
A Multi-Analytical Approach to Infer Mineral–Microbial Interactions Applied to Petroglyph Sites in the Negev Desert of Israel

 , , and
, , and
Abstract
:1. Introduction
2. Materials and Methods
2.1. Study Location and Sampling
2.2. DNA Extraction, Whole Genome Amplification (WGA), Library Preparation and Sequencing
2.3. Sequence Analysis
2.4. Optical Microscopy (OM)
2.5. X-ray Fluorescence (XRF)
2.6. Scanning Electron Microscopy Coupled with Energy Dispersive X-ray Spectroscopy (SEM–EDX)
2.7. Micro-Raman Spectroscopy
3. Results
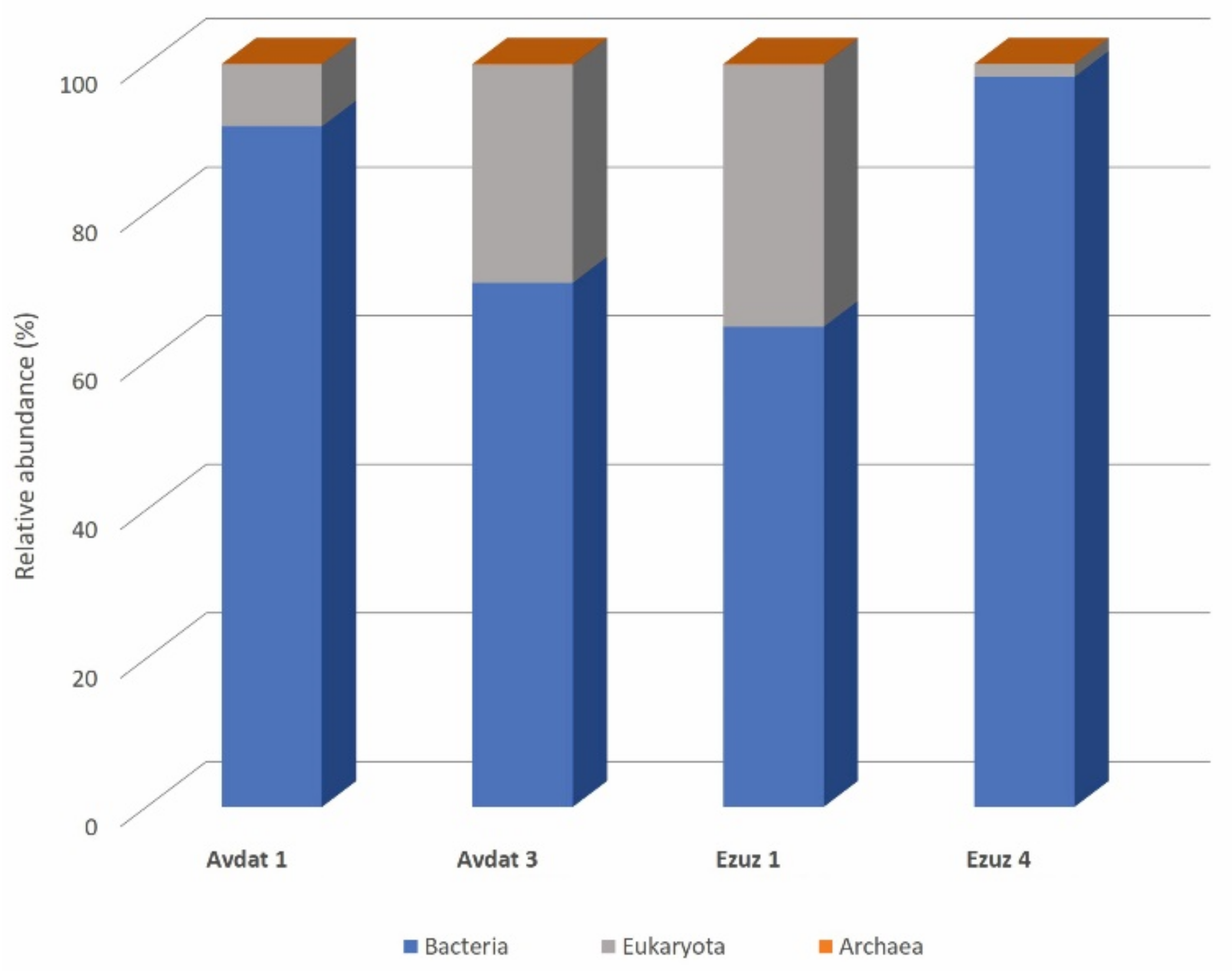
3.1. Metagenomic Analysis
3.1.1. DNA Yield and Sequencing Analysis
3.1.2. Microbiomes of the Stone Samples
3.1.3. Bacterial Communities
3.1.4. Eukaryotic Communities
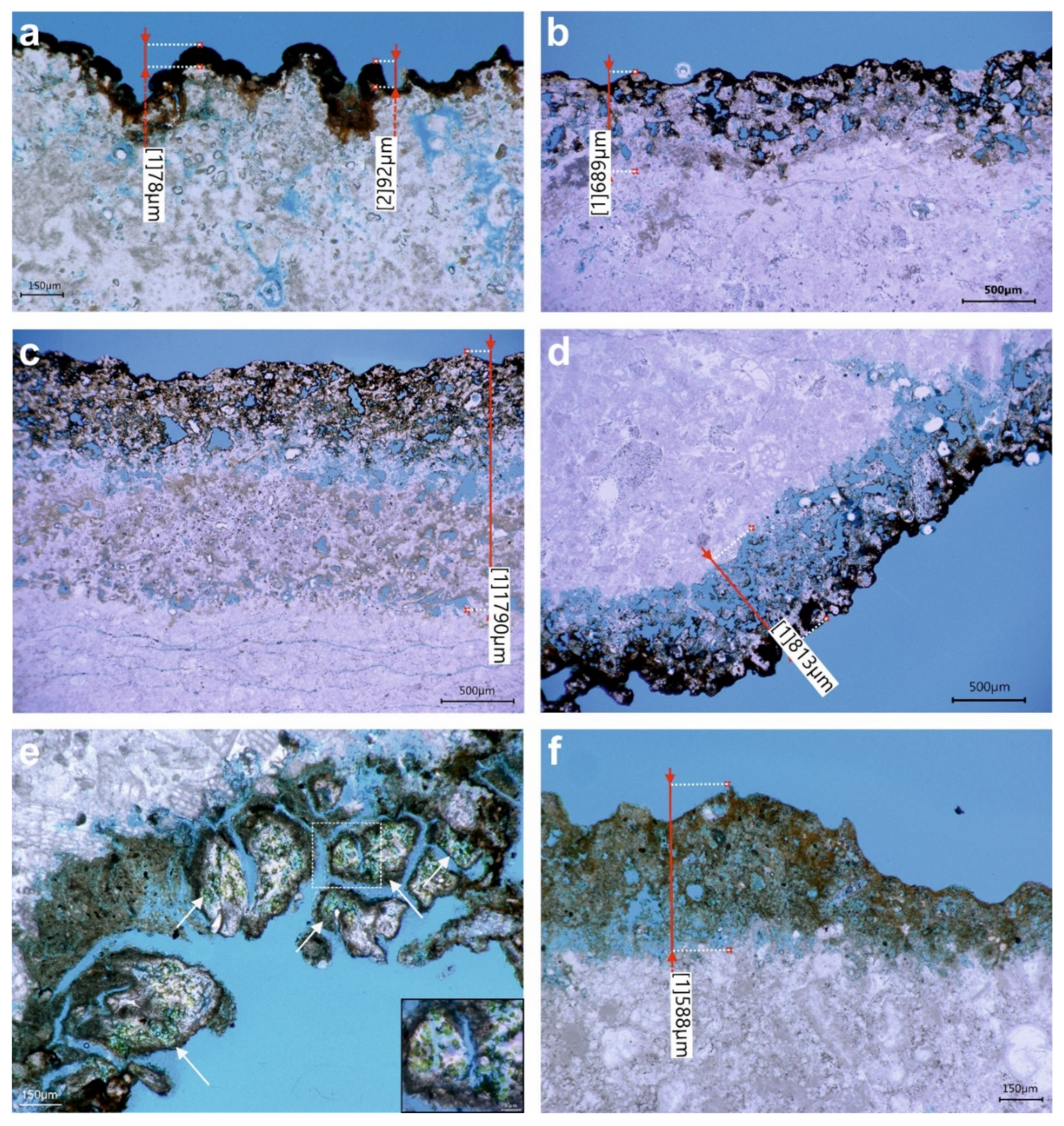
3.2. Digital Optical Microscopy
3.3. X-ray Fluorescence Spectroscopy (XRF)
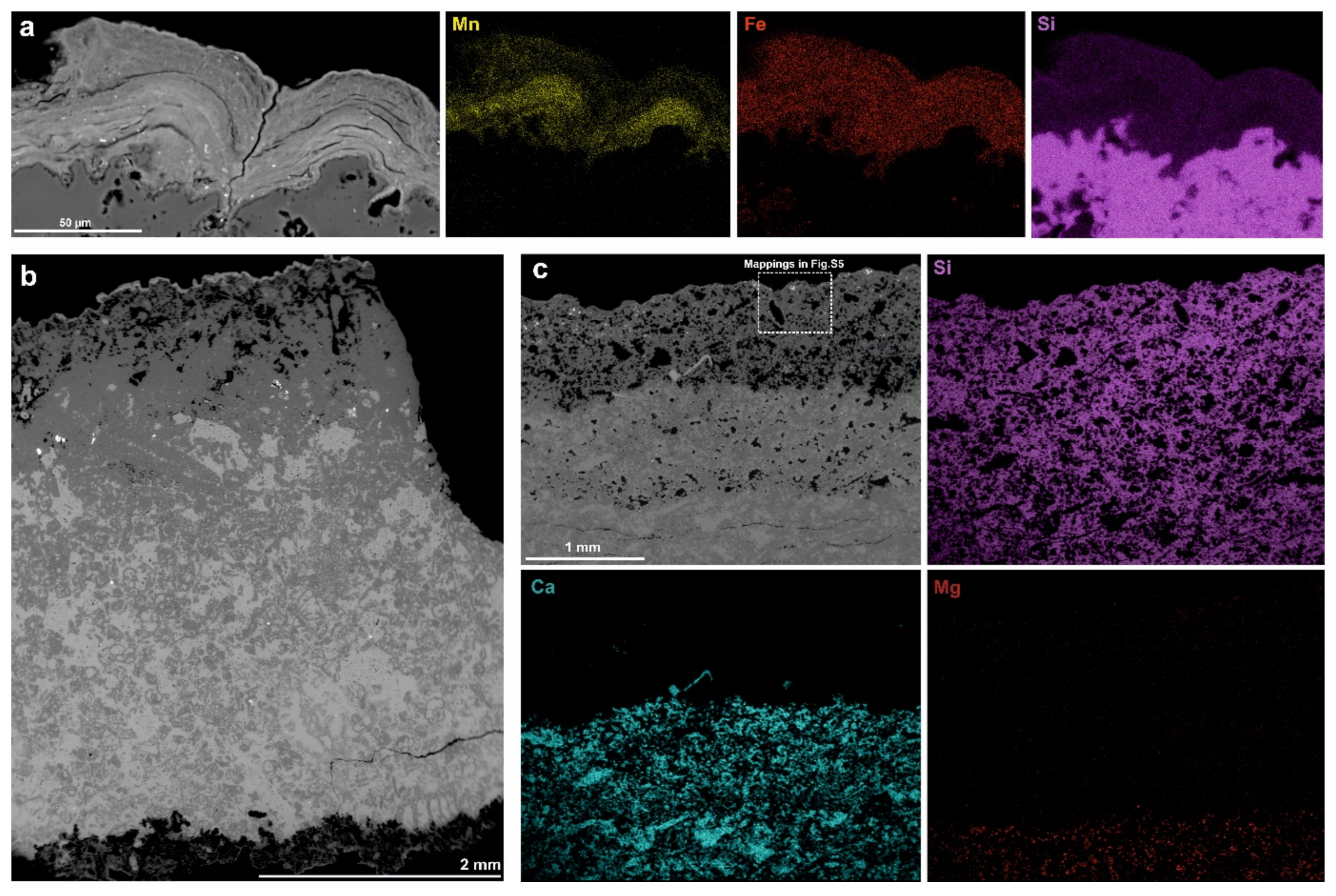
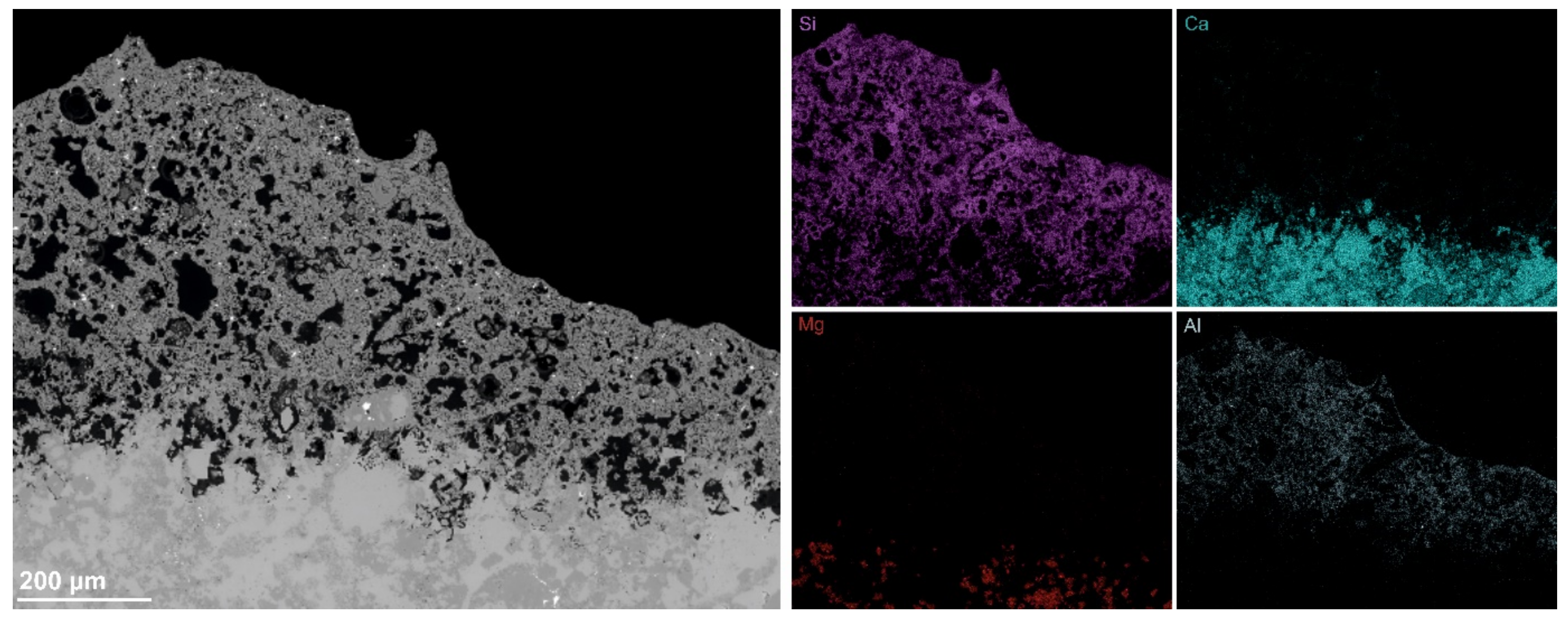
3.4. Scanning Electron Microscopy Coupled with EDX Microanalysis (SEM–EDX)
3.5. Micro-Raman Spectroscopy
4. Discussion
5. Conclusions
- most of the microorganisms detected by metagenomic analysis are taxa adapted to live in harsh environmental condition such as the Negev desert, with a high resistance to radiation and desiccation;
- weathering of the stone beneath the black crust was associated with the potential of the species detected to leach the calcareous matrix of the rock in order to find optimal conditions of growth while being sheltered by the black desert varnish;
- in sample EZ4, the orange layer is probably not a different kind of coating, but it is actually a weathered area of the bedrock in which the black varnish is no longer present; therefore, we hypothesize that this sample is in a later stage of weathering compared to the others;
- we acknowledge the possible microbial involvement in the origin of the crust, as previously reported by many authors in the literature, and, on the other hand, what we observe in our study are patterns of deterioration in the bedrock and in the black varnish, feasibly connected to microbial action. The two hypotheses can be viewed in terms of succession of biogeochemical processes and of microbial communities.
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Aubert, M.; Lebe, R.; Oktaviana, A.A.; Tang, M.; Burhan, B.; Hamrullah; Jusdi, A.; Abdullah; Hakim, B.; Zhao, J.-X.; et al. Earliest hunting scene in prehistoric art. Nature 2019, 576, 442–445. [Google Scholar] [CrossRef] [PubMed]
- Brumm, A.; Oktaviana, A.A.; Burhan, B.; Hakim, B.; Lebe, R.; Zhao, J.-X.; Sulistyarto, P.H.; Ririmasse, M.; Adhityatama, S.; Sumantri, I.; et al. Oldest cave art found in Sulawesi. Sci. Adv. 2021, 7, eabd4648. [Google Scholar] [CrossRef] [PubMed]
- Francis, A.; Loendorf, L.L. Ancient Visions: Petroglyphs and Pictographs of the Wind River and Bighorn Country, Wyoming and Montana; University of Utah Press: Salt Lake City, UT, USA, 2004. [Google Scholar]
- Bednarik, R.G. Pleistocene Palaeoart of Africa. Arts 2013, 2, 6–34. [Google Scholar] [CrossRef] [Green Version]
- Dorn, R.I. Anthropogenic Interactions with Rock Varnish. In Biogeochemical Cycles: Ecological Drivers and Environmental Impact; John Wiley & Sons: New York, NY, USA, 2020; pp. 267–283. [Google Scholar]
- Smith, B.W.; Black, J.L.; Hoerle, S.; Ferland, M.A.; Diffey, S.M.; Neumann, J.T.; Geisler, T. The Impact of Industrial Pollution on the Rock Art of Murujuga, Western Australia. J. Aust. Rock Art Res. Assoc. 2022, 39, 3–14. [Google Scholar]
- Neumann, J.T.; Black, J.L.; Hœrlé, S.; Smith, B.W.; Watkins, R.; Lagos, M.; Ziegler, A.; Geisler, T. Artificial weathering of rock types bearing petroglyphs from Murujuga, Western Australia. Herit. Sci. 2022, 10, 77. [Google Scholar] [CrossRef]
- Fernandes, A.P.B. The Conservation Programme of the Côa Valley Archaeological Park: Philosophy, Objectives and Action. Conserv. Manag. Archaeol. Sites 2007, 9, 71–96. [Google Scholar] [CrossRef]
- Siegesmund, S.; Snethlage, R. Stone in Architecture. Properties, Durability, 5th ed.; Springer: Berlin/Heidelberg, Germany, 2014. [Google Scholar]
- Gorbushina, A.A.; Krumbein, W. Role of Microorganisms in Wear Down of Rocks and Minerals. Microorg. Soils Roles Genes. Funct. 2005, 3, 59–84. [Google Scholar] [CrossRef]
- Sterflinger, K.; Krumbein, W.E.; Lellau, T.; Rullkötter, J. Microbially mediated orange patination of rock surfaces. Anc. Biomol. 1999, 3, 51–65. [Google Scholar]
- Steiger, M.; Charola, E.; Sterflinger, K. Weathering and Deterioration. In Stone in Architecture: Properties, Durability, 5th ed; Springer: Berlin/Heidelberg, Germany, 2014; pp. 225–316. [Google Scholar]
- Louati, M.; Ennis, N.J.; Ghodhbane-Gtari, F.; Hezbri, K.; Sevigny, J.L.; Fahnestock, M.F.; Cherif-Silini, H.; Bryce, J.G.; Tisa, L.S.; Gtari, M. Elucidating the ecological networks in stone-dwelling microbiomes. Environ. Microbiol. 2020, 22, 1467–1480. [Google Scholar] [CrossRef]
- Eisenberg-Degen, D.; Rosen, S.A. Chronological Trends in Negev Rock Art: The Har Michia Petroglyphs as a Test Case. Arts 2013, 2, 225–252. [Google Scholar] [CrossRef] [Green Version]
- Eisenberg-Degen, D.; Nash, G. Hunting and Gender as Reflected in the Central Negev Rock Art, Israel. Time Mind 2014, 7, 259–277. [Google Scholar] [CrossRef]
- Nir, I.; Barak, H.; Kramarsky-Winter, E.; Kushmaro, A. Seasonal diversity of the bacterial communities associated with petroglyphs sites from the Negev Desert, Israel. Ann. Microbiol. 2019, 69, 1079–1086. [Google Scholar] [CrossRef]
- Perry, R.S.; Adams, J.B. Desert varnish: Evidence of cyclic deposition of manganese. Nature 1978, 276, 489–491. [Google Scholar] [CrossRef]
- Martínez-Pabello, P.U.; Villalobos, C.; Sedov, S.; Solleiro-Rebolledo, E.; Solé, J.; Pi-Puig, T.; Chávez-Vergara, B.; Díaz-Ortega, J.; Gubin, A. Rock varnish as a natural canvas for rock art in La Proveedora, northwestern Sonoran Desert (Mexico): Integrating archaeological and geological evidences. Quat. Int. 2021, 572, 74–87. [Google Scholar] [CrossRef]
- Goldsmith, Y.; Stein, M.; Enzel, Y. From dust to varnish: Geochemical constraints on rock varnish formation in the Negev Desert, Israel. Geochim. Cosmochim. Acta 2014, 126, 97–111. [Google Scholar] [CrossRef]
- Andreae, M.O.; Al-Amri, A.; Andreae, C.M.; Guagnin, M.; Jochum, K.P.; Stoll, B.; Weis, U. Archaeometric studies on the petroglyphs and rock varnish at Kilwa and Sakaka, northern Saudi Arabia. Arab. Archaeol. Epigr. 2020, 31, 219–244. [Google Scholar] [CrossRef]
- Harmon, R.S.; Khashchevskaya, D.; Morency, M.; Owen, L.A.; Jennings, M.; Knott, J.R.; Dortch, J.M. Analysis of Rock Varnish from the Mojave Desert by Handheld Laser-Induced Breakdown Spectroscopy. Molecules 2021, 26, 5200. [Google Scholar] [CrossRef]
- Otter, L.M.; Macholdt, D.S.; Jochum, K.P.; Stoll, B.; Weis, U.; Weber, B.; Scholz, D.; Haug, G.H.; Al-Amri, A.M.; Andreae, M.O. Geochemical insights into the relationship of rock varnish and adjacent mineral dust fractions. Chem. Geol. 2020, 551, 119775. [Google Scholar] [CrossRef]
- Andreae, M.O.; Al-Amri, A.; Andreae, T.W.; Garfinkel, A.; Haug, G.; Jochum, K.P.; Stoll, B.; Weis, U. Geochemical studies on rock varnish and petroglyphs in the Owens and Rose Valleys, California. PLoS ONE 2020, 15, e0235421. [Google Scholar] [CrossRef]
- Macholdt, D.; Jochum, K.; Pöhlker, C.; Arangio, A.; Förster, J.-D.; Stoll, B.; Weis, U.; Weber, B.; Müller, M.; Kappl, M.; et al. Characterization and differentiation of rock varnish types from different environments by microanalytical techniques. Chem. Geol. 2017, 459, 91–118. [Google Scholar] [CrossRef]
- Lang-Yona, N.; Maier, S.; Macholdt, D.S.; Müller-Germann, I.; Yordanova, P.; Rodriguez-Caballero, E.; Jochum, K.P.; Al-Amri, A.; Andreae, M.O.; Fröhlich-Nowoisky, J.; et al. Insights into microbial involvement in desert varnish formation retrieved from metagenomic analysis. Environ. Microbiol. Rep. 2018, 10, 264–271. [Google Scholar] [CrossRef] [PubMed]
- Dorn, R.I.; Oberlander, T.M. Microbial Origin of Desert Varnish. Science 1981, 213, 1245–1247. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lingappa, U.F.; Yeager, C.M.; Sharma, A.; Lanza, N.L.; Morales, D.P.; Xie, G.; Atencio, A.D.; Chadwick, G.L.; Monteverde, D.R.; Magyar, J.S.; et al. An ecophysiological explanation for manganese enrichment in rock varnish. Proc. Natl. Acad. Sci. USA 2021, 118, e2025188118. [Google Scholar] [CrossRef] [PubMed]
- Esposito, A.; Ahmed, E.; Ciccazzo, S.; Sikorski, J.; Overmann, J.; Holmström, S.J.M.; Brusetti, L. Comparison of Rock Varnish Bacterial Communities with Surrounding Non-Varnished Rock Surfaces: Taxon-Specific Analysis and Morphological Description. Microb. Ecol. 2015, 70, 741–750. [Google Scholar] [CrossRef] [PubMed]
- Wierzchos, J.; de los Ríos, A.; Ascaso, C. Microorganisms in desert rocks: The edge of life on Earth. Int. Microbiol. 2012, 15, 173–183. [Google Scholar] [CrossRef] [Green Version]
- Northup, D.E.; Snider, J.R.; Spilde, M.N.; Porter, M.L.; Van De Kamp, J.L.; Boston, P.J.; Nyberg, A.M.; Bargar, J.R. Diversity of rock varnish bacterial communities from Black Canyon, New Mexico. J. Geophys. Res. Biogeosci. 2010, 115, G2. [Google Scholar] [CrossRef] [Green Version]
- Knight, K.; St. Clair, L.L.; Gardner, J. Lichen Biodeterioration at Inscription Rock, El Morro National Monument, Ramah, New Mexico, USA. In Biodeterioration of Stone Surfaces; St. Clair, L.L., Seaward, M.R.D., Eds.; Springer: Dordrecht, The Netherlands, 2004; pp. 129–163. [Google Scholar]
- Dandridge, D.E. Lichens: The Challenge for Rock Art Conservation. Ph.D. Thesis, Texas A & M University, College Station, TX, USA, 2007. [Google Scholar]
- Nir, I.; Barak, H.; Kramarsky-Winter, E.; Kushmaro, A.; Ríos, A. Microscopic and biomolecular complementary approaches to characterize bioweathering processes at petroglyph sites from the Negev Desert, Israel. Environ. Microbiol. 2022, 24, 967–980. [Google Scholar] [CrossRef]
- Evenari, M.; Shanan, L.; Tadmor, N. The Negev: The Challenge of a Desert; Harvard University Press: Cambridge, MA, USA, 1982. [Google Scholar]
- Hillel, D. Negev Land, Water, and Life in a Desert Environment; Praeger: New York, NY, USA, 1982. [Google Scholar]
- Kidron, G.J. Altitude dependent dew and fog in the Negev Desert, Israel. Agric. For. Meteorol. 1999, 96, 1–8. [Google Scholar] [CrossRef]
- Piñar, G.; Poyntner, C.; Lopandic, K.; Tafer, H.; Sterflinger, K. Rapid diagnosis of biological colonization in cultural artefacts using the MinION nanopore sequencing technology. Int. Biodeterior. Biodegrad. 2020, 148, 104908. [Google Scholar] [CrossRef]
- RRUFF Database. Available online: https://rruff.info/ (accessed on 3 May 2022).
- Caggiani, M.; Cosentino, A.; Mangone, A. Pigments Checker version 3.0, a handy set for conservation scientists: A free online Raman spectra database. Microchem. J. 2016, 129, 123–132. [Google Scholar] [CrossRef]
- Hanesch, M. Raman spectroscopy of iron oxides and (oxy)hydroxides at low laser power and possible applications in environmental magnetic studies. Geophys. J. Int. 2009, 177, 941–948. [Google Scholar] [CrossRef]
- Bruins, J.H.; Petrusevski, B.; Slokar, Y.M.; Kruithof, J.C.; Kennedy, M.D. Manganese removal from groundwater: Characterization of filter media coating. Desalin. Water Treat. 2015, 55, 1851–1863. [Google Scholar] [CrossRef]
- Jehlička, J.; Edwards, H.; Oren, A. Bacterioruberin and salinixanthin carotenoids of extremely halophilic Archaea and Bacteria: A Raman spectroscopic study. Spectrochim. Acta Part A Mol. Biomol. Spectrosc. 2013, 106, 99–103. [Google Scholar] [CrossRef]
- Piñar, G.; Sclocchi, M.C.; Pinzari, F.; Colaizzi, P.; Graf, A.; Sebastiani, M.L.; Sterflinger, K. The Microbiome of Leonardo da Vinci’s Drawings: A Bio-Archive of Their History. Front. Microbiol. 2020, 11, 593401. [Google Scholar] [CrossRef]
- Planý, M.; Pinzari, F.; Šoltys, K.; Kraková, L.; Cornish, L.; Pangallo, D.; Jungblut, A.D.; Little, B. Fungal-induced atmospheric iron corrosion in an indoor environment. Int. Biodeterior. Biodegrad. 2021, 159, 105204. [Google Scholar] [CrossRef]
- Grottoli, A.; Beccaccioli, M.; Zoppis, E.; Fratini, R.S.; Schifano, E.; Santarelli, M.L.; Uccelletti, D.; Reverberi, M. Nanopore Sequencing and Bioinformatics for Rapidly Identifying Cultural Heritage Spoilage Microorganisms. Front. Mater. 2020, 7, 14. [Google Scholar] [CrossRef] [Green Version]
- Krumbein, W.E.; Jens, K. Biogenic rock varnishes of the negev desert (Israel) an ecological study of iron and manganese transformation by cyanobacteria and fungi. Oecologia 1981, 50, 25–38. [Google Scholar] [CrossRef]
- Vikram, S.; Guerrero, L.D.; Makhalanyane, T.P.; Le, P.; Seely, M.; Cowan, D.A. Metagenomic analysis provides insights into functional capacity in a hyperarid desert soil niche community. Environ. Microbiol. 2015, 18, 1875–1888. [Google Scholar] [CrossRef]
- Hirschberg, J.; Chamovitz, D. Carotenoids in Cyanobacteria BT-The Molecular Biology of Cyanobacteria; Bryant, D.A., Ed.; Springer: Dordrecht, The Netherlands, 1994; pp. 559–579. [Google Scholar]
- Perera, I.; Subashchandrabose, S.R.; Venkateswarlu, K.; Naidu, R.; Megharaj, M. Consortia of cyanobacteria/microalgae and bacteria in desert soils: An underexplored microbiota. Appl. Microbiol. Biotechnol. 2018, 102, 7351–7363. [Google Scholar] [CrossRef]
- Lacap, D.C.; Warren-Rhodes, K.A.; McKay, C.P.; Pointing, S.B. Cyanobacteria and chloroflexi-dominated hypolithic colonization of quartz at the hyper-arid core of the Atacama Desert, Chile. Extremophiles 2010, 15, 31–38. [Google Scholar] [CrossRef] [Green Version]
- Mishra, S. Chapter 1-Cyanobacterial imprints in diversity and phylogeny. In Advances in Cyanobacterial Biology; Singh, P.K., Kumar, A., Singh, V.K., Shrivastava, C.B., Eds.; Academic Press: Cambridge, MA, USA, 2020; pp. 1–15. [Google Scholar]
- Abed, R.M.; Al Kharusi, S.; Schramm, A.; Robinson, M.D. Bacterial diversity, pigments and nitrogen fixation of biological desert crusts from the Sultanate of Oman. FEMS Microbiol. Ecol. 2010, 72, 418–428. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wierzchos, J.; Ascaso, C.; McKay, C.P. Endolithic Cyanobacteria in Halite Rocks from the Hyperarid Core of the Atacama Desert. Astrobiology 2006, 6, 415–422. [Google Scholar] [CrossRef] [PubMed]
- Wierzchos, J.; DiRuggiero, J.; Vítek, P.; Artieda, O.; Souza-Egipsy, V.; Škaloud, P.; Tisza, M.; Davila, A.F.; Vílchez, C.; Garbayo, I.; et al. Adaptation strategies of endolithic chlorophototrophs to survive the hyperarid and extreme solar radiation environment of the Atacama Desert. Front. Microbiol. 2015, 6, 934. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mergelov, N.; Mueller, C.W.; Prater, I.; Shorkunov, I.; Dolgikh, A.; Zazovskaya, E.; Shishkov, V.; Krupskaya, V.; Abrosimov, K.; Cherkinsky, A.; et al. Alteration of rocks by endolithic organisms is one of the pathways for the beginning of soils on Earth. Sci. Rep. 2018, 8, 3367. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ríos, A.D.L.; Ascaso, C. Contributions of in situ microscopy to the current understanding of stone biodeterioration. Int. Microbiol. 2005, 8, 181–188. [Google Scholar]
- Grbic, M.L.; Vukojevic, J.; Subakov-Simic, G.; Krizmanic, J.; Stupar, M. Biofilm forming cyanobacteria, algae and fungi on two historic monuments in Belgrade, Serbia. Arch. Biol. Sci. 2010, 62, 625–631. [Google Scholar] [CrossRef]
- Genderjahn, S.; Lewin, S.; Horn, F.; Schleicher, A.; Mangelsdorf, K.; Wagner, D. Living Lithic and Sublithic Bacterial Communities in Namibian Drylands. Microorganisms 2021, 9, 235. [Google Scholar] [CrossRef]
- Ferreira, A.C.; Nobre, M.F.; Moore, E.; Rainey, F.A.; Battista, J.R.; Da Costa, M.S. Characterization and radiation resistance of new isolates of Rubrobacter radiotolerans and Rubrobacter xylanophilus. Extremophiles 1999, 3, 235–238. [Google Scholar] [CrossRef]
- Webb, K.M.; DiRuggiero, J. Role of Mn2+ and Compatible Solutes in the Radiation Resistance of Thermophilic Bacteria and Archaea. Archaea 2012, 2012, 845756. [Google Scholar] [CrossRef] [Green Version]
- Piñar, G.; Ettenauer, J.; Sterflinger, K. ‘La vie en ros’: A review of the rosy discoloration of subsurface monuments. In The Conservation of Subterranean Cultural Heritage; Saiz-Jimenez, C., Ed.; CRC Press: Boca Raton, FL, USA, 2014; pp. 113–124. [Google Scholar] [CrossRef]
- Laiz, L.; Miller, A.Z.; Jurado, V.; Akatova, E.V.; Sanchez-Moral, S.; Gonzalez, J.M.; Dionísio, A.; Macedo, M.F.; Sáiz-Jiménez, C. Isolation of five Rubrobacter strains from biodeteriorated monuments. Naturwissenschaften 2009, 96, 71–79. [Google Scholar] [CrossRef]
- Tescari, M.; Visca, P.; Frangipani, E.; Bartoli, F.; Rainer, L.; Caneva, G. Celebrating centuries: Pink-pigmented bacteria from rosy patinas in the House of Bicentenary (Herculaneum, Italy). J. Cult. Heritage 2018, 34, 43–52. [Google Scholar] [CrossRef]
- Maeng, S.; Park, Y.; Lee, S.E.; Han, J.H.; Cha, I.-T.; Lee, K.-E.; Lee, B.-H.; Kim, M.K. Pontibacter pudoricolor sp. nov., and Pontibacter russatus sp. nov. radiation-resistant bacteria isolated from soil. Int. J. Gen. Mol. Microbiol. 2020, 113, 1361–1369. [Google Scholar] [CrossRef]
- Zhang, J.-Y.; Liu, X.-Y.; Liu, S.-J. Adhaeribacter terreus sp. nov., isolated from forest soil. Int. J. Syst. Evol. Microbiol. 2009, 59, 1595–1598. [Google Scholar] [CrossRef] [Green Version]
- Klassen, J.L.; Foght, J.M. Differences in Carotenoid Composition among Hymenobacter and Related Strains Support a Tree-Like Model of Carotenoid Evolution. Appl. Environ. Microbiol. 2008, 74, 2016–2022. [Google Scholar] [CrossRef] [Green Version]
- Sterflinger, K.; Tesei, D.; Zakharova, K. Fungi in hot and cold deserts with particular reference to microcolonial fungi. Fungal Ecol. 2012, 5, 453–462. [Google Scholar] [CrossRef]
- Sterflinger, K.; De Baere, R.; De Hoog, G.; De Wachter, R.; Krumbein, W.E.; Haase, G. Coniosporium perforans and C. apollinis, two new rock-inhabiting fungi isolated from marble in the Sanctuary of Delos (Cyclades, Greece). Int. J. Gen. Mol. Microbiol. 1997, 72, 349–363. [Google Scholar] [CrossRef]
- Coleine, C.; Stajich, J.E.; de Los Ríos, A.; Selbmann, L. Beyond the extremes: Rocks as ultimate refuge for fungi in drylands. Mycologia 2020, 113, 108–133. [Google Scholar] [CrossRef]
- Engel, C.; Robert, S. Chemical data on desert varnish. Bull. Geol. Soc. Am. 1958, 69, 487–518. [Google Scholar] [CrossRef]
- Liu, T.; Broecker, W.S. How fast does rock varnish grow? Geology 2000, 28, 183. [Google Scholar] [CrossRef]
- Malherbe, C.; Ingley, R.; Hutchinson, I.; Edwards, H.; Carr, A.; Harris, L.; Boom, A. Biogeological Analysis of Desert Varnish Using Portable Raman Spectrometers. Astrobiology 2015, 15, 442–452. [Google Scholar] [CrossRef]
- Ren, G.; Yan, Y.; Nie, Y.; Lu, A.; Wu, X.; Li, Y.; Wang, C.; Ding, H. Natural Extracellular Electron Transfer Between Semiconducting Minerals and Electroactive Bacterial Communities Occurred on the Rock Varnish. Front. Microbiol. 2019, 10, 293. [Google Scholar] [CrossRef]
- Edwards, H.G.M.; Moody, C.A.; Villar, S.E.J.; Mancinelli, R. Raman spectroscopy of desert varnishes and their rock substrata. J. Raman Spectrosc. 2004, 35, 475–479. [Google Scholar] [CrossRef]
- Skipper, P.J.A.; Skipper, L.K.; Dixon, R.A. A metagenomic analysis of the bacterial microbiome of limestone, and the role of associated biofilms in the biodeterioration of heritage stone surfaces. Sci. Rep. 2022, 12, 1–19. [Google Scholar] [CrossRef]
- Ríos, A.D.L.; Cámara, B.; del Cura, M.G.; Rico, V.J.; Galván, V.; Ascaso, C. Deteriorating effects of lichen and microbial colonization of carbonate building rocks in the Romanesque churches of Segovia (Spain). Sci. Total Environ. 2009, 407, 1123–1134. [Google Scholar] [CrossRef]
- Subrahmanyam, G.; Vaghela, R.; Bhatt, N.P.; Archana, G. Carbonate-Dissolving Bacteria from ‘Miliolite’, a Bioclastic Limestone, from Gopnath, Gujarat, Western India. Microbes Environ. 2012, 27, 334–337. [Google Scholar] [CrossRef] [Green Version]
- Uroz, S.; Calvaruso, C.; Turpault, M.; Sarniguet, A.; de Boer, W.; Leveau, J.; Frey-Klett, P. Efficient mineral weathering is a distinctive functional trait of the bacterial genus Collimonas. Soil Biol. Biochem. 2009, 41, 2178–2186. [Google Scholar] [CrossRef]
- Lian, B.; Chen, Y.; Zhu, L.; Yang, R. Effect of Microbial Weathering on Carbonate Rocks. Earth Sci. Front. 2008, 15, 90–99. [Google Scholar] [CrossRef]
- dos Santos, S.I.P.; Yang, Y.; Rosa, M.T.G.; Rodrigues, M.A.A.; De La Tour, C.B.; Sommer, S.; Teixeira, M.; Carrondo, M.A.; Cloetens, P.; Abreu, I.A.; et al. The interplay between Mn and Fe in Deinococcus radiodurans triggers cellular protection during paraquat-induced oxidative stress. Sci. Rep. 2019, 9, 17217. [Google Scholar] [CrossRef]
- Bernardini, S.; Bellatreccia, F.; Columbu, A.; Vaccarelli, I.; Pellegrini, M.; Jurado, V.; Del Gallo, M.; Saiz-Jimenez, C.; Sodo, A.; Millo, C.; et al. Morpho-Mineralogical and Bio-Geochemical Description of Cave Manganese Stromatolite-Like Patinas (Grotta del Cervo, Central Italy) and Hints on Their Paleohydrological-Driven Genesis. Front. Earth Sci. 2021, 9, 642667. [Google Scholar] [CrossRef]
- Saiz-Jimenez, C.; Miller, A.Z.; Martin-Sanchez, P.M.; Hernández-Mariné, M. Uncovering the origin of the black stains in Lascaux Cave in France. Environ. Microbiol. 2012, 14, 3220–3231. [Google Scholar] [CrossRef] [Green Version]
- Barboza, N.R.; Amorim, S.S.; Santos, P.A.; Reis, F.D.; Cordeiro, M.M.; Guerra-Sá, R.; Leão, V.A. Indirect Manganese Removal by Stenotrophomonas sp. and Lysinibacillus sp. Isolated from Brazilian Mine Water. BioMed Res. Int. 2015, 2015, 925972. [Google Scholar] [CrossRef] [PubMed] [Green Version]










{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Sample | Petroglyph Site | Brief Description of the Crust |
|---|---|---|
| AV1 | Avdat | Very dark crust covering a reddish layer |
| AV2 | Avdat | Very dark crust covering a reddish layer |
| AV3 | Avdat | Thinner black crust, intermixed with reddish layer |
| EZ1 | Ezuz | Very dark crust covering a reddish layer |
| EZ2 | Ezuz | Very dark crust covering a reddish layer |
| EZ3 | Ezuz | Dark crust covering a reddish layer |
| EZ4 | Ezuz | Orange thick crust |
| Elements * | Black Varnish (%) | Reddish Layer (%) | Bedrock (%) |
|---|---|---|---|
| Mg | 1.1 | 1.1 | 2.0 |
| Al | 8.4 | 7.0 | 3.2 |
| Si | 66.1 | 73.4 | 48.9 |
| P | 1.1 | 0.5 | 0 |
| S | 1.0 | 0.3 | 0.1 |
| K | 1.1 | 1.1 | 0.4 |
| Ca | 8.3 | 9.9 | 40.9 |
| Ti | 0.6 | 0.5 | 0.1 |
| Mn | 3.1 | 0.8 | 0.1 |
| Fe | 8.0 | 4.7 | 1.2 |
| Ba | 0.3 | 0 | 0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Rabbachin, L.; Piñar, G.; Nir, I.; Kushmaro, A.; Pavan, M.J.; Eitenberger, E.; Waldherr, M.; Graf, A.; Sterflinger, K. A Multi-Analytical Approach to Infer Mineral–Microbial Interactions Applied to Petroglyph Sites in the Negev Desert of Israel. Appl. Sci. 2022, 12, 6936. https://doi.org/10.3390/app12146936
Rabbachin L, Piñar G, Nir I, Kushmaro A, Pavan MJ, Eitenberger E, Waldherr M, Graf A, Sterflinger K. A Multi-Analytical Approach to Infer Mineral–Microbial Interactions Applied to Petroglyph Sites in the Negev Desert of Israel. Applied Sciences. 2022; 12(14):6936. https://doi.org/10.3390/app12146936
Chicago/Turabian StyleRabbachin, Laura, Guadalupe Piñar, Irit Nir, Ariel Kushmaro, Mariela J. Pavan, Elisabeth Eitenberger, Monika Waldherr, Alexandra Graf, and Katja Sterflinger. 2022. "A Multi-Analytical Approach to Infer Mineral–Microbial Interactions Applied to Petroglyph Sites in the Negev Desert of Israel" Applied Sciences 12, no. 14: 6936. https://doi.org/10.3390/app12146936