Weaning Mice and Adult Mice Exhibit Differential Carbon Tetrachloride-Induced Acute Hepatotoxicity

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animal and Treatments
2.2. CCl4-Induced Hepatotoxicity
2.3. Immunoblotting Analysis
2.4. Hepatic Lipid Peroxidation Induced by CCl4
2.5. Measurement of Hepatic Protein Oxidation
2.6. Determination of Levels of Hepatic cysteine, GSH, and Taurine
2.7. Statistical Analysis
3. Results
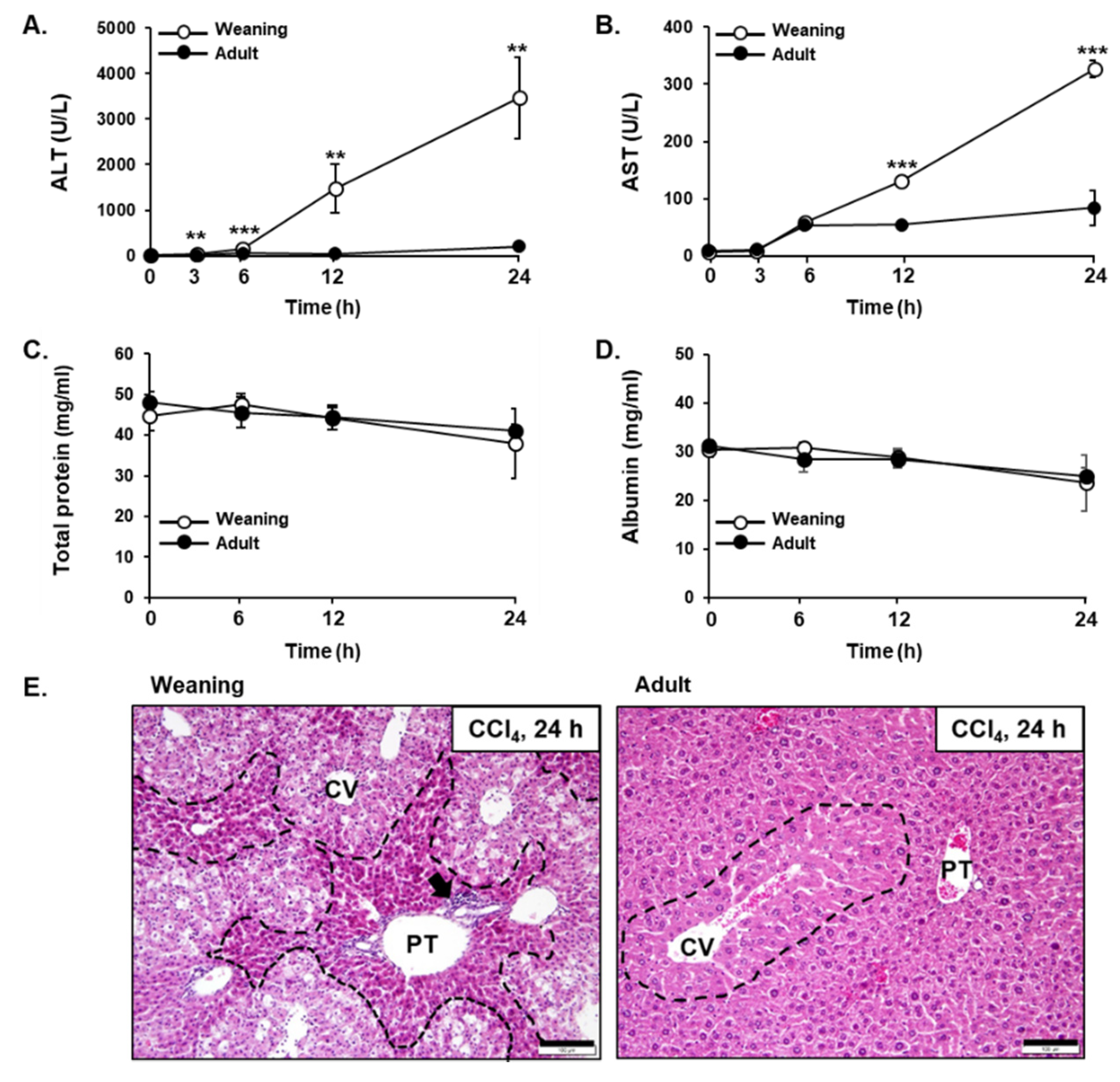
3.1. CCl4-Induced Liver Injuries in Weaning Mice and Adult Mice
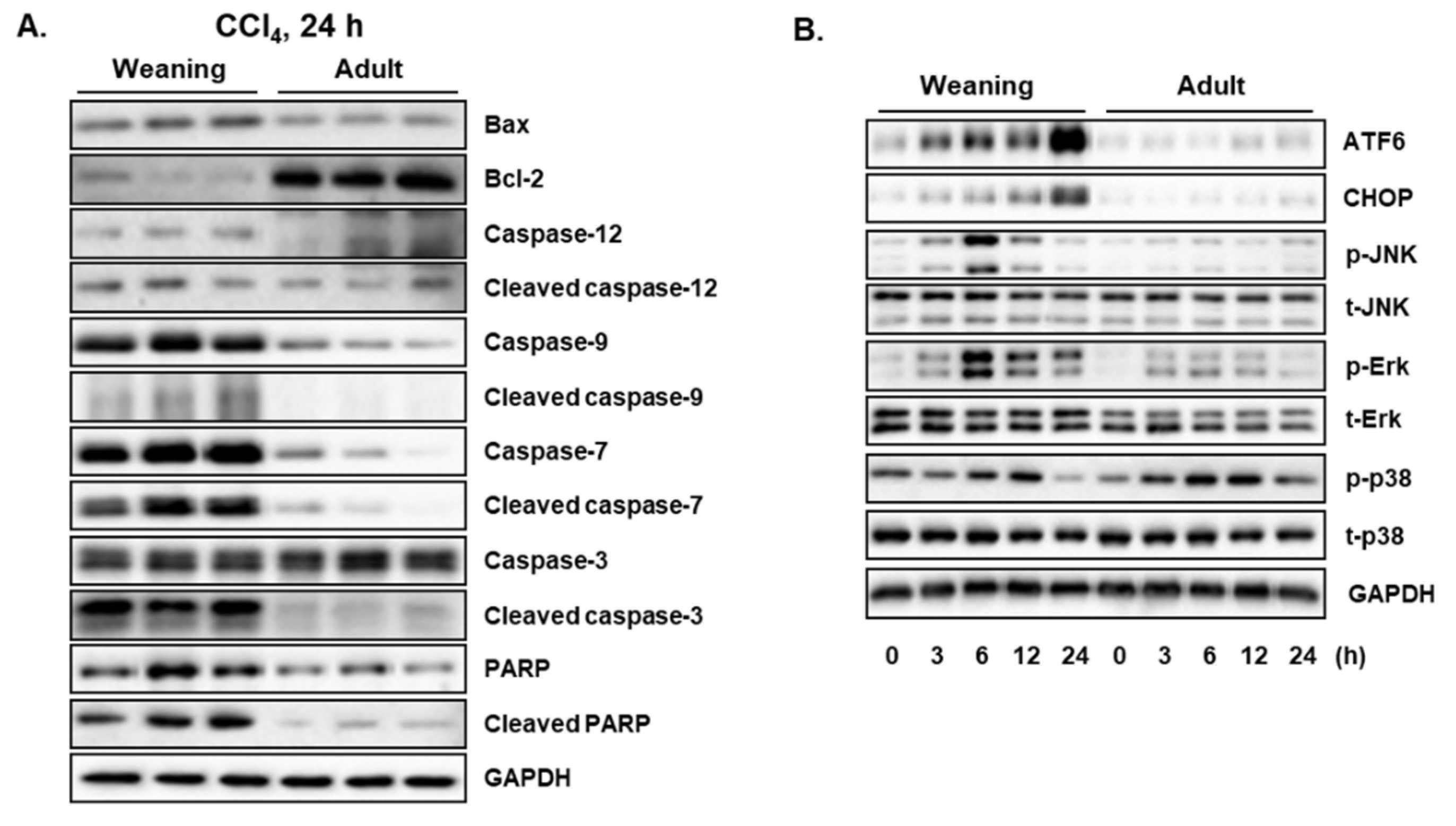
3.2. CCl4-Induced Hepatic Apoptosis and Endoplasmic Reticulum (ER) Stress in Weaning Mice and Adult Mice
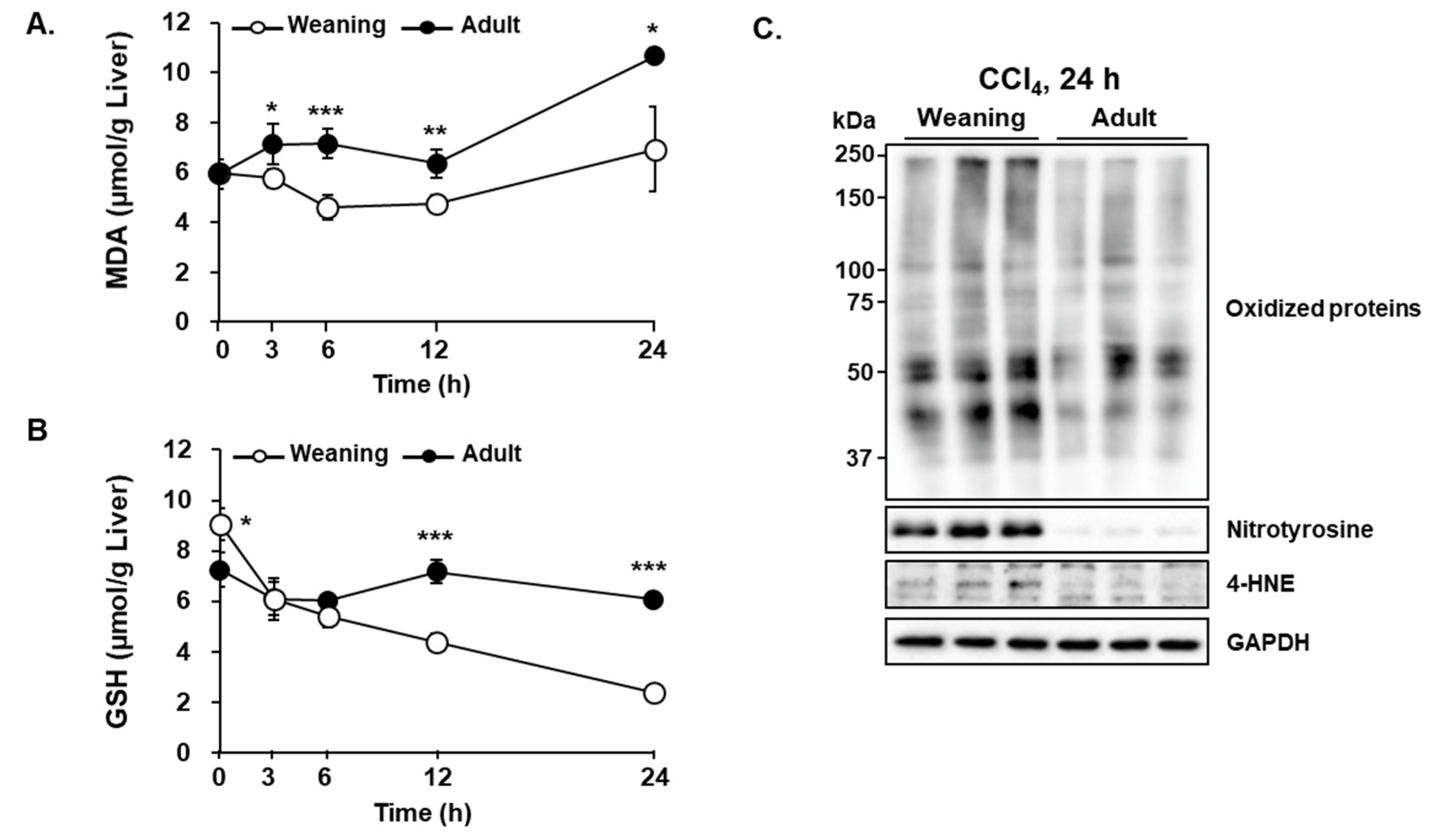
3.3. CCl4-Induced Oxidative Stress in the Liver of Weaning Mice and Adult Mice
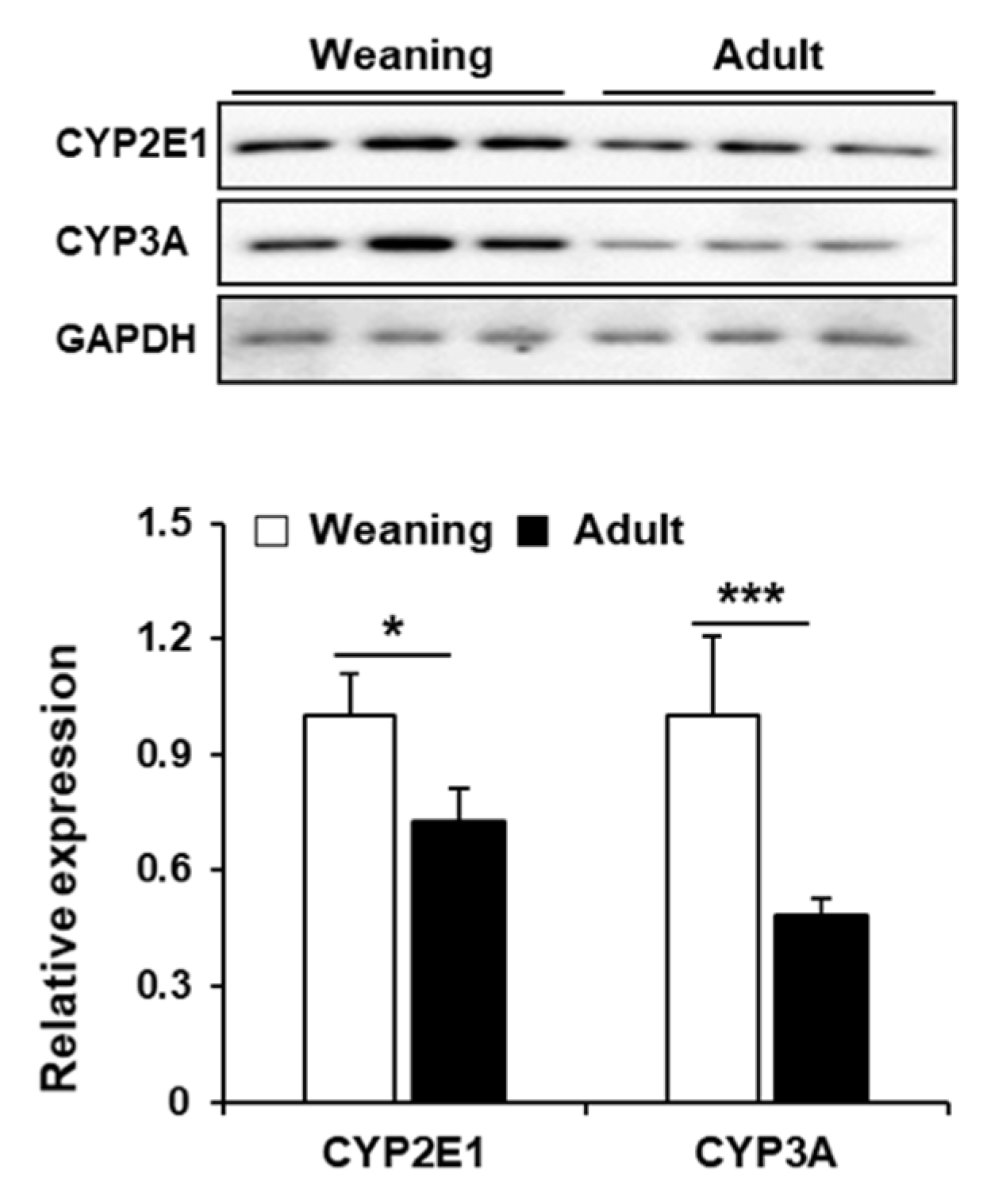
3.4. Hepatic CYP Levels in Weaning Mice and Adult Mice
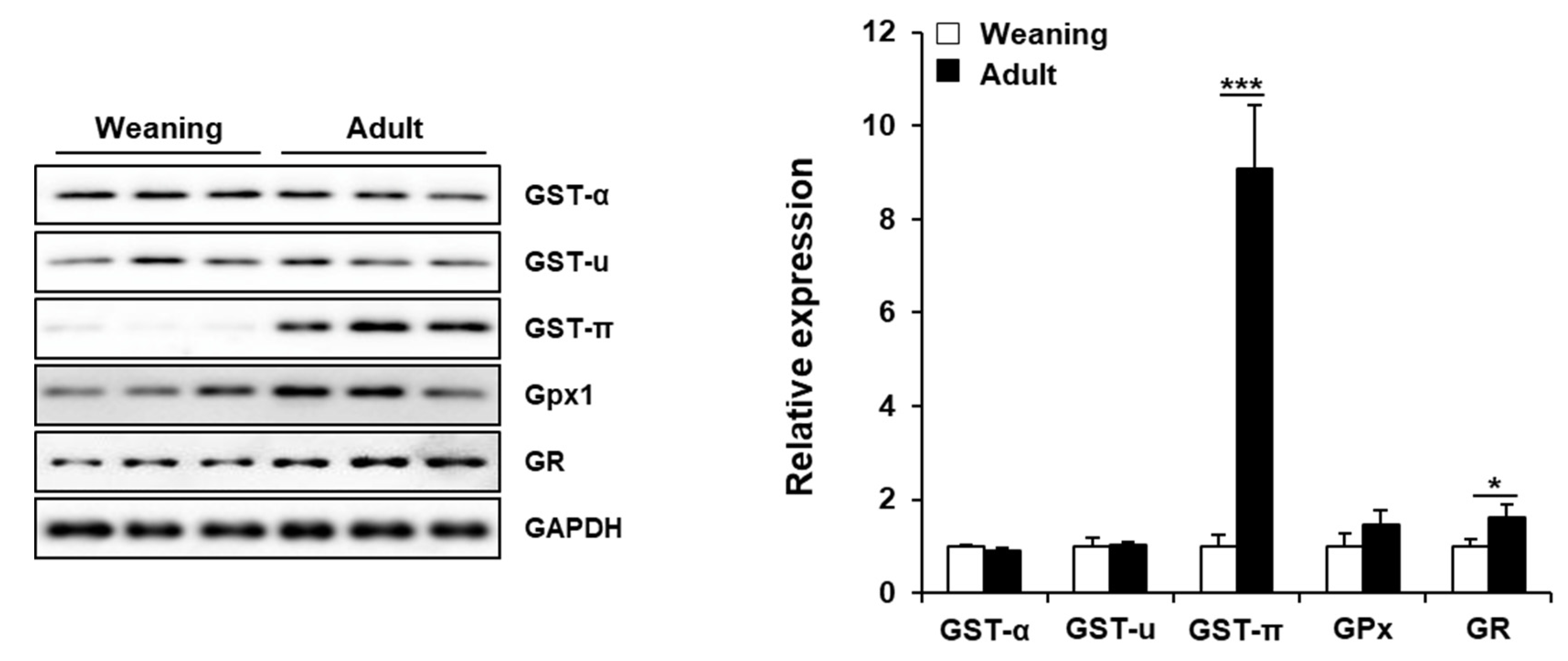
3.5. Hepatic Enzymes Involved in GSH Metabolism in Weaning Mice and Adult Mice
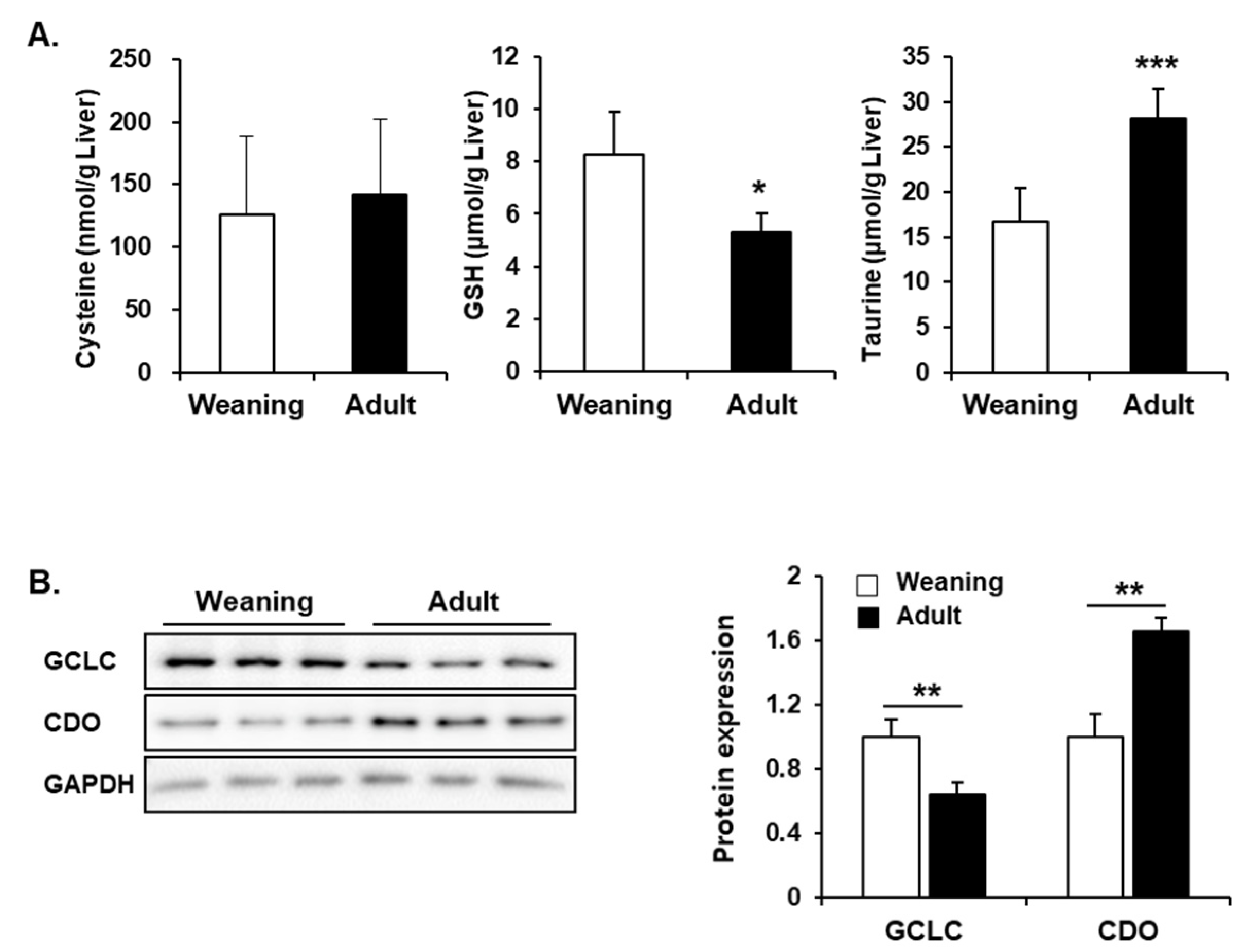
3.6. Hepatic Synthesis of GSH and Taurine from Cysteine in Weaning Mice and Adult Mice
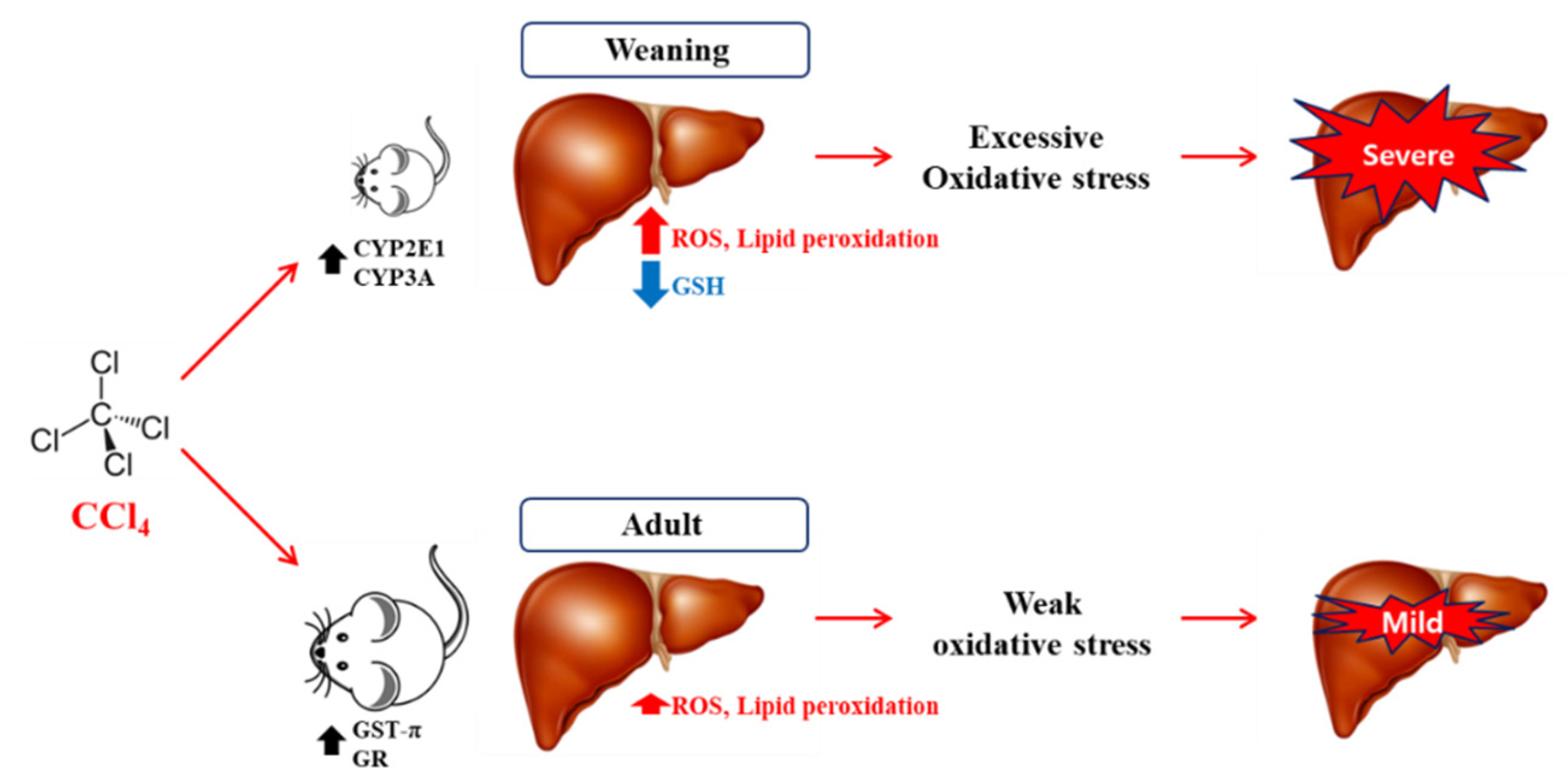
4. Discussion
5. Conclusions
Author Contributions
Funding
Conflicts of Interest
References
- Croom, E. Metabolism of xenobiotics of human environments. Prog. Mol. Biol. Transl. Sci. 2012, 112, 31–88. [Google Scholar] [CrossRef] [PubMed]
- Gu, X.; Manautou, J.E. Molecular mechanisms underlying chemical liver injury. Expert Rev. Mol. Med. 2012, 14, e4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Corsini, A.; Bortolini, M. Drug-induced liver injury: The role of drug metabolism and transport. J. Clin. Pharmacol. 2013, 53, 463–474. [Google Scholar] [CrossRef] [PubMed]
- Shi, Q.; Yang, X.; Greenhaw, J.J.; Salminen, A.T.; Russotti, G.M.; Salminen, W.F. Drug-Induced Liver Injury in Children: Clinical Observations, Animal Models, and Regulatory Status. Int. J. Toxicol. 2017, 36, 365–379. [Google Scholar] [CrossRef] [PubMed]
- Chalasani, N.; Bjornsson, E. Risk factors for idiosyncratic drug-induced liver injury. Gastroenterology 2010, 138, 2246–2259. [Google Scholar] [CrossRef] [Green Version]
- Sotaniemi, E.A.; Arranto, A.J.; Pelkonen, O.; Pasanen, M. Age and cytochrome P450-linked drug metabolism in humans: An analysis of 226 subjects with equal histopathologic conditions. Clin. Pharmacol. Ther. 1997, 61, 331–339. [Google Scholar] [CrossRef]
- Parkinson, A.; Mudra, D.R.; Johnson, C.; Dwyer, A.; Carroll, K.M. The effects of gender, age, ethnicity, and liver cirrhosis on cytochrome P450 enzyme activity in human liver microsomes and inducibility in cultured human hepatocytes. Toxicol. Appl. Pharmacol. 2004, 199, 193–209. [Google Scholar] [CrossRef]
- Rikans, L.E.; Moore, D.R. Acetaminophen hepatotoxicity in aging rats. Drug Chem. Toxicol. 1988, 11, 237–247. [Google Scholar] [CrossRef]
- Palomero, J. Effects of aging on the susceptibility to the toxic effects of cyclosporin A in rats. Changes in liver glutathione and antioxidant enzymes. Free Radic. Biol. Med. 2001, 30, 836–845. [Google Scholar] [CrossRef]
- Faa, G.; Ekstrom, J.; Castagnola, M.; Gibo, Y.; Ottonello, G.; Fanos, V. A developmental approach to drug-induced liver injury in newborns and children. Curr. Med. Chem. 2012, 19, 4581–4594. [Google Scholar] [CrossRef] [Green Version]
- Bryant, A.E., III; Dreifuss, F.E. Valproic acid hepatic fatalities. III. U.S. experience since 1986. Neurology 1996, 46, 465–469. [Google Scholar] [CrossRef] [PubMed]
- Rivkees, S.A.; Szarfman, A. Dissimilar hepatotoxicity profiles of propylthiouracil and methimazole in children. J. Clin. Endocrinol. Metab. 2010, 95, 3260–3267. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mach, J.; Huizer-Pajkos, A.; Cogger, V.C.; McKenzie, C.; Le Couteur, D.G.; Jones, B.E.; de Cabo, R.; Hilmer, S.N. The effect of aging on acetaminophen pharmacokinetics, toxicity and Nrf2 in Fischer 344 rats. J. Gerontol. Biol. Sci. Med. Sci. 2014, 69, 387–397. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mach, J.; Huizer-Pajkos, A.; Mitchell, S.J.; McKenzie, C.; Phillips, L.; Kane, A.; Jones, B.; de Cabo, R.; Cogger, V.; Le Couteur, D.G.; et al. The effect of ageing on isoniazid pharmacokinetics and hepatotoxicity in Fischer 344 rats. Fundam. Clin. Pharmacol. 2016, 30, 23–34. [Google Scholar] [CrossRef] [PubMed]
- Brautbar, N.; Williams, J., II. Industrial solvents and liver toxicity: Risk assessment, risk factors and mechanisms. Int. J. Hyg. Environ. Health 2002, 205, 479–491. [Google Scholar] [CrossRef] [PubMed]
- Weber, L.W.; Boll, M.; Stampfl, A. Hepatotoxicity and mechanism of action of haloalkanes: Carbon tetrachloride as a toxicological model. Crit. Rev. Toxicol. 2003, 33, 105–136. [Google Scholar] [CrossRef]
- Jahn, F.; Reuter, A.; Karge, E.; Danz, M.; Klinger, W. Age dependent different influence of carbon tetrachloride on biotransformation of xenobiotics, glutathione content, lipid peroxidation and histopathology of rat liver. Exp. Toxicol. Pathol. 1993, 45, 101–107. [Google Scholar] [CrossRef]
- Rikans, L.E.; Hornbrook, K.R.; Cai, Y. Carbon tetrachloride hepatotoxicity as a function of age in female Fischer 344 rats. Mech. Ageing Dev. 1994, 76, 89–99. [Google Scholar] [CrossRef]
- Dalu, A.; Warbritton, A.; Bucci, T.J.; Mehendale, H.M. Age-related susceptibility to chlordecone-potentiated carbon tetrachloride hepatotoxicity and lethality is due to hepatic quiescence. Pediatr. Res. 1995, 38, 140–148. [Google Scholar] [CrossRef] [Green Version]
- Collins, B.H.; Holzknecht, Z.E.; Lynn, K.A.; Sempowski, G.D.; Smith, C.C.; Liu, S.; Parker, W.; Rockey, D.C. Association of age-dependent liver injury and fibrosis with immune cell populations. Liver Int. 2013, 33, 1175–1186. [Google Scholar] [CrossRef] [Green Version]
- Hong, I.H.; Lewis, K.; Iakova, P.; Jin, J.; Sullivan, E.; Jawanmardi, N.; Timchenko, L.; Timchenko, N. Age-associated change of C/EBP family proteins causes severe liver injury and acceleration of liver proliferation after CCl4 treatments. J. Biol. Chem. 2014, 289, 1106–1118. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Reitman, S.; Frankel, S. A colorimetric method for the determination of serum glutamic oxalacetic and glutamic pyruvic transaminases. Am. J. Clin. Pathol. 1957, 28, 56–63. [Google Scholar] [CrossRef] [PubMed]
- Volpi, N.; Tarugi, P. Improvement in the high-performance liquid chromatography malondialdehyde level determination in normal human plasma. J. Chromatogr. B Biomed. Sci. Appl. 1998, 713, 433–437. [Google Scholar] [CrossRef]
- Lee, Y.H.; Kim, S.H.; Lee, S.; Kim, K.M.; Jung, J.C.; Son, T.G.; Ki, S.H.; Seo, W.D.; Kwak, J.H.; Hong, J.T.; et al. Antioxidant Effect of Barley Sprout Extract via Enhancement of Nuclear Factor-Erythroid 2 Related Factor 2 Activity and Glutathione Synthesis. Nutrients 2017, 9, 1252. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kim, S.H.; Kwon, D.Y.; Kwak, J.H.; Lee, S.; Lee, Y.H.; Yun, J.; Son, T.G.; Jung, Y.S. Tunicamycin-Induced ER Stress is Accompanied with Oxidative Stress via Abrogation of Sulfur Amino Acids Metabolism in the Liver. Int. J. Mol. Sci. 2018, 19, 4114. [Google Scholar] [CrossRef] [Green Version]
- Liu, H.; Wang, Z.; Nowicki, M.J. Caspase-12 mediates carbon tetrachloride-induced hepatocyte apoptosis in mice. World J. Gastroenterol. 2014, 20, 18189–18198. [Google Scholar] [CrossRef]
- Vannuvel, K.; Renard, P.; Raes, M.; Arnould, T. Functional and morphological impact of ER stress on mitochondria. J. Cell Physiol. 2013, 228, 1802–1818. [Google Scholar] [CrossRef]
- Iurlaro, R.; Munoz-Pinedo, C. Cell death induced by endoplasmic reticulum stress. FEBS J. 2016, 283, 2640–2652. [Google Scholar] [CrossRef] [Green Version]
- Suh, Y. Cell signaling in aging and apoptosis. Mech. Ageing Dev. 2002, 123, 881–890. [Google Scholar] [CrossRef]
- Kim, E.K.; Choi, E.J. Pathological roles of MAPK signaling pathways in human diseases. Biochim. Biophys Acta 2010, 1802, 396–405. [Google Scholar] [CrossRef] [Green Version]
- Hayes, J.D.; Flanagan, J.U.; Jowsey, I.R. Glutathione transferases. Annu. Rev. Pharmacol. Toxicol. 2005, 45, 51–88. [Google Scholar] [CrossRef] [PubMed]
- Forman, H.J.; Zhang, H.; Rinna, A. Glutathione: Overview of its protective roles, measurement, and biosynthesis. Mol. Aspects Med. 2009, 30, 1–12. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Arana, A.; Morton, N.S.; Hansen, T.G. Treatment with paracetamol in infants. Acta Anaesthesiol. Scand. 2001, 45, 20–29. [Google Scholar] [CrossRef] [PubMed]
- O’Donnell, J.T.; Marks, D.H.; Danese, P.; O’Donnell, J.J., III. Drug-induced liver disease: Primer for the primary care physician. Dis. Mon. 2014, 60, 55–104. [Google Scholar] [CrossRef]
- Erden-Inal, M.; Sunal, E.; Kanbak, G. Age-related changes in the glutathione redox system. Cell Biochem. Funct. 2002, 20, 61–66. [Google Scholar] [CrossRef]
- Wong, F.W.; Chan, W.Y.; Lee, S.S. Resistance to carbon tetrachloride-induced hepatotoxicity in mice which lack CYP2E1 expression. Toxicol. Appl. Pharmacol. 1998, 153, 109–118. [Google Scholar] [CrossRef]
- Zangar, R.C.; Benson, J.M.; Burnett, V.L.; Springer, D.L. Cytochrome P450 2E1 is the primary enzyme responsible for low-dose carbon tetrachloride metabolism in human liver microsomes. Chem. Biol. Interact. 2000, 125, 233–243. [Google Scholar] [CrossRef]
- Veith, A.; Moorthy, B. Role of Cytochrome P450s in the Generation and Metabolism of Reactive Oxygen Species. Curr. Opin. Toxicol. 2018, 7, 44–51. [Google Scholar] [CrossRef]
- Oinonen, T.; Lindros, K.O. Zonation of hepatic cytochrome P-450 expression and regulation. Biochem. J. 1998, 329, 17–35. [Google Scholar] [CrossRef] [Green Version]
- Kietzmann, T. Metabolic zonation of the liver: The oxygen gradient revisited. Redox Biol. 2017, 11, 622–630. [Google Scholar] [CrossRef]
- Vyskocilova, E.; Szotakova, B.; Skalova, L.; Bartikova, H.; Hlavacova, J.; Bousova, I. Age-related changes in hepatic activity and expression of detoxification enzymes in male rats. Biomed. Res. Int. 2013, 2013, 408573. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lu, Y.; Zhang, C.; Chen, Y.H.; Wang, H.; Zhang, Z.H.; Chen, X.; Xu, D.X. Immature mice are more susceptible than adult mice to acetaminophen-induced acute liver injury. Sci. Rep. 2017, 7, 42736. [Google Scholar] [CrossRef] [Green Version]
- Vieira, I.; Sonnier, M.; Cresteil, T. Developmental expression of CYP2E1 in the human liver. Hypermethylation control of gene expression during the neonatal period. Eur. J. Biochem. 1996, 238, 476–483. [Google Scholar] [CrossRef] [PubMed]
- Johnson, T.N. The development of drug metabolising enzymes and their influence on the susceptibility to adverse drug reactions in children. Toxicology 2003, 192, 37–48. [Google Scholar] [CrossRef]
- Iracheta-Vellve, A.; Petrasek, J.; Gyongyosi, B.; Satishchandran, A.; Lowe, P.; Kodys, K.; Catalano, D.; Calenda, C.D.; Kurt-Jones, E.A.; Fitzgerald, K.A.; et al. Endoplasmic Reticulum Stress-induced Hepatocellular Death Pathways Mediate Liver Injury and Fibrosis via Stimulator of Interferon Genes. J. Biol. Chem. 2016, 291, 26794–26805. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jang, I.; Chae, K.; Cho, J. Effects of age and strain on small intestinal and hepatic antioxidant defense enzymes in Wistar and Fisher 344 rats. Mech. Ageing Dev. 2001, 122, 561–570. [Google Scholar] [CrossRef]
- Xu, S.; Hou, D.; Liu, J.; Ji, L. Age-associated changes in GSH S-transferase gene/proteins in livers of rats. Redox Rep. 2018, 23, 213–218. [Google Scholar] [CrossRef]
- Adler, V.; Yin, Z.; Fuchs, S.Y.; Benezra, M.; Rosario, L.; Tew, K.D.; Pincus, M.R.; Sardana, M.; Henderson, C.J.; Wolf, C.R.; et al. Regulation of JNK signaling by GSTp. EMBO J. 1999, 18, 1321–1334. [Google Scholar] [CrossRef]
- Tew, K.D.; Townsend, D.M. Regulatory functions of glutathione S-transferase P1-1 unrelated to detoxification. Drug Metab. Rev. 2011, 43, 179–193. [Google Scholar] [CrossRef] [Green Version]
- Bernardini, S.; Bernassola, F.; Cortese, C.; Ballerini, S.; Melino, G.; Motti, C.; Bellincampi, L.; Iori, R.; Federici, G. Modulation of GST P1-1 activity by polymerization during apoptosis. J. Cell Biochem. 2000, 77, 645–653. [Google Scholar] [CrossRef]
- Ferguson, G.; Bridge, W. Glutamate cysteine ligase and the age-related decline in cellular glutathione: The therapeutic potential of gamma-glutamylcysteine. Arch. Biochem. Biophys. 2016, 593, 12–23. [Google Scholar] [CrossRef] [PubMed]
- Liu, R.; Choi, J. Age-associated decline in gamma-glutamylcysteine synthetase gene expression in rats. Free Radic Biol. Med. 2000, 28, 566–574. [Google Scholar] [CrossRef]
- Toroser, D.; Sohal, R.S. Age-associated perturbations in glutathione synthesis in mouse liver. Biochem. J. 2007, 405, 583–589. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.; Sheng, Y.; Ji, L. The age-related change of glutathione antioxidant system in mice liver. Toxicol. Mech. Methods 2013, 23, 396–401. [Google Scholar] [CrossRef] [PubMed]
- Schuller-Levis, G.B.; Park, E. Taurine: New implications for an old amino acid. FEMS Microbiol. Lett. 2003, 226, 195–202. [Google Scholar] [CrossRef]
- Eppler, B.; Dawson, R., Jr. Cysteine sulfinate decarboxylase and cysteine dioxygenase activities do not correlate with strain-specific changes in hepatic and cerebellar taurine content in aged rats. Mech. Ageing Dev. 1999, 110, 57–72. [Google Scholar] [CrossRef]
- Eppler, B.; Dawson, R., Jr. Dietary taurine manipulations in aged male Fischer 344 rat tissue: Taurine concentration, taurine biosynthesis, and oxidative markers. Biochem. Pharmacol. 2001, 62, 29–39. [Google Scholar] [CrossRef]








© 2020 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Jeong, T.B.; Kwon, D.; Son, S.W.; Kim, S.H.; Lee, Y.-H.; Seo, M.-S.; Kim, K.S.; Jung, Y.-S. Weaning Mice and Adult Mice Exhibit Differential Carbon Tetrachloride-Induced Acute Hepatotoxicity. Antioxidants 2020, 9, 201. https://doi.org/10.3390/antiox9030201
Jeong TB, Kwon D, Son SW, Kim SH, Lee Y-H, Seo M-S, Kim KS, Jung Y-S. Weaning Mice and Adult Mice Exhibit Differential Carbon Tetrachloride-Induced Acute Hepatotoxicity. Antioxidants. 2020; 9(3):201. https://doi.org/10.3390/antiox9030201
Chicago/Turabian StyleJeong, Tae Bin, Doyoung Kwon, Seung Won Son, Sou Hyun Kim, Yun-Hee Lee, Min-Soo Seo, Kil Soo Kim, and Young-Suk Jung. 2020. "Weaning Mice and Adult Mice Exhibit Differential Carbon Tetrachloride-Induced Acute Hepatotoxicity" Antioxidants 9, no. 3: 201. https://doi.org/10.3390/antiox9030201