
Comparison of Milk Kefirs Obtained from Cow’s, Ewe’s and Goat’s Milk: Antioxidant Role of Microbial-Derived Exopolysaccharides

 ,
,  , ,
, ,  ,
,  ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Milk Kefir Fermentation
2.2. Microbiological Characterization
2.3. Physicochemical and Rheological Characterization
2.4. Sugars, Ethanol, and Organic Acids Quantification
2.5. Total and Individual Free Amino Acids (FAAs)
2.6. Purification of EPS
2.7. DPPH· Radical Scavenging Activity
2.8. Ferric Reducing Antioxidant Power (FRAP)
2.9. ABTS·+ Radical Scavenging Activity
2.10. Fourier Transform Infrared Spectroscopy (FT-IR)
2.11. Monosaccharide Composition of EPS
2.12. Scanning Electron Microscopy (SEM)
2.13. Statistical Analysis
3. Results and Discussion
3.1. Microbiological, Physicochemical, and Rheological Characterization of Milk Kefirs Obtained from Cow, Ewe, and Goat Milk
3.2. Microbial Metabolites
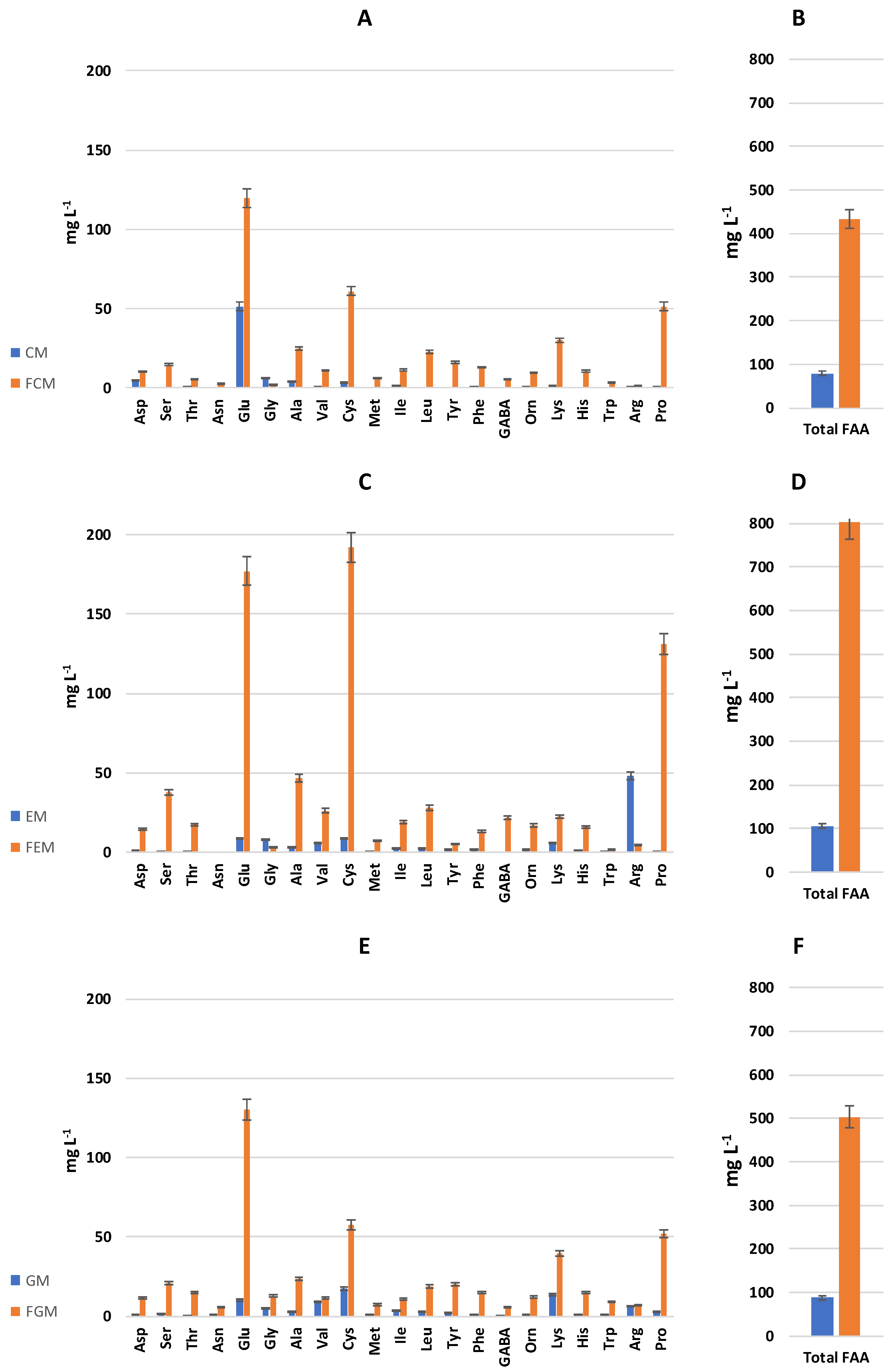
3.3. FAA Profiles
3.4. EPS Purification and Quantification
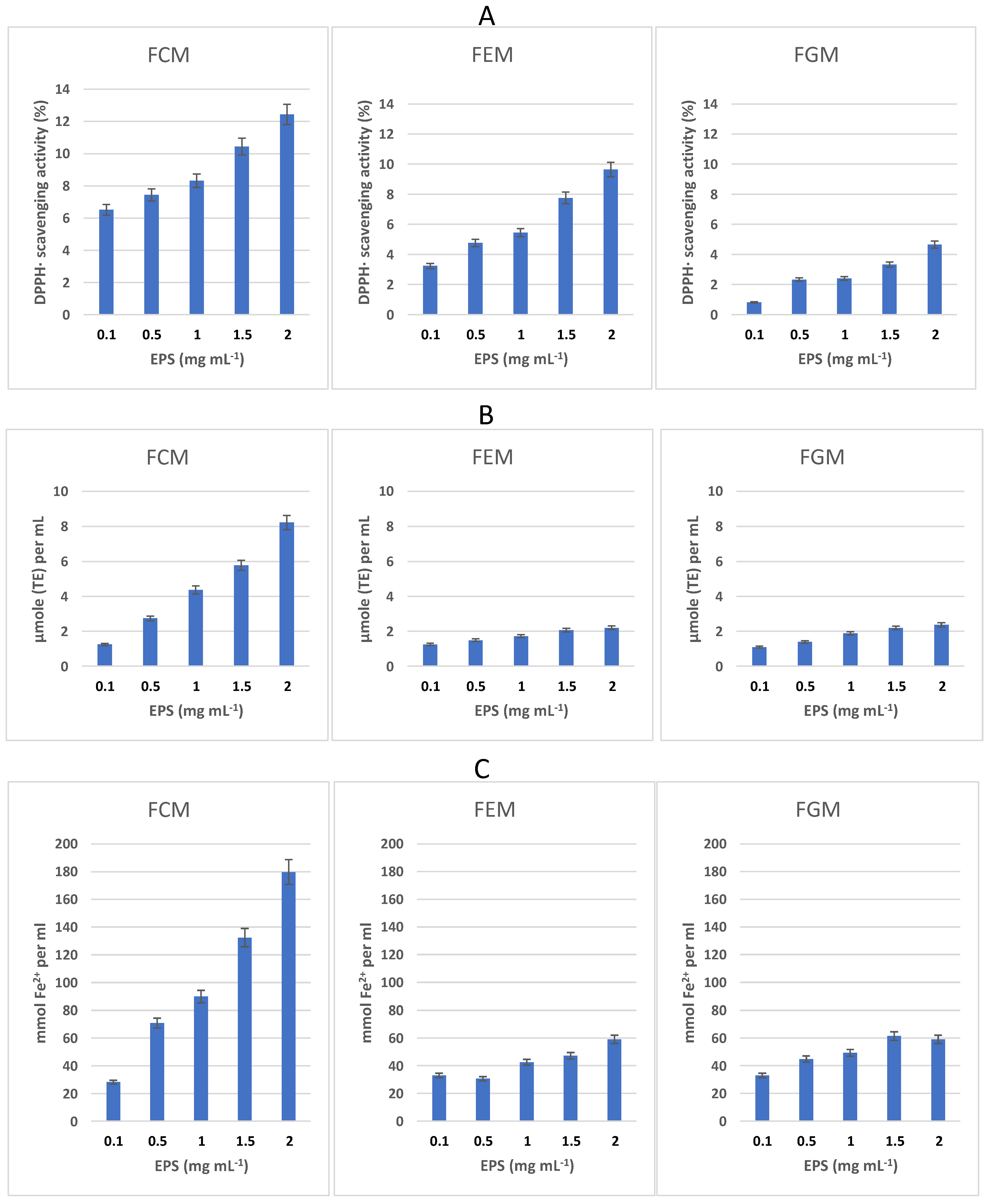
3.5. Antioxidant Properties of EPS
3.6. Monosaccharide Composition of EPS
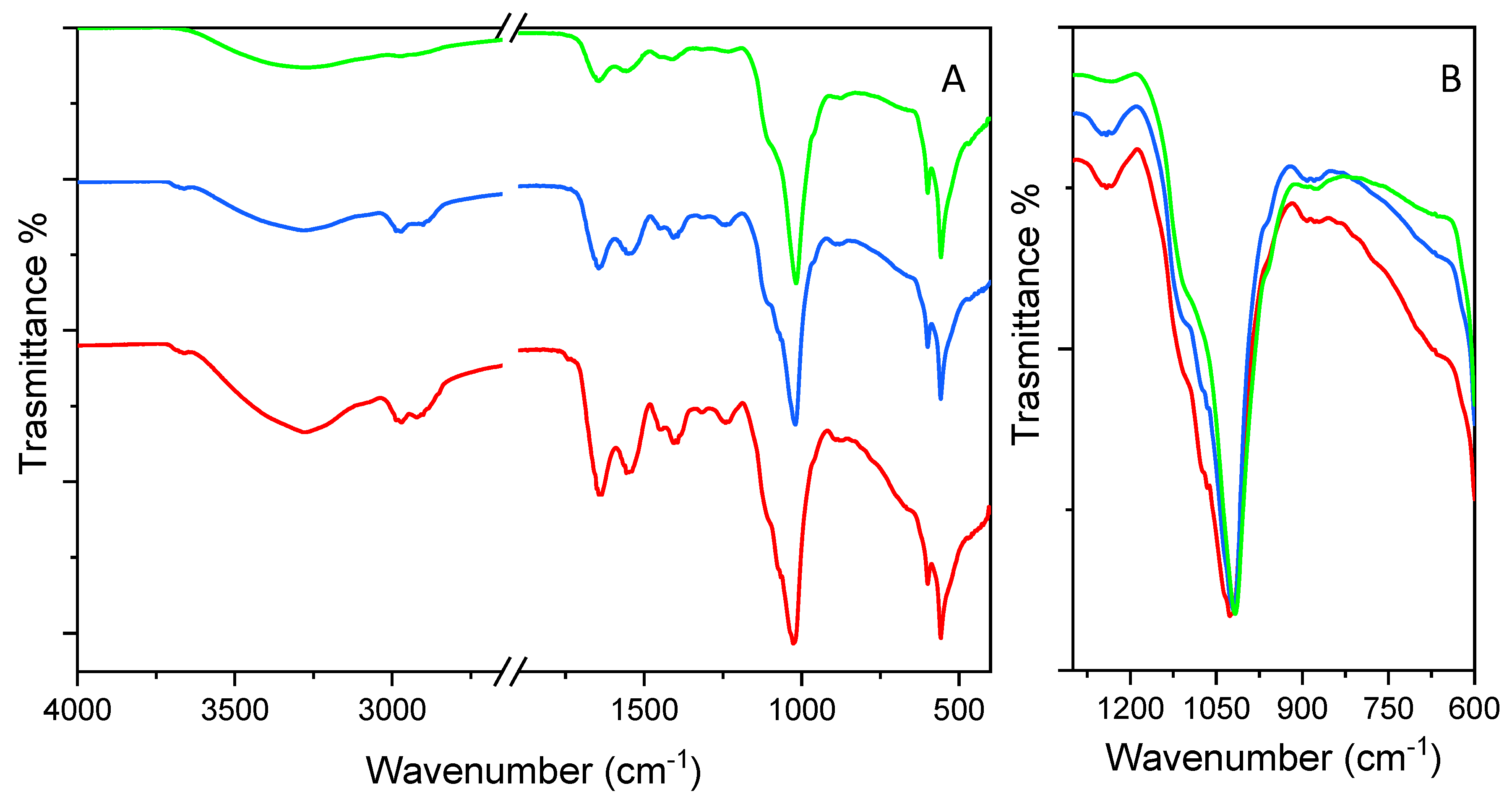
3.7. FT-IR
3.8. Microstructure of Freeze-Dried EPS
4. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Farag, M.A.; Jomaa, S.A.; Abd El-Wahed, A.; El-Seedi, H.R. The many faces of kefir fermented dairy products: Quality characteristics, flavour chemistry, nutritional value, health benefits, and safety. Nutrients 2020, 12, 346. [Google Scholar] [CrossRef] [PubMed]
- Izquierdo-González, J.J.; Amil-Ruiz, F.; Zazzu, S.; Sánchez-Lucas, R.; Fuentes-Almagro, C.A.; Rodríguez-Ortega, M.J. Proteomic analysis of goat milk kefir: Profiling the fermentation-time dependent protein digestion and identification of potential peptides with biological activity. Food Chem. 2019, 295, 456–465. [Google Scholar] [CrossRef] [PubMed]
- Wang, H.; Sun, X.; Song, X.; Guo, M. Effects of kefir grains from different origins on proteolysis and volatile profile of goat milk kefir. Food Chem. 2021, 339, 128099. [Google Scholar] [CrossRef] [PubMed]
- El, S.N.; Karakaya, S.; Simsek, S.; Dupont, D.; Menfaatli, E.; Eker, A.T. In vitro digestibility of goat milk and kefir with a new standardised static digestion method (INFOGEST cost action) and bioactivities of the resultant peptides. Food Funct. 2015, 6, 2322–2330. [Google Scholar]
- Dalabasmaz, S.; De la Torre, E.P.; Gensberger-Reigl, S.; Pischetsrieder, M.; Rodríguez-Ortega, M.J. Identification of Potential Bioactive Peptides in Sheep Milk Kefir through Peptidomic Analysis at Different Fermentation Times. Foods 2023, 12, 2974. [Google Scholar] [CrossRef] [PubMed]
- Ferreira, I.M.P.L.V.O.; Pinho, O.; Monteiro, D.; Faria, S.; Cruz, S.; Perreira, A.; Tavares, P. Effect of kefir grains on proteolysis of major milk proteins. J. Dairy Sci. 2010, 93, 27–31. [Google Scholar] [CrossRef]
- Gentry, B.; Cazón, P.; O’Brien, K. A comprehensive review of the production, beneficial properties, and applications of kefiran, the kefir grain exopolysaccharide. Int. Dairy J. 2023, 144, 105691. [Google Scholar] [CrossRef]
- De Carvalho, A.P.A.; Conte-Junior, C.A. Food-derived biopolymer kefiran composites, nanocomposites and nanofibers: Emerging alternatives to food packaging and potentials in nanomedicine. Trends Food Sci. Technol. 2021, 116, 370–386. [Google Scholar] [CrossRef]
- Bengoa, A.A.; Dueñas, M.T.; Prieto, A.; Garrote, G.L.; Abraham, A.G. Exopolysaccharide-producing Lacticaseibacillus paracasei strains isolated from kefir as starter for functional dairy products. Front. Microbiol. 2023, 14, 1110177. [Google Scholar] [CrossRef]
- Jiang, S.J.; Qian, F.; Ren, X.H.; Mu, G.Q. Studies on the preliminary characterization of a novel exopolysaccharide produced by Streptococcus thermophilus strain from Tibetan kefir grain. Adv. Mat. Res. 2013, 690, 1374–1377. [Google Scholar] [CrossRef]
- Wang, L.; Gu, Y.; Zheng, X.; Zhang, Y.; Deng, K.; Wu, T.; Cheng, H. Analysis of physicochemical properties of exopolysaccharide from Leuconostoc mesenteroides strain XR1 and its application in fermented milk. LWT 2021, 146, 111449. [Google Scholar] [CrossRef]
- Chen, Z.; Shi, J.; Yang, X.; Nan, B.; Liu, Y.; Wang, Z. Chemical and physical characteristics and antioxidant activities of the exopolysaccharide produced by Tibetan kefir grains during milk fermentation. Int. Dairy J. 2015, 43, 15–21. [Google Scholar] [CrossRef]
- Wang, W.; Ju, Y.; Liu, N.; Shi, S.; Hao, L. Structural characteristics of microbial exopolysaccharides in association with their biological activities: A review. Chem. Biol. Technol. Agric. 2023, 10, 137. [Google Scholar] [CrossRef]
- Adebayo-Tayo, B.; Ishola, R.; Oyewunmi, T. Characterization, antioxidant and immunomodulatory potential on exopolysaccharide produced by wild type and mutant Weissella confusa strains. Biotechnol. Rep. 2018, 9, e00271. [Google Scholar] [CrossRef]
- Zhao, Z.Y.; Zhang, Q.; Li, Y.F.; Dong, L.L.; Liu, S.L. Optimization of ultrasound extraction of Alisma orientalis polysaccharides by response surface methodology and their antioxidant activities. Carbohydr. Polym. 2015, 119, 101–109. [Google Scholar] [CrossRef]
- Jahanbin, K.; Gohari, A.R.; Moini, S.; Emam-Djomeh, Z.; Masi, P. Isolation, structural characterization and antioxidant activity of a new water-soluble polysaccharide from Acanthophyllum bracteatum roots. Int. J. Biol. Macromol. 2011, 49, 567–572. [Google Scholar] [CrossRef]
- Zhang, Z.; Lv, G.; He, W.; Shi, L.; Pan, H.; Fan, L. Effects of extraction methods on the antioxidant activities of polysaccharides obtained from Flammulina velutipes. Carbohydr. Polym. 2013, 98, 1524–1531. [Google Scholar] [CrossRef]
- Hasheminya, S.M.; Dehghannya, J. Novel ultrasound-assisted extraction of kefiran biomaterial, a prebiotic exopolysaccharide, and investigation of its physicochemical, antioxidant and antimicrobial properties. Mater. Chem. Phys. 2020, 243, 122645. [Google Scholar] [CrossRef]
- Andrew, M.; Jayaraman, G. Structural features of microbial exopolysaccharides in relation to their antioxidant activity. Carbohydr. Res. 2020, 487, 107881. [Google Scholar] [CrossRef] [PubMed]
- Buran, İ.; Akal, C.; Ozturkoglu-Budak, S.; Yetisemiyen, A. Rheological, sensorial and volatile profiles of synbiotic kefirs produced from cow and goat milk containing varied probiotics in combination with fructooligosaccharide. LWT 2021, 148, 111591. [Google Scholar] [CrossRef]
- Al-Kaisy, Q.H.; Al-Saadi, J.S.; Al-Rikabi, A.K.J.; Altemimi, A.B.; Hesarinejad, M.A.; Abedelmaksoud, T.G. Exploring the health benefits and functional properties of goat milk proteins. Food Sci. Nutr. 2023, 11, 5641–5656. [Google Scholar] [CrossRef]
- Flis, Z.; Molik, E. Importance of Bioactive Substances in Sheep’s Milk in Human Health. Int. J. Mol. Sci. 2021, 22, 4364. [Google Scholar] [CrossRef]
- Prado, M.R.; Blandón, L.M.; Vandenberghe, L.P.; Rodrigues, C.; Castro, G.R.; Thomaz-Soccol, V.; Soccol, C.R. Milk kefir: Composition, microbial cultures, biological activities, and related products. Front. Microbiol. 2015, 6, 1177. [Google Scholar] [CrossRef] [PubMed]
- Zhong, Z.; Hou, Q.; Kwok, L.; Yu, Z.; Zheng, Y.; Sun, Z.; Zhang, H. Bacterial microbiota compositions of naturally fermented milk are shaped by both geographic origin and sample type. J. Dairy Sci. 2016, 99, 7832–7841. [Google Scholar] [CrossRef] [PubMed]
- Kim, Y.H.; Kim, J.U.; Oh, S.J.; Kim, Y.J.; Kim, M.H.; Kim, S.H. Technical optimization of culture conditions for the production of exopolysaccharide (EPS) by Lactobacillus rhamnosus ATCC 9595. Food Sci. Biotechnol. 2008, 17, 587–593. [Google Scholar]
- Wang, X.; Li, W.; Xu, M.; Tian, J.; Li, W. The microbial diversity and biofilm-forming characteristic of two traditional Tibetan kefir grains. Foods 2021, 11, 12. [Google Scholar] [CrossRef] [PubMed]
- Luang-In, V.; Deeseenthum, S. Exopolysaccharide-producing isolates from Thai milk kefir and their antioxidant activities. LWT 2016, 73, 592–601. [Google Scholar] [CrossRef]
- M’hir, S.; Rtibi, K.; Ayed, L.; Hamdi, M.; Marzouki, L.; Sebai, H. Evaluation de l’effet protecteur du lait de chèvre fermenté par le kefir et enrichi à la caroube contre l’ulcère gastrique induit par l’étahnol chez le rat. Int. J. Adv. Res. 2019, 7, 1019–1028. [Google Scholar] [CrossRef]
- Kim, D.-H.; Jeong, D.; Song, K.-Y.; Seo, K.-H. Comparison of traditional and backslopping methods for kefir fermentation based on physicochemical and microbiological characteristics. LWT 2018, 97, 503–507. [Google Scholar] [CrossRef]
- Alves, E.; Ntungwe, E.N.; Gregório, J.; Rodrigues, L.M.; Pereira-Leite, C.; Caleja, C.; Pereira, E.; Barros, L.; Aguilar-Vilas, M.V.; Rosado, C.; et al. Characterization of kefir produced in household conditions: Physicochemical and nutritional profile, and storage stability. Foods. 2021, 10, 1057. [Google Scholar] [CrossRef]
- Tlais, A.Z.A.; Kanwal, S.; Filannino, P.; Albiac, M.A.; Gobbetti, M.; Di Cagno, R. Effect of sequential or ternary starters-assisted fermentation on the phenolic and glucosinolate profiles of sauerkraut in comparison with spontaneous fermentation. Food Res. Int. 2022, 156, 111116. [Google Scholar] [CrossRef] [PubMed]
- Verni, M.; Dingeo, C.; Rizzello, C.G.; Pontonio, E. Lactic Acid Bacteria Fermentation and Endopeptidase Treatment Improve the Functional and Nutritional Features of Arthrospira platensis. Front. Microbiol. 2021, 12, 744437. [Google Scholar] [CrossRef]
- Bouallegue, A.; Casillo, A.; Chaari, F.; La Gatta, A.; Lanzetta, R.; Corsaro, M.M.; Bachoual, R.; Ellouz-Chaabouni, S. Levan from a new isolated Bacillus subtilis AF17: Purification, structural analysis and antioxidant activities. Int. J. Biol. Macromol. 2020, 144, 316–324. [Google Scholar] [CrossRef] [PubMed]
- Groth, S.; Budke, C.; Neugart, S.; Ackermann, S.; Kappenstein, F.S.; Daum, D.; Rohn, S. Influence of a selenium biofortification on antioxidant properties and phenolic compounds of apples (Malus domestica). Antioxidants 2020, 9, 187. [Google Scholar] [CrossRef] [PubMed]
- Di Cagno, R.; De Angelis, M.; Limitone, A.; Minervini, F.; Carnevali, P.; Corsetti, A.; Gäenzle, M.; Ciati, R.; Gobbetti, M. Glucan and fructan production by sourdough Weissella cibaria and Lactobacillus plantarum. J. Agric. Food Chem. 2006, 54, 9873–9881. [Google Scholar] [CrossRef] [PubMed]
- M’hir, S. (National Institute of Applied Sciences and Technology (INSAT), Tunis, Tunisia). Unpublished microbiological data. 2023. [Google Scholar]
- Öner, Z.; Karahan, A.G.; Çakmakçı, M.L. Effects of different milk types and starter cultures on kefir. GIDA 2015, 35, 177–182. [Google Scholar]
- Guangsen, T.; Xiang, L.; Jiahu, G. Microbial diversity and volatile metabolites of kefir prepared by different milk types. CYTA J. Food 2021, 19, 399–407. [Google Scholar] [CrossRef]
- Horne, D.S. Casein interactions: Casting light on the black boxes, the structure in dairy products. Int. Dairy J. 1998, 8, 171–177. [Google Scholar] [CrossRef]
- Tamime, A.Y.; Robinson, R.K. Tamime and Robinson’s Yoghurt—Science and Technology, 3rd ed.; Woodhead Publishing Ltd.: Cambridge, UK, 2007; pp. 13–161. [Google Scholar]
- Tamime, A.Y.; Robinson, R.K. Background to manufacturing practice. In Yoghurt: Science and Technology; Tamime, A.Y., Robinson, R.K., Eds.; Woodhead Publishing Ltd.: Cambridge, UK, 1999; pp. 11–128. [Google Scholar]
- Yang, T.; Wu, K.; Wang, F.; Liang, X.; Liu, Q.; Li, G.; Li, Q. Effect of exopolysaccharides from lactic acid bacteria on the texture and microstructure of buffalo yoghurt. Int. Dairy J. 2014, 34, 252–256. [Google Scholar] [CrossRef]
- Amatayakul, T.; Halmos, A.L.; Sherkat, F.; Shah, N.P. Physical characteristics of yoghurts made using exopolysaccharide-producing starter cultures and varying casein to whey protein ratios. Int. Dairy J. 2006, 16, 40–51. [Google Scholar] [CrossRef]
- Ruas-Madiedo, P.; Tuinier, R.; Kanning, M.; Zoon, P. Role of exopolysaccharides produced by Lactococcus lactis subsp. cremoris on the viscosity of fermented milks. Int. Dairy J. 2002, 12, 689–695. [Google Scholar]
- Koroleva, N.S. Starters for fermented milks. Section 4, Kefir and Kumys starters. Bulletion IDF 1975, 227, 35–40. [Google Scholar]
- Saygili, D.; Döner, D.; İçier, F.; Karagözlü, C. Rheological properties and microbiological characteristics of kefir produced from different milk types. Food Sci. Technol. 2021, 42, e32520. [Google Scholar] [CrossRef]
- Rizzello, C.G.; Portincasa, P.; Montemurro, M.; Di Palo, D.M.; Lorusso, M.P.; De Angelis, M.; Bonfrate, L.; Genot, B.; Gobbetti, M. Sourdough Fermented Breads are More Digestible than Those Started with Baker’s Yeast Alone: An In Vivo Challenge Dissecting Distinct Gastrointestinal Responses. Nutrients 2019, 11, 2954. [Google Scholar] [CrossRef] [PubMed]
- Gamba, R.R.; Yamamoto, S.; Abdel-Hamid, M.; Sasaki, T.; Michihata, T.; Koyanagi, T.; Enomoto, T. Chemical, Microbiological, and Functional Characterization of Kefir Produced from Cow’s Milk and Soy Milk. Int. J. Microbiol. 2020, 2020, e7019286. [Google Scholar] [CrossRef]
- Collar, C.; Mascarós, A.F.; Barber, C.B. Amino Acid Metabolism by Yeasts and Lactic Acid Bacteria during Bread Dough Fermentation. J. Food Sci. 1992, 57, 1423–1427. [Google Scholar] [CrossRef]
- Grønnevik, H.; Falstad, M.; Narvhus, J.A. Microbiological and chemical properties of Norwegian kefir during storage. Int. Dairy J. 2011, 21, 601–606. [Google Scholar] [CrossRef]
- Ziarno, M.; Hasalliu, R.; Cwalina, A. Effect of the Addition of Milk Protein Preparations on Selected Quality Parameters and Nutritional Characteristics of Kefir. Appl. Sci. 2021, 11, 966. [Google Scholar] [CrossRef]
- Rimada, P.S.; Abraham, A.G. Polysaccharide production by kefir grains during whey fermentation. J. Dairy Res. 2021, 68, 653–661. [Google Scholar] [CrossRef]
- Wang, Y.; Ahmed, Z.; Feng, W.; Li, C.; Song, S. Physicochemical properties of exopolysaccharide produced by Lactobacillus kefiranofaciens ZW3 isolated from Tibet kefir. Int. J. Biol. Macromol. 2008, 43, 283–288. [Google Scholar] [CrossRef] [PubMed]
- Radhouani, H.; Gonçalves, C.; Maia, F.R.; Oliveira, J.M.; Reis, R.L. Biological performance of a promising Kefiran-biopolymer with potential in regenerative medicine applications: A comparative study with hyaluronic acid. J. Mater. Sci. Mater. Med. 2018, 29, 1–10. [Google Scholar] [CrossRef]
- Blandón, L.M.; Noseda, M.D.; Islan, G.A.; Castro, G.R.; De Melo Pereira, G.V.; Thomaz-Soccol, V.; Soccol, C.R. Optimization of culture conditions for kefiran production in whey: The structural and biocidal properties of the resulting polysaccharide. Bioact. Carbohydr. Diet. Fibre 2018, 16, 14–21. [Google Scholar] [CrossRef]
- Xiao, L.; Xu, D.; Tang, N.; Rui, X.; Zhang, Q.; Chen, X.; Li, W. Biosynthesis of exopolysaccharide and structural characterization by Lacticaseibacillus paracasei ZY-1 isolated from Tibetan kefir. Food Chem. 2021, 3, 100054. [Google Scholar] [CrossRef] [PubMed]
- Koçak, Ç.; Demiralay, E.Ç.; Özarslan, S.; Aydoğdu, N.S.; Taş, T.K. Determination of monosaccharide composition of kefiran using HPLC. Mljekarstvo 2021, 71, 49–59. [Google Scholar] [CrossRef]
- Ahmed, Z.; Wang, Y.; Anjum, N.; Ahmad, A.; Khan, S.T. Characterization of exopolysaccharide produced by Lactobacillus kefiranofaciens ZW3 isolated from Tibet kefir–Part II. Food Hydrocoll. 2013, 30, 343–350. [Google Scholar] [CrossRef]
- Ahmed, Z.; Wang, Y.; Anjum, N.; Ahmad, H.; Ahmad, A.; Raza, M. Characterization of new exopolysaccharides produced by coculturing of L. kefiranofaciens with yoghurt strains. Int. J. Biol. Macromol. 2013, 59, 377–383. [Google Scholar] [CrossRef] [PubMed]
- Zannini, E.; Lynch, K.M.; Nyhan, L.; Sahin, A.W.; O’Riordan, P.; Luk, D.; Arendt, E.K. Influence of substrate on the fermentation characteristics and culture-dependent microbial composition of water kefir. Fermentation 2022, 9, 28. [Google Scholar] [CrossRef]
- Li, W.; Ji, J.; Chen, X.; Jiang, M.; Rui, X.; Dong, M. Structural elucidation and antioxidant activities of exopolysaccharides from Lactobacillus helveticus MB2-1. Carbohydr. Polym. 2014, 102, 351–359. [Google Scholar] [CrossRef] [PubMed]
- Qiao, D.; Ke, C.; Hu, B.; Luo, J.; Ye, H.; Sun, Y.; Zeng, X. Antioxidant activities of polysaccharides from Hyriopsis cumingii. Carbohydr. Polym. 2009, 78, 199–204. [Google Scholar] [CrossRef]
- Qiao, D.; Liu, J.; Ke, C.; Sun, Y.; Ye, H.; Zeng, X. Structural characterization of polysaccharides from Hyriopsis cumingii. Carbohydr. Polym. 2010, 82, 1184–1190. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| TTA (mL 0.1 M NaOH per 10 mL) | pH | Viscosity (mPa·s) | L* | a* | b* | ΔE*ab | |
|---|---|---|---|---|---|---|---|
| CM | 0.2 ± 0.1 c | 6.58 ± 0.03 b | 1.11 ± 0.08 c | 68.17 ± 1.52 b | −6.20 ± 0.18 a | 12.65 ± 0.88 b | - |
| EM | 0.2 ± 0.1 c | 6.61 ± 0.02 b | 1.15 ± 0.11 c | 78.03 ± 0.71 a | −6.82 ± 0.10 b | 12.70 ± 0.16 b | - |
| GM | 0.2 ± 0.1 c | 6.73 ± 0.02 a | 1.13 ± 0.12 c | 70.12 ± 0.49 b | −7.01 ± 0.14 b | 10.02 ± 0.17 c | - |
| FCM | 0.9 ± 0.1 b | 4.39 ± 0.04 c | 1.59 ± 0.15 ab | 76.97 ± 0.51 a | −6.85 ± 0.11 b | 17.50 ± 0.14 a | 10.07 ± 2.22 a |
| FEM | 2.1 ± 0.3 a | 4.06 ± 0.05 e | 1.37 ± 0.17 bc | 76.96 ± 0.42 a | −6.11 ± 0.18 a | 11.96 ± 0.30 b | 1.48 ± 0.34 b |
| FGM | 1.0 ± 0.2 b | 4.19 ± 0.07 d | 1.62 ± 0.13 ab | 69.87 ± 1.07 b | −6.30 ± 0.14 a | 10.57 ± 0.17 c | 1.05 ± 0.13 b |
| Lactose | Glucose | Galactose | Xylitol | Lactic Acid | Acetic Acid | Ethanol | |
|---|---|---|---|---|---|---|---|
| CM | 49.94 ± 1.77 a | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| EM | 45.63 ± 0.39 b | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| GM | 42.74 ± 0.38 c | n.d. | n.d. | n.d. | n.d. | n.d. | n.d. |
| FCM | 33.20 ± 3.47 d | n.d. | 0.80 ± 0.05 | 0.09 ± 0.01 b | 15.21 ± 0.91 b | 0.35 ± 0.02 b | n.d. |
| FEM | 23.04 ± 0.39 e | 0.88 ± 0.02 | 0.88 ± 0.02 | 0.23 ± 0.01 a | 21.80 ± 0.51 a | 0.43 ± 0.01 a | n.d. |
| FGM | 24.19 ± 1.87 e | n.d. | 0.89 ± 0.05 | 0.08 ± 0.01 b | 14.06 ± 0.98 b | 0.29 ± 0.01 c | n.d. |
| FCM (cm−1) | FEM (cm−1) | FGM (cm−1) | |
|---|---|---|---|
| νs OH monomer H-bonded | 3660 | 3660 | |
| νs OH dimer H-bond | 3278 | 3278 | 3278 |
| νs CH3 | 2986 | 2986 | 2986 |
| νs CH2 | 2970 | 2970 | 2970 |
| νas CH3 | 2922 | 2922 | 2922 |
| νas CH2 | 2901 | 2901 | 2901 |
| C=O carboxylic group | 1730 | ||
| δ H2O/ν C=O amide I and δ N-H amide II | 1643 | 1643 | 1643 |
| νas COO−/δ N-H amide II coupled with ν C-N | 1547 | 1547 | 1547 |
| δs CH2 | 1454 | 1454 | 1454 |
| νs COO−/ν C-N | 1406 | 1406 | 1406 |
| amide III and C-O stretching mode | 1318 1244 | 1318 1244 | 1318 1244 |
| C-C-O stretching modes/O-P=O modes | 1100, 1080, 1024 | 1100, 1022 | 1100, 1080, 1022 |
| δOH | 950 | 950 | 950 |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2024 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
M’hir, S.; Ayed, L.; De Pasquale, I.; Fanizza, E.; Tlais, A.Z.A.; Comparelli, R.; Verni, M.; Latronico, R.; Gobbetti, M.; Di Cagno, R.; et al. Comparison of Milk Kefirs Obtained from Cow’s, Ewe’s and Goat’s Milk: Antioxidant Role of Microbial-Derived Exopolysaccharides. Antioxidants 2024, 13, 335. https://doi.org/10.3390/antiox13030335
M’hir S, Ayed L, De Pasquale I, Fanizza E, Tlais AZA, Comparelli R, Verni M, Latronico R, Gobbetti M, Di Cagno R, et al. Comparison of Milk Kefirs Obtained from Cow’s, Ewe’s and Goat’s Milk: Antioxidant Role of Microbial-Derived Exopolysaccharides. Antioxidants. 2024; 13(3):335. https://doi.org/10.3390/antiox13030335
Chicago/Turabian StyleM’hir, Sana, Lamia Ayed, Ilaria De Pasquale, Elisabetta Fanizza, Ali Zein Alabiden Tlais, Roberto Comparelli, Michela Verni, Rosanna Latronico, Marco Gobbetti, Raffaella Di Cagno, and et al. 2024. "Comparison of Milk Kefirs Obtained from Cow’s, Ewe’s and Goat’s Milk: Antioxidant Role of Microbial-Derived Exopolysaccharides" Antioxidants 13, no. 3: 335. https://doi.org/10.3390/antiox13030335