
Placental Malfunction, Fetal Survival and Development Caused by Sow Metabolic Disorder: The Impact of Maternal Oxidative Stress
 , and
, and
Abstract
:1. Introduction
2. Maternal Energy and Metabolism Changes during Pregnancy
2.1. Early Pregnancy
2.2. The Second Trimester of Pregnancy
2.3. Late Pregnancy
2.4. Progressive Oxidative Stress during Pregnancy of Sows
3. Placenta and Oxidative Stress
3.1. The Structural Characteristics of Placenta
3.2. Mechanism of Oxidative Stress in Placenta
3.3. Effects of Oxidative Stress on Placental Function
4. Effect of Oxidative Stress on Fetus
4.1. Fetal Development during Pregnancy
4.2. Effects of Maternal Oxidative Stress on Offspring in Sows
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Guan, R.; Zhou, X.; Cai, H.; Qian, X.; Xin, X.; Li, X. Study on the influence of different production factors on PSY and its correlation. Porc. Health Manag. 2022, 8, 9. [Google Scholar] [CrossRef]
- Lavery, A.; Lawlor, P.G.; Magowan, E.; Miller, H.M.; O’Driscoll, K.; Berry, D.P. An association analysis of sow parity, live-weight and back-fat depth as indicators of sow productivity. Animal 2019, 13, 622–630. [Google Scholar] [CrossRef]
- Oliviero, C.; Junnikkala, S.; Peltoniemi, O. The challenge of large litters on the immune system of the sow and the piglets. Reprod. Domest. Anim. 2019, 54 (Suppl. S3), 12–21. [Google Scholar] [CrossRef] [PubMed]
- Quesnel, H.; Brossard, L.; Valancogne, A.; Quiniou, N. Influence of some sow characteristics on within-litter variation of piglet birth weight. Animal 2008, 2, 1842–1849. [Google Scholar] [CrossRef] [PubMed]
- Yuan, T.L.; Zhu, Y.H.; Shi, M.; Li, T.T.; Li, N.; Wu, G.Y.; Bazer, F.W.; Zang, J.J.; Wang, F.L.; Wang, J.J. Within-litter variation in birth weight: Impact of nutritional status in the sow. J. Zhejiang Univ. Sci. B 2015, 16, 417–435. [Google Scholar] [CrossRef] [PubMed]
- Xue, Y.; Guo, C.; Hu, F.; Zhu, W.; Mao, S. Maternal undernutrition induces fetal hepatic lipid metabolism disorder and affects the development of fetal liver in a sheep model. FASEB J. 2019, 33, 9990–10004. [Google Scholar] [CrossRef] [PubMed]
- Furse, S.; Fernandez-Twinn, D.S.; Chiarugi, D.; Koulman, A.; Ozanne, S.E. Lipid Metabolism Is Dysregulated before, during and after Pregnancy in a Mouse Model of Gestational Diabetes. Int. J. Mol. Sci. 2021, 22, 7452. [Google Scholar] [CrossRef]
- Johns, E.C.; Denison, F.C.; Norman, J.E.; Reynolds, R.M. Gestational Diabetes Mellitus: Mechanisms, Treatment, and Complications. Trends Endocrinol. Metab. 2018, 29, 743–754. [Google Scholar] [CrossRef]
- Liu, L.X.; Arany, Z. Maternal cardiac metabolism in pregnancy. Cardiovasc. Res. 2014, 101, 545–553. [Google Scholar] [CrossRef]
- Wu, G.; Bazer, F.W.; Wallace, J.M.; Spencer, T.E. Board-invited review: Intrauterine growth retardation: Implications for the animal sciences. J. Anim. Sci. 2006, 84, 2316–2337. [Google Scholar] [CrossRef]
- Roseboom, T.J.; Painter, R.C.; de Rooij, S.R.; van Abeelen, A.F.M.; Veenendaal, M.V.E.; Osmond, C.; Barker, D.J.P. Effects of famine on placental size and efficiency. Placenta 2011, 32, 395–399. [Google Scholar] [CrossRef] [PubMed]
- Sadovsky, Y.; Mouillet, J.F.; Ouyang, Y.; Bayer, A.; Coyne, C.B. The Function of TrophomiRs and Other MicroRNAs in the Human Placenta. Cold Spring Harb. Perspect. Med. 2015, 5, a023036. [Google Scholar] [CrossRef] [PubMed]
- Tarrade, A.; Panchenko, P.; Junien, C.; Gabory, A. Placental contribution to nutritional programming of health and diseases: Epigenetics and sexual dimorphism. J. Exp. Biol. 2015, 218, 50–58. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Huang, Z.; Xiong, W.; Ye, H.; Deng, J.; Yin, Y. A review of the amino acid metabolism in placental function response to fetal loss and low birth weight in pigs. J. Anim. Sci. Biotechnol. 2022, 13, 28. [Google Scholar] [CrossRef] [PubMed]
- Pinto, C.R.F. Impact of the corpus luteum on survival of the developing embryo and early pregnancy in mares. Theriogenology 2020, 150, 374–381. [Google Scholar] [CrossRef]
- Langendijk, P. Latest Advances in Sow Nutrition during Early Gestation. Animals 2021, 11, 1720. [Google Scholar] [CrossRef]
- Anderson, L.L.; Dyck, G.W.; Mori, H.; Henricks, D.M.; Melampy, R.M. Ovarian function in pigs following hypophysial stalk transection or hypophysectomy. Am. J. Physiol. 1967, 212, 1188–1194. [Google Scholar] [CrossRef] [PubMed]
- Tast, A.; Love, R.J.; Clarke, I.J.; Evans, G. Effects of active and passive gonadotrophin-releasing hormone immunization on recognition and establishment of pregnancy in pigs. Reprod. Fertil. Dev. 2000, 12, 277–282. [Google Scholar] [CrossRef]
- Qian, Z.; Zhu, H.; Lv, Y.; Liu, H.; Bao, E. Impact of exogenous adrenocorticotropic hormone on gelatinase expression and steroidogenesis in the newly formed corpus luteum in sows. Livest. Sci. 2018, 207, 68–74. [Google Scholar] [CrossRef]
- Leung, K.C.; Xu, A.; Craig, M.E.; Martin, A.; Lam, K.S.; O’Sullivan, A.J. Adiponectin isoform distribution in women--relationship to female sex steroids and insulin sensitivity. Metabolism 2009, 58, 239–245. [Google Scholar] [CrossRef]
- Alminana, C.; Heath, P.R.; Wilkinson, S.; Sanchez-Osorio, J.; Cuello, C.; Parrilla, I.; Gil, M.A.; Vazquez, J.L.; Vazquez, J.M.; Roca, J.; et al. Early developing pig embryos mediate their own environment in the maternal tract. PLoS ONE 2012, 7, e33625. [Google Scholar] [CrossRef] [PubMed]
- Lain, K.Y.; Catalano, P.M. Metabolic changes in pregnancy. Clin. Obstet. Gynecol. 2007, 50, 938–948. [Google Scholar] [CrossRef] [PubMed]
- Napso, T.; Yong, H.E.J.; Lopez-Tello, J.; Sferruzzi-Perri, A.N. The Role of Placental Hormones in Mediating Maternal Adaptations to Support Pregnancy and Lactation. Front. Physiol. 2018, 9, 1091. [Google Scholar] [CrossRef]
- Jindal, R.; Cosgrove, J.R.; Foxcroft, G.R. Progesterone mediates nutritionally induced effects on embryonic survival in gilts. J. Anim. Sci. 1997, 75, 1063–1070. [Google Scholar] [CrossRef] [PubMed]
- Mattos, F.; Canavessi, A.M.O.; Wiltbank, M.C.; Bastos, M.R.; Lemes, A.P.; Mourao, G.B.; Susin, I.; Coutinho, L.L.; Sartori, R. Investigation of mechanisms involved in regulation of progesterone catabolism using an overfed versus underfed ewe-lamb model. J. Anim. Sci. 2017, 95, 5537–5546. [Google Scholar] [CrossRef] [PubMed]
- Fraser, R.B.; Waite, S.L.; Wood, K.A.; Martin, K.L. Impact of hyperglycemia on early embryo development and embryopathy: In vitro experiments using a mouse model. Hum. Reprod. 2007, 22, 3059–3068. [Google Scholar] [CrossRef]
- Dicks, N.; Gutierrez, K.; Currin, L.; de Macedo, M.P.; Glanzner, W.G.; Mondadori, R.G.; Michalak, M.; Agellon, L.B.; Bordignon, V. Tauroursodeoxycholic acid/TGR5 signaling promotes survival and early development of glucose-stressed porcine embryos. Biol. Reprod. 2021, 105, 76–86. [Google Scholar] [CrossRef]
- Leal, D.F.; Muro, B.B.D.; Nichi, M.; Almond, G.W.; Viana, C.H.C.; Vioti, G.; Carnevale, R.F.; Garbossa, C.A.P. Effects of post-insemination energy content of feed on embryonic survival in pigs: A systematic review. Anim. Reprod. Sci. 2019, 205, 70–77. [Google Scholar] [CrossRef]
- Virolainen, J.V.; Tast, A.; Sorsa, A.; Love, R.J.; Peltoniemi, O.A. Changes in feeding level during early pregnancy affect fertility in gilts. Anim. Reprod. Sci. 2004, 80, 341–352. [Google Scholar] [CrossRef]
- Carrion-Lopez, M.J.; Madrid, J.; Martinez, S.; Hernandez, F.; Orengo, J. Effects of the feeding level in early gestation on body reserves and the productive and reproductive performance of primiparous and multiparous sows. Res. Vet. Sci. 2022, 148, 42–51. [Google Scholar] [CrossRef]
- Piao, L.G.; Ju, W.S.; Long, H.F.; Kim, Y.Y. Effects of Various Feeding Methods for Gestating Gilts on Reproductive Performance and Growth of Their Progeny. Asian Australas J. Anim. Sci. 2010, 23, 1354–1363. [Google Scholar] [CrossRef]
- Cerisuelo, A.; Sala, R.; Gasa, J.; Carrión, D.; Coma, J.; Chapinal, N.; Baucells, M.D. Effects of extra feeding in mid-pregnancy for three successive parities on lean sows’ productive performance and longevity. Can. J. Anim. Sci. 2010, 90, 521–528. [Google Scholar] [CrossRef]
- Scott, E.M.; Feig, D.S.; Murphy, H.R.; Law, G.R.; Grp, C.C. Continuous Glucose Monitoring in Pregnancy: Importance of Analyzing Temporal Profiles to Understand Clinical Outcomes. Diabetes Care 2020, 43, 1178–1184. [Google Scholar] [CrossRef] [PubMed]
- Hedderson, M.M.; Ferrara, A.; Sacks, D.A. Gestational diabetes mellitus and lesser degrees of pregnancy hyperglycemia: Association with increased risk of spontaneous preterm birth. Obstet. Gynecol. 2003, 102, 850–856. [Google Scholar] [CrossRef]
- Kerssen, A.; de Valk, H.W.; Visser, G.H.A. Increased second trimester maternal glucose levels are related to extremely large-for-gestational-age infants in women with type 1 diabetes. Diabetes Care 2007, 30, 1069–1074. [Google Scholar] [CrossRef] [PubMed]
- Riskin-Mashiah, S.; Damti, A.; Younes, G.; Auslander, R. Normal fasting plasma glucose levels during pregnancy: A hospital-based study. J. Perinat. Med. 2011, 39, 209–211. [Google Scholar] [CrossRef]
- George, P.B.; England, D.C.; Siers, D.G.; Stanton, H.C. Diabetogenic effects of pregnancy in sows on plasma glucose and insulin release. J. Anim. Sci. 1978, 46, 1694–1700. [Google Scholar] [CrossRef]
- Yang, Y.; Heo, S.; Jin, Z.; Yun, J.; Shinde, P.; Choi, J.; Yang, B.; Chae, B. Effects of dietary energy and lysine intake during late gestation and lactation on blood metabolites, hormones, milk composition and reproductive performance in multiparous sows. Arch. Anim. Nutr. 2008, 62, 10–21. [Google Scholar] [CrossRef]
- Goncalves, M.A.D.; Gourley, K.M.; Dritz, S.S.; Tokach, M.D.; Bello, N.M.; DeRouchey, J.M.; Woodworth, J.C.; Goodband, R.D. Effects of amino acids and energy intake during late gestation of high-performing gilts and sows on litter and reproductive performance under commercial conditions. J. Anim. Sci. 2016, 94, 1993–2003. [Google Scholar] [CrossRef]
- Wang, W.H.; Wang, Z.J.; Ming, D.X.; Huang, C.Y.; Xu, S.; Li, Z.; Wang, Z.Y.; Liu, H.; Zeng, X.F.; Wang, F.L. Effect of maternal dietary starch-to-fat ratio and daily energy intake during late pregnancy on the performance and lipid metabolism of primiparous sows and newborn piglets. J. Anim. Sci. 2022, 100, skac033. [Google Scholar] [CrossRef]
- Zhou, Y.; Xu, T.; Cai, A.; Wu, Y.; Wei, H.; Jiang, S.; Peng, J. Excessive backfat of sows at 109 d of gestation induces lipotoxic placental environment and is associated with declining reproductive performance. J. Anim. Sci. 2018, 96, 250–257. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.H.; Zhang, X.M.; Zhou, Y.F.; Wang, C.; Xiong, J.; Guo, L.L.; Wang, L.; Jiang, S.W.; Peng, J. Effect of increasing feed intake during late gestation on piglet performance at parturition in commercial production enterprises. Anim. Reprod. Sci. 2020, 218, 106477. [Google Scholar] [CrossRef] [PubMed]
- Trujillo, M.L.; Spuch, C.; Carro, E.; Senaris, R. Hyperphagia and central mechanisms for leptin resistance during pregnancy. Endocrinology 2011, 152, 1355–1365. [Google Scholar] [CrossRef] [PubMed]
- Ma, C.; Gao, Q.; Zhang, W.; Azad, M.A.K.; Kong, X. Alterations in the Blood Parameters and Fecal Microbiota and Metabolites during Pregnant and Lactating Stages in Bama Mini Pigs as a Model. Mediat. Inflamm. 2020, 2020, 8829072. [Google Scholar] [CrossRef] [PubMed]
- Saleri, R.; Sabbioni, A.; Cavalli, V.; Superchi, P. Monitoring blood plasma leptin and lactogenic hormones in pregnant sows. Animal 2015, 9, 629–634. [Google Scholar] [CrossRef] [PubMed]
- Miller, H.M.; Foxcroft, G.R.; Aherne, F.X. Increasing feed intake in late gestation does not affect plasma progesterone concentration in the sow. Theriogenology 2004, 62, 1618–1626. [Google Scholar] [CrossRef] [PubMed]
- Li, Q.; Yang, S.; Chen, F.; Guan, W.; Zhang, S. Nutritional strategies to alleviate oxidative stress in sows. Anim. Nutr. 2022, 9, 60–73. [Google Scholar] [CrossRef]
- Phillippe, M. Telomeres, oxidative stress, and timing for spontaneous term and preterm labor. Am. J. Obs. Gynecol. 2022, 227, 148–162. [Google Scholar] [CrossRef]
- Zhao, Y.; Kim, S.W. Oxidative stress status and reproductive performance of sows during gestation and lactation under different thermal environments. Asian-Australas J. Anim. Sci. 2020, 33, 722–731. [Google Scholar] [CrossRef]
- Zhao, Y.; Flowers, W.L.; Saraiva, A.; Yeum, K.J.; Kim, S.W. Effect of social ranks and gestation housing systems on oxidative stress status, reproductive performance, and immune status of sows. J. Anim. Sci. 2013, 91, 5848–5858. [Google Scholar] [CrossRef]
- Ostrenko, K.; Nekrasov, R.; Ovcharova, A.; Lemiasheuski, V.; Kutin, I. The Effect of Lithium Salt with Ascorbic Acid on the Antioxidant Status and Productivity of Gestating Sows. Animals 2022, 12, 915. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Wei, H.; Ao, J.; Long, G.; Peng, J. Inclusion of Konjac Flour in the Gestation Diet Changes the Gut Microbiota, Alleviates Oxidative Stress, and Improves Insulin Sensitivity in Sows. Appl. Environ. Microbiol. 2016, 82, 5899–5909. [Google Scholar] [CrossRef] [PubMed]
- Tan, C.; Wei, H.; Sun, H.; Ao, J.; Long, G.; Jiang, S.; Peng, J. Effects of Dietary Supplementation of Oregano Essential Oil to Sows on Oxidative Stress Status, Lactation Feed Intake of Sows, and Piglet Performance. Biomed. Res. Int. 2015, 2015, 525218. [Google Scholar] [CrossRef] [PubMed]
- Berchieri-Ronchi, C.B.; Kim, S.W.; Zhao, Y.; Correa, C.R.; Yeum, K.J.; Ferreira, A.L. Oxidative stress status of highly prolific sows during gestation and lactation. Animal 2011, 5, 1774–1779. [Google Scholar] [CrossRef]
- Guo, G.; Zhou, T.; Ren, F.; Sun, J.; Deng, D.; Huang, X.; Wassie, T.; Qazi, I.H.; Wu, X. Effect of Maternal Catalase Supplementation on Reproductive Performance, Antioxidant Activity and Mineral Transport in Sows and Piglets. Animals 2022, 12, 828. [Google Scholar] [CrossRef]
- Hu, J.; Yan, P. Effects of Backfat Thickness on Oxidative Stress and Inflammation of Placenta in Large White Pigs. Vet. Sci. 2022, 9, 302. [Google Scholar] [CrossRef]
- Cheng, C.; Wu, X.; Zhang, X.; Zhang, X.; Peng, J. Obesity of Sows at Late Pregnancy Aggravates Metabolic Disorder of Perinatal Sows and Affects Performance and Intestinal Health of Piglets. Animals 2019, 10, 49. [Google Scholar] [CrossRef]
- Pawar, A.S.; Zhu, X.Y.; Eirin, A.; Tang, H.; Jordan, K.L.; Woollard, J.R.; Lerman, A.; Lerman, L.O. Adipose Tissue Remodeling in a Novel Domestic Porcine Model of Diet-Induced Obesity. Obesity 2015, 23, 399–407. [Google Scholar] [CrossRef]
- Fujimori, M.; Franca, E.L.; Morais, T.C.; Fiorin, V.; de Abreu, L.C.; Honorio-Franca, A.C. Cytokine and adipokine are biofactors can act in blood and colostrum of obese mothers. Biofactors 2017, 43, 243–250. [Google Scholar] [CrossRef]
- Jarvie, E.; Hauguel-de-Mouzon, S.; Nelson, S.M.; Sattar, N.; Catalano, P.M.; Freeman, D.J. Lipotoxicity in obese pregnancy and its potential role in adverse pregnancy outcome and obesity in the offspring. Clin. Sci. 2010, 119, 123–129. [Google Scholar] [CrossRef] [Green Version]
- Catalano, P.M. Obesity, insulin resistance, and pregnancy outcome. Reproduction 2010, 140, 365–371. [Google Scholar] [CrossRef] [PubMed]
- Tian, L.; Dong, S.S.; Hu, J.; Yao, J.J.; Yan, P.S. The effect of maternal obesity on fatty acid transporter expression and lipid metabolism in the full-term placenta of lean breed swine. J. Anim. Physiol. Anim. Nutr. 2018, 102, e242–e253. [Google Scholar] [CrossRef] [PubMed]
- Fowden, A.L.; Camm, E.J.; Sferruzzi-Perri, A.N. Effects of Maternal Obesity on Placental Phenotype. Curr. Vasc. Pharmacol. 2021, 19, 113–131. [Google Scholar] [CrossRef] [PubMed]
- Li, J.W.; Hu, J.; Wei, M.; Guo, Y.Y.; Yan, P.S. The Effects of Maternal Obesity on Porcine Placental Efficiency and Proteome. Animals 2019, 9, 546. [Google Scholar] [CrossRef] [PubMed]
- Hu, R.; Tan, J.; Li, Z.; Wang, L.; Shi, M.; Li, B.; Liu, M.; Yuan, X.; He, J.; Wu, X. Effect of dietary resveratrol on placental function and reproductive performance of late pregnancy sows. Front. Nutr. 2022, 9, 1001031. [Google Scholar] [CrossRef]
- Cristofolini, A.; Fiorimanti, M.; Campos, M.; Sanchis, E.; Diaz, T.; Moschetti, E.; Merkis, C. Morphometric study of the porcine placental vascularization. Reprod. Domest. Anim. 2018, 53, 217–225. [Google Scholar] [CrossRef]
- Leiser, R.; Kaufmann, P. Placental structure: In a comparative aspect. Exp. Clin. Endocrinol. 1994, 102, 122–134. [Google Scholar] [CrossRef]
- Bidarimath, M.; Tayade, C. Pregnancy and spontaneous fetal loss: A pig perspective. Mol. Reprod. Dev. 2017, 84, 856–869. [Google Scholar] [CrossRef]
- Bazer, F.W.; Johnson, G.A. Pig blastocyst-uterine interactions. Differentiation 2014, 87, 52–65. [Google Scholar] [CrossRef]
- Biensen, N.J.; Wilson, M.E.; Ford, S.P. The impact of either a Meishan or Yorkshire uterus on Meishan or Yorkshire fetal and placental development to days 70, 90, and 110 of gestation. J. Anim. Sci. 1998, 76, 2169–2176. [Google Scholar] [CrossRef] [Green Version]
- Vallet, J.L.; Miles, J.R.; Freking, B.A. Development of the pig placenta. Soc. Reprod. Fertil. Suppl. 2009, 66, 265–279. [Google Scholar] [CrossRef]
- Myatt, L.; Cui, X. Oxidative stress in the placenta. Histochem. Cell Biol. 2004, 122, 369–382. [Google Scholar] [CrossRef] [PubMed]
- Pereira, R.D.; De Long, N.E.; Wang, R.C.; Yazdi, F.T.; Holloway, A.C.; Raha, S. Angiogenesis in the placenta: The role of reactive oxygen species signaling. Biomed. Res. Int. 2015, 2015, 814543. [Google Scholar] [CrossRef] [PubMed]
- Jauniaux, E.; Poston, L.; Burton, G.J. Placental-related diseases of pregnancy: Involvement of oxidative stress and implications in human evolution. Hum. Reprod. Update 2006, 12, 747–755. [Google Scholar] [CrossRef] [PubMed]
- Watson, A.L.; Skepper, J.N.; Jauniaux, E.; Burton, G.J. Susceptibility of human placental syncytiotrophoblastic mitochondria to oxygen-mediated damage in relation to gestational age. J. Clin. Endocrinol. Metab. 1998, 83, 1697–1705. [Google Scholar] [CrossRef]
- Watson, A.L.; Palmer, M.E.; Jauniaux, E.; Burton, G.J. Variations in expression of copper/zinc superoxide dismutase in villous trophoblast of the human placenta with gestational age. Placenta 1997, 18, 295–299. [Google Scholar] [CrossRef]
- Watson, A.L.; Skepper, J.N.; Jauniaux, E.; Burton, G.J. Changes in concentration, localization and activity of catalase within the human placenta during early gestation. Placenta 1998, 19, 27–34. [Google Scholar] [CrossRef]
- Kampmann, U.; Knorr, S.; Fuglsang, J.; Ovesen, P. Determinants of Maternal Insulin Resistance during Pregnancy: An Updated Overview. J. Diabetes Res. 2019, 2019, 5320156. [Google Scholar] [CrossRef]
- Heerwagen, M.J.; Stewart, M.S.; de la Houssaye, B.A.; Janssen, R.C.; Friedman, J.E. Transgenic increase in N-3/n-6 Fatty Acid ratio reduces maternal obesity-associated inflammation and limits adverse developmental programming in mice. PLoS ONE 2013, 8, e67791. [Google Scholar] [CrossRef]
- Strakovsky, R.S.; Pan, Y.X. A decrease in DKK1, a WNT inhibitor, contributes to placental lipid accumulation in an obesity-prone rat model. Biol. Reprod. 2012, 86, 81. [Google Scholar] [CrossRef]
- Song, T.; Lu, J.; Deng, Z.; Xu, T.; Yang, Y.; Wei, H.; Li, S.; Jiang, S.; Peng, J. Maternal obesity aggravates the abnormality of porcine placenta by increasing N(6)-methyladenosine. Int. J. Obes. 2018, 42, 1812–1820. [Google Scholar] [CrossRef] [PubMed]
- Liang, T.; Jinglong, X.; Shusheng, D.; Aiyou, W. Maternal obesity stimulates lipotoxicity and up-regulates inflammatory signaling pathways in the full-term swine placenta. Anim. Sci. J. 2018, 89, 1310–1322. [Google Scholar] [CrossRef] [PubMed]
- Saben, J.; Lindsey, F.; Zhong, Y.; Thakali, K.; Badger, T.M.; Andres, A.; Gomez-Acevedo, H.; Shankar, K. Maternal obesity is associated with a lipotoxic placental environment. Placenta 2014, 35, 171–177. [Google Scholar] [CrossRef] [PubMed]
- Saben, J.; Zhong, Y.; Gomez-Acevedo, H.; Thakali, K.M.; Borengasser, S.J.; Andres, A.; Shankar, K. Early growth response protein-1 mediates lipotoxicity-associated placental inflammation: Role in maternal obesity. Am. J. Physiol. Endocrinol. Metab. 2013, 305, E1–E14. [Google Scholar] [CrossRef] [PubMed]
- Zhou, Y.; Xu, T.; Wu, Y.; Wei, H.; Peng, J. Oxidative Stress and Inflammation in Sows with Excess Backfat: Up-Regulated Cytokine Expression and Elevated Oxidative Stress Biomarkers in Placenta. Animals 2019, 9, 796. [Google Scholar] [CrossRef]
- Tian, L.; Liu, G.; Kang, Z.; Yan, P. Microtubule Affinity-Regulating Kinase 4 Promotes Oxidative Stress and Mitochondrial Dysfunction by Activating NF-kappaB and Inhibiting AMPK Pathways in Porcine Placental Trophoblasts. Biomedicines 2022, 10, 165. [Google Scholar] [CrossRef]
- Xie, C.; Wu, X.; Long, C.; Wang, Q.; Fan, Z.; Li, S.; Yin, Y. Chitosan oligosaccharide affects antioxidant defense capacity and placental amino acids transport of sows. BMC Vet. Res. 2016, 12, 243. [Google Scholar] [CrossRef]
- Myatt, L.; Kossenjans, W.; Sahay, R.; Eis, A.; Brockman, D. Oxidative stress causes vascular dysfunction in the placenta. J. Matern.-Fetal Med. 2000, 9, 79–82. [Google Scholar]
- Mele, J.; Muralimanoharan, S.; Maloyan, A.; Myatt, L. Impaired mitochondrial function in human placenta with increased maternal adiposity. Am. J. Physiol. Endocrinol. Metab. 2014, 307, E419–E425. [Google Scholar] [CrossRef]
- Hastie, R.; Lappas, M. The effect of pre-existing maternal obesity and diabetes on placental mitochondrial content and electron transport chain activity. Placenta 2014, 35, 673–683. [Google Scholar] [CrossRef]
- Prater, M.R.; Laudermilch, C.L.; Liang, C.; Holladay, S.D. Placental oxidative stress alters expression of murine osteogenic genes and impairs fetal skeletal formation. Placenta 2008, 29, 802–808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mutinati, M.; Piccinno, M.; Roncetti, M.; Campanile, D.; Rizzo, A.; Sciorsci, R. Oxidative stress during pregnancy in the sheep. Reprod. Domest. Anim. 2013, 48, 353–357. [Google Scholar] [CrossRef] [PubMed]
- Sato, T.; Takino, J.-I.; Nagamine, K.; Nishio, K.; Hori, T. RASGRP2 Suppresses Apoptosis via Inhibition of ROS Production in Vascular Endothelial Cells. Sci. World J. 2019, 2019, 4639165. [Google Scholar] [CrossRef] [PubMed]
- Gourvas, V.; Dalpa, E.; Konstantinidou, A.; Vrachnis, N.; Spandidos, D.A.; Sifakis, S. Angiogenic factors in placentas from pregnancies complicated by fetal growth restriction (review). Mol. Med. Rep. 2012, 6, 23–27. [Google Scholar] [CrossRef] [PubMed]
- Huang, S.; Wu, Z.; Huang, Z.; Hao, X.; Zhang, L.; Hu, C.; Wei, J.; Deng, J.; Tan, C. Maternal supply of cysteamine alleviates oxidative stress and enhances angiogenesis in porcine placenta. J. Anim. Sci. Biotechnol. 2021, 12, 91. [Google Scholar] [CrossRef]
- Dumolt, J.H.; Powell, T.L.; Jansson, T. Placental Function and the Development of Fetal Overgrowth and Fetal Growth Restriction. Obstet. Gynecol. Clin. North Am. 2021, 48, 247–266. [Google Scholar] [CrossRef]
- Aye, I.L.; Keelan, J.A. Placental ABC transporters, cellular toxicity and stress in pregnancy. Chem. Biol. Interact. 2013, 203, 456–466. [Google Scholar] [CrossRef]
- Hu, C.; Yang, Y.; Deng, M.; Yang, L.; Shu, G.; Jiang, Q.; Zhang, S.; Li, X.; Yin, Y.; Tan, C.; et al. Placentae for Low Birth Weight Piglets Are Vulnerable to Oxidative Stress, Mitochondrial Dysfunction, and Impaired Angiogenesis. Oxidative Med. Cell. Longev. 2020, 2020, 8715412. [Google Scholar] [CrossRef]
- Lappas, M.; Andrikopoulos, S.; Permezel, M. Hypoxanthine-xanthine oxidase down-regulates GLUT1 transcription via SIRT1 resulting in decreased glucose uptake in human placenta. J. Endocrinol. 2012, 213, 49–57. [Google Scholar] [CrossRef]
- Jia, X.; Cao, Y.; Ye, L.; Liu, X.; Huang, Y.; Yuan, X.; Lu, C.; Xu, J.; Zhu, H. Vitamin D stimulates placental L-type amino acid transporter 1 (LAT1) in preeclampsia. Sci. Rep. 2022, 12, 4651. [Google Scholar] [CrossRef] [PubMed]
- Glazier, J.D.; Cetin, I.; Perugino, G.; Ronzoni, S.; Grey, A.M.; Mahendran, D.; Marconi, A.M.; Pardi, G.; Sibley, C.P. Association between the activity of the system A amino acid transporter in the microvillous plasma membrane of the human placenta and severity of fetal compromise in intrauterine growth restriction. Pediatr. Res. 1997, 42, 514–519. [Google Scholar] [CrossRef] [PubMed]
- Teixeira, S.A.; Marques, D.B.D.; Costa, T.C.; Oliveira, H.C.; Costa, K.A.; Carrara, E.R.; da Silva, W.; Guimaraes, J.D.; Neves, M.M.; Ibelli, A.M.G.; et al. Transcription Landscape of the Early Developmental Biology in Pigs. Animals 2021, 11, 1443. [Google Scholar] [CrossRef] [PubMed]
- Ramos-Ibeas, P.; Sang, F.; Zhu, Q.; Tang, W.W.C.; Withey, S.; Klisch, D.; Wood, L.; Loose, M.; Surani, M.A.; Alberio, R. Pluripotency and X chromosome dynamics revealed in pig pre-gastrulating embryos by single cell analysis. Nat. Commun. 2019, 10, 500. [Google Scholar] [CrossRef] [PubMed]
- Che, L.; Yang, Z.; Xu, M.; Zhang, Z.; Liu, P.; Xu, S.; Che, L.; Lin, Y.; Fang, Z.; Feng, B.; et al. Dietary energy intake affects fetal survival and development during early and middle pregnancy in Large White and Meishan gilts. Anim. Nutr. (Zhongguo Xu Mu Shou Yi Xue Hui) 2015, 1, 152–159. [Google Scholar] [CrossRef] [PubMed]
- Cao, G.; Tao, F.; Xin, L.; Li, Z.; Zhou, X. Effects of maternal serine supplementation on high-fat diet-induced oxidative stress and epigenetic changes in promoters of glutathione synthesis-related genes in offspring. J. Funct. Foods 2018, 47, 316–324. [Google Scholar] [CrossRef]
- Pereira, A.C.; Martel, F. Oxidative stress in pregnancy and fertility pathologies. Cell Biol. Toxicol. 2014, 30, 301–312. [Google Scholar] [CrossRef]
- Hussain, T.; Murtaza, G.; Metwally, E.; Kalhoro, D.H.; Kalhoro, M.S.; Rahu, B.A.; Sahito, R.G.A.; Yin, Y.; Yang, H.S.; Chughtai, M.I.; et al. The Role of Oxidative Stress and Antioxidant Balance in Pregnancy. Mediat. Inflamm. 2021, 2021, 9962860. [Google Scholar] [CrossRef]
- Hernandez-Trejo, M.; Montoya-Estrada, A.; Torres-Ramos, Y.; Espejel-Nunez, A.; Guzman-Grenfell, A.; Morales-Hernandez, R.; Tolentino-Dolores, M.; Laresgoiti-Servitje, E. Oxidative stress biomarkers and their relationship with cytokine concentrations in overweight/obese pregnant women and their neonates. BMC Immunol. 2017, 18, 3. [Google Scholar] [CrossRef]
- Sharma, D.; Shastri, S.; Sharma, P. Intrauterine Growth Restriction: Antenatal and Postnatal Aspects. Clin. Med. Insights. Pediatr. 2016, 10, 67–83. [Google Scholar] [CrossRef]
- Biri, A.; Bozkurt, N.; Turp, A.; Kavutcu, M.; Himmetoglu, O.; Durak, I. Role of oxidative stress in intrauterine growth restriction. Gynecol. Obs. Investig. 2007, 64, 187–192. [Google Scholar] [CrossRef]
- Zhang, Q.Q.; Li, J.; Cao, M.; Li, Y.; Zhuo, Y.; Fang, Z.F.; Che, L.Q.; Xu, S.Y.; Feng, B.; Lin, Y.; et al. Dietary supplementation of Bacillus subtilis PB6 improves sow reproductive performance and reduces piglet birth intervals. Anim. Nutr. 2020, 6, 278–287. [Google Scholar] [CrossRef] [PubMed]


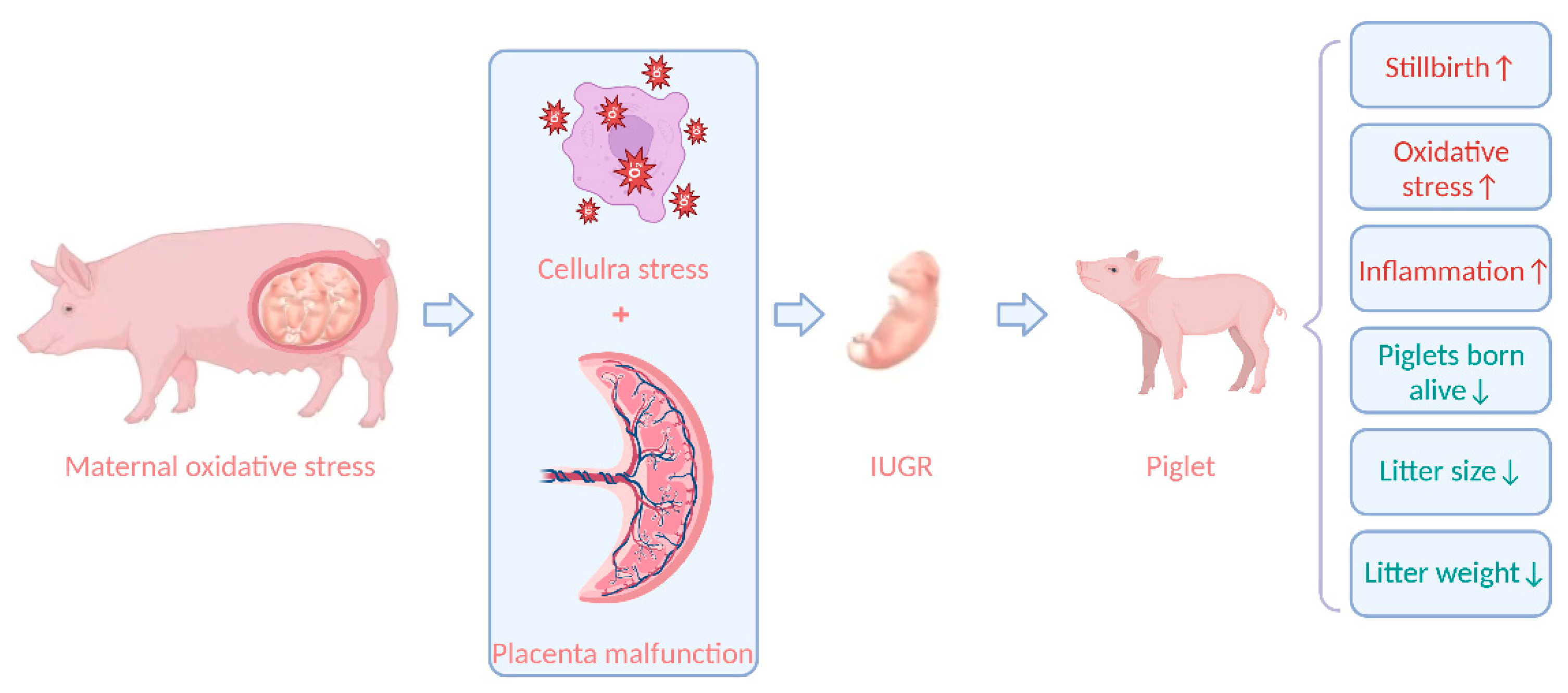{kind=link}
| Items | Early Pregnancy | The Second Trimester of Pregnancy | Late Pregnancy | References | ||
|---|---|---|---|---|---|---|
| Oxidative stress markers | Plasma | 8-OHdG | 0.71 ng/mL | 1.00 ng/mL | 1.07 ng/mL | Zhao et al. [49] |
| - | 0.72 ng/mL | 0.69–0.95 ng/mL | Zhao et al. [50] | |||
| MDA | - | 5.50 μmol/L | 5.79–5.67 μmol/L | Zhao et al. [50] | ||
| - | 6.35 nmol/mL | 7.20 nmol/ml | Ostrenko et al. [51] | |||
| Serum | ROS | About 1000 RLU | About 1000 RLU | About 3000 RLU | Tan et al. [52,53] | |
| TBARS | About 1.5 nmol/mL | About 1.5 nmol/mL | About 1 nmol/mL | Tan et al. [52] | ||
| 8-OHdG | About 21 ng/mL | About 35 ng/mL | About 40 ng/mL | Tan et al. [52] | ||
| Antioxidant capacity | Plasma | α-tocopherol | 7.14 μmol/L | 6.10 μmol/L | 3.07 μmol/L | Berchieri-Ronchi et al. [54] |
| Retinol | 1.10 μmol/L | 1.08 μmol/L | 0.57 μmol/L | Berchieri-Ronchi et al. [54] | ||
| GSH-Px | - | 2376 units | 2346 units | Ostrenko et al. [51] | ||
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yang, X.; Hu, R.; Shi, M.; Wang, L.; Yan, J.; Gong, J.; Zhang, Q.; He, J.; Wu, S. Placental Malfunction, Fetal Survival and Development Caused by Sow Metabolic Disorder: The Impact of Maternal Oxidative Stress. Antioxidants 2023, 12, 360. https://doi.org/10.3390/antiox12020360
Yang X, Hu R, Shi M, Wang L, Yan J, Gong J, Zhang Q, He J, Wu S. Placental Malfunction, Fetal Survival and Development Caused by Sow Metabolic Disorder: The Impact of Maternal Oxidative Stress. Antioxidants. 2023; 12(2):360. https://doi.org/10.3390/antiox12020360
Chicago/Turabian StyleYang, Xizi, Ruizhi Hu, Mingkun Shi, Long Wang, Jiahao Yan, Jiatai Gong, Qianjin Zhang, Jianhua He, and Shusong Wu. 2023. "Placental Malfunction, Fetal Survival and Development Caused by Sow Metabolic Disorder: The Impact of Maternal Oxidative Stress" Antioxidants 12, no. 2: 360. https://doi.org/10.3390/antiox12020360