

Vitamin B12 Ameliorates the Pathological Phenotypes of Multiple Parkinson’s Disease Models by Alleviating Oxidative Stress
 , and
, and 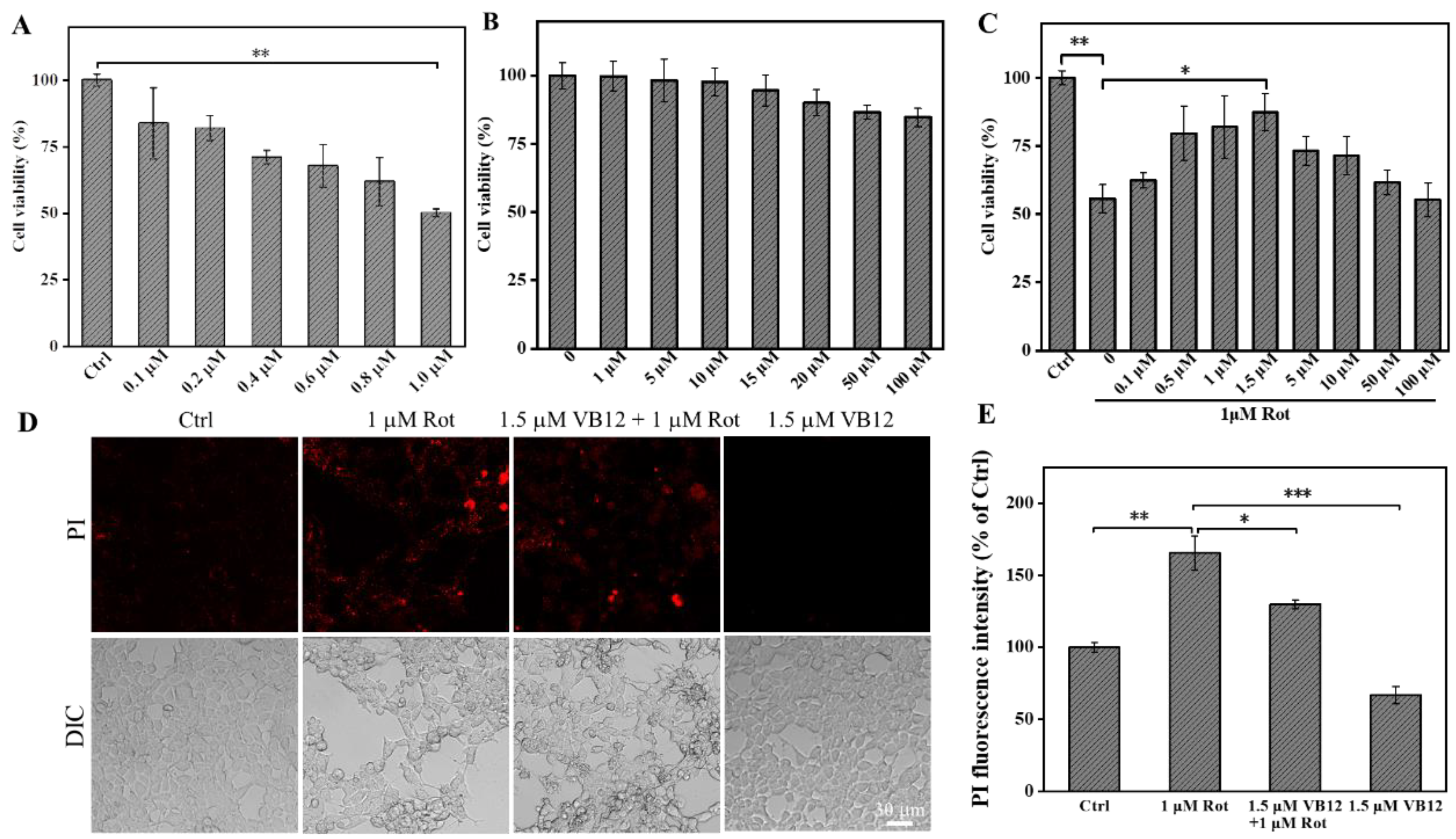{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Materials
2.2. Cell Culture and VB12 Treatments
2.3. Cell Viability Assays
2.4. Intracellular ROS Content, PI, SOD, GSH and ATP Assays
2.5. Western Blotting
2.6. Effects of VB12 on the Behavior of C. elegans PD Model
2.7. Effect of VB12 on the Mouse PD Model
2.8. Statistical Analysis
3. Results
3.1. Effect of VB12 on PD Cell Viability
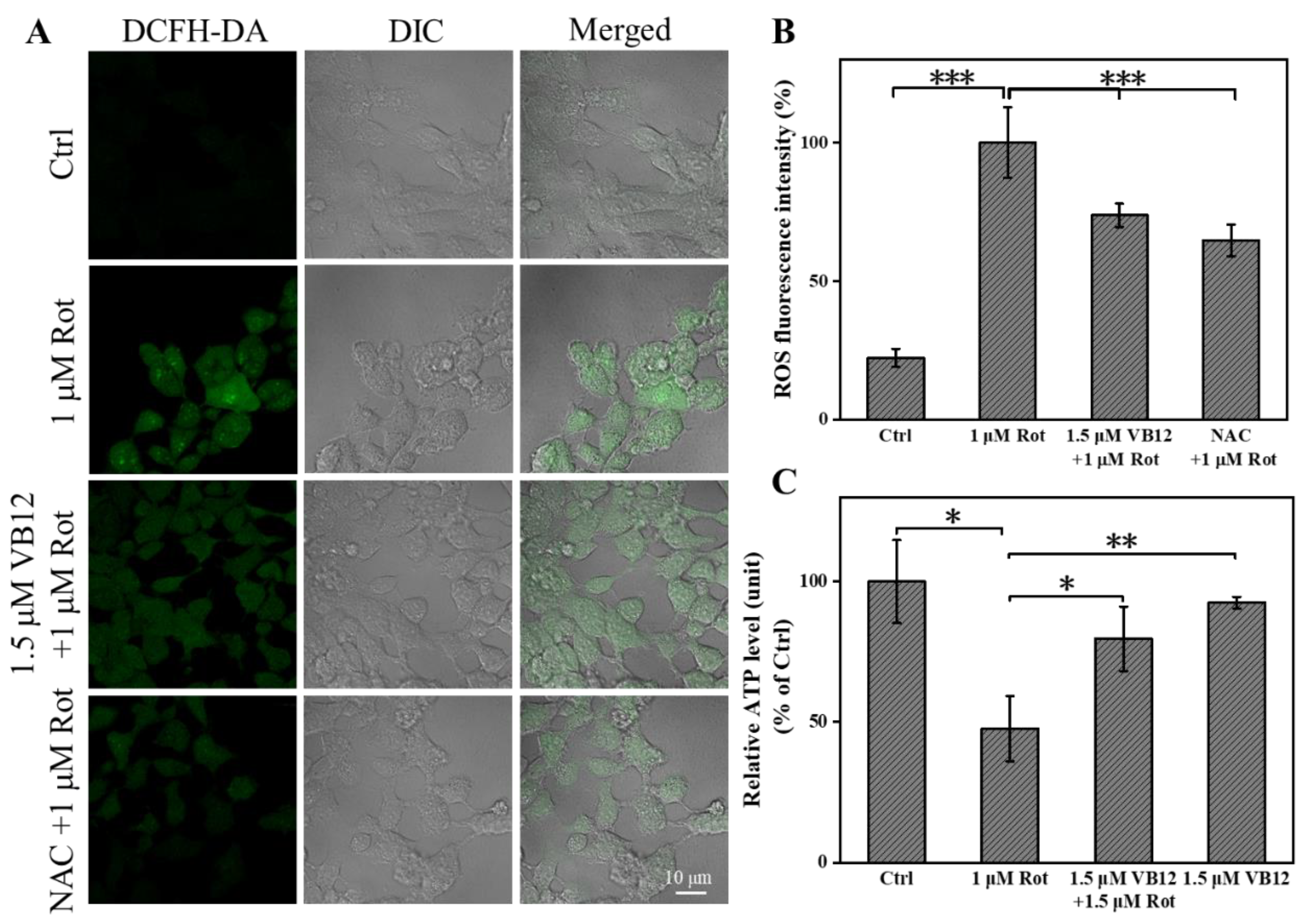
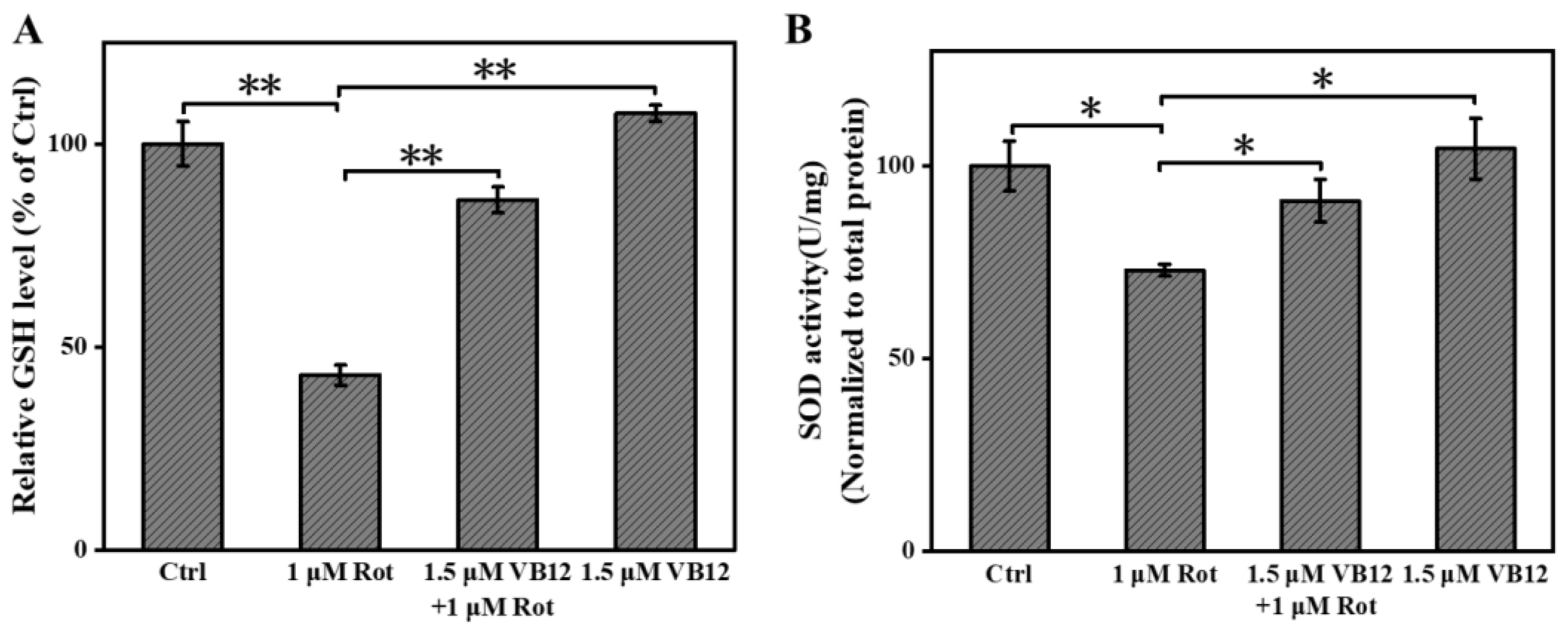
3.2. VB12 Relieves Oxidative Stress in the Rot-Induced Cellular PD Model
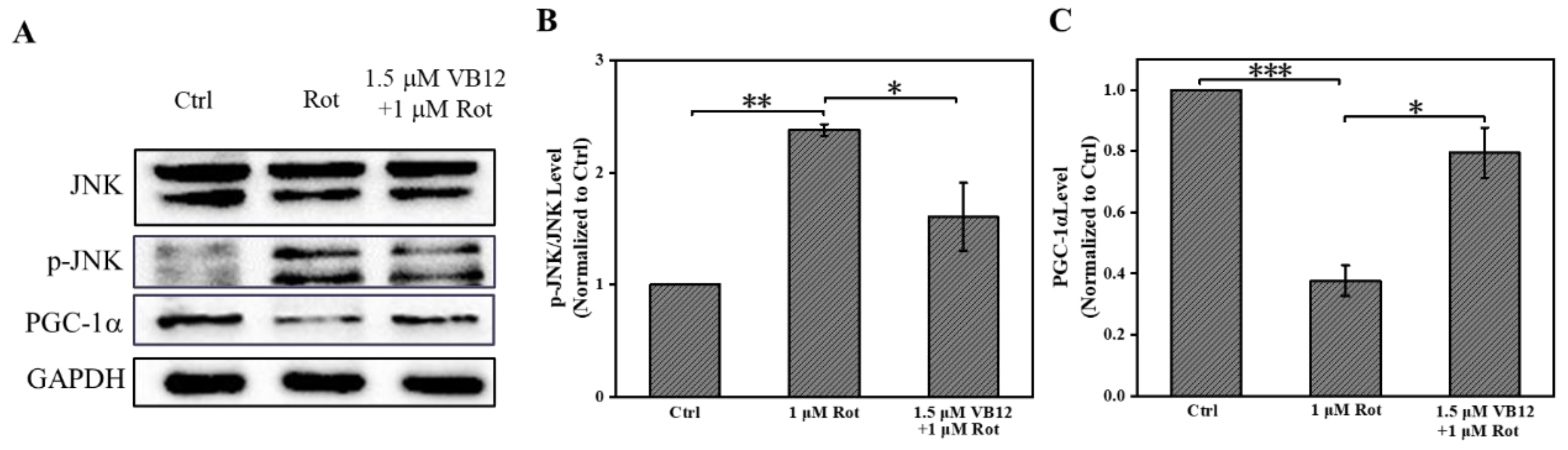
3.3. VB12 Affects Oxidative-Related Proteins in Rot-Treated SH-SY5Y Cells
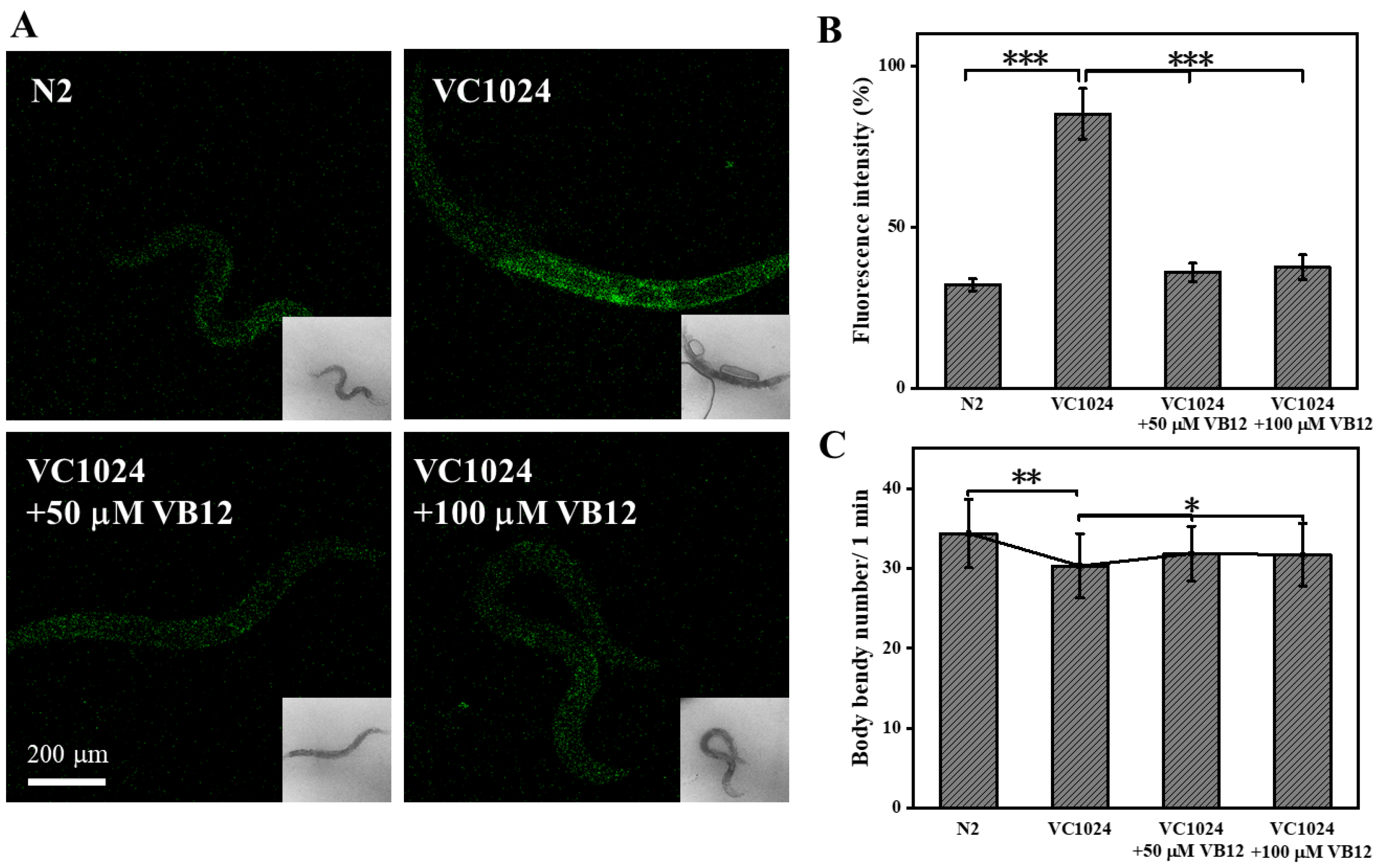
3.4. VB12 Reduces ROS and Ameliorates Movement Deficiency in the C. elegans PD Model
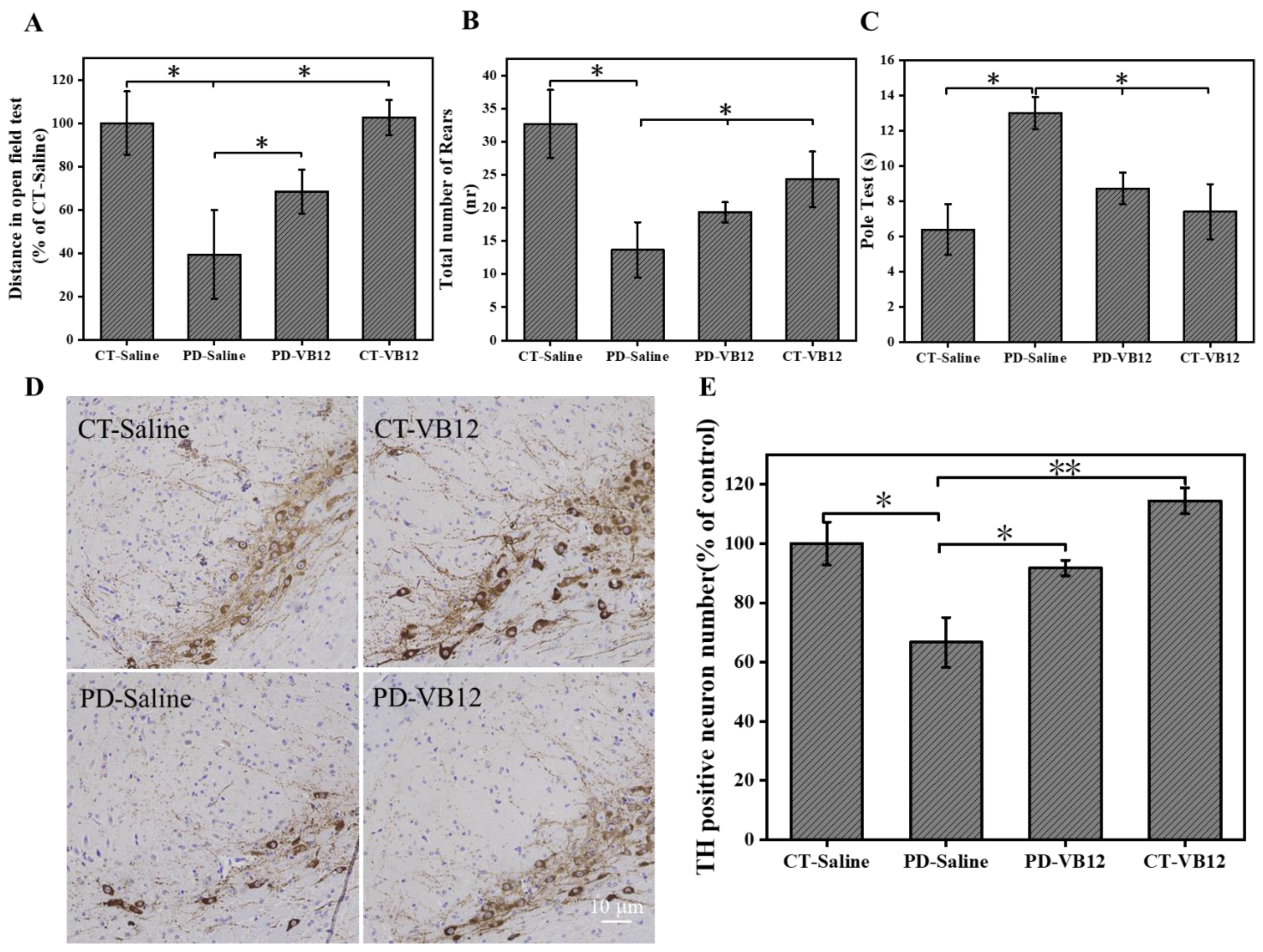
3.5. VB12 Ameliorates Movement Deficits and DA Neuron Loss in the MPTP-Induced PD Model
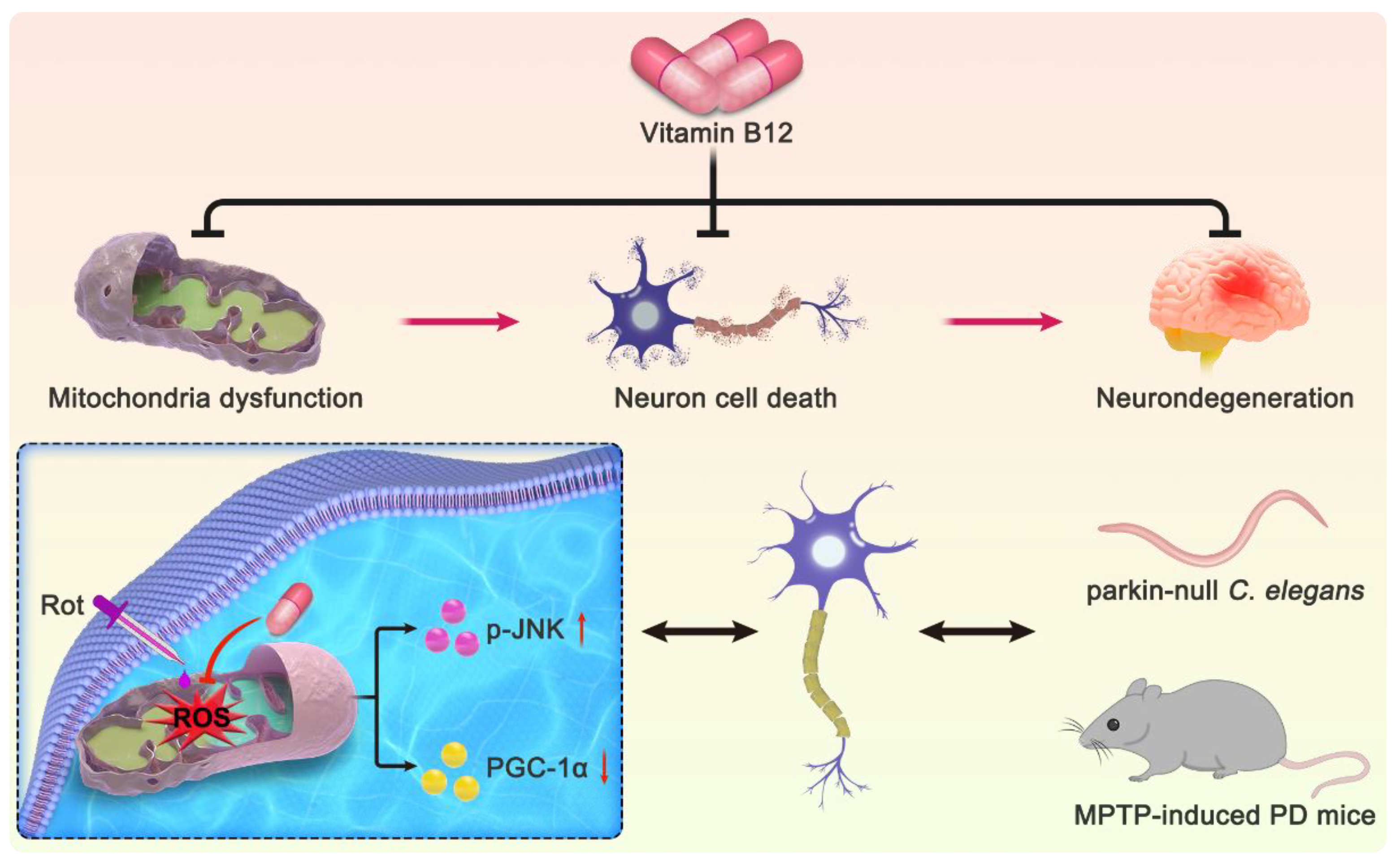
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Beitz, J.M. Parkinson’s disease: A review. Front. Biosci. (Sch. Ed.) 2014, 6, 65–74. [Google Scholar] [CrossRef]
- Ascherio, A.; Schwarzschild, M.A. The epidemiology of parkinson’s disease: Risk factors and prevention. Lancet Neurol. 2016, 15, 1257–1272. [Google Scholar]
- Raza, C.; Anjum, R.; Shakeel, N.U.A. Parkinson’s disease: Mechanisms, translational models and management strategies. Life Sci. 2019, 226, 77–90. [Google Scholar] [CrossRef]
- Reich, S.G.; Savitt, J.M. Parkinson’s Disease. Med. Clin. N. Am. 2019, 103, 337–350. [Google Scholar] [CrossRef]
- Jurcau, A. Insights into the pathogenesis of neurodegenerative diseases: Focus on mitochondrial dysfunction and oxidative stress. Int. J. Mol. Sci. 2021, 22, 11847. [Google Scholar] [CrossRef]
- Zhou, Z.D.; Xie, S.P.; Saw, W.T.; Ho, P.G.H.; Wang, H.; Lei, Z.; Yi, Z.; Tan, E.K. The therapeutic implications of tea polyphenols against dopamine (DA) neuron degeneration in parkinson’s Disease (PD). Cells 2019, 8, 911. [Google Scholar] [CrossRef] [Green Version]
- Marino, B.L.B.; de Souza, L.R.; Sousa, K.P.A.; Ferreira, J.V.; Padilha, E.C.; da Silva, C.H.T.P.; Taft, C.A.; Hage-Melim, L.I.S. Parkinson’s disease: A review from pathophysiology to treatment. Mini Rev. Med. Chem. 2020, 20, 754–767. [Google Scholar] [CrossRef]
- Puspita, L.; Chung, S.Y.; Shim, J.-W. Oxidative stress and cellular pathologies in Parkinson’s disease. Mol Brain 2017, 10, 53. [Google Scholar] [CrossRef] [Green Version]
- Trist, B.G.; Hare, D.J.; Double, K.L. Oxidative stress in the aging substantia nigra and the etiology of parkinson’s disease. Aging Cell 2019, 18, e13031. [Google Scholar] [CrossRef] [Green Version]
- Blesa, J.; Trigo-Damas, I.; Quiroga-Varela, A.; Jackson-Lewis, V.R. Oxidative stress and parkinson’s disease. Front. Neuroanat. 2015, 9, 91. [Google Scholar] [CrossRef] [Green Version]
- Dias, V.; Junn, E.; Mouradian, M.M. The role of oxidative stress in parkinson’s disease. J. Parkinson’s Dis. 2013, 3, 461–491. [Google Scholar] [CrossRef]
- Yang, S.; Lian, G. ROS and diseases: Role in metabolism and energy supply. Mol. Cell. Biochem. 2020, 467, 1–12. [Google Scholar] [CrossRef] [PubMed]
- Chia, S.J.; Tan, E.-K.; Chao, Y.-X. Historical perspective: Models of parkinson’s disease. Int. J. Mol. Sci. 2020, 21, 2464. [Google Scholar] [CrossRef] [Green Version]
- Bisbal, M.; Sanchez, M. Neurotoxicity of the pesticide Rot on neuronal polarization: A mechanistic approach. Neural Regen. Res. 2019, 14, 762–766. [Google Scholar] [CrossRef] [PubMed]
- Luo, R.; Zhu, L.; Zeng, Z.; Zhou, R.; Zhang, J.; Xiao, S.; Bi, W. Dl-butylphthalide inhibits Rot-induced oxidative stress in microglia via regulation of the Keap1/Nrf2/HO-1 signaling pathway. Exp. Ther. Med. 2021, 21, 597. [Google Scholar] [CrossRef]
- Inden, M.; Kitamura, Y.; Abe, M.; Tamaki, A.; Takata, K.; Taniguchi, T. Parkinsonian Rot mouse model: Reevaluation of long-term administration of Rot in C57BL/6 mice. Biol. Pharm. Bull. 2011, 34, 92–96. [Google Scholar] [CrossRef] [Green Version]
- AlShimemeri, S.; Di Luca, D.G.; Fox, S.H. MPTP parkinsonism and implications for understanding parkinson’s disease. Mov. Disord. Clin. Pract. 2022, 9, 42–47. [Google Scholar] [CrossRef]
- Hunt, J.; Coulson, E.J.; Rajnarayanan, R.; Oster, H.; Videnovic, A.; Rawashdeh, O. Sleep and circadian rhythms in parkinson’s disease and preclinical models. Mol. Neurodegener. 2022, 17, 2. [Google Scholar]
- Liang, Z.; Currais, A.; Soriano-Castell, D.; Schubert, D.; Maher, P. Natural products targeting mitochondria: Emerging therapeutics for age-associated neurological disorders. Pharmacol. Ther. 2021, 221, 107749. [Google Scholar]
- Wulansari, N.; Kim, E.-H.; Sulistio, Y.A.; Rhee, Y.-H.; Song, J.-J.; Lee, S.-H. Vitamin C-induced epigenetic modifications in donor NSCs establish midbrain marker expressions critical for cell-based therapy in parkinson’s disease. Stem Cell Rep. 2017, 9, 1192–1206. [Google Scholar] [CrossRef] [Green Version]
- Zhang, H.-J.; Zhang, J.-R.; Mao, C.-J.; Li, K.; Wang, F.; Chen, J.; Liu, C.-F. Relationship between 25-Hydroxyvitamin D, bone density, and parkinson’s disease symptoms. Acta Neurol. Scand. 2019, 140, 274–280. [Google Scholar] [CrossRef]
- Lee, P.; Ulatowski, L.M. Vitamin E: Mechanism of transport and regulation in the CNS. IUBMB Life 2019, 71, 424–429. [Google Scholar] [CrossRef]
- Zhao, X.; Zhang, M.; Li, C.; Jiang, X.; Su, Y.; Zhang, Y. Benefits of vitamins in the treatment of parkinson’s disease. Oxid. Med. Cell. Longev. 2019, 2019, 9426867. [Google Scholar] [CrossRef]
- Sun, Y.; Pham, A.N.; Waite, T.D. The effect of vitamin C and iron on dopamine-mediated free radical generation: Implications to parkinson’s disease. Dalton Trans. 2018, 47, 4059–4069. [Google Scholar] [CrossRef]
- Hinman, A.; Holst, C.R.; Latham, J.C.; Bruegger, J.J.; Ulas, G.; McCusker, K.P.; Amagata, A.; Davis, D.; Hoff, K.G.; Kahn-Kirby, A.H.; et al. Vitamin E hydroquinone is an endogenous regulator of ferroptosis via redox control of 15-lipoxygenase. PLoS ONE 2018, 13, e0201369. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abdalkader, M.; Lampinen, R.; Kanninen, K.M.; Malm, T.M.; Liddell, J.R. Targeting Nrf2 to suppress ferroptosis and mitochondrial dysfunction in neurodegeneration. Front. Neurosci. 2018, 12, 466. [Google Scholar] [CrossRef] [Green Version]
- Agarwal, P.; Wang, Y.; Buchman, A.S.; Holland, T.M.; Bennett, D.A.; Morris, M.C. Dietary antioxidants associated with slower progression of parkinsonian signs in older adults. Nutr. Neurosci. 2020, 25, 550–557. [Google Scholar] [CrossRef]
- Hribar, C.A.; Cobbold, P.H.; Church, F.C. Potential role of vitamin D in the elderly to resist COVID-19 and to slow progression of parkinson’s disease. Brain Sci. 2020, 10, 284. [Google Scholar] [CrossRef]
- Luthra, N.S.; Marcus, A.H.; Hills, N.K.; Christine, C.W. VB12 measurements across neurodegenerative disorders. J. Clin. Mov. Disord. 2020, 7, 3. [Google Scholar] [CrossRef]
- McCarter, S.J.; Stang, C.; Turcano, P.; Mielke, M.M.; Ali, F.; Bower, J.H.; Savica, R. Higher VB12 level at parkinson’s disease diagnosis is associated with lower risk of future dementia. Parkinsonism Relat. Disord. 2020, 73, 19–22. [Google Scholar] [CrossRef]
- Ozcan, O.; Cosar, A. Homocysteine and VB12 levels related to MRI white matter abnormalities in parkinson’s disease dementia. Neuro-Degener. Dis. 2013, 12, 164. [Google Scholar] [CrossRef] [PubMed]
- Madenci, G.; Bilen, S.; Arli, B.; Saka, M.; Ak, F. Serum iron, VB12 and folic acid levels in parkinson’s disease. Neurochem. Res. 2012, 37, 1436–1441. [Google Scholar] [CrossRef] [PubMed]
- Dong, B.; Wu, R. Plasma homocysteine, folate and vitamin B12 levels in parkinson’s disease in China: A meta-analysis. Clin. Neurol. Neurosurg. 2020, 188, 105587. [Google Scholar] [CrossRef]
- McCaddon, A. VB12 in neurology and ageing; clinical and genetic aspects. Biochimie 2013, 95, 1066–1076. [Google Scholar] [CrossRef] [PubMed]
- Schaffner, A.; Li, X.; Gomez-Llorente, Y.; Leandrou, E.; Memou, A.; Clemente, N.; Yao, C.; Afsari, F.; Zhi, L.; Pan, N.; et al. Vitamin B modulates parkinson’s disease LRRK2 kinase activity through allosteric regulation and confers neuroprotection. Cell Res. 2019, 29, 313–329. [Google Scholar] [CrossRef] [Green Version]
- Martínez, M.-A.; Rodríguez, J.-L.; Lopez-Torres, B.; Martínez, M.; Martínez-Larrañaga, M.-R.; Maximiliano, J.-E.; Anadón, A.; Ares, I. Use of human neuroblastoma SH-SY5Y cells to evaluate glyphosate-induced effects on oxidative stress, neuronal development and cell death signaling pathways. Environ. Int. 2020, 135, 105414. [Google Scholar] [CrossRef]
- Ramalingam, M.; Huh, Y.-J.; Lee, Y.-I. The impairments of α-Synuclein and mechanistic target of rapamycin in Rot-induced SH-SY5Y cells and mice model of parkinson’s disease. Front. Neurosci. 2019, 13, 1028. [Google Scholar] [CrossRef] [Green Version]
- Zou, J. Zhang, Y.; Sun, J.; Wang, X.; Tu, H.; Geng, S.; Liu, R.; Chen, Y.; Bi, Z. Deoxyelephantopin induces reactive oxygen species-mediated apoptosis and autophagy in human osteosarcoma cells. Cell. Physiol. Biochem. 2017, 42, 1812–1821. [Google Scholar] [CrossRef] [Green Version]
- Zhang, L.; Xu, M.; Ren, Q.; Liu, G.; Meng, S.; Xiahou, K.; Zhang, Y.; Jiang, N.; Zhou, W. Human induced pluripotent stem cell-derived neural cells from Alzheimer’s disease patients exhibited different susceptibility to oxidative stress. Stem. Cells Dev. 2020, 29, 1444–1456. [Google Scholar] [CrossRef]
- Liu, Y.; Fu, Y.; Hu, X.; Chen, S.; Miao, J.; Wang, Y.; Zhou, Y.; Zhang, Y. Caveolin-1 knockdown increases the therapeutic sensitivity of lung cancer to cisplatin-induced apoptosis by repressing parkin-related mitophagy and activating the ROCK1 pathway. J. Cell. Physiol. 2020, 235, 1197–1208. [Google Scholar] [CrossRef]
- He, H.; Wang, L.; Qiao, Y.; Yang, B.; Yin, D.; He, M. Epigallocatechin-3-gallate pretreatment alleviates doxorubicin-induced ferroptosis and cardiotoxicity by upregulating AMPKα2 and activating adaptive autophagy. Redox Biol. 2021, 48, 102185. [Google Scholar] [CrossRef] [PubMed]
- Jiang, X.J.; Wu, Y.Q.; Ma, R.; Chang, Y.M.; Li, L.L.; Zhu, J.H.; Liu, G.P.; Li, G. PINK1 alleviates cognitive impairments via attenuating pathological tau aggregation in a mouse model of tauopathy. Front. Cell. Dev. Biol. 2022, 9, 736267. [Google Scholar] [CrossRef] [PubMed]
- Zhao, Z.; Ning, J.; Bao, X.-Q.; Shang, M.; Ma, J.; Li, G.; Zhang, D. Fecal microbiota transplantation protects rotenone-induced parkinson’s disease mice by suppressing inflammation mediated by the lipopolysaccharide-TLR4 signaling pathway through the microbiota-gut-brain axis. Microbiome 2021, 9, 226. [Google Scholar] [CrossRef] [PubMed]
- Jiang, P.-E.; Lang, Q.-H.; Yu, Q.-Y.; Tang, X.-Y.; Liu, Q.-Q.; Li, X.-Y.; Feng, X.-Z. Behavioral assessments of spontaneous locomotion in a murine MPTP-induced parkinson’s disease model. J. Vis. Exp. 2019, 143, e58653. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bai, H.; Ding, Y.; Li, X.; Kong, D.; Xin, C.; Yang, X.; Zhang, C.; Rong, Z.; Yao, C.; Lu, S.; et al. Polydatin protects SH-SY5Y in models of parkinson’s disease by promoting Atg5-mediated but parkin-independent autophagy. Neurochem. Int. 2020, 134, 104671. [Google Scholar] [CrossRef]
- Hadipour, E.; Fereidoni, M.; Tayarani-Najaran, Z. Betanin Attenuates oxidative stress induced by 6-OHDA in PC12 cells via SAPK/JNK and PI3 K pathways. Neurochem. Res. 2020, 45, 395–403. [Google Scholar] [CrossRef]
- Sun, K.; Jing, X.; Guo, J.; Yao, X.; Guo, F. Mitophagy in degenerative joint diseases. Autophagy 2021, 17, 2082–2092. [Google Scholar] [CrossRef]
- Zhang, Y.; Li, H.; Jin, S.; Lu, Y.; Peng, Y.; Zhao, L.; Wang, X. Cannabidiol protects against Alzheimer’s disease in C. elegans via ROS scavenging activity of its phenolic hydroxyl groups. Eur. J. Pharmacol. 2022, 919, 174829. [Google Scholar] [CrossRef]
- Thirugnanam, T.; Santhakumar, K. Chemically induced models of parkinson’s disease. Comp. Biochem. Physiol. C Toxicol. Pharm. 2022, 252, 109213. [Google Scholar] [CrossRef]
- Jankovic, J.; Tan, E.K. Parkinson’s disease: Etiopathogenesis and treatment. J. Neurol. Neurosurg. Psychiatry 2020, 91, 795–808. [Google Scholar] [CrossRef]
- Collier, T.J.; Kanaan, N.M.; Kordower, J.H. Aging and parkinson’s disease: Different sides of the same coin? Mov. Disord. Off. J. Mov. Disord. Soc. 2017, 32, 983–990. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhang, K.; Huang, Q.; Deng, S.; Yang, Y.; Li, J.; Wang, S. Mechanisms of TLR4-mediated autophagy and nitroxidative Stress. Front. Cell. Infect. Microbiol. 2021, 11, 766590. [Google Scholar] [CrossRef] [PubMed]
- Sies, H. Oxidative stress: A concept in redox biology and medicine. Redox Biol. 2015, 4, 180–183. [Google Scholar] [CrossRef] [PubMed]
- Pissadaki, E.K.; Bolam, J.P. The energy cost of action potential propagation in dopamine neurons: Clues to susceptibility in parkinson’s disease. Front. Comput. Neurosci. 2013, 7, 13. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Elkamhawy, A.; Woo, J.; Gouda, N.A.; Kim, J.; Nada, H.; Roh, E.J.; Park, K.D.; Cho, J.; Lee, K. Melatonin analogues potently inhibit MAO-B and protect PC12 cells against oxidative Stress. Antioxidants 2021, 10, 1604. [Google Scholar] [CrossRef]
- Kim, H.D.; Lee, J.Y.; Park, J.-Y.; Kim, D.H.; Kang, M.H.; Seong, H.-A.; Seo, K.H.; Ji, Y.-J. Neuroprotective effects of flower extract against oxidative stress-induced apoptosis in neuronal cells and mice. Antioxidants 2021, 10, 951. [Google Scholar] [CrossRef]
- Wang, T.; Li, C.; Han, B.; Wang, Z.; Meng, X.; Zhang, L.; He, J.; Fu, F. Neuroprotective effects of danshensu on rotenone-induced parkinson’s disease models in vitro and in vivo. BMC Complement Med. 2020, 20, 20. [Google Scholar] [CrossRef] [Green Version]
- Asanuma, M.; Miyazaki, I. Antioxidants in astrocytes as target of neuroprotection for parkinson’s disease. Nihon Yakurigaku Zasshi 2021, 156, 14–20. [Google Scholar] [CrossRef]
- Dong, W.; Luo, B.; Qiu, C.; Jiang, X.; Shen, B.; Zhang, L.; Liu, W.; Zhang, W. TRIM3 attenuates apoptosis in parkinson’s disease by activating PI3K/AKT signal pathway. Aging 2020, 13, 735–749. [Google Scholar] [CrossRef]
- Malko, P.; Ding, R.; Jiang, L.-H. TRPM2 channel in oxidative stress-induced mitochondrial dysfunction and apoptotic cell death. Adv. Protein Chem. Struct. Biol. 2021, 125, 51–72. [Google Scholar]
- Stern, M.; McNew, J.A. A transition to degeneration triggered by oxidative stress in degenerative disorders. Mol. Psychiatry 2021, 26, 736–746. [Google Scholar] [CrossRef] [PubMed]
- Ahmed, T.; Zulfiqar, A.; Arguelles, S.; Rasekhian, M.; Nabavi, S.F.; Silva, A.S.; Nabavi, S.M. Map kinase signaling as therapeutic target for neurodegeneration. Pharmacol. Res. 2020, 160, 105090. [Google Scholar] [CrossRef] [PubMed]
- Muhammad, T.; Ikram, M.; Ullah, R.; Rehman, S.U.; Kim, M.O. Hesperetin, a citrus flavonoid, attenuates LPS-induced neuroinflammation, apoptosis and memory impairments by modulating TLR4/NF-κB signaling. Nutrients 2019, 11, 648. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Song, Z.; Zhang, Y.; Zhang, H.; Rajendran, R.S.; Wang, R.; Hsiao, C.-D.; Li, J.; Xia, Q.; Liu, K. Isoliquiritigenin triggers developmental toxicity and oxidative stress-mediated apoptosis in zebrafish embryos/larvae via Nrf2-HO1/JNK-ERK/mitochondrion pathway. Chemosphere 2020, 246, 125727. [Google Scholar] [CrossRef]
- Caldwell, K.A.; Willicott, C.W.; Caldwell, G.A. Modelling neurodegeneration in Caenorhabditis elegans. Dis. Model Mech. 2020, 13, dmm046110. [Google Scholar] [CrossRef]
- He, C.-L.; Tang, Y.; Wu, J.-M.; Long, T.; Yu, L.; Teng, J.-F.; Qiu, W.-Q.; Pan, R.; Yu, C.-L.; Qin, D.-L.; et al. Chlorogenic acid delays the progression of parkinson’s disease via autophagy induction in Caenorhabditis elegans. Nutr. Neurosci. 2021, 19, 1–14. [Google Scholar] [CrossRef]
- Sharma, S.; Trivedi, S.; Pandey, T.; Ranjan, S.; Trivedi, M.; Pandey, R. Wedelolactone Mitigates parkinsonism via alleviating oxidative stress and mitochondrial dysfunction through NRF2/SKN-1. Mol. Neurobiol. 2021, 58, 65–77. [Google Scholar] [CrossRef]
- Chang, C.-H.; Wei, C.-C.; Ho, C.-T.; Liao, V.H.-C. N-γ-(L-glutamyl)-L-selenomethionine shows neuroprotective effects against parkinson’s disease associated with SKN-1/Nrf2 and TRXR-1 in Caenorhabditis elegans. Phytomedicine 2021, 92, 153733. [Google Scholar] [CrossRef]
- Rui, W.; Li, S.; Xiao, H.; Xiao, M.; Shi, J. Baicalein attenuates neuroinflammation by inhibiting NLRP3/caspase-1/GSDMD pathway in MPTP induced mice model of parkinson’s disease. Int. J. Neuropsychopharmacol. 2020, 23, 762–773. [Google Scholar] [CrossRef]
- Fan, Z.; Liang, Z.; Yang, H.; Pan, Y.; Zheng, Y.; Wang, X. Tenuigenin protects dopaminergic neurons from inflammation by suppressing NLRP3 inflammasome activation in microglia. J. Neuroinflamm. 2017, 14, 256. [Google Scholar] [CrossRef] [Green Version]
- Wang, K.; Lu, C.; Wang, T.; Qiao, C.; Lu, L.; Wu, D.; Lu, M.; Chen, R.; Fan, L.; Tang, J. Hyperoside suppresses NLRP3 inflammasome in parkinson’s disease via pituitary adenylate cyclase-activating polypeptide. Neurochem. Int. 2022, 152, 105254. [Google Scholar] [CrossRef] [PubMed]
- Gao, M.-R.; Wang, M.; Jia, Y.-Y.; Tian, D.-D.; Liu, A.; Wang, W.-J.; Yang, L.; Chen, J.-Y.; Yang, Q.; Liu, R.; et al. Echinacoside protects dopaminergic neurons by inhibiting NLRP3/Caspase-1/IL-1β signaling pathway in MPTP-induced parkinson’s disease model. Brain Res. Bull. 2020, 164, 55–64. [Google Scholar] [CrossRef] [PubMed]
- Roth, W. Mohamadzadeh, M. Vitamin B12 and gut-brain homeostasis in the pathophysiology of ischemic stroke. EBioMedicine 2021, 73, 103676. [Google Scholar] [CrossRef] [PubMed]
- Christine, C.W.; Auinger, P.; Saleh, N.; Tian, M.; Bottiglieri, T.; Arning, E.; Tran, N.K.; Ueland, P.M.; Green, R. Relationship of cerebrospinal fluid vitamin B12 status markers with parkinson’s disease progression. Mov. Disord. 2020, 35, 1466–1471. [Google Scholar] [CrossRef]







Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Wu, Y.; Zhao, Z.; Yang, N.; Xin, C.; Li, Z.; Xu, J.; Ma, B.; Lim, K.-L.; Li, L.; Wu, Q.; et al. Vitamin B12 Ameliorates the Pathological Phenotypes of Multiple Parkinson’s Disease Models by Alleviating Oxidative Stress. Antioxidants 2023, 12, 153. https://doi.org/10.3390/antiox12010153
Wu Y, Zhao Z, Yang N, Xin C, Li Z, Xu J, Ma B, Lim K-L, Li L, Wu Q, et al. Vitamin B12 Ameliorates the Pathological Phenotypes of Multiple Parkinson’s Disease Models by Alleviating Oxidative Stress. Antioxidants. 2023; 12(1):153. https://doi.org/10.3390/antiox12010153
Chicago/Turabian StyleWu, Yue, Zhongting Zhao, Naidi Yang, Chenqi Xin, Zheng Li, Jiajia Xu, Bo Ma, Kah-Leong Lim, Lin Li, Qiong Wu, and et al. 2023. "Vitamin B12 Ameliorates the Pathological Phenotypes of Multiple Parkinson’s Disease Models by Alleviating Oxidative Stress" Antioxidants 12, no. 1: 153. https://doi.org/10.3390/antiox12010153