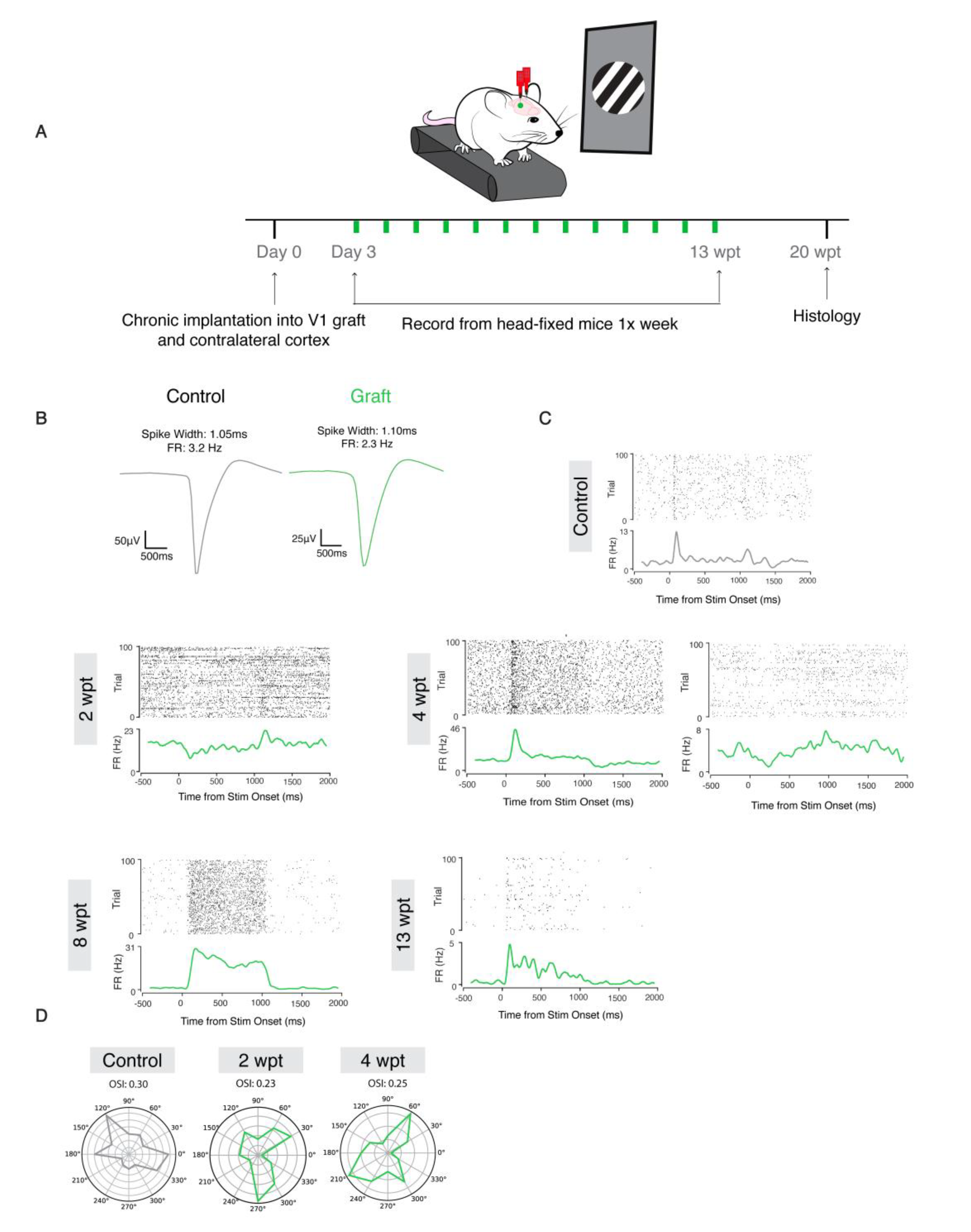
2.9. In Vivo Electrophysiology
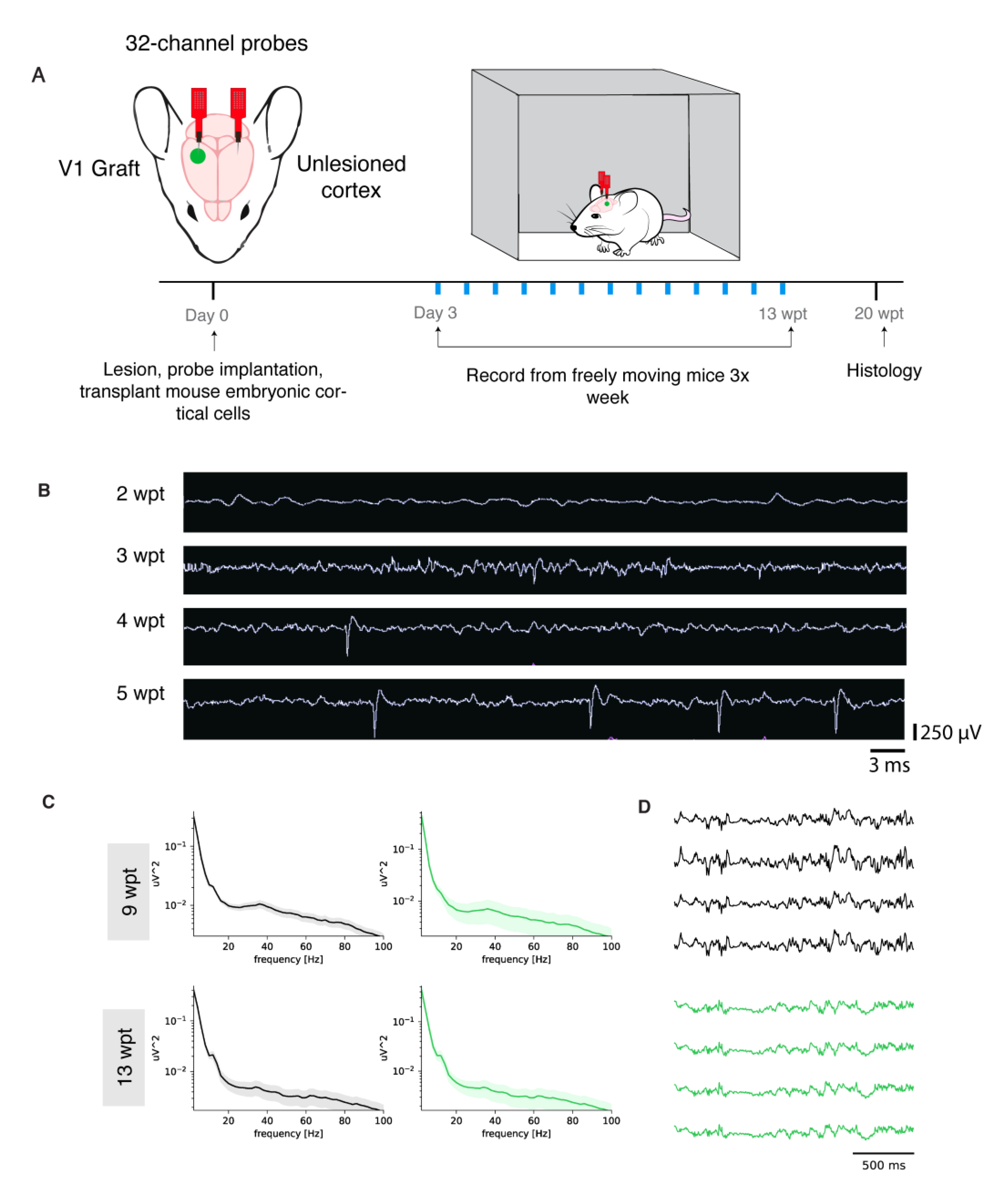
2.10. In Vivo Freely-Moving Electrophysiology Recordings
2.11. Visual Stimulation
2.12. In Vivo Electrophysiology Analysis
2.13. Single Unit Activity Analysis in Head Fixed Recordings
2.14. Local Field Potential Computation in Freely-Moving Recordings
2.15. Statistical Analyses
3. Results
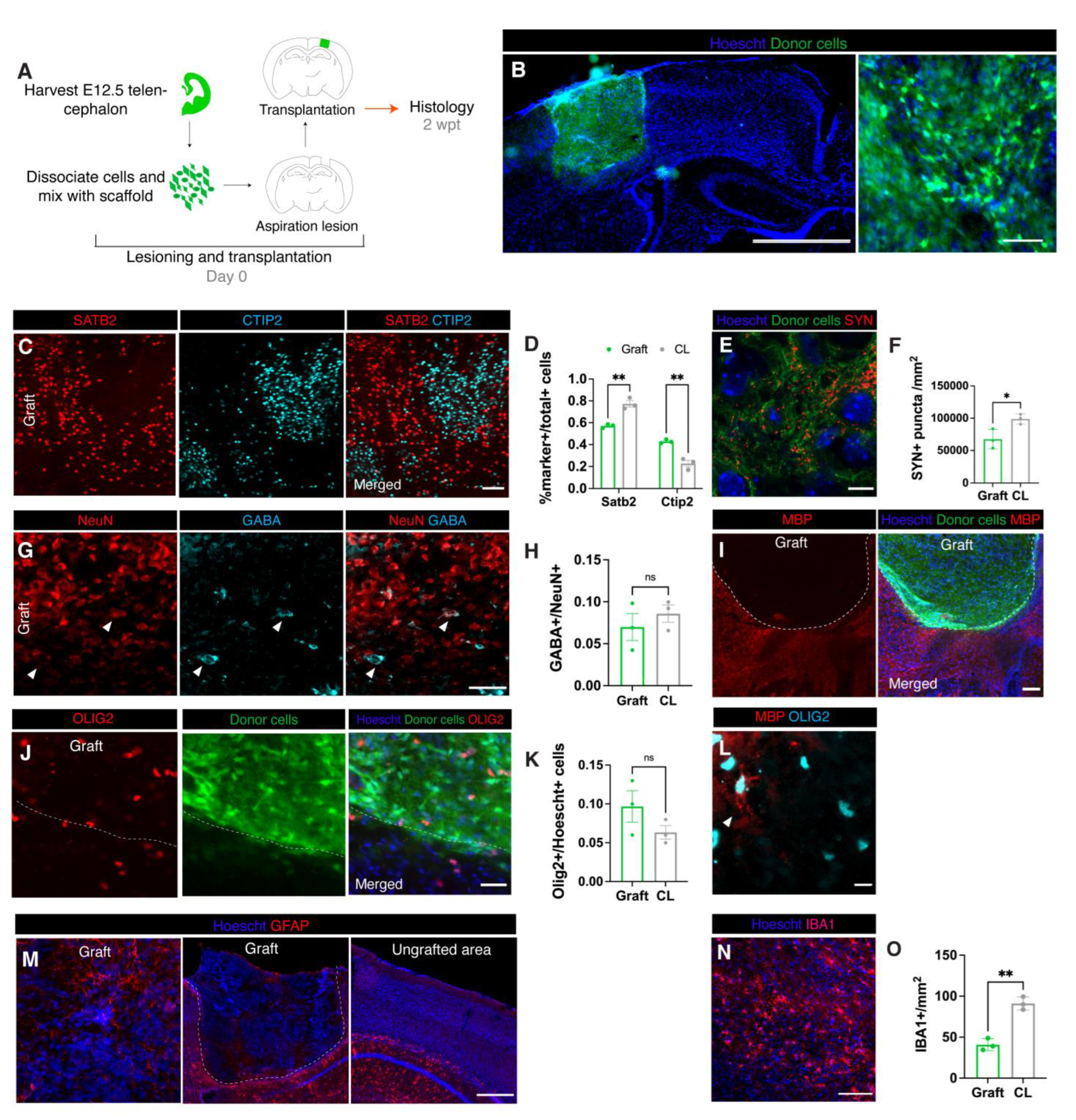
3.1. Graft Integrity Is Dependent on Scaffold Dilution
3.2. Donor Neural Precursor Cells Survive and Differentiate into Several Cortical Cell Types
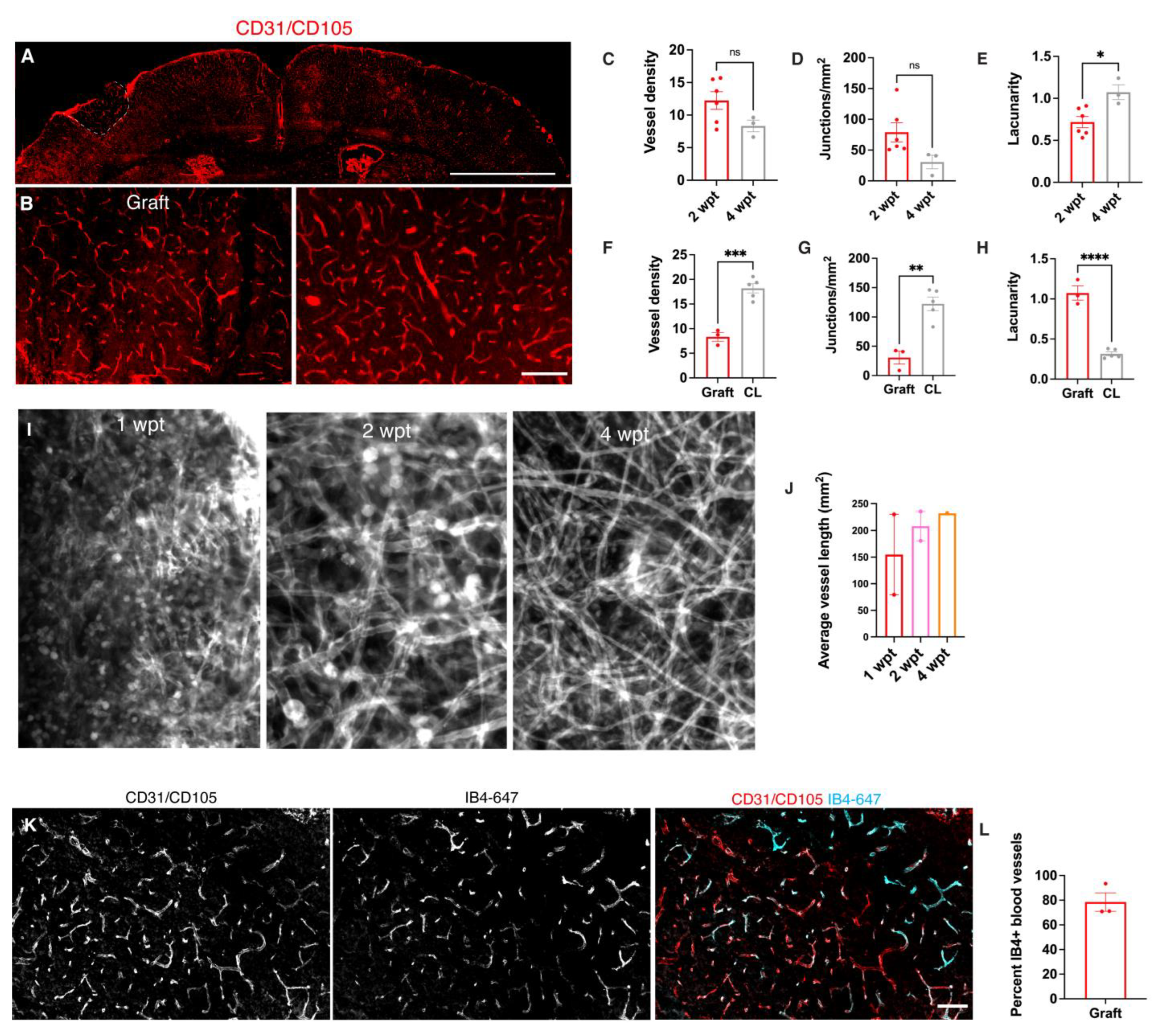
3.3. Transplants Become Vascularized with Vessels and Perfused with Blood
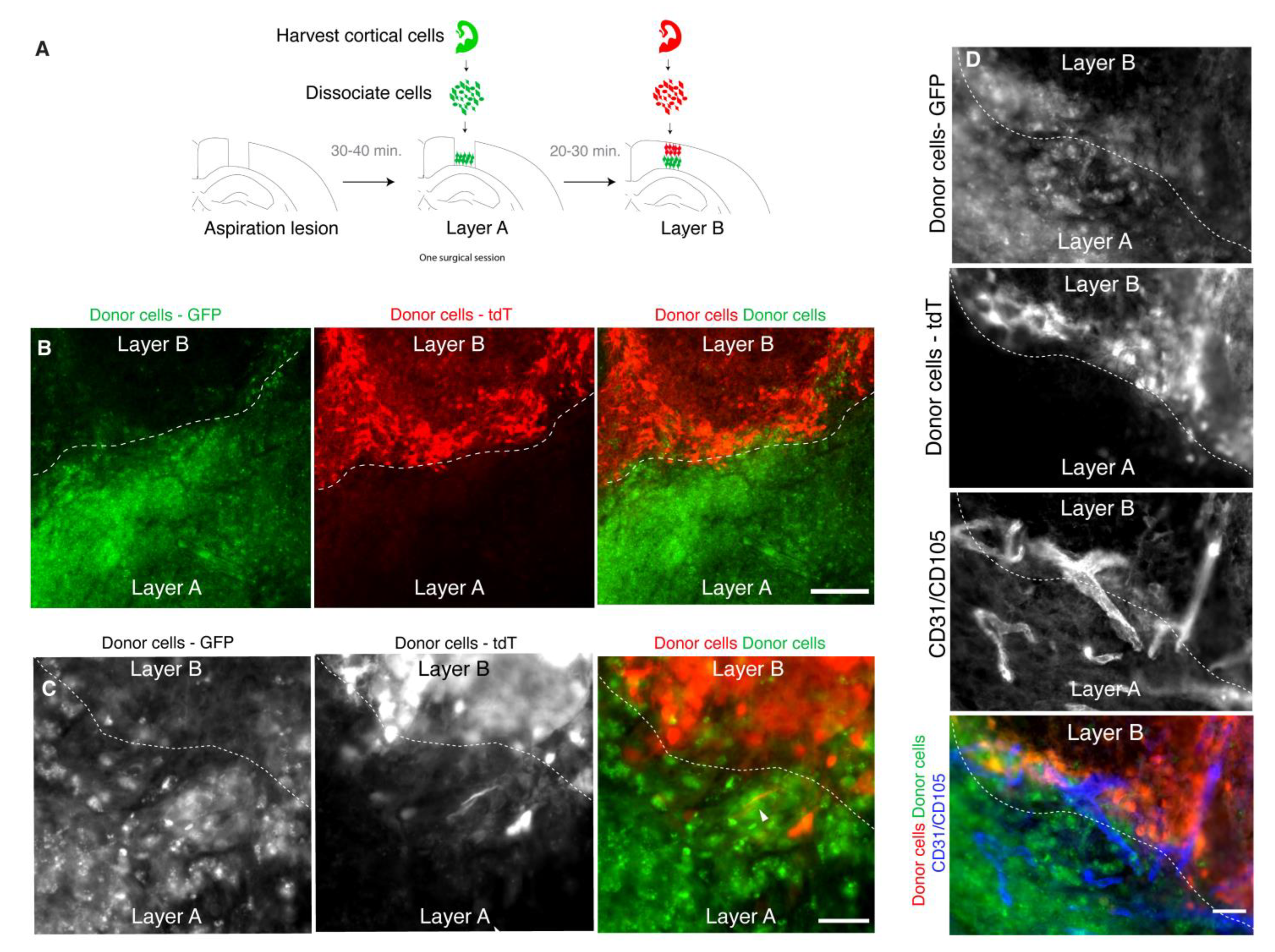
3.4. Donor Cells Can Be Layered in Lesion Sites
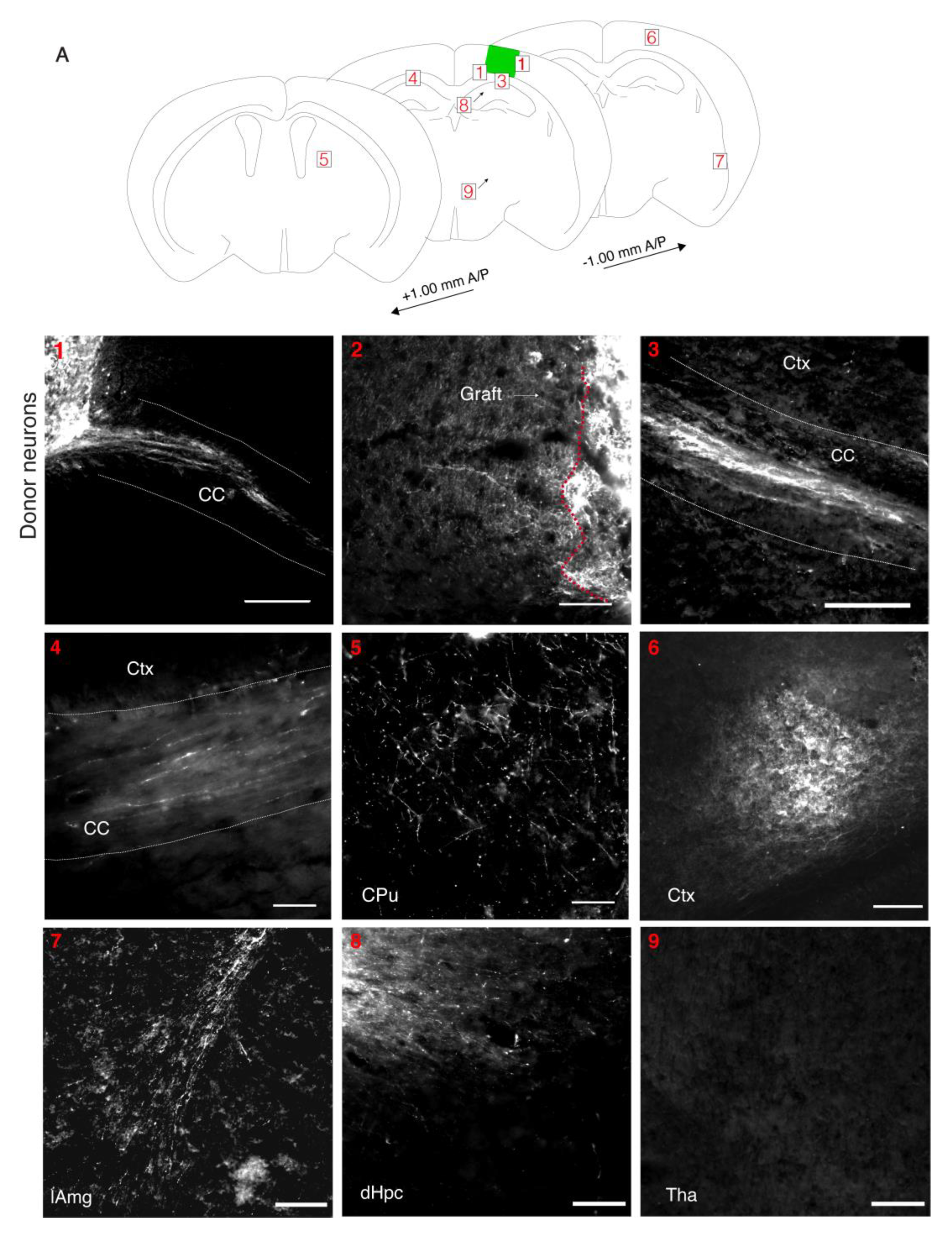
3.5. Donor Neurons Project to Appropriate Targets in Host Brains
3.6. Transplanted Neurons Become Electrophysiologically Active and Respond to Visual Stimuli
4. Discussion
Supplementary Materials
Author Contributions
Funding
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Linaro, D.; Vermaercke, B.; Iwata, R.; Ramaswamy, A.; Libé-Philippot, B.; Boubakar, L.; Davis, B.A.; Wierda, K.; Davie, K.; Poovathingal, S.; et al. Xenotransplanted Human Cortical Neurons Reveal Species-Specific Development and Functional Integration into Mouse Visual Circuits. Neuron 2019, 104, 972–986.e6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Espuny-Camacho, I.; Michelsen, K.A.; Linaro, D.; Bilheu, A.; Acosta-Verdugo, S.; Herpoel, A.; Giugliano, M.; Gaillard, A.; Vanderhaeghen, P. Human Pluripotent Stem-Cell-Derived Cortical Neurons Integrate Functionally into the Lesioned Adult Murine Visual Cortex in an Area-Specific Way. Cell Rep. 2018, 23, 2732–2743. [Google Scholar] [CrossRef] [Green Version]
- Falkner, S.; Grade, S.; Dimou, L.; Conzelmann, K.K.; Bonhoeffer, T.; Götz, M.; Hübener, M. Transplanted embryonic neurons integrate into adult neocortical circuits. Nature 2016, 539, 248–253. [Google Scholar] [CrossRef] [PubMed]
- Mansour, A.A.; Gonçalves, J.T.; Bloyd, C.W.; Li, H.; Fernandes, S.; Quang, D.; Johnston, S.; Parylak, S.L.; Jin, X.; Gage, F.H. An in vivo model of functional and vascularized human brain organoids. Nat. Biotechnol. 2018, 36, 432–441. [Google Scholar] [CrossRef] [PubMed]
- Michelsen, K.A.; Acosta-Verdugo, S.; Benoit-Marand, M.; Espuny-Camacho, I.; Gaspard, N.; Saha, B.; Gaillard, A.; Vanderhaeghen, P. Area-specific reestablishment of damaged circuits in the adult cerebral cortex by cortical neurons derived from mouse embryonic stem cells. Neuron 2015, 85, 982–997. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palma-Tortosa, S.; Tornero, D.; Hansen, M.G.; Monni, E.; Hajy, M.; Kartsivadze, S.; Aktay, S.; Tsupykov, O.; Parmar, M.; Deisseroth, K.; et al. Activity in grafted human iPS cell–derived cortical neurons integrated in stroke-injured rat brain regulates motor behavior. Proc. Natl. Acad. Sci. USA 2020, 117, 9094–9100. [Google Scholar] [CrossRef]
- Tornero, D.; Tsupykov, O.; Granmo, M.; Rodriguez, C.; Grønning-Hansen, M.; Thelin, J.; Smozhanik, E.; Laterza, C.; Wattananit, S.; Ge, R.; et al. Synaptic inputs from stroke-injured brain to grafted human stem cell-derived neurons activated by sensory stimuli. Brain 2017, 140, 692–706. [Google Scholar] [CrossRef] [Green Version]
- Henriques, D.; Moreira, R.; Schwamborn, J.; Pereira de Almeida, L.; Mendonça, L.S. Successes and Hurdles in Stem Cells Application and Production for Brain Transplantation. Front. Neurosci. 2019, 13, 1194. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.P.; Jeyakumar, M.; Gonzalez, R.; Takahashi, H.; Lee, P.J.; Baek, R.C.; Clark, D.; Rose, H.; Fu, G.; Clarke, J.; et al. Stem cells act through multiple mechanisms to benefit mice with neurodegenerative metabolic disease. Nat. Med. 2007, 13, 439–447. [Google Scholar] [CrossRef]
- Pham, M.T.; Pollock, K.M.; Rose, M.D.; Cary, W.A.; Stewart, H.R.; Zhou, P.; Nolta, J.A.; Waldau, B. Generation of human vascularized brain organoids. Neuroreport 2018, 29, 588–593. [Google Scholar] [CrossRef]
- Dong, X.; Xu, S.B.; Chen, X.; Tao, M.; Tang, X.Y.; Fang, K.H.; Xu, M.; Pan, Y.; Chen, Y.; He, S.; et al. Human cerebral organoids establish subcortical projections in the mouse brain after transplantation. Mol. Psychiatry 2021, 26, 2964–2976. [Google Scholar] [CrossRef]
- Saga, Y.; Miyagawa-Tomita, S.; Takagi, A.; Kitajima, S.; Miyazaki, J.I.; Inoue, T. MesP1 is expressed in the heart precursor cells and required for the formation of a single heart tube. Development 1999, 126, 3437–3447. [Google Scholar] [CrossRef] [PubMed]
- Zudaire, E.; Gambardella, L.; Kurcz, C.; Vermeren, S. A computational tool for quantitative analysis of vascular networks. PLoS ONE 2011, 6, e27385. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Schneider, C.A.; Rasband, W.S.; Eliceiri, K.W. NIH Image to ImageJ: 25 years of image analysis. Nat. Methods 2012, 9, 671–675. [Google Scholar] [CrossRef] [PubMed]
- Luan, L.; Wei, X.; Zhao, Z.; Siegel, J.J.; Potnis, O.; Tuppen, C.A.; Lin, S.; Kazmi, S.; Fowler, R.A.; Holloway, S.; et al. Ultraflexible nanoelectronic probes form reliable, glial scar–free neural integration. Sci. Adv. 2017, 3, e1601966. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yin, R.; Noble, B.C.; He, F.; Zolotavin, P.; Rathore, H.; Jin, Y.; Sevilla, N.; Xie, C.; Luan, L. Chronic co-implantation of ultraflexible neural electrodes and a cranial window. Neurophotonics 2022, 9, 032204. [Google Scholar] [CrossRef] [PubMed]
- Pachitariu, M.; Steinmetz, N.; Kadir, S.; Carandini, M.; Harris, K. Fast and accurate spike sorting of high-channel count probes with KiloSort. In Proceedings of the Advances in Neural Information Processing Systems 29, Annual Conference on Neural Information Processing Systems 2016, Barcelona, Spain, 5–10 December 2016; pp. 1–9. [Google Scholar]
- Rossant, C.; Harris, K.D. Hardware-accelerated interactive data visualization for neuroscience in Python. Front. Neuroinform. 2013, 7, 36. [Google Scholar] [CrossRef]
- Batista-Brito, R.; Vinck, M.; Ferguson, K.A.; Chang, J.T.; Laubender, D.; Lur, G.; Mossner, J.M.; Hernandez, V.G.; Ramakrishnan, C.; Deisseroth, K.; et al. Developmental Dysfunction of VIP Interneurons Impairs Cortical Circuits. Neuron 2017, 95, 884–895.e9. [Google Scholar] [CrossRef] [PubMed]
- Daviaud, N.; Friedel, R.H.; Zou, H. Vascularization and engraftment of transplanted human cerebral organoids in mouse cortex. eNeuro 2018, 5, 1–18. [Google Scholar] [CrossRef] [Green Version]
- Wang, J.; Chu, R.; Ni, N.; Nan, G. The effect of Matrigel as scaffold material for neural stem cell transplantation for treating spinal cord injury. Sci. Rep. 2020, 10, 2576. [Google Scholar] [CrossRef] [Green Version]
- Yang, G.; Mahadik, B.; Choi, J.Y.; Fisher, J.P. Vascularization in tissue engineering: Fundamentals and state-of-art. Prog. Biomed. Eng. 2020, 2, 012002. [Google Scholar] [CrossRef] [PubMed]
- Chen, V.S.; Morrison, J.P.; Southwell, M.; Foley, J.F.; Bolon, B.; Elmore, S.A. Histology Atlas of the Developing Prenatal and Postnatal Mouse Central Nervous System, with Emphasis on Prenatal Days E7.5 to E18.5. Toxicol. Pathol. 2017, 45, 705–744. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Krzyspiak, J.; Yan, J.; Ghosh, H.S.; Galinski, B.; Lituma, P.J.; Alvina, K.; Quezada, A.; Kee, S.; Grońska-Pęski, M.; De Tai, Y.; et al. Donor-derived vasculature is required to support neocortical cell grafts after stroke. Stem Cell Res. 2022, 59, 102642. [Google Scholar] [CrossRef] [PubMed]
- Liaudanskaya, V.; Jgamadze, D.; Berk, A.N.; Bischoff, D.J.; Gu, B.J.; Hawks-Mayer, H.; Whalen, M.J.; Chen, H.I.; Kaplan, D.L. Engineering advanced neural tissue constructs to mitigate acute cerebral inflammation after brain transplantation in rats. Biomaterials 2019, 192, 510–522. [Google Scholar] [CrossRef] [PubMed]
- Fang, W.Q.; Chen, W.W.; Jiang, L.; Liu, K.; Yung, W.H.; Fu, A.K.Y.; Ip, N.Y. Overproduction of Upper-Layer Neurons in the Neocortex Leads to Autism-like Features in Mice. Cell Rep. 2014, 9, 1635–1643. [Google Scholar] [CrossRef] [Green Version]
- Wiedenmann, B.; Franke, W.W. Identification and localization of synaptophysin, an integral membrane glycoprotein of Mr 38,000 characteristic of presynaptic vesicles. Cell 1985, 41, 1017–1028. [Google Scholar] [CrossRef]
- Williams, R.H.; Riedemann, T. Development, diversity and death of mge-derived cortical interneurons. Int. J. Mol. Sci. 2021, 22, 9297. [Google Scholar] [CrossRef]
- Jäkel, S.; Dimou, L. Glial cells and their function in the adult brain: A journey through the history of their ablation. Front. Cell. Neurosci. 2017, 11, 24. [Google Scholar] [CrossRef] [Green Version]
- Clarke, L.E.; Barres, B.A. Emerging roles of astrocytes in neural circuit development. Nat. Rev. Neurosci. 2013, 14, 311–321. [Google Scholar] [CrossRef] [Green Version]
- Sweeney, M.D.; Zhao, Z.; Montagne, A.; Nelson, A.R.; Zlokovic, B.V. Blood-brain barrier: From physiology to disease and back. Physiol. Rev. 2019, 99, 21–78. [Google Scholar] [CrossRef]
- Hinkle, D.A.; Baldwin, S.A.; Scheff, S.W.; Wise, P.M. GFAP and S100β expression in the cortex and hippocampus in response to mild cortical contusion. J. Neurotrauma 1997, 14, 729–738. [Google Scholar] [CrossRef] [PubMed]
- Arnold, T.; Betsholtz, C. The importance of microglia in the development of the vasculature in the central nervous system. Vasc. Cell 2013, 5, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- da Fonseca, A.C.C.; Matias, D.; Garcia, C.; Amaral, R.; Geraldo, L.H.; Freitas, C.; Lima, F.R.S. The impact of microglial activation on blood-brain barrier in brain diseases. Front. Cell. Neurosci. 2014, 8, 362. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lenz, K.M.; Nelson, L.H. Microglia and beyond: Innate immune cells as regulators of brain development and behavioral function. Front. Immunol. 2018, 9, 698. [Google Scholar] [CrossRef] [Green Version]
- Andreone, B.J.; Lacoste, B.; Gu, C. Neuronal and Vascular Interactions. Annu. Rev. Neurosci. 2015, 38, 25–46. [Google Scholar] [CrossRef] [Green Version]
- Gould, D.J.; Vadakkan, T.J.; Poche, R.A.; Dickinson, M.E. Multifractal and Lacunarity Analysis of Microvascular Morphology and Remodeling. Microcirculation 2011, 18, 136–151. [Google Scholar] [CrossRef]
- Fenlon, L.R.; Suárez, R.; Richards, L.J. The anatomy, organisation and development of contralateral callosal projections of the mouse somatosensory cortex. Brain Neurosci. Adv. 2017, 1, 2398212817694888. [Google Scholar] [CrossRef] [Green Version]
- Rajasethupathy, P.; Sankaran, S.; Marshel, J.H.; Kim, C.K.; Ferenczi, E.; Lee, S.Y.; Berndt, A.; Ramakrishnan, C.; Jaffe, A.; Lo, M.; et al. Projections from neocortex mediate top-down control of memory retrieval. Nature 2015, 526, 653–659. [Google Scholar] [CrossRef] [Green Version]
- Zakiewicz, I.M.; Bjaalie, J.G.; Leergaard, T.B. Brain-wide map of efferent projections from rat barrel cortex. Front. Neuroinform. 2014, 8, 5. [Google Scholar] [CrossRef] [Green Version]
- Mcdonald, A.J. Cortical pathways to the mammalian amygdala. Prog. Neurobiol. 1998, 55, 257–332. [Google Scholar] [CrossRef]
- Shen, J.; Colonnese, M.T. Development of activity in the mouse visual cortex. J. Neurosci. 2016, 36, 12259–12275. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ballout, N.; Frappé, I.; Péron, S.; Jaber, M.; Zibara, K.; Gaillard, A. Development and maturation of embryonic cortical neurons grafted into the damaged adult motor cortex. Front. Neural Circ. 2016, 10, 55. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Real, R.; Peter, M.; Trabalza, A.; Khan, S.; Smith, M.A.; Dopp, J.; Barnes, S.J.; Momoh, A.; Strano, A.; Volpi, E.; et al. In vivo modeling of human neuron dynamics and Down syndrome. Science 2018, 362, eaau1810. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, A.L.; Bennett, N.K.; Francis, N.L.; Halikere, A.; Clarke, S.; Moore, J.C.; Hart, R.P.; Paradiso, K.; Wernig, M.; Kohn, J.; et al. Generation and transplantation of reprogrammed human neurons in the brain using 3D microtopographic scaffolds. Nat. Commun. 2016, 7, 10862. [Google Scholar] [CrossRef] [PubMed]
- Agirman, G.; Broix, L.; Nguyen, L. Cerebral cortex development: An outside-in perspective. FEBS Lett. 2017, 591, 3978–3992. [Google Scholar] [CrossRef] [Green Version]
- Auger, F.A.; Gibot, L.; Lacroix, D. The Pivotal Role of Vascularization in Tissue Engineering. Annu. Rev. Biomed. Eng. 2013, 15, 177–200. [Google Scholar] [CrossRef]
- Hawkins, B.T.; Davis, T.P. The blood-brain barrier in health and disease. Pharmacol. Rev. 2005, 57, 173–185. [Google Scholar] [CrossRef]
- De Filippis, L.; Delia, D. Hypoxia in the regulation of neural stem cells. Cell. Mol. Life Sci. 2011, 68, 2831–2844. [Google Scholar] [CrossRef]
- Giandomenico, S.L.; Lancaster, M.A. Probing human brain evolution and development in organoids. Curr. Opin. Cell Biol. 2017, 44, 36–43. [Google Scholar] [CrossRef]
- Espuny-Camacho, I.; Michelsen, K.A.; Gall, D.; Linaro, D.; Hasche, A.; Bonnefont, J.; Bali, C.; Orduz, D.; Bilheu, A.; Herpoel, A.; et al. Pyramidal Neurons Derived from Human Pluripotent Stem Cells Integrate Efficiently into Mouse Brain Circuits In Vivo. Neuron 2013, 77, 440–456. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Figure | 1D | 1F | 1H | 1K | 1O | 2C-E | 2F-H | 2I,J | 2L | |
|---|---|---|---|---|---|---|---|---|---|---|
| Number of host mice | 3 * | 3 * | 3 * | 3 * | 3 * | 9 | 8 * | 5 | 3 | |
| Figure | 3 | 4 | 5 | 6 | S1A | S1B | S1C | S2C,D | S2E | Total |
| Number of host mice | 2 | 9 | 2 * | 3 * | 3 | 80 | 6 | 19 | 10 | 200 |
| Table | Company | Catalog No. | Species | Dilution |
|---|---|---|---|---|
| Anti-CD105 | Biolegend | 120402 | Rat | 1:50 * |
| Anti-CD31 | BD Pharmingen | 553370 | Rat | 1:50 * |
| Anti-CTIP2 | abcam | ab18465 | Rat | 1:500 * |
| Anti-GABA | Millipore-Sigma | ab2052 | Rabbit | 1:1000 |
| Anti-GFAP | Invitrogen | 13-0300 | Rat | 1:500 |
| Anti-GFP | ThermoFisher | A11122 | Rabbit | 1:250 |
| Hoechst 33342 | Invitrogen | H3570 | 1:1000 | |
| IB4-647 | ThermoFisher | I32450 | 1 μg/μL | |
| Anti-Iba1 | Wako | 019-19741 | Rabbit | 1:100 |
| Anti-MBP | abcam | ab7349 | Rat | 1:50 |
| Anti-NeuN | Synaptic Systems | 266004 | Guinea Pig | 1:500 |
| Anti-OLIG2 | Millipore-Sigma | ab9610 | Rabbit | 1:50 |
| Anti-SATB2 | abcam | ab92446 | Rabbit | 1:500 * |
| Anti-Synaptophysin | abcam | ab32127 | Rabbit | 1:500 * |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Quezada, A.; Ward, C.; Bader, E.R.; Zolotavin, P.; Altun, E.; Hong, S.; Killian, N.J.; Xie, C.; Batista-Brito, R.; Hébert, J.M. An In Vivo Platform for Rebuilding Functional Neocortical Tissue. Bioengineering 2023, 10, 263. https://doi.org/10.3390/bioengineering10020263
Quezada A, Ward C, Bader ER, Zolotavin P, Altun E, Hong S, Killian NJ, Xie C, Batista-Brito R, Hébert JM. An In Vivo Platform for Rebuilding Functional Neocortical Tissue. Bioengineering. 2023; 10(2):263. https://doi.org/10.3390/bioengineering10020263
Chicago/Turabian StyleQuezada, Alexandra, Claire Ward, Edward R. Bader, Pavlo Zolotavin, Esra Altun, Sarah Hong, Nathaniel J. Killian, Chong Xie, Renata Batista-Brito, and Jean M. Hébert. 2023. "An In Vivo Platform for Rebuilding Functional Neocortical Tissue" Bioengineering 10, no. 2: 263. https://doi.org/10.3390/bioengineering10020263