MiR155-5p Inhibits Cell Migration and Oxidative Stress in Vascular Smooth Muscle Cells of Spontaneously Hypertensive Rats

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Culture of Primary VSMCs
2.3. Boyden Chamber Assay
2.4. Wound-Healing Assay
2.5. Transfection of miR155-5p Mimic and Inhibitor
2.6. MiR155-5p Overexpression and Ang II Treatment in VSMCs
2.7. Western Blot Analysis
2.8. Measurement of miR155-5p Level with qPCR
2.9. Measurement of ACE mRNA Level with qPCR
2.10. DHE Fluorescence Staining
2.11. Measurement of NAD(P)H Oxidase Activity
2.12. Statistics and Data Analysis
3. Results
3.1. Effects of miR155-5p Mimic and Inhibitor on VSMC Migration
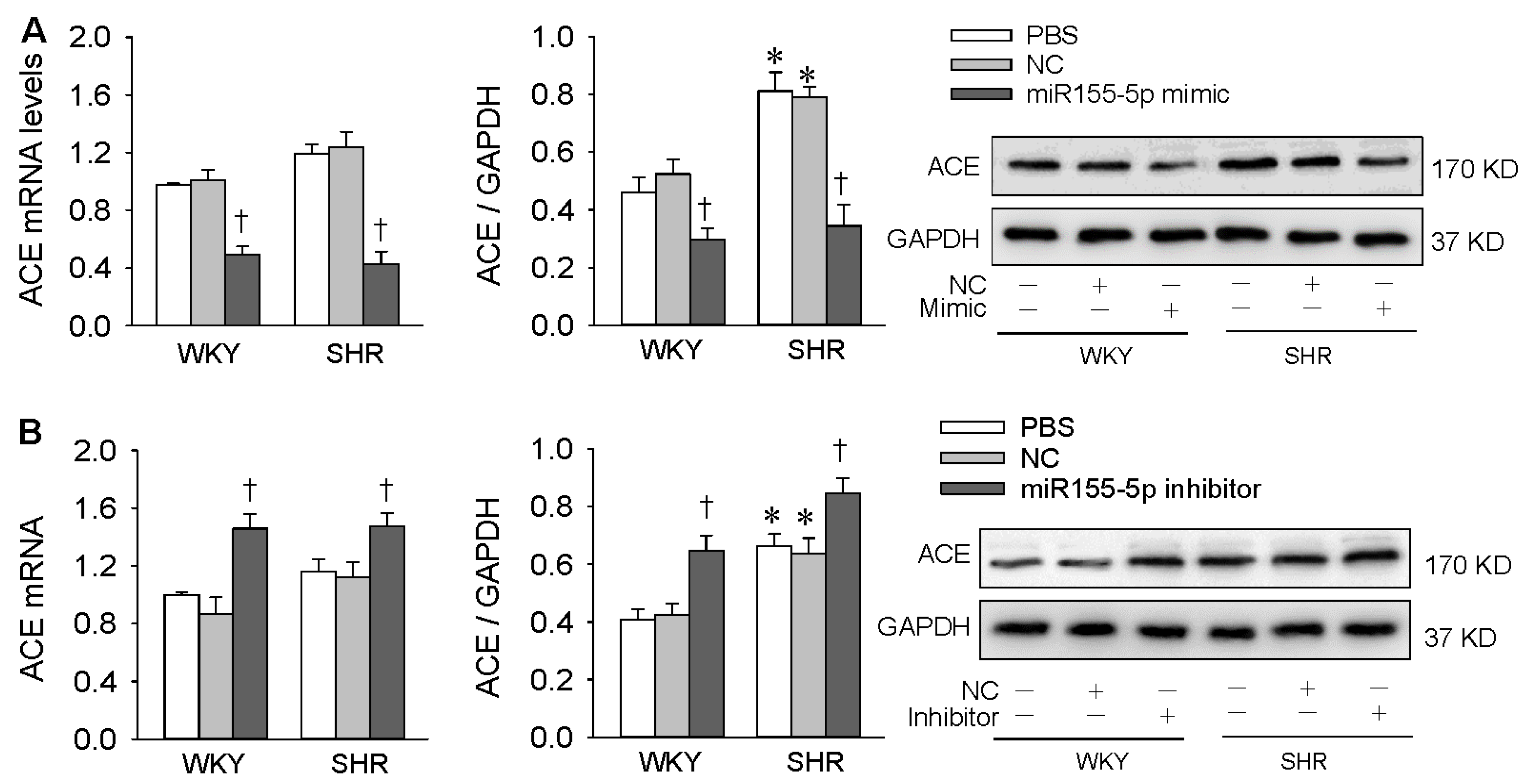
3.2. Effects of miR155-5p Mimic and Inhibitor on ACE Expression
3.3. Effects of miR155-5p Mimic on Oxidative Stress
3.4. Effects of miR155-5p Mimic on IL-1β and TNF-α Levels
3.5. Effects of miR155-5p Overexpression on Ang II-Induced VSMC Migration
3.6. Effects of miR155-5p Overexpression on Ang II-Induced Oxidative Stress and Inflammation
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Conflicts of Interest
Data Availability
References
- Cai, X. Regulation of smooth muscle cells in development and vascular disease: Current therapeutic strategies. Expert Rev. Cardiovasc. Ther. 2006, 4, 789–800. [Google Scholar] [CrossRef]
- Chou, C.C.; Wang, C.P.; Chen, J.H.; Lin, H.H. Anti-atherosclerotic effect of hibiscus leaf polyphenols against tumor necrosis factor-a-induced abnormal vascular smooth muscle cell migration and proliferation. Antioxidants 2019, 8, 620. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Tajsic, T.; Morrell, N.W. Smooth muscle cell hypertrophy, proliferation, migration and apoptosis in pulmonary hypertension. Compr. Physiol. 2011, 1, 295–317. [Google Scholar] [PubMed]
- Willis, A.I.; Pierre-Paul, D.; Sumpio, B.E.; Gahtan, V. Vascular smooth muscle cell migration: Current research and clinical implications. Vasc. Endovascular. Surg. 2004, 38, 11–23. [Google Scholar] [CrossRef] [PubMed]
- Obradovic, M.; Essack, M.; Zafirovic, S.; Sudar-Milovanovic, E.; Bajic, V.P.; Van, N.C.; Trpkovic, A.; Stanimirovic, J.; Bajic, V.B.; Isenovic, E.R. Redox control of vascular biology. Biofactors 2019, 2019, 3764252. [Google Scholar] [CrossRef]
- Pellegrino, D.; La, R.D.; Marrone, A. Oxidative imbalance and kidney damage: New study perspectives from animal models to hospitalized patients. Antioxidants 2019, 8, 594. [Google Scholar] [CrossRef] [Green Version]
- Schieber, M.; Chandel, N.S. ROS function in redox signaling and oxidative stress. Curr. Biol. 2014, 24, R453–R462. [Google Scholar] [CrossRef] [Green Version]
- Sena, C.M.; Leandro, A.; Azul, L.; Seica, R.; Perry, G. Vascular oxidative stress: Impact and therapeutic approaches. Front. Physiol. 2018, 9, 1668. [Google Scholar] [CrossRef] [Green Version]
- Durgin, B.G.; Straub, A.C. Redox control of vascular smooth muscle cell function and plasticity. Lab. Investig. 2018, 98, 1254–1262. [Google Scholar] [CrossRef]
- Huetsch, J.C.; Suresh, K.; Shimoda, L.A. Regulation of smooth muscle cell proliferation by NADPH oxidases in pulmonary hypertension. Antioxidants 2019, 8, 56. [Google Scholar] [CrossRef] [Green Version]
- Ismaeel, A.; Brumberg, R.S.; Kirk, J.S.; Papoutsi, E.; Farmer, P.J.; Bohannon, W.T.; Smith, R.S.; Eidson, J.L.; Sawicki, I.; Koutakis, P. Oxidative stress and arterial dysfunction in peripheral artery disease. Antioxidants 2018, 7, 145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guzik, T.J.; Touyz, R.M. Oxidative Stress, Inflammation, and Vascular Aging in Hypertension. Hypertension 2017, 70, 660–667. [Google Scholar] [CrossRef] [PubMed]
- Solak, Y.; Afsar, B.; Vaziri, N.D.; Aslan, G.; Yalcin, C.E.; Covic, A.; Kanbay, M. Hypertension as an autoimmune and inflammatory disease. Hypertens. Res. 2016, 39, 567–573. [Google Scholar] [CrossRef]
- Brito, R.; Castillo, G.; Gonzalez, J.; Valls, N.; Rodrigo, R. Oxidative stress in hypertension: Mechanisms and therapeutic opportunities. Exp. Clin. Endocrinol. Diabetes 2015, 123, 325–335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hartmann, D.; Fiedler, J.; Sonnenschein, K.; Just, A.; Pfanne, A.; Zimmer, K.; Remke, J.; Foinquinos, A.; Butzlaff, M.; Schimmel, K.; et al. MicroRNA-based therapy of GATA2-deficient vascular disease. Circulation 2016, 134, 1973–1990. [Google Scholar] [CrossRef] [PubMed]
- Liu, Y.; Pan, Q.; Zhao, Y.; He, C.; Bi, K.; Chen, Y.; Zhao, B.; Chen, Y.; Ma, X. MicroRNA-155 regulates ROS production, NO generation, apoptosis and multiple functions of human brain microvessel endothelial cells under physiological and pathological conditions. J. Cell Biochem. 2015, 116, 2870–2881. [Google Scholar] [CrossRef] [PubMed]
- Ren, X.S.; Tong, Y.; Qiu, Y.; Ye, C.; Wu, N.; Xiong, X.Q.; Wang, J.J.; Han, Y.; Zhou, Y.B.; Zhang, F.; et al. MiR155-5p in adventitial fibroblasts-derived extracellular vesicles inhibits vascular smooth muscle cell proliferation via suppressing angiotensin-converting enzyme expression. J. Extracell. Vesicles. 2019, 9, 1698795. [Google Scholar] [CrossRef] [Green Version]
- Tong, Y.; Ye, C.; Ren, X.S.; Qiu, Y.; Zang, Y.H.; Xiong, X.Q.; Wang, J.J.; Chen, Q.; Li, Y.H.; Kang, Y.M.; et al. Exosome-mediated transfer of ACE (angiotensin-converting enzyme) from adventitial fibroblasts of spontaneously hypertensive rats promotes vascular smooth muscle cell migration. Hypertension 2018, 72, 881–888. [Google Scholar] [CrossRef] [PubMed]
- Ling, L.; Chen, D.; Tong, Y.; Zang, Y.H.; Ren, X.S.; Zhou, H.; Qi, X.H.; Chen, Q.; Li, Y.H.; Kang, Y.M.; et al. Fibronectin type III domain containing 5 attenuates NLRP3 inflammasome activation and phenotypic transformation of adventitial fibroblasts in spontaneously hypertensive rats. J. Hypertens. 2018, 36, 1104–1114. [Google Scholar] [CrossRef] [PubMed]
- Sun, H.J.; Zhao, M.X.; Ren, X.S.; Liu, T.Y.; Chen, Q.; Li, Y.H.; Kang, Y.M.; Wang, J.J.; Zhu, G.Q. Salusin-b promotes vascular smooth muscle cell migration and intimal hyperplasia after vascular injury via ROS/NFkB/MMP-9 pathway. Antioxid. Redox. Signal. 2016, 24, 1045–1057. [Google Scholar] [CrossRef]
- Sheu, M.L.; Shen, C.C.; Chen, Y.S.; Chiang, C.K. Ochratoxin A induces ER stress and apoptosis in mesangial cells via a NADPH oxidase-derived reactive oxygen species-mediated calpain activation pathway. Oncotarget 2017, 8, 19376–19388. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ciarcia, R.; Damiano, S.; Florio, A.; Spagnuolo, M.; Zacchia, E.; Squillacioti, C.; Mirabella, N.; Florio, S.; Pagnini, U.; Garofano, T.; et al. The protective effect of apocynin on cyclosporine A-induced hypertension and nephrotoxicity in rats. J. Cell Biochem. 2015, 116, 1848–1856. [Google Scholar] [CrossRef] [PubMed]
- Xu, S.; Zhao, Y.; Jin, C.; Yu, L.; Ding, F.; Fu, G.; Zhu, J. PKC/NADPH oxidase are involved in the protective effect of pioglitazone in high homocysteine-induced paracrine dyfunction in endothelial progenitor cells. Am. J. Transl. Res. 2017, 9, 1037–1048. [Google Scholar] [PubMed]
- Nguyen Dinh, C.A.; Montezano, A.C.; Burger, D.; Touyz, R.M. Angiotensin II, NADPH oxidase, and redox signaling in the vasculature. Antioxid. Redox. Signal. 2013, 19, 1110–1120. [Google Scholar] [CrossRef] [Green Version]
- Sifi, A.; di-Bessalem, S.; Laraba-Djebari, F. Role of angiotensin II and angiotensin type-1 receptor in scorpion venom-induced cardiac and aortic tissue inflammation. Exp. Mol. Pathol. 2017, 102, 32–40. [Google Scholar] [CrossRef]
- Montezano, A.C.; Callera, G.E.; Yogi, A.; He, Y.; Tostes, R.C.; He, G.; Schiffrin, E.L.; Touyz, R.M. Aldosterone and angiotensin II synergistically stimulate migration in vascular smooth muscle cells through c-Src-regulated redox-sensitive RhoA pathways. Arterioscler. Thromb. Vasc. Biol. 2008, 28, 1511–1518. [Google Scholar] [CrossRef] [Green Version]
- Wang, Y.; Ji, L.; Jiang, R.; Zheng, L.; Liu, D. Oxidized high-density lipoprotein induces the proliferation and migration of vascular smooth muscle cells by promoting the production of ROS. J. Atheroscler. Thromb. 2014, 21, 204–216. [Google Scholar] [CrossRef] [Green Version]
- Kim, S.Y.; Lee, J.G.; Cho, W.S.; Cho, K.H.; Sakong, J.; Kim, J.R.; Chin, B.R.; Baek, S.H. Role of NADPH oxidase-2 in lipopolysaccharide-induced matrix metalloproteinase expression and cell migration. Immunol. Cell Biol. 2010, 88, 197–204. [Google Scholar] [CrossRef]
- Garcia-Redondo, A.B.; Aguado, A.; Briones, A.M.; Salaices, M. NADPH oxidases and vascular remodeling in cardiovascular diseases. Pharmacol. Res. 2016, 114, 110–120. [Google Scholar] [CrossRef]
- Siti, H.N.; Kamisah, Y.; Kamsiah, J. The role of oxidative stress, antioxidants and vascular inflammation in cardiovascular disease (a review). Vascul. Pharmacol. 2015, 71, 40–56. [Google Scholar] [CrossRef]
- Chen, X.H.; Ruan, C.C.; Ge, Q.; Ma, Y.; Xu, J.Z.; Zhang, Z.B.; Lin, J.R.; Chen, D.R.; Zhu, D.L.; Gao, P.J. Deficiency of complement C3a and C5a receptors prevents angiotensin II-induced hypertension via regulatory T cells. Circ. Res. 2018, 122, 970–983. [Google Scholar] [CrossRef] [PubMed]
- Kumar, N.; Liao, T.D.; Romero, C.A.; Maheshwari, M.; Peterson, E.L.; Carretero, O.A. Thymosin beta4 deficiency exacerbates renal and cardiac injury in angiotensin-II-induced hypertension. Hypertension 2018, 71, 1133–1142. [Google Scholar] [CrossRef] [PubMed]
- Song, C.Y.; Khan, N.S.; Liao, F.F.; Wang, B.; Shin, J.S.; Bonventre, J.V.; Malik, K.U. Brain cytosolic phospholipase A2alpha mediates angiotensin II-induced hypertension and reactive oxygen species production in male mice. Am. J. Hypertens. 2018, 31, 622–629. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Haudenschild, C.C.; Grunwald, J.; Chobanian, A.V. Effects of hypertension on migration and proliferation of smooth muscle in culture. Hypertension 1985, 7, I101–I104. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grunwald, J.; Chobanian, A.V.; Haudenschild, C.C. Smooth muscle cell migration and proliferation: Atherogenic mechanisms in hypertension. Atherosclerosis 1987, 67, 215–221. [Google Scholar] [CrossRef]
- Belo, V.A.; Guimaraes, D.A.; Castro, M.M. Matrix metalloproteinase 2 as a potential mediator of vascular smooth muscle cell migration and chronic vascular remodeling in hypertension. J. Vasc. Res. 2015, 52, 221–231. [Google Scholar] [CrossRef]
- Rudemiller, N.P.; Crowley, S.D. Interactions between the immune and the renin-angiotensin systems in hypertension. Hypertension 2016, 68, 289–296. [Google Scholar] [CrossRef]
- Mirabito Colafella, K.M.; Danser, A.H.J. RFecent advances in angiotensin research. Hypertension 2017, 69, 994–999. [Google Scholar] [CrossRef]
- Eguchi, S.; Kawai, T.; Scalia, R.; Rizzo, V. Understanding angiotensin II type 1 receptor signaling in vascular pathophysiology. Hypertension 2018, 71, 804–810. [Google Scholar] [CrossRef]
- Sinha, N.; Dabla, P.K. Oxidative stress and antioxidants in hypertension-a current review. Curr. Hypertens. Rev. 2015, 11, 132–142. [Google Scholar] [CrossRef]
- Meijles, D.N.; Pagano, P.J. Nox and Inflammation in the Vascular Adventitia. Hypertension 2016, 67, 14–19. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhuo, J.L. SH2B3 (LNK) as a novel link of immune signaling, inflammation, and hypertension in Dahl salt-sensitive hypertensive rats. Hypertension 2015, 65, 989–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Marchio, P.; Guerra-Ojeda, S.; Vila, J.M.; Aldasoro, M.; Victor, V.M.; Mauricio, M.D. Targeting Early Atherosclerosis: A Focus on Oxidative Stress and Inflammation. Oxid. Med. Cell Longev. 2019, 2019, 8563845. [Google Scholar] [CrossRef] [PubMed]
- Steven, S.; Frenis, K.; Oelze, M.; Kalinovic, S.; Kuntic, M.; Bayo Jimenez, M.T.; Vujacic-Mirski, K.; Helmstadter, J.; Kroller-Schon, S.; Munzel, T.; et al. Vascular Inflammation and Oxidative Stress: Major Triggers for Cardiovascular Disease. Oxid. Med. Cell Longev. 2019, 2019, 7092151. [Google Scholar] [CrossRef] [Green Version]
- Sturza, A.; Popoiu, C.M.; Ionica, M.; Duicu, O.M.; Olariu, S.; Muntean, D.M.; Boia, E.S. Monoamine Oxidase-Related Vascular Oxidative Stress in Diseases Associated with Inflammatory Burden. Oxid. Med. Cell Longev. 2019, 2019, 8954201. [Google Scholar] [CrossRef]
- Pacurari, M.; Kafoury, R.; Tchounwou, P.B.; Ndebele, K. The Renin-Angiotensin-aldosterone system in vascular inflammation and remodeling. Int. J. Inflam. 2014, 2014, 689360. [Google Scholar] [CrossRef]
- Chen, D.; Zang, Y.H.; Qiu, Y.; Zhang, F.; Chen, A.D.; Wang, J.J.; Chen, Q.; Li, Y.H.; Kang, Y.M.; Zhu, G.Q. BCL6 Attenuates Proliferation and Oxidative Stress of Vascular Smooth Muscle Cells in Hypertension. Oxid. Med. Cell Longev. 2019, 2019, 5018410. [Google Scholar] [CrossRef] [Green Version]
- Peng, K.; Lu, X.; Wang, F.; Nau, A.; Chen, R.; Zhou, S.F.; Yang, T. Collecting duct (pro)renin receptor targets ENaC to mediate angiotensin II-induced hypertension. Am. J. Physiol. Renal Physiol. 2017, 312, F245–F253. [Google Scholar] [CrossRef]
- Xue, B.; Beltz, T.G.; Yu, Y.; Guo, F.; Gomez-Sanchez, C.E.; Hay, M.; Johnson, A.K. Central interactions of aldosterone and angiotensin II in aldosterone- and angiotensin II-induced hypertension. Am. J. Physiol. Heart Circ. Physiol. 2011, 300, H555–H564. [Google Scholar] [CrossRef] [Green Version]
- Hicks, B.M.; Filion, K.B.; Yin, H.; Sakr, L.; Udell, J.A.; Azoulay, L. Angiotensin converting enzyme inhibitors and risk of lung cancer: Population based cohort study. BMJ 2018, 363, k4209. [Google Scholar] [CrossRef] [Green Version]







© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, N.; Ye, C.; Zheng, F.; Wan, G.-W.; Wu, L.-L.; Chen, Q.; Li, Y.-H.; Kang, Y.-M.; Zhu, G.-Q. MiR155-5p Inhibits Cell Migration and Oxidative Stress in Vascular Smooth Muscle Cells of Spontaneously Hypertensive Rats. Antioxidants 2020, 9, 204. https://doi.org/10.3390/antiox9030204
Wu N, Ye C, Zheng F, Wan G-W, Wu L-L, Chen Q, Li Y-H, Kang Y-M, Zhu G-Q. MiR155-5p Inhibits Cell Migration and Oxidative Stress in Vascular Smooth Muscle Cells of Spontaneously Hypertensive Rats. Antioxidants. 2020; 9(3):204. https://doi.org/10.3390/antiox9030204
Chicago/Turabian StyleWu, Nan, Chao Ye, Fen Zheng, Guo-Wei Wan, Lu-Lu Wu, Qi Chen, Yue-Hua Li, Yu-Ming Kang, and Guo-Qing Zhu. 2020. "MiR155-5p Inhibits Cell Migration and Oxidative Stress in Vascular Smooth Muscle Cells of Spontaneously Hypertensive Rats" Antioxidants 9, no. 3: 204. https://doi.org/10.3390/antiox9030204