
Screening for Antibacterial Activity of French Mushrooms against Pathogenic and Multidrug Resistant Bacteria

 ,
,
Abstract
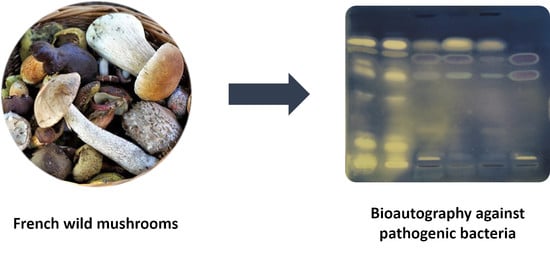
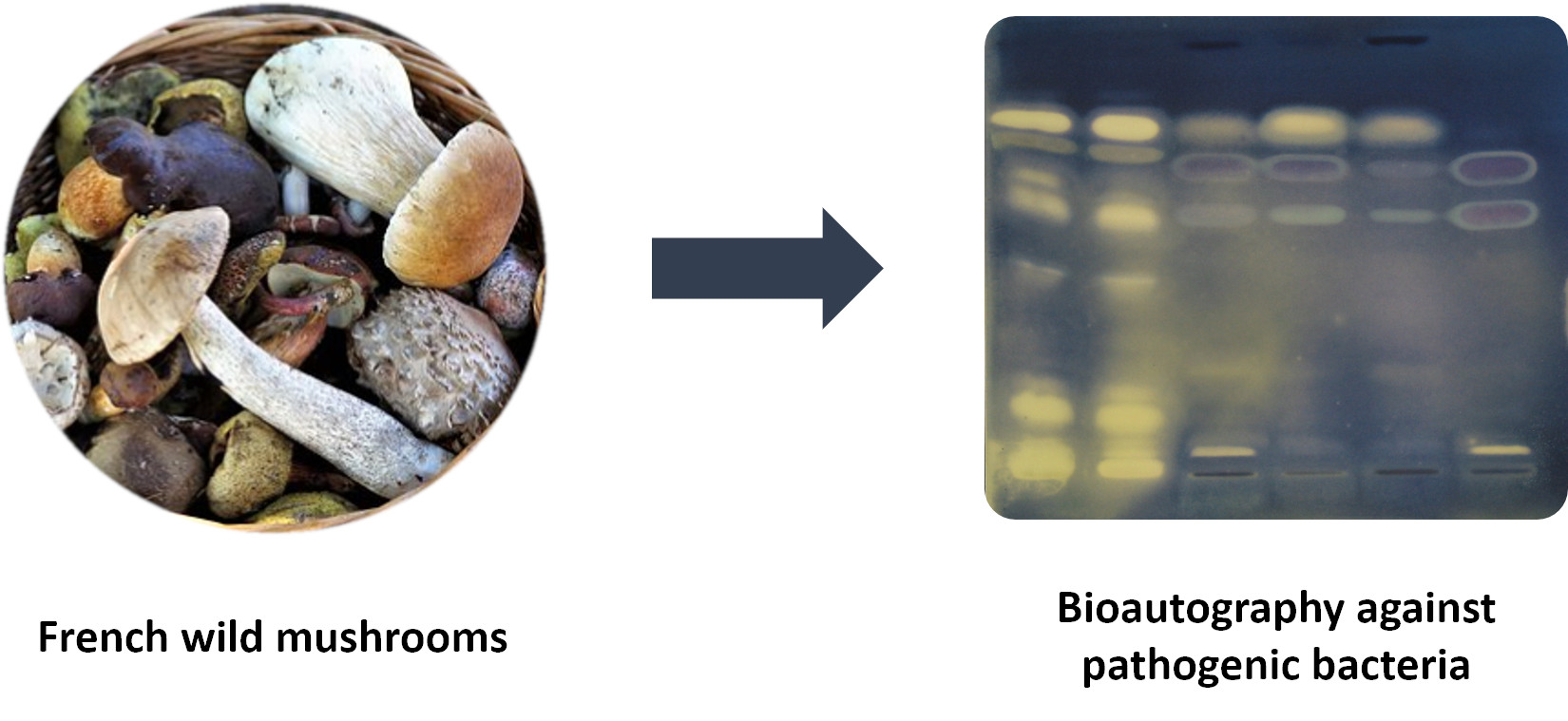
:
1. Introduction
2. Materials and Methods
2.1. Mushrooms
2.2. Chemicals
2.3. Extracts Preparation
2.4. Thin-Layer Chromatography
2.5. Bacterial Culture
2.5.1. Wild-Type Bacteria
2.5.2. Multidrug Resistant Bacterial Strains
2.6. TLC-Direct Bioautography
2.7. Purification and Identification of Stearic Acid
3. Results
3.1. Mushroom Collection
3.2. Antibacterial Activities of Mushroom Extracts against Wild-Type Bacteria
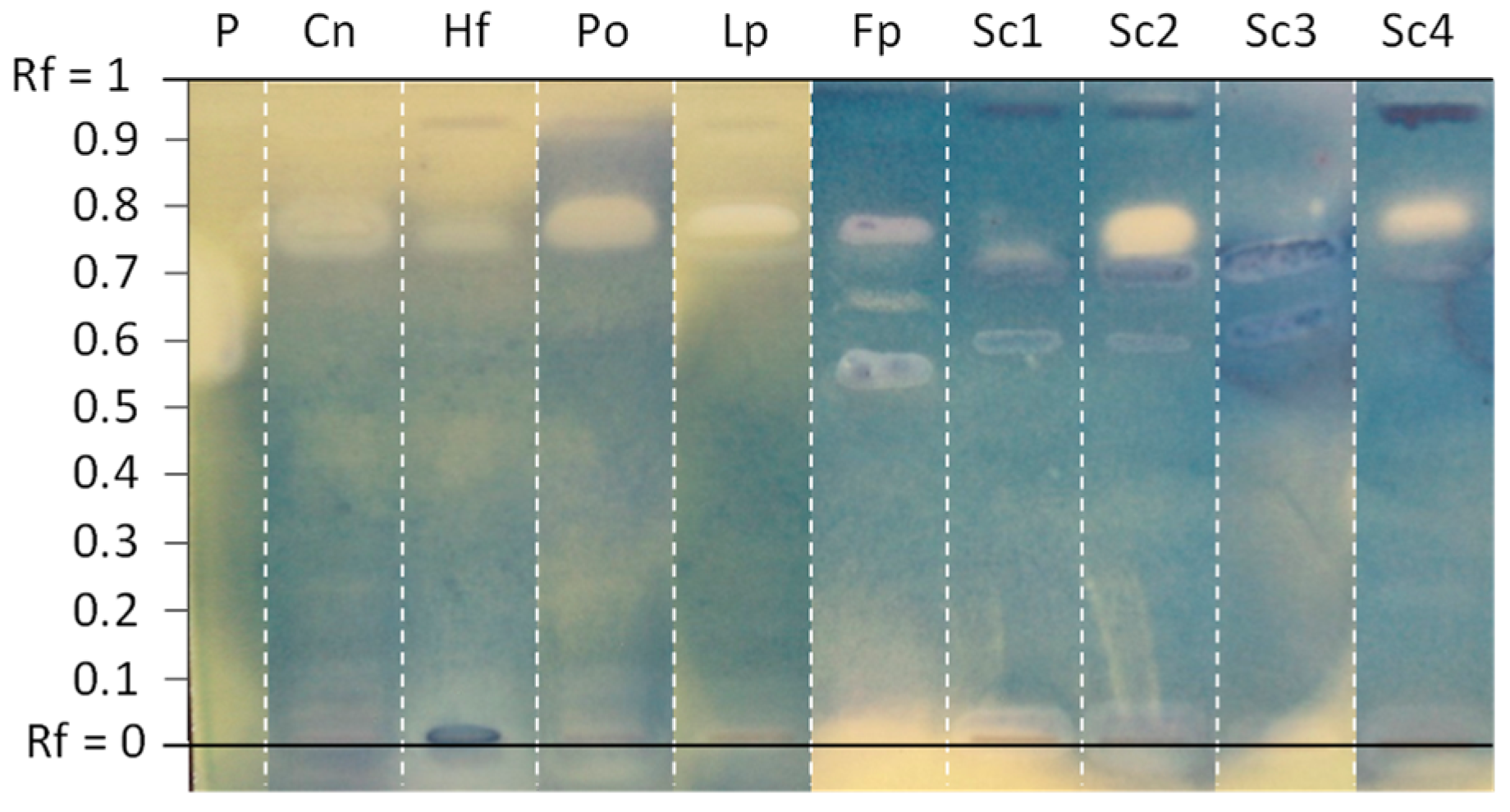
- Fomitopsis pinicola (Polyporales) was selected because both polar and intermediate-polarity extracts were shown to contain several molecules affecting the growth of the five wild-type bacteria.
- The sulphur tuft (Hypholoma fasciculare, Agaricales) was selected because its MeOH extract was one of the few active against P. aeruginosa, with at least four antibacterial compounds.
- Clitocybe nebularis (Agaricales) was included in the bioassay as its EtOAc extract was one of the most active against E. coli, as assessed by the total number of inhibition bands.
- The oyster mushroom (Pleurotus ostreatus, Agaricales) was included for further investigation because of the activity profile of its EtOAc extract against Gram-negative bacteria, and above all to its availability as an edible mushroom.
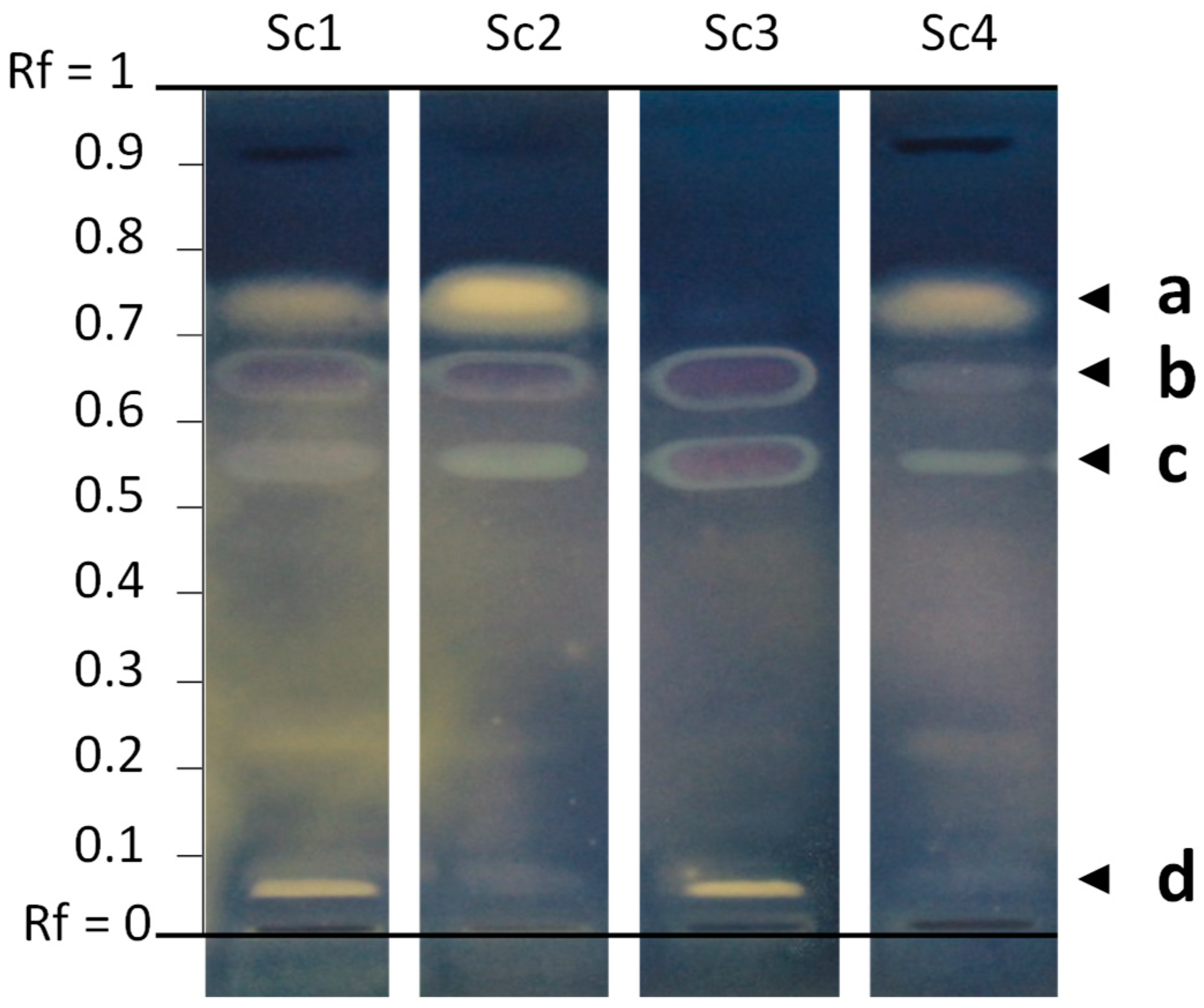
- Regarding the common earthball (Scleroderma citrinum, Boletales), the EtOAc extract of the first specimen (#1) was one of the most active in terms of number of inhibition zones against all bacteria. In addition to the three initial samples screened against reference bacteria, a fourth sample harvested in 2019 was included in the screening against MDR strains.
- Finally, the peppery milkcap (Lactifluus piperatus, Russulales)—that has been used for the extraction of stearic acid—was also included in the bioautography assays against MDR strains.
3.3. Antibacterial Activity against MDR Bacterial Strain
3.3.1. Antibacterial Activity against MDR S. aureus Strains
3.3.2. Antibacterial Activity against MDR E. coli Strains
3.4. Identification of Stearic Acid
4. Discussion
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Antibiotic | Class of Antibiotics | SA1 | SA2 | SA6 | SA8 |
|---|---|---|---|---|---|
| Oxacillin | Penicillins | S | R | R | R |
| Ceftobiprol | Cephalosporin-5G | S | S | S | S |
| Amikacin | Aminoglycosides | R | R | R | R |
| Gentamicin | Aminoglycosides | S | S | S | S |
| Tobramycin | Aminoglycosides | S | R | R | S |
| Ofloxacin | Fluoroquinolones | S | R | R | S |
| Levofloxacin | Fluoroquinolones | S | R | R | S |
| Erythromycin | Macrolides | R | S | S | R |
| Clindamycin | Lincosamides | S | S | ND | R |
| Pristinamycin | Streptogramins | S | S | S | S |
| Tigecycline | Glycylcyclines | S | S | S | S |
| Trimethoprim/sulfamethoxazole | Sulfamide/diaminopyrimidine | S | S | S | S |
| Fosfomycin | Phosphonic acids | S | S | S | S |
| Linezolid | Oxazolidinones | S | S | S | S |
| Rifampicin | Ansamycins | S | S | S | S |
| Fusidic acid | Fusidanes | R | S | R | S |
| Vancomycin | Glycopeptide | ND | ND | S | ND |
| Teicoplanin | Glycopeptide | ND | ND | S | ND |
| Antibiotic | Class of Antibiotics | EC2 | EC5 | EC6 | EC8 |
|---|---|---|---|---|---|
| Amoxicillin | Aminopenicillins | R | R | R | R |
| Amoxicillin/clavulanic acid | Aminopenicillins/β-lactamase inhibitor | R | R | S | R |
| Mecillinam | Amidinopenicillins | S | S | S | R |
| Ticarcillin | Carboxypenicillins | R | R | R | ND |
| Ticarcillin/clavulanic acid | Carboxypenicillins/β-lactamase inhibitor | R | ND | S | ND |
| Temocillin | Carboxypenicillins | ND | ND | ND | ND |
| Piperacillin | Ureidopenicillins | R | R | S | ND |
| Piperacillin/tazobactam | Ureidopenicillins/β-lactamase inhibitor | S | S | S | R |
| Cefoxitin | Cephamycins | S | S | S | S |
| Cefixime | Cephalosporin-3G | ND | R | S | S |
| Ceftriaxone | Cephalosporin-3G | ND | R | S | S |
| Cefotaxime | Cephalosporin-3G | R | ND | ND | ND |
| Ceftazidime | Cephalosporin-3G | R | R | S | S |
| Cefepime | Cephalosporin-4G | R | R | ND | ND |
| Aztreonam | Monobactams | R | R | ND | ND |
| Ertapenem | Carbapenems | S | S | S | S |
| Imipenem | Carbapenems | S | ND | ND | ND |
| Meropenem | Carbapenems | S | S | ND | ND |
| Amikacin | Aminoglycosides | S | S | S | S |
| Gentamicin | Aminoglycosides | R | R | R | R |
| Tobramycin | Aminoglycosides | R | R | ND | ND |
| Nalidixic acid | Quinolones | ND | R | S | R |
| Ciprofloxacin | Fluoroquinolones | R | S | R | R |
| Levofloxacin | Fluoroquinolones | R | R | ND | ND |
| Ofloxacin | Fluoroquinolones | R | R | S | R |
| Trimethoprim/sulfamethoxazole | Sulfamide/diaminopyrimidine | R | R | R | S |
| Fosfomycin | Phosphonic acids | S | R | S | S |
| Nitrofurantoin | Nitrofurans | S | S | S | S |
| E. faecalis | S. aureus | S. epidermidis | E. coli | P. aeruginosa | ||
|---|---|---|---|---|---|---|
| Mushroom Species | Extract | Rf Values | Rf Values | Rf Values | Rf Values | Rf Values |
| Agaricus bisporus | EtOAc | 0.45, 0.81 | 0.41, 0.59, 0.75 | 0.49, 0.81 | 0.06, 0.43, 0.73–0.78 | / |
| MeOH | 0, 0.08, 0.15, 0.75, 0.85 | 0.85 | 0.88 | 0.15, 0.85 | / | |
| Amanita citrina | EtOAc | 0.28, 0.45, 0.78, 0.80–0.84 | 0, 0.40–0.44, 0.70–0.94 | 0.10, 0.49, 0.79–0.84 | 0.25–0.28, 0.41–0.45, 0.86 | 0.78–0.83 |
| MeOH | 0.08, 0.85 | 0.85 | 0.92 | 0.02, 0.70 | / | |
| Amanita muscaria | EtOAc | 0.09, 0.26, 0.37, 0.43, 0.54, 0.58–0.63, 0.75, 0.78–0.83 | 0.04, 0.08, 0.10, 0.63, 0.68, 0.72–0.76, 0.77–0.80 | 0.03, 0.73 | 0–0.10, 0.76 | 0.70 |
| MeOH | 0.80, 0.86 | 0, 0.48 | 0.11, 0.59 | 0.70 | / | |
| Armillaria cepistipesvar. pseudobulbosa | EtOAc | 0.09, 0.26, 0.37, 0.43, 0.54, 0.58–0.63, 0.75, 0.78–0.83 | 0.04, 0.08, 0.10, 0.63, 0.68, 0.72–0.76, 0.77–0.80 | 0.03, 0.73 | 0–0.10, 0.76 | 0.70 |
| MeOH | 0.80, 0.86 | 0, 0.48 | 0.11, 0.59 | 0.70 | / | |
| Armillaria ostoyae | EtOAc | 0.79 | 0.06, 0.09, 0.15, 0.78 | 0.09, 0.80 | 0.04, 0.10, 0.75 | / |
| MeOH | / | / | 0 | / | / | |
| Bolbitius variicolor | EtOAc | 0.47–0.50, 0.84–0.87 | 0.78 | 0.75–0.83 | 0.69 | / |
| MeOH | 0, 0.26, 0.66–0.73 | 0 | 0, 0.80 | 0.03 | / | |
| Clitocybe nebularis | EtOAc | 0.05, 0.75–0.80 | 0.06, 0.15, 0.40, 0,70–0.78 | 0.04, 0.06, 0.10, 0.21, 0.48, 0.65, 0.76–0.84 | 0.03, 0.04, 0.06, 0.18, 0.25, 0.43, 0.73–0.80 | 0.16, 0.79 |
| MeOH | / | / | / | / | / | |
| Cortinarius semisanguineus | EtOAc | 0.63–0.69 | 0–0.05, 0.31, 0.85–0.94 | 0–0.05, 0.31, 0.85–0.94 | / | / |
| MeOH | / | / | 0.11, 0.58 | / | / | |
| Gymnopus luxurians | EtOAc | 0.03, 0.30, 0.50, 0.73–0.78 | / | / | 0.75–0.85 | 0.75–0.81 |
| MeOH | / | / | 0.13, 0.60 | / | / | |
| Hypholoma fasciculare | EtOAc | 0–0.05, 0.48–0.52, 0.73, 0.84–0.87 | 0–0.05, 0.75–0.79 | 0–0.05, 0.60, 0.74 | 0.05, 0.70 | 0 |
| MeOH | 0.04, 0.05, 0.10, 0.34, 0.38 | 0.05, 0.29, 0.39 | 0.20 | / | 0.04, 0.21, 0.31–0.35, 0.41 | |
| Macrolepiota procera | EtOAc | 0.03, 0.06, 0.49, 0.56, 0.80–0.84 | 0.75–0,79, 0.83–0.89 | / | / | / |
| MeOH | / | / | 0.11, 0.59 | / | / | |
| Mycena rosea | EtOAc | 0.04, 0.55, 0.79, 0.81 | 0.04, 0.06, 0.15, 0.70–0.79 | 0.05, 0.10, 0.21, 0.78–0.84 | 0.25, 0.73–0.80 | 0.16, 0.79 |
| MeOH | / | / | / | / | / | |
| Parasola auricoma | EtOAc | 0.09, 0.28, 0.79, 0.81 | 0.05, 0.08, 0.15, 0.41, 0.71–0.94 | 0.21, 0.79–0.83 | 0.44, 0.73–0.78 | 0.79 |
| MeOH | / | / | 0 | / | / | |
| Pleurotus ostreatus | EtOAc | 0.65, 0.78–0.83 | 0.06, 0.15, 0.59, 0.70–0.79 | 0.06, 0.10, 0.13, 0.21, 0.68, 0.79–0.84 | 0.41, 0.63, 0.74–0.80 | 0.79 |
| MeOH | / | / | 0 | / | / | |
| Volvopluteus gloiocephalus | EtOAc | 0.03, 0.08, 0.16, 0.29, 0.38, 0.41, 0.49–0.50, 0.59, 0.64, 0.71, 0.81–0.83 | 0.04, 0.15, 0.55, 0.64–0.66, 0.73–0.75, 0.76–0.81 | 0.61, 0.74, 0.76 | 0.13, 0.43–0.50, 0.69–0.80 | 0.05, 0.76–0.80 |
| MeOH | 0 | / | 0 | / | / |
| E. faecalis | S. aureus | S. epidermidis | E. coli | P. aeruginosa | ||
|---|---|---|---|---|---|---|
| Mushroom Species | Extract | Rf Values | Rf Values | Rf Values | Rf Values | Rf Values |
| Hygrophoropsis aurantiaca | EtOAc | 0.03, 0.08, 0.10, 0.45, 0.79–0.81 | 0.80 | 0.65, 0.80 | 0.03, 0.76 | / |
| MeOH | 0.59 | / | 0, 0.56 | / | / | |
| Neoboletus erythropus #1 | EtOAc | 0, 0.14, 0.41–0.45, 0.53, 0.65, 0.79–0.83 | 0.73–0.76, 0.77–0.81 | 0.63, 0.70 | 0.66, 0.78 | / |
| MeOH | / | / | / | / | / | |
| Neoboletus erythropus #2 | EtOAc | 0.01, 0.10, 0.42, 0.69–0.81 | 0.04, 0.06, 0.45–0.48, 0.60–0.65, 0.69–0.75, 0.89 | 0.04, 0.06, 0.45–0.48, 0.60–0.65, 0.69–0.75, 0.86–0.89 | 0.75–0.79 | 0–0.03, 0.75–0.80 |
| MeOH | / | 0–0.04, 0.70, 0.93 | 0.44 | / | / | |
| Paxillus involutus | EtOAc | 0.03–0.10, 0.15, 0.20, 0.30–0.33, 0.48–0.51, 0.54–0.61, 0.65, 0.71, 0.79–0.86 | 0.04, 0.69–0.73, 0.75–0.79 | 0.79 | 0.80 | 0.70 |
| MeOH | / | / | 0.90 | / | / | |
| Scleroderma citrinum #1 | EtOAc | 0.01, 0.03, 0.04–0.05, 0.15, 0.48–0.50, 0.60–0.63, 0.71, 0.75, 0.79–0.84 | 0, 0.02, 0.04, 0.05, 0.74, 0.79 | 0.05, 0.64, 0.78 | 0.61, 0.71 | 0.05, 0.11, 0.25, 0.43 |
| MeOH | / | / | 0.89 | / | / | |
| Scleroderma citrinum #2 | EtOAc | 0.05–0.08, 0.44, 0.57, 0.63–0.66, 0.69, 0.73, 0.78–0.83 | 0.02, 0.05, 0.56–0.61, 0.65–0.68, 0.71–0.73, 0.76–0.81 | 0.63, 0.68–0.77 | 0.63, 0.69–0.78 | 0.60, 0.69 |
| MeOH | / | / | 0.89 | / | / | |
| Scleroderma citrinum #3 | EtOAc | 0.05, 0.41, 0.49, 0.70 | 0.02, 0.43, 0.61, 0.79 | 0.03 | 0.45, 0.63, 0.71 | / |
| MeOH | / | 0, 0.45, 0.73, 0.77, 0.94 | 0 | 0.13 | / |
| E. faecalis | S. aureus | S. epidermidis | E. coli | P. aeruginosa | ||
|---|---|---|---|---|---|---|
| Mushroom Species | Extract | Rf Values | Rf Values | Rf Values | Rf Values | Rf Values |
| Morchella esculenta #1 | EtOAc | 0.80 | 0.65, 0.77 | 0.84 | / | 0.70 |
| MeOH | 0.65 | 0, 0.30, 0.55–0.56, 0.73 | / | 0.56 | / | |
| Morchella esculenta #2 | EtOAc | 0.03, 0.14, 0.22, 0.78 | / | 0.04, 0.06, 0.10, 0.23 | 0.04, 0.16 | 0.09, 0.19 |
| MeOH | / | 0.69 | / | / | / | |
| Peziza vesiculosa | EtOAc | 0.43, 0.51, 0.58, 0.63, 0.78–0.83 | 0.73–0.76, 0.77–0.81 | 0.74 | 0.78 | / |
| MeOH | 0, 0.64–0.72 | 0, 0.59, 0.81, 0.94 | 0, 0.80 | 0 | 0, 0.88 |
| E. faecalis | S. aureus | S. epidermidis | E. coli | P. aeruginosa | ||
|---|---|---|---|---|---|---|
| Mushroom Species | Extract | Rf Values | Rf Values | Rf Values | RF Values | Rf Values |
| Fomitopsis pinicola | EtOAc | 0.01, 0.03, 0.04, 0.08, 0.15, 0.21, 0.30, 0.37, 0.41, 0.53, 0.56, 0.62, 0.66, 0.71, 0.82, 0.89 | 0–0.05, 0.08, 0.21–0.26, 0.30, 0.48, 0.58, 0.64, 0.73, 0.75–0.79 | 0.04, 0.05, 0.06, 0.41, 0.53, 0.58, 0.63, 0.73–0.76, 0.83 | 0.03, 0.44, 0.64, 0.74 | 0.45, 0.55, 0.63, 0.71–0.74 |
| MeOH | 0.31, 0.34, 0.38, 0.41, 0.50, 0.56, 0.60, 0.75, 0.81 | 0.28–0.35, 0.38, 0.45, 0.60–0.73, 0.76 | 0.28, 0.33, 0.40, 0.53, 0.56, 0.70, 0.71–0.76, 0.80 | 0.24, 0.27, 0.33, 0.39, 0.48, 0.57, 0.62 | 0.03, 0.08, 0.33–0.38, 0.40–0.45, 0.46–0.49, 0.51, 0.54–0.58, 0.59, 0.63, 0.66 | |
| Ganoderma applanatum | EtOAc | 0.19, 0.29, 0.58, 0.82–0.86 | 0.28, 0.54, 0.76–0.81 | 0.75 | 0.72 | / |
| MeOH | 0–0.05 | 0–0.06, 0.63, 0.75–0.81, 0.85–0.90 | 0 | 0.06, 0.23 | / | |
| Meripilus giganteus | EtOAc | 0.03, 0.08, 0.19, 0.45, 0.55, 0.66, 0.80–0.83 | 0, 0.05, 0.55, 0.81 | 0.63, 0.74 | 0.03, 0.74 | / |
| MeOH | / | / | 0 | 0 | / | |
| Polyporus umbellatus | EtOAc | 0.03, 0.08, 0.18, 0.28, 0.43–0.45, 0.60, 0.68, 0.79–0.83 | 0.05, 0.68, 0.81 | 0.18, 0.25, 0.43, 0.81 | 0.03, 0.43, 0.71–0.81 | / |
| MeOH | 0.75, 0.78 | 0 | 0.11, 0.59 | / | / | |
| Trametes versicolor | EtOAc | 0.28, 0.79–0.84 | 0.16, 0.43, 0.60–0.69, 0.75–0.84 | 0.18, 0.21, 0.66, 0.81 | 0.31, 0.73–0.79 | / |
| MeOH | / | 0.18 | 0.24 | / | / |
| E. faecalis | S. aureus | S. epidermidis | E. coli | P. aeruginosa | ||
|---|---|---|---|---|---|---|
| Mushroom Species | Extract | Rf Values | Rf Values | Rf Values | Rf Values | Rf Values |
| Lactarius helvus | EtOAc | 0.04, 0.22, 0.41, 0.75–0.84 | 0.06, 0.15, 0.22, 0.40, 0.61–0.66, 0.75–0.83 | 0.06, 0.77, 0.83–0.89 | 0.01, 0.04, 0.08, 0.24, 0.41, 0.69 | 0.66–0.70, 0.80–0.84 |
| MeOH | / | 0 | 0 | 0 | / | |
| Lactifluus piperatus | EtOAc | 0.05, 0.43, 0.80–0.85 | 0.77–0.83, 0.86 | 0.25, 0.40, 0.76 | 0.48, 0.81 | 0.75–0.76 |
| MeOH | 0.03, 0.79 | / | / | / | / | |
| Russula integra | EtOAc | 0.03, 0.06, 0.08, 0.16, 0.28, 0.44–0.46, 0, 80–0.86 | 0.76–0.81 | 0.25, 0.40, 0.76 | 0.80 | 0.79 |
| MeOH | 0, 0.86 | / | 0–0.05, 0.79–0.84 | / | / | |
| Russula lepida | EtOAc | 0, 0.16–0.20, 0.29, 0.68, 0.79–0.83 | 0, 0.05, 0.08, 0.19, 0.60, 0.66, 0.70, 0.79–0.85 | 0.81–0.84 | 0.04, 0.19, 0.54, 0.60, 0.78 | 0.19, 0.81–0.86 |
| MeOH | / | / | 0, 0.68 | / | / | |
| Scutiger pes-caprae | EtOAc | 0.03, 0.07, 0.15, 0.31, 0.50, 0.68, 0.80–0.86 | 0.75, 0.80–0.85 | 0.05, 0.64, 0.76–0.81 | 0.80 | / |
| MeOH | 0.01 | 0 | 0–0.06 | 0.95 | / |
References
- Centers for Disease Control and Prevention (U.S.). Antibiotic Resistance Threats in the United States; Centers for Disease Control and Prevention: Atlanta, GA, USA, 2019.
- Murray, C.J.; Ikuta, K.S.; Sharara, F.; Swetschinski, L.; Aguilar, G.R.; Gray, A.; Han, C.; Bisignano, C.; Rao, P.; Wool, E.; et al. Global Burden of Bacterial Antimicrobial Resistance in 2019: A Systematic Analysis. Lancet 2022, 399, 629–655. [Google Scholar] [CrossRef]
- World Health Organization. Global Priority List of Antibiotic-Resistant Bacteria to Guide Research, Discovery, and Development of New Antibiotics; World Health Organization: Geneva, Switzerland, 2017; p. 7. [Google Scholar]
- Hamers, V.; Huguet, C.; Bourjot, M.; Urbain, A. Antibacterial Compounds from Mushrooms: A Lead to Fight ESKAPEE Pathogenic Bacteria? Planta Med. 2020, 87, 351–367. [Google Scholar] [CrossRef] [PubMed]
- Breitenbach, J.; Kränzlin, F. Champignons de Suisse—Contribution à La Connaissance de La Flore Fongique de Suisse; Mykologia Luzern: Lucerne, Switzerland, 1981. [Google Scholar]
- Eyssartier, G.; Roux, P. Guide Des Champignons—France et Europe, 4th ed.; Éditions Belin: Paris, France, 2017. [Google Scholar]
- EUCAST MIC Distributions and the Setting of Epidemiological Cutoff (ECOFF) Values. Standard Operating Procedure 10.1 2019. Available online: https://www.eucast.org/fileadmin/src/media/PDFs/EUCAST_files/EUCAST_SOPs/EUCAST_SOP_10.1_MIC_distributions_and_epidemiological_cut-off_value__ECOFF__setting_20191130.pdf (accessed on 6 June 2021).
- Köck, R.; Becker, K.; Cookson, B.; van Gemert-Pijnen, J.E.; Harbarth, S.; Kluytmans, J.; Mielke, M.; Peters, G.; Skov, R.L.; Struelens, M.J.; et al. Methicillin-Resistant Staphylococcus aureus (MRSA): Burden of Disease and Control Challenges in Europe. Eurosurveillance 2010, 15, 19688. [Google Scholar] [CrossRef] [PubMed]
- Morel, S.; Vitou, M.; Masnou, A.; Jumas-Bilak, E.; Rapior, S.; Licznar-Fajardo, P. Antibacterial Activity of Wild Mushrooms from France. Int. J. Med. Mushrooms 2021, 23, 79–89. [Google Scholar] [CrossRef]
- Clericuzio, M.; Bivona, M.; Gamalero, E.; Bona, E.; Novello, G.; Massa, N.; Dovana, F.; Marengo, E.; Robotti, E. A Systematic Study of the Antibacterial Activity of Basidiomycota Crude Extracts. Antibiotics 2021, 10, 1424. [Google Scholar] [CrossRef] [PubMed]
- Urbain, A.; Simões-Pires, C.A. Thin-Layer Chromatography for the Detection and Analysis of Bioactive Natural Products. In Encyclopedia of Analytical Chemistry; American Cancer Society: Atlanta, GA, USA, 2020; pp. 1–29. ISBN 978-0-470-02731-8. [Google Scholar]
- Walton, J. The Cyclic Peptide Toxins of Amanita and Other Poisonous Mushrooms; Springer International Publishing: Cham, Switzerland, 2018; ISBN 978-3-319-76821-2. [Google Scholar]
- Casillas-Vargas, G.; Ocasio-Malavé, C.; Medina, S.; Morales-Guzmán, C.; Del Valle, R.G.; Carballeira, N.M.; Sanabria-Ríos, D.J. Antibacterial Fatty Acids: An Update of Possible Mechanisms of Action and Implications in the Development of the next-Generation of Antibacterial Agents. Prog. Lipid Res. 2021, 82, 101093. [Google Scholar] [CrossRef] [PubMed]
- Desbois, A.P.; Smith, V.J. Antibacterial Free Fatty Acids: Activities, Mechanisms of Action and Biotechnological Potential. Appl. Microbiol. Biotechnol. 2010, 85, 1629–1642. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Cheung Lam, A.H.; Sandoval, N.; Wadhwa, R.; Gilkes, J.; Do, T.Q.; Ernst, W.; Chiang, S.-M.; Kosina, S.; Howard Xu, H.; Fujii, G.; et al. Assessment of Free Fatty Acids and Cholesteryl Esters Delivered in Liposomes as Novel Class of Antibiotic. BMC Res. Notes 2016, 9, 337. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grienke, U.; Zöll, M.; Peintner, U.; Rollinger, J.M. European Medicinal Polypores—A Modern View on Traditional Uses. J. Ethnopharmacol. 2014, 154, 564–583. [Google Scholar] [CrossRef] [PubMed]
- Grienke, U.; Foster, P.A.; Zwirchmayr, J.; Tahir, A.; Rollinger, J.M.; Mikros, E. 1H NMR-MS-Based Heterocovariance as a Drug Discovery Tool for Fishing Bioactive Compounds out of a Complex Mixture of Structural Analogues. Sci. Rep. 2019, 9, 11113. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdullah, S.; Oh, Y.S.; Kwak, M.-K.; Chong, K. Biophysical Characterization of Antibacterial Compounds Derived from Pathogenic Fungi Ganoderma Boninense. J. Microbiol. 2021, 59, 164–174. [Google Scholar] [CrossRef] [PubMed]
- Liu, X.-T.; Winkler, A.L.; Schwan, W.R.; Volk, T.J.; Rott, M.A.; Monte, A. Antibacterial Compounds from Mushrooms I: A Lanostane-Type Triterpene and Prenylphenol Derivatives from Jahnoporus Hirtus and Albatrellus Flettii and Their Activities Against Bacillus Cereus and Enterococcus faecalis. Planta Med. 2010, 76, 182–185. [Google Scholar] [CrossRef] [PubMed]





| Order | Family | Species | Harvest Information * |
|---|---|---|---|
| Agaricales | Agaricaceae | Agaricus bisporus | 2016 February, from supermarket |
| Macrolepiota procera | 2015 October, Fuschloch | ||
| Amanitaceae | Amanita citrina | 2014 October, Haguenau | |
| Amanita muscaria | 2015 October, Dabo | ||
| Bolbitiaceae | Bolbitius variicolor | 2016 April, Illkirch | |
| Cortinariaceae | Cortinarius semisanguineus | 2015 November, Wisches | |
| Mycenaceae | Mycena rosea | 2017 Octpber, Ohlungen | |
| Omphalotaceae | Gymnopus luxurians | 2016 June, Illkirch | |
| Physalacriaceae | Armillaria cepistipes var. pseudobulbosa | 2015 October, Illkirch | |
| Armillaria ostoyae | 2018 October, Floessplatz | ||
| Pleurotaceae | Pleurotus ostreatus | 2018 November, Barr | |
| Pluteaceae | Volvopluteus gloiocephalus | 2016 April, Illkirch | |
| Psathyrellaceae | Parasola auricoma | 2016 June, Illkirch | |
| Strophariaceae | Hypholoma fasciculare | 2015 October, Fuschloch | |
| Tricholomataceae | Clitocybe nebularis | 2017 October, Ried | |
| Boletales | Boletaceae | Neoboletus erythropus #1 | 2018 October, La Fischhutte |
| Neoboletus erythropus #2 | 2017 August, Saint-Jean-Saverne | ||
| Hygrophoropsidaceae | Hygrophoropsis auriantaca | 2016 October, Hohbuhl | |
| Paxillaceae | Paxillus involutus | 2015 October, Dabo | |
| Sclerodermataceae | Scleroderma citrinum #1 | 2018 October, Balbronn | |
| Scleroderma citrinum #2 | 2018 October, Floessplatz | ||
| Scleroderma citrinum #3 | 2018 October, Floessplatz | ||
| Scleroderma citrinum #4 | 2019 September, Balbronn | ||
| Pezizales | Morchellaceae | Morchella esculenta #1 | 2019 April, Erstein |
| Morchella esculenta #2 | 2019 April, Plobsheim | ||
| Pezizaceae | Peziza vesiculosa | 2016 April, Illkirch | |
| Polyporales | Fomitopsidaceae | Fomitopsis pinicola | 2014 October, Barr |
| Ganodermataceae | Ganoderma applanatum | 2018 October, Balbronn | |
| Meripilaceae | Meripilus giganteus | 2017 August, Saint-Jean-Saverne | |
| Polyporaceae | Polyporus umbellatus | 2017 July, Still | |
| Trametes versicolor | 2018 August, Saint-Jean-Saverne | ||
| Russulales | Albatrellaceae | Scutiger pes-caprae | 2018 April, Fackental |
| Russulaceae | Lactarius helvus | 2017 September, Haguenau | |
| Lactifluus piperatus | 2017 July, Still | ||
| Russula integra | 2017 August, Still | ||
| Russula lepida | 2017 August, Saint-Jean-Saverne |
| EtOAc Extracts | MeOH Extracts | ||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| Order | Mushroom Species | Ef | Sa | Se | Ec | Pa | Ef | Sa | Se | Ec | Pa |
| Agaricales | Agaricus bisporus | 2 | 3 | 2 | 3 | 0 | 5 | 1 | 1 | 2 | 0 |
| Amanita citrina | 4 | 3 | 3 | 3 | 1 | 2 | 1 | 1 | 2 | 0 | |
| Amanita muscaria | 8 | 7 | 2 | 2 | 1 | 2 | 2 | 2 | 1 | 0 | |
| Armillaria cepistipes var. pseudobulbosa | 12 | 5 | 5 | 1 | 0 | 1 | 1 | 1 | 0 | 0 | |
| Armillaria ostoyae | 1 | 4 | 2 | 3 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Bolbitius variicolor | 2 | 1 | 2 | 1 | 0 | 3 | 1 | 2 | 1 | 0 | |
| Clitocybe nebularis | 2 | 4 | 7 | 7 | 2 | 0 | 0 | 0 | 0 | 0 | |
| Cortinarius semisanguineus | 1 | 3 | 3 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | |
| Gymnopus luxurians | 4 | 0 | 0 | 1 | 1 | 0 | 0 | 2 | 0 | 0 | |
| Hypholoma fasciculare | 4 | 2 | 3 | 2 | 1 | 5 | 3 | 1 | 0 | 4 | |
| Macrolepiota procera | 5 | 2 | 0 | 0 | 0 | 0 | 0 | 2 | 0 | 0 | |
| Mycena rosea | 4 | 4 | 4 | 2 | 2 | 0 | 0 | 0 | 0 | 0 | |
| Parasola auricoma | 4 | 5 | 2 | 2 | 1 | 0 | 0 | 1 | 0 | 0 | |
| Pleurotus ostreatus | 2 | 4 | 6 | 3 | 1 | 0 | 0 | 1 | 0 | 0 | |
| Volvopluteus gloiocephalus | 11 | 6 | 3 | 3 | 2 | 0 | 0 | 1 | 0 | 0 | |
| Boletales | Hygrophoropsis aurantiaca | 5 | 1 | 2 | 2 | 0 | 1 | 0 | 2 | 0 | 0 |
| Neoboletus erythropus #1 | 6 | 2 | 2 | 2 | 0 | 0 | 0 | 0 | 0 | 0 | |
| Neoboletus erythropus #2 | 4 | 6 | 6 | 1 | 2 | 0 | 3 | 1 | 0 | 0 | |
| Paxillus involutus | 9 | 3 | 1 | 1 | 1 | 0 | 0 | 1 | 0 | 0 | |
| Scleroderma citrinum #1 | 9 | 6 | 3 | 2 | 4 | 0 | 0 | 1 | 0 | 0 | |
| Scleroderma citrinum #2 | 7 | 6 | 2 | 2 | 2 | 0 | 0 | 1 | 0 | 0 | |
| Scleroderma citrinum #3 | 4 | 4 | 1 | 3 | 0 | 0 | 5 | 1 | 1 | 0 | |
| Pezizales | Morchella esculenta #1 | 1 | 2 | 1 | 0 | 1 | 1 | 4 | 0 | 1 | 0 |
| Morchella esculenta #2 | 4 | 0 | 4 | 2 | 2 | 0 | 1 | 0 | 0 | 0 | |
| Peziza vesiculosa | 5 | 2 | 1 | 1 | 0 | 2 | 4 | 2 | 1 | 2 | |
| Polyporales | Fomitopsis pinicola | 14 | 10 | 4 | 5 | 3 | 5 | 1 | 8 | 3 | 3 |
| Ganoderma applanatum | 4 | 3 | 1 | 1 | 0 | 1 | 4 | 1 | 2 | 0 | |
| Meripilus giganteus | 7 | 4 | 2 | 2 | 0 | 0 | 0 | 2 | 1 | 0 | |
| Polyporus umbellatus | 8 | 3 | 4 | 3 | 0 | 2 | 1 | 2 | 0 | 0 | |
| Trametes versicolor | 2 | 4 | 4 | 2 | 0 | 0 | 1 | 1 | 0 | 0 | |
| Russulales | Lactarius helvus | 4 | 6 | 3 | 6 | 2 | 0 | 1 | 1 | 1 | 0 |
| Lactifluus piperatus | 3 | 2 | 3 | 2 | 1 | 2 | 0 | 0 | 0 | 0 | |
| Russula integra | 7 | 1 | 3 | 1 | 1 | 2 | 0 | 2 | 0 | 0 | |
| Russula lepida | 5 | 8 | 1 | 5 | 2 | 0 | 0 | 2 | 0 | 0 | |
| Scutiger pes-caprae | 7 | 2 | 3 | 1 | 0 | 1 | 1 | 1 | 1 | 0 | |
| Mushroom Species | Extract | SA1 | SA2 | SA6 | SA8 |
|---|---|---|---|---|---|
| Clitocybe nebularis | EtOAc | 0.75 | 0.25, 0.45, 0.50, 0.75 | 0.75 | 0.40, 0.75 |
| MeOH | / | / | / | / | |
| Hypholoma fasciculare | EtOAc | 0.01, 0.75 | 0.01, 0.75 | 0.75 | 0.01, 0.75 |
| MeOH | / | 0.08 | / | 0.08 | |
| Pleurotus ostreatus | EtOAc | 0.75 | 0.25, 0.65, 0.75 | 0.40, 0.65, 0.75 | 0.10, 0.20, 0.65, 0.75 |
| MeOH | / | / | / | / | |
| Lactifluus piperatus | EtOAc | 0.75 | 0.75 | 0.75 | 0.01, 0.75 |
| MeOH | / | 0.70 | 0.05, 0.70 | 0.70 | |
| Fomitopsis pinicola | EtOAc | 0.01, 0.10, 0.35, 0.50, 0.70, 0.75, 0.85 | 0.01, 0.05, 0.08, 0.10, 0.18, 0.40, 0.55, 0.70, 0.75 | 0.01, 0.45, 0.62, 0.70, 0.75 | 0.01, 0.08, 0.40, 0.55, 0.65, 0.70, 0.75 |
| MeOH | 0.55, 0.65, 0.70, 0.80 | 0.60, 0.70, 0.78, 0.85 | 0.60, 0.70, 0.78, 0.85 | 0.50, 0.65, 0.70, 0.85 | |
| Scleroderma citrinum #1 | EtOAc | 0.08, 0.50, 0.60, 0.70 | 0.08, 0.55, 0.65, 0.75 | 0.08, 0.60, 0.70, 0.75 | 0.08, 0.50, 0.60, 0.70, 0.75 |
| MeOH | 0.01 | 0.01 | 0.01 | 0.01 | |
| Scleroderma citrinum #2 | EtOAc | 0.50, 0.60, 0.70 | 0.55, 0.65, 0.75 | 0.60, 0.70, 0.75 | 0.60, 0.70, 0.75 |
| MeOH | 0.90 | 0.50, 0.90 | 0.01 | 0.01 | |
| Scleroderma citrinum #3 | EtOAc | 0.08, 0.50, 0.60, | 0.08, 0.55, 0.65 | 0.08, 0.60, 0.70 | 0.08, 0.60, 0.70 |
| MeOH | 0.01 | 0.50 | 0.01, 0.35, 0.80 | 0.01 | |
| Scleroderma citrinum #4 | EtOAc | 0.60, 0.70 | 0.55, 0.65, 0.75 | 0.60, 0.70, 0.75 | 0.60, 0.70, 0.75 |
| MeOH | / | / | 0.01 | 0.01 |
| Mushroom Species | Extract | EC2 | EC5 | EC6 | EC8 |
|---|---|---|---|---|---|
| Clitocybe nebularis | EtOAc | 0.78 | 0.78 | 0.78 | 0.78 |
| MeOH | / | / | / | / | |
| Hypholoma fasciculare | EtOAc | / | 0.78 | 0.78 | 0.78 |
| MeOH | / | / | / | / | |
| Pleurotus ostreatus | EtOAc | 0.78 | 0.78 | 0.40, 0.62 | 0.78 |
| MeOH | / | / | / | / | |
| Lactifluus piperatus | EtOAc | 0.78 | 0.78 | 0.78 | 0.78 |
| MeOH | / | / | / | / | |
| Fomitopsis pinicola | EtOAc | / | 0.55, 0.65, 0.78 | 0.40, 0.55 | 0.55 |
| MeOH | 0.70, 0.85 | 0.65 | 0.50, 0.60, 0.62, 0.80 | 0.55, 0.65, 0.70, 0.82 | |
| Scleroderma citrinum #1 | EtOAc | 0.60, 0.70 | 0.01, 0.60 | 0.60, 0.78 | 0.70 |
| MeOH | 0.01 | / | 0.40 | 0.01 | |
| Scleroderma citrinum #2 | EtOAc | 0.60, 0.70, 0.75 | 0.01, 0.60, 0.78 | 0.60, 0.78 | 0.60, 0.70, 0.75 |
| MeOH | 0.90 | / | 0.40 | 0.42 | |
| Scleroderma citrinum #3 | EtOAc | 0.60, 0.70 | 0.60 | 0.60 | 0.60 |
| MeOH | 0.40, 0.45 | / | 0.40 | 0.40 | |
| Scleroderma citrinum #4 | EtOAc | 0.75 | 0.78 | / | 0.75 |
| MeOH | 0.01 | / | / | 0.01 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Huguet, C.; Bourjot, M.; Bellanger, J.-M.; Prévost, G.; Urbain, A. Screening for Antibacterial Activity of French Mushrooms against Pathogenic and Multidrug Resistant Bacteria. Appl. Sci. 2022, 12, 5229. https://doi.org/10.3390/app12105229
Huguet C, Bourjot M, Bellanger J-M, Prévost G, Urbain A. Screening for Antibacterial Activity of French Mushrooms against Pathogenic and Multidrug Resistant Bacteria. Applied Sciences. 2022; 12(10):5229. https://doi.org/10.3390/app12105229
Chicago/Turabian StyleHuguet, Clément, Mélanie Bourjot, Jean-Michel Bellanger, Gilles Prévost, and Aurélie Urbain. 2022. "Screening for Antibacterial Activity of French Mushrooms against Pathogenic and Multidrug Resistant Bacteria" Applied Sciences 12, no. 10: 5229. https://doi.org/10.3390/app12105229