
Impulse Noise Induced Hidden Hearing Loss, Hair Cell Ciliary Changes and Oxidative Stress in Mice

 , , , , , and
, , , , , and 
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
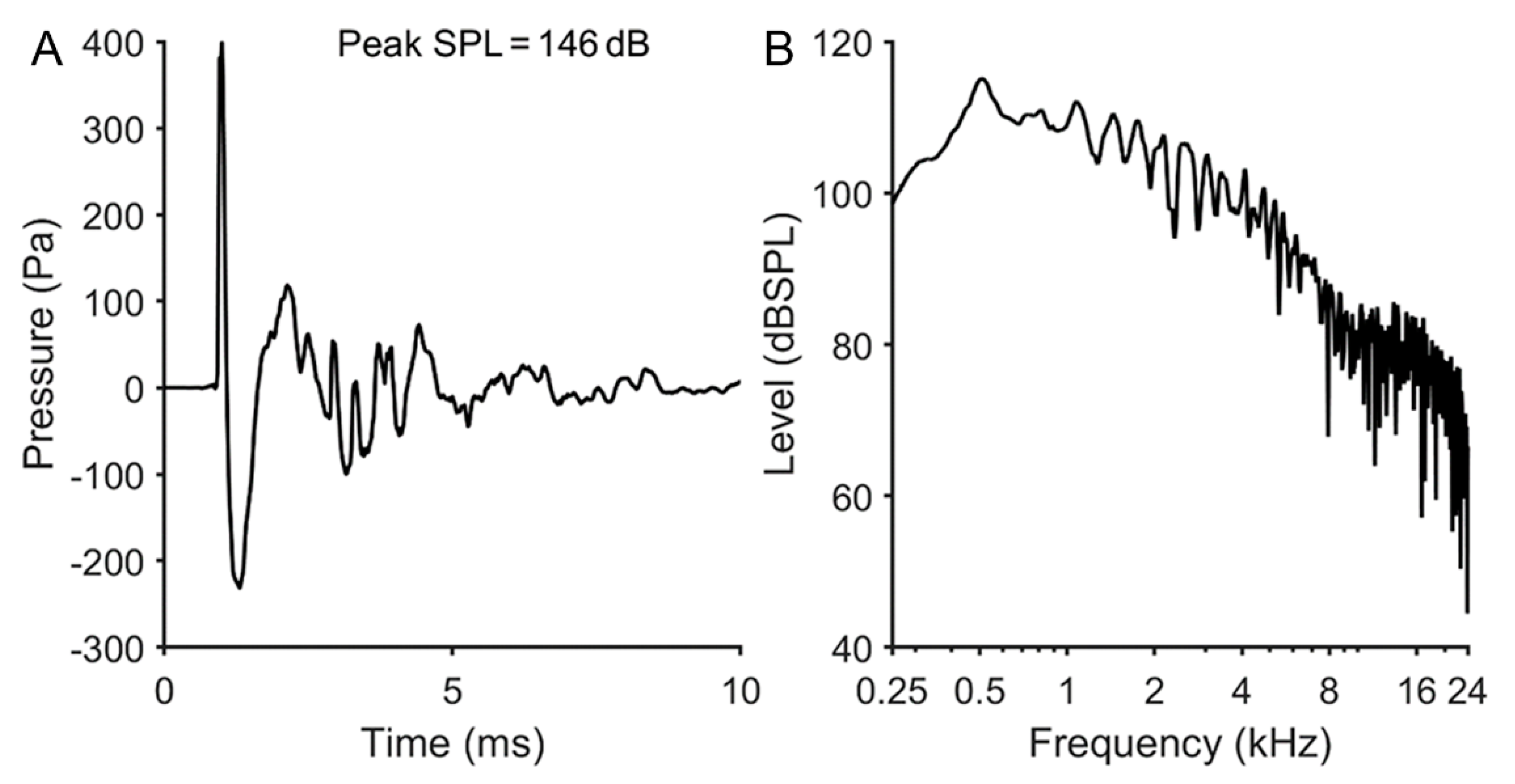
2.2. Impulse Exposures
2.3. Functional Hearing Assessments
2.3.1. Distortion-Product Otoacoustic Emission (DPOAEs)
2.3.2. Auditory Brainstem Response (ABRs)
2.4. Morphological Assessments
2.4.1. Counting of Sensory Hair Cells
2.4.2. Ultrastructural Analysis
2.4.3. Counting of the Auditory Nerve-Fiber Terminals
2.4.4. Counting of Ribbon Synapses of IHCs
2.5. Molecular Assessment
2.5.1. Enzymatic Activities and Lipid Peroxidation
2.5.2. Immunocytochemistry
2.6. Statistics
3. Results
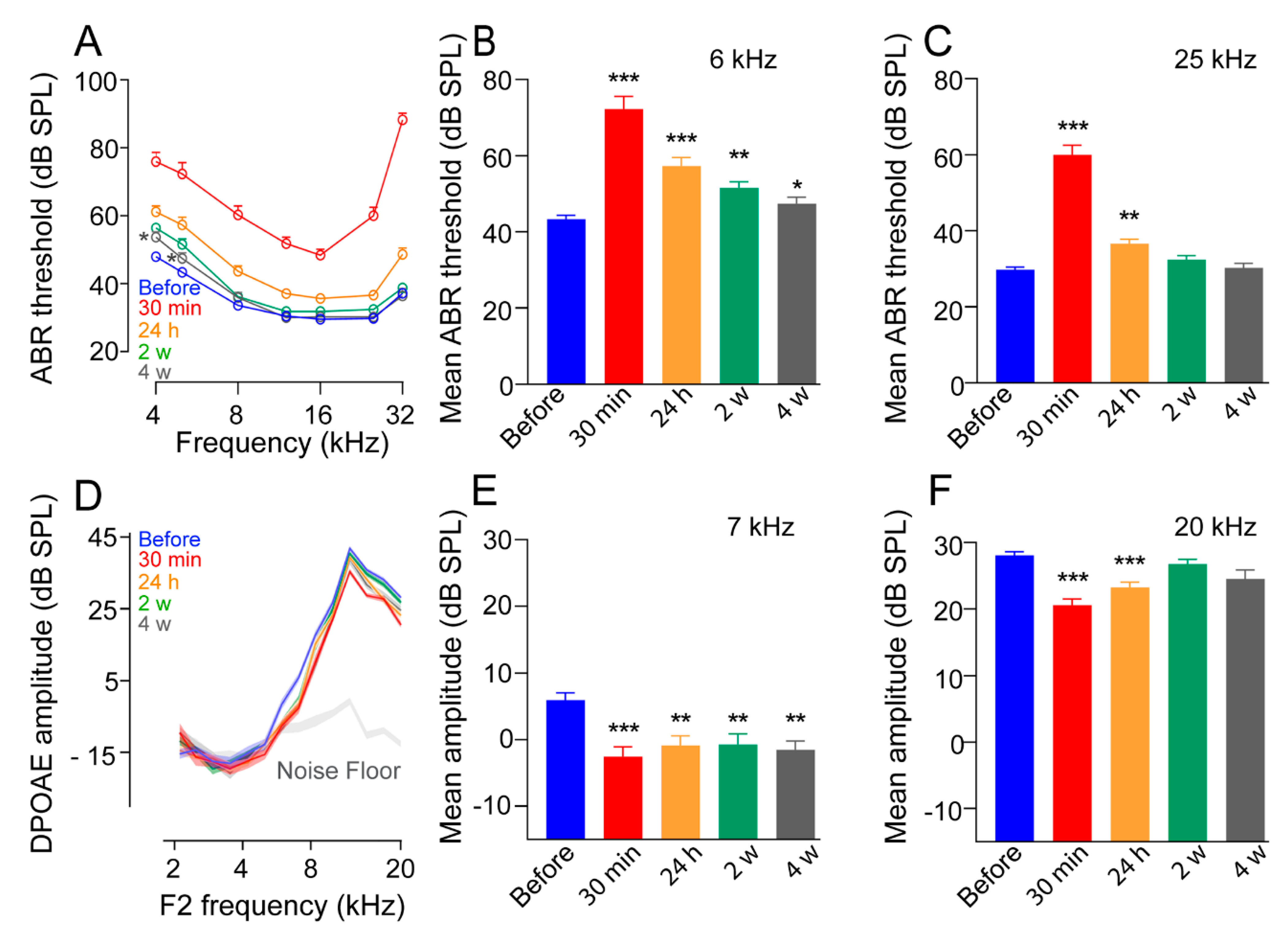
3.1. Impulse Noise Induced Reversible Threshold Shifts at the Higher Frequencies
3.2. Alteration of Distortion-Product Otoacoustic Emissions
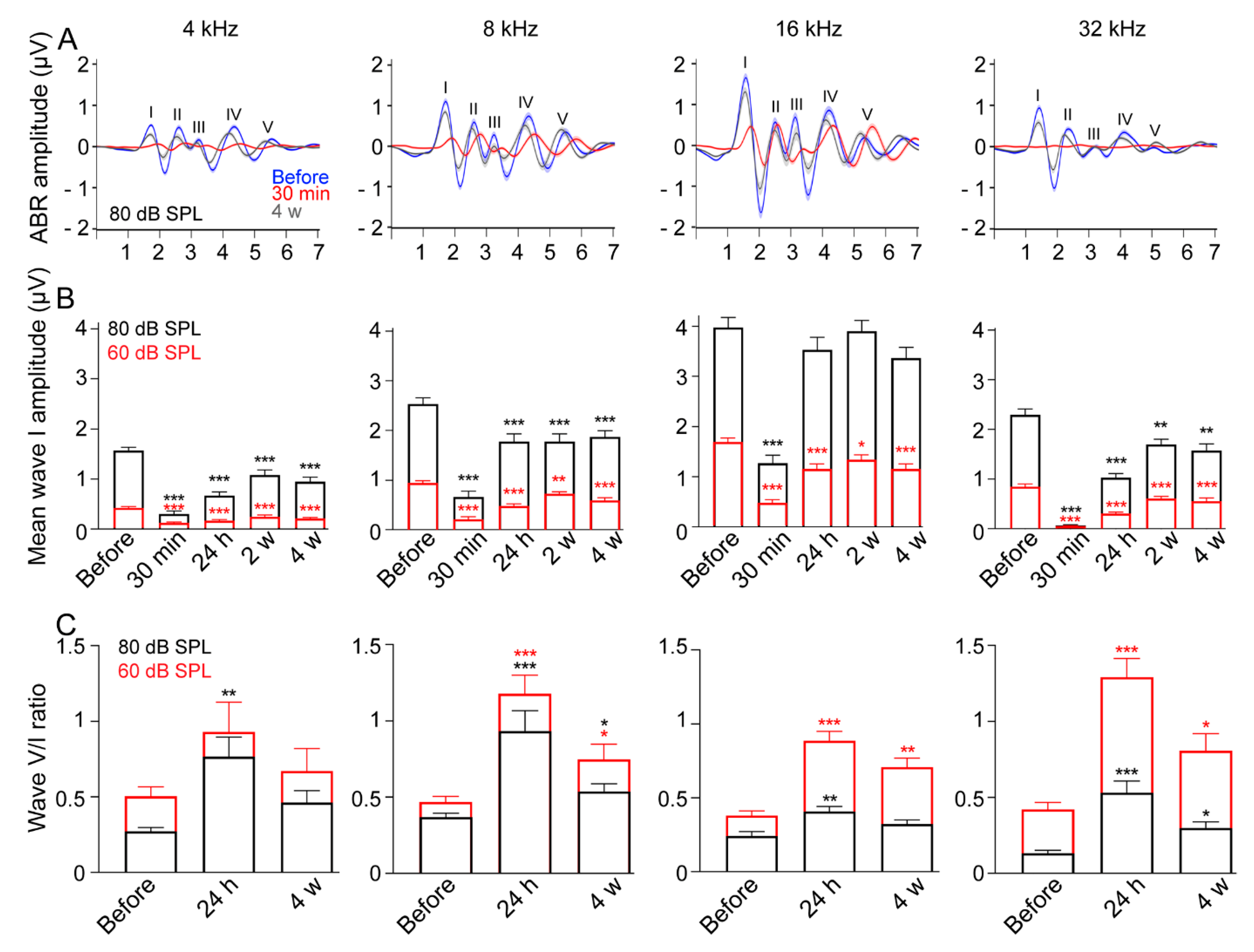
3.3. Reduced ABR Wave-I Amplitude and Elevated Central Gain following Exposure
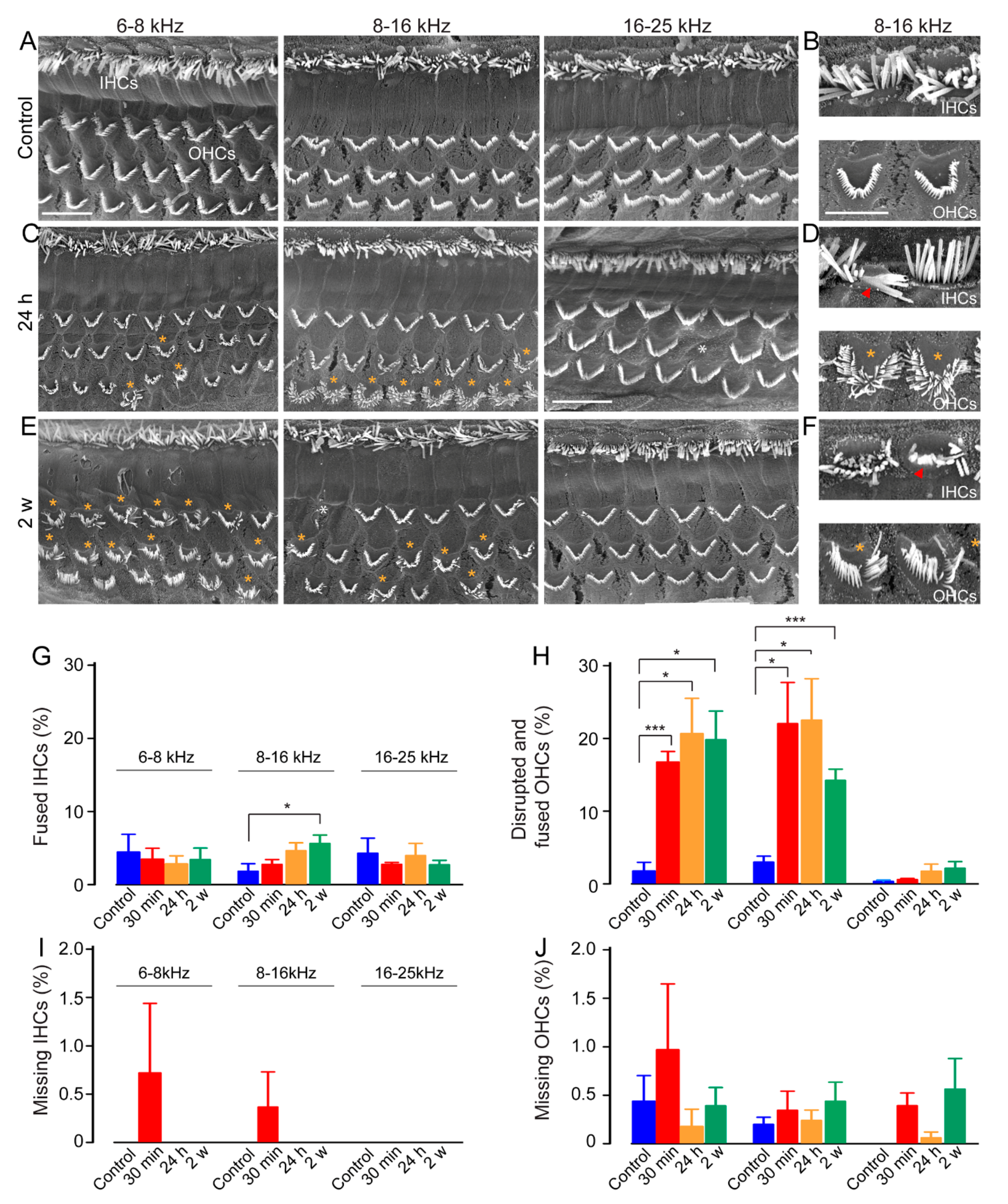
3.4. Disturbances in Stereociliary Bundle Morphology of the Outer Hair Cells
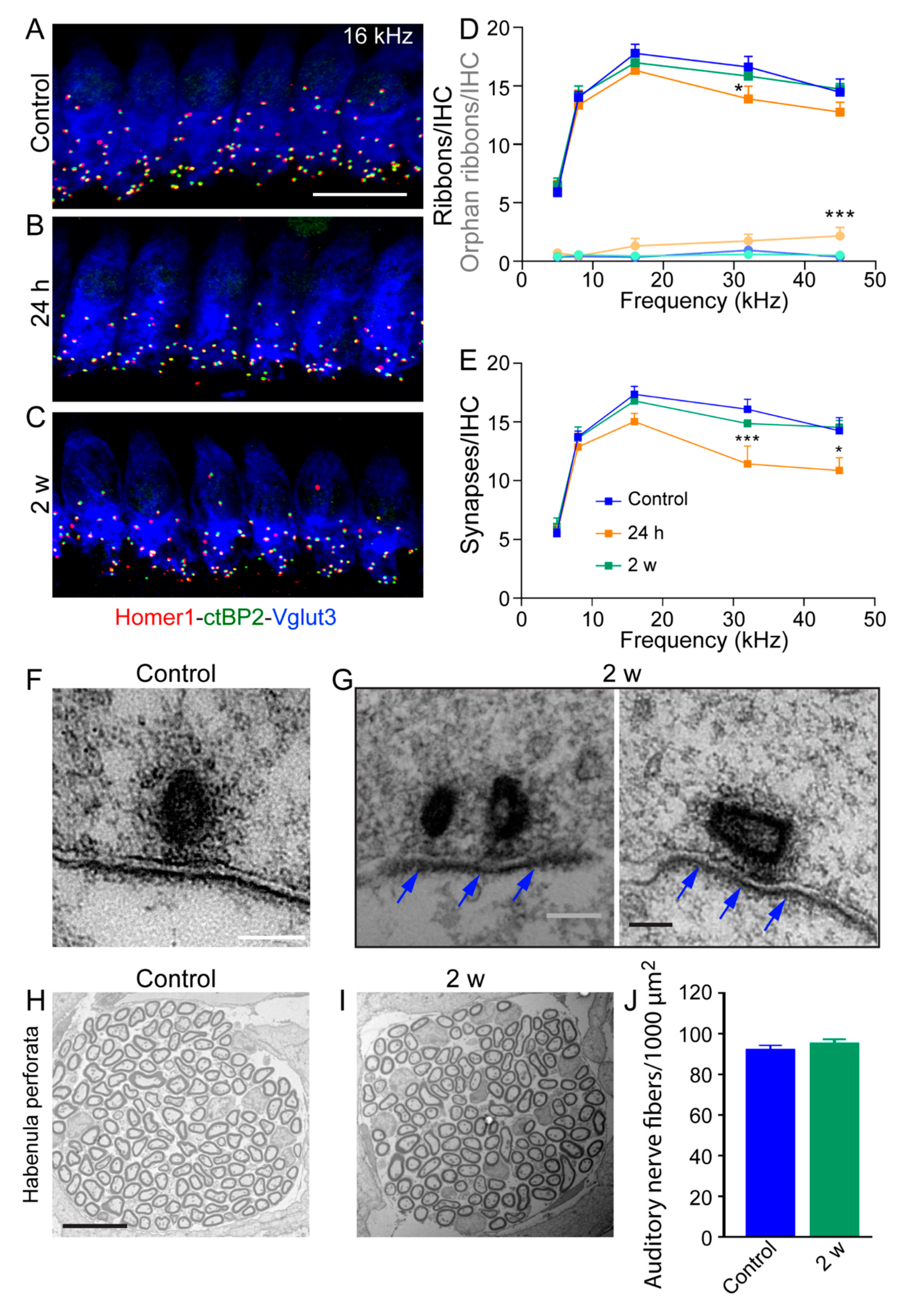
3.5. Reversible and Moderate Loss of IHC Ribbon Synapses
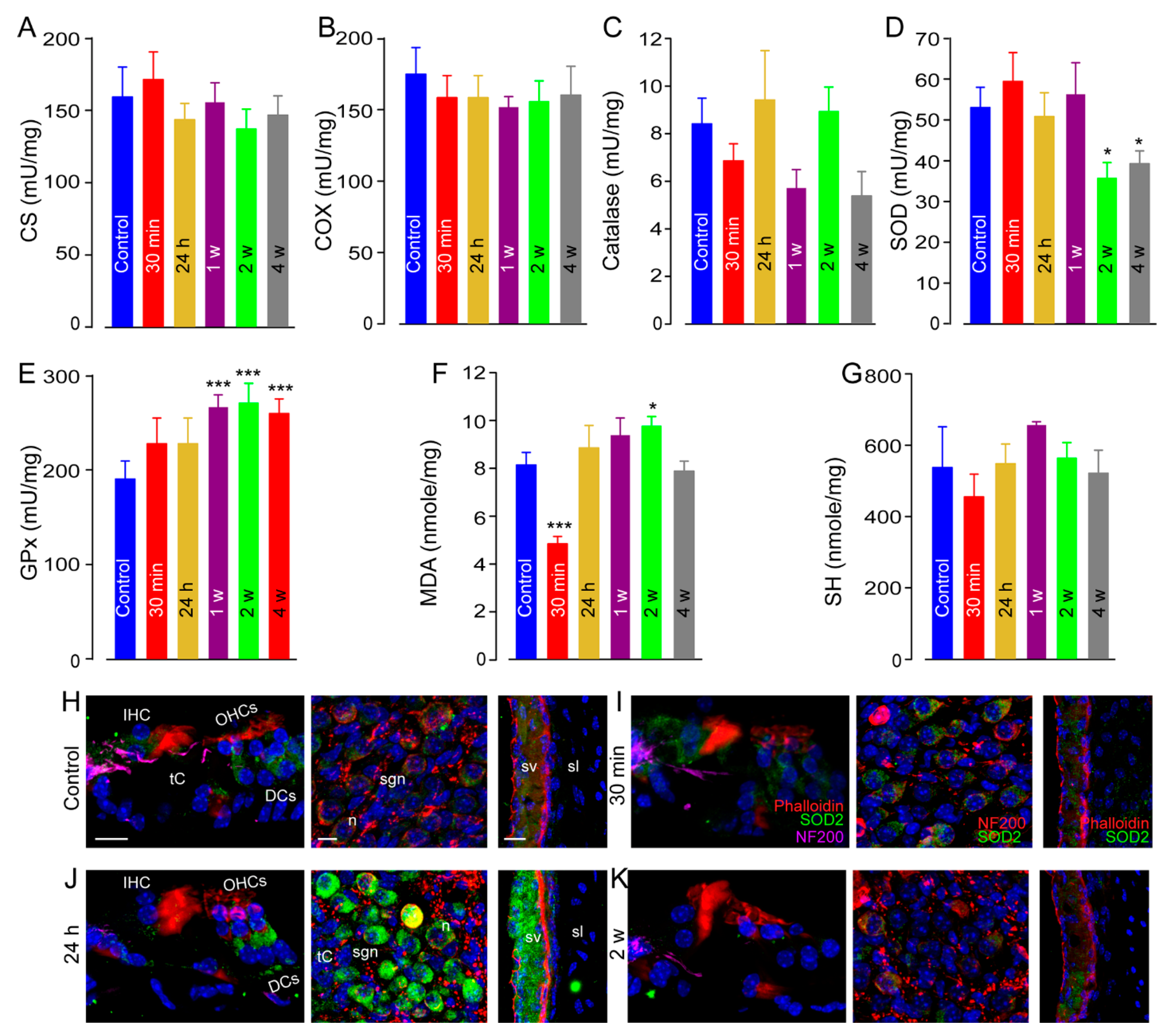
3.6. Oxidative Stress
4. Discussion
4.1. Impulse Noise Did Not Induce Eardrum Rupture
4.2. Reversible Shifts of ABR Thresholds and Reduction of DPOAE Amplitude at Higher, but Not at Lower, Frequencies
4.3. Persistent Reduction of ABR Wave-I Amplitude and Elevated Central Gain
4.4. Oxidative Stress
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Nelson, E.G.; Hinojosa, R. Presbycusis: A Human Temporal Bone Study of Individuals With Downward Sloping Audiometric Patterns of Hearing Loss and Review of the Literature. Laryngoscope 2006, 116, 1–12. [Google Scholar] [CrossRef]
- Stanbury, M.; Rafferty, A.P.; Rosenman, K. Prevalence of Hearing Loss and Work-Related Noise-Induced Hearing Loss in Michigan. J. Occup. Environ. Med. 2008, 50, 72–79. [Google Scholar] [CrossRef]
- Spoendlin, H. Histopathology of noise deafness. J. Otolaryngol. 1985, 14, 282–286. [Google Scholar] [PubMed]
- Miller, E.F.; Graybiel, A. LXIII A Comparison of Ocular Counter-Rolling Movements between Normal Persons and Deaf Subjects with Bilateral Labyrinthine Defects. Ann. Otol. Rhinol. Laryngol. 1963, 72, 885–893. [Google Scholar] [CrossRef]
- Gao, W.-Y.; Ding, D.-L.; Zheng, X.-Y.; Ruan, F.-M.; Liu, Y.-J. A comparison of changes in the stereocilia between temporary and permanent hearing losses in acoustic trauma. Hear. Res. 1992, 62, 27–41. [Google Scholar] [CrossRef]
- Robertson, D. Functional significance of dendritic swelling after loud sounds in the guinea pig cochlea. Hear. Res. 1983, 9, 263–278. [Google Scholar] [CrossRef]
- Pujol, R.; Puel, J.-L. Excitotoxicity, Synaptic Repair, and Functional Recovery in the Mammalian Cochlea: A Review of Recent Findings. Ann. N. Y. Acad. Sci. 1999, 884, 249–254. [Google Scholar] [CrossRef]
- Liberman, M.C. Noise-induced and age-related hearing loss: New perspectives and potential therapies. F1000Research 2017, 6, 927. [Google Scholar] [CrossRef]
- Liberman, M.C. Noise-Induced Hearing Loss: Permanent Versus Temporary Threshold Shifts and the Effects of Hair Cell Versus Neuronal Degeneration. Adv. Exp. Med. Biol. 2016, 875, 1–7. [Google Scholar] [CrossRef] [PubMed]
- Kujawa, S.G.; Liberman, M.C. Adding Insult to Injury: Cochlear Nerve Degeneration after Temporary Noise-Induced Hearing Loss. J. Neurosci. 2009, 29, 14077–14085. [Google Scholar] [CrossRef] [Green Version]
- Kujawa, S.G.; Liberman, M.C. Synaptopathy in the noise-exposed and aging cochlea: Primary neural degeneration in acquired sensorineural hearing loss. Hear. Res. 2015, 330, 191–199. [Google Scholar] [CrossRef] [Green Version]
- Bauer, C.; Brozoski, T.J.; Myers, K.S. Acoustic injury and TRPV1 expression in the cochlear spiral ganglion. Int. Tinnitus J. 2007, 13, 21–28. [Google Scholar] [PubMed]
- Knipper, M.; Van Dijk, P.; Nunes, I.; Rüttiger, L.; Zimmermann, U. Advances in the neurobiology of hearing disorders: Recent developments regarding the basis of tinnitus and hyperacusis. Prog. Neurobiol. 2013, 111, 17–33. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, K.A.; Jeffers, P.W.; Lall, K.; Liberman, M.C.; Kujawa, S.G. Aging after Noise Exposure: Acceleration of Cochlear Synaptopathy in Recovered Ears. J. Neurosci. 2015, 35, 7509–7520. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sergeyenko, Y.; Lall, K.; Liberman, M.C.; Kujawa, S.G. Age-Related Cochlear Synaptopathy: An Early-Onset Contributor to Auditory Functional Decline. J. Neurosci. 2013, 33, 13686–13694. [Google Scholar] [CrossRef]
- Furman, A.C.; Kujawa, S.G.; Liberman, M.C. Noise-induced cochlear neuropathy is selective for fibers with low spontaneous rates. J. Neurophysiol. 2013, 110, 577–586. [Google Scholar] [CrossRef]
- Hickman, T.T.; Smalt, C.; Bobrow, J.; Quatieri, T.; Liberman, M.C. Blast-induced cochlear synaptopathy in chinchillas. Sci. Rep. 2018, 8, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Yankaskas, K.; Hammill, T.; Packer, M.; Zuo, J. Editorial: Auditory injury—A military perspective. Hear. Res. 2017, 349, 1–3. [Google Scholar] [CrossRef]
- Helfer, T.M.; Canham-Chervak, M.; Canada, S.; Mitchener, T.A. Epidemiology of Hearing Impairment and Noise-Induced Hearing Injury Among U.S. Military Personnel, 2003–2005. Am. J. Prev. Med. 2010, 38, S71–S77. [Google Scholar] [CrossRef]
- Lie, A.; Skogstad, M.; Johannessen, H.A.; Tynes, T.; Mehlum, I.S.; Nordby, K.-C.; Engdahl, B.; Tambs, K. Occupational noise exposure and hearing: A systematic review. Int. Arch. Occup. Environ. Health 2016, 89, 351–372. [Google Scholar] [CrossRef] [Green Version]
- Cave, K.M.; Cornish, E.M.; Chandler, D.W. Blast Injury of the Ear: Clinical Update from the Global War on Terror. Mil. Med. 2007, 172, 726–730. [Google Scholar] [CrossRef] [Green Version]
- Tepe, V.; Smalt, C.; Nelson, J.; Quatieri, T.; Pitts, K. Hidden Hearing Injury: The Emerging Science and Military Relevance of Cochlear Synaptopathy. Mil. Med. 2017, 182, e1785–e1795. [Google Scholar] [CrossRef] [Green Version]
- Swan, A.; Nelson, J.; Swiger, B.; Jaramillo, C.; Eapen, B.; Packer, M.; Pugh, M. Prevalence of hearing loss and tinnitus in Iraq and Afghanistan Veterans: A Chronic Effects of Neurotrauma Consortium study. Hear. Res. 2017, 349, 4–12. [Google Scholar] [CrossRef] [PubMed]
- Batrel, C.; Huet, A.; Hasselmann, F.; Wang, J.; Desmadryl, G.; Nouvian, R.; Puel, J.-L.; Bourien, J. Mass Potentials Recorded at the Round Window Enable the Detection of Low Spontaneous Rate Fibers in Gerbil Auditory Nerve. PLoS ONE 2017, 12, e0169890. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Dib, M.; Lenoir, M.; Vago, P.; Eybalin, M.; Hameg, A.; Pujol, R.; Puel, J.-L. Riluzole rescues cochlear sensory cells from acoustic trauma in the guinea-pig. Neuroscience 2002, 111, 635–648. [Google Scholar] [CrossRef]
- Wang, J.; Puel, J.-L. Toward Cochlear Therapies. Physiol. Rev. 2018, 98, 2477–2522. [Google Scholar] [CrossRef] [PubMed]
- Wang, J.; Van De Water, T.R.; Bonny, C.; De Ribaupierre, F.; Puel, J.-L.; Zine, A. A Peptide Inhibitor of c-Jun N-Terminal Kinase Protects against Both Aminoglycoside and Acoustic Trauma-Induced Auditory Hair Cell Death and Hearing Loss. J. Neurosci. 2003, 23, 8596–8607. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liberman, M.C. Hidden Hearing Loss. Sci. Am. 2015, 313, 48–53. [Google Scholar] [CrossRef] [PubMed]
- Qin, J.; Wu, Q.; Walker, J.; Campbell, K.C.; Fox, D. Development of a digital noise exposure system for research on noise induced hearing loss. In Proceedings of the Meetings on Acoustics, 167th Meeting of the Acoustical Society of America (ASA), Providence, RI, USA, 5–9 May 2014. [Google Scholar]
- Gorga, M.P.; Johnson, T.A.; Kaminski, J.R.; Beauchaine, K.L.; Garner, C.A.; Neely, S. Using a Combination of Click- and Tone Burst–Evoked Auditory Brain Stem Response Measurements to Estimate Pure-Tone Thresholds. Ear Hear. 2006, 27, 60–74. [Google Scholar] [CrossRef] [Green Version]
- Ladrech, S.; Wang, J.; Boukhaddaoui, H.; Puel, J.L.; Eybalin, M.; Lenoir, M. Differential expression of PKC beta II in the rat organ of Corti. Eur. J. Neurosci. 2007, 26, 2922–2930. [Google Scholar] [CrossRef] [Green Version]
- Benkafadar, N.; Menardo, J.; Bourien, J.; Nouvian, R.; François, F.; Decaudin, D.; Maiorano, D.; Puel, J.-L.; Wang, J. Reversible p53 inhibition prevents cisplatin ototoxicity without blocking chemotherapeutic efficacy. EMBO Mol. Med. 2017, 9, 7–26. [Google Scholar] [CrossRef] [PubMed]
- Casas, F.; Pessemesse, L.; Grandemange, S.; Seyer, P.; Gueguen, N.; Baris, O.; Lepourry, L.; Cabello, G.; Wrutniak-Cabello, C. Overexpression of the Mitochondrial T3 Receptor p43 Induces a Shift in Skeletal Muscle Fiber Types. PLoS ONE 2008, 3, e2501. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Beers, R.F.; Sizer, I.W. A Spectrophotometric Method for Measuring the Breakdown of Hydrogen Peroxide by Catalase. J. Biol. Chem. 1952, 195, 133–140. [Google Scholar] [CrossRef]
- Marklund, S. Spectrophotometric study of spontaneous disproportionation of superoxide anion radical and sensitive direct assay for superoxide dismutase. J. Biol. Chem. 1976, 251, 7504–7507. [Google Scholar] [CrossRef]
- Janssen, A.J.M.; Trijbels, F.J.M.; Sengers, R.C.A.; Smeitink, J.A.M.; Heuvel, L.P.V.D.; Wintjes, L.T.M.; Stoltenborg-Hogenkamp, B.J.M.; Rodenburg, R. Spectrophotometric Assay for Complex I of the Respiratory Chain in Tissue Samples and Cultured Fibroblasts. Clin. Chem. 2007, 53, 729–734. [Google Scholar] [CrossRef]
- Wharton, D.C.; Tzagoloff, A. studies on the electron transfer system. lvii. the near infrared absorption band of cytochrome oxidase. J. Biol. Chem. 1964, 239, 2036–2041. [Google Scholar] [CrossRef]
- Rustin, P.; Chretien, D.; Bourgeron, T.; Gérard, B.; Rotig, A.; Saudubray, J.; Munnich, A. Biochemical and molecular investigations in respiratory chain deficiencies. Clin. Chim. Acta 1994, 228, 35–51. [Google Scholar] [CrossRef]
- Menardo, J.; Tang, Y.; Ladrech, S.; Lenoir, M.; Casas, F.; Michel, C.; Bourien, J.; Ruel, J.; Rebillard, G.; Maurice, T.; et al. Oxidative Stress, Inflammation, and Autophagic Stress as the Key Mechanisms of Premature Age-Related Hearing Loss in SAMP8 Mouse Cochlea. Antioxid. Redox Signal. 2012, 16, 263–274. [Google Scholar] [CrossRef] [PubMed]
- Bourien, J.; Tang, Y.; Batrel, C.; Huet, A.; Lenoir, M.; Ladrech, S.; Desmadryl, G.; Nouvian, R.; Puel, J.-L.; Wang, J. Contribution of auditory nerve fibers to compound action potential of the auditory nerve. J. Neurophysiol. 2014, 112, 1025–1039. [Google Scholar] [CrossRef] [Green Version]
- Shi, L.; Liu, L.; He, T.; Guo, X.; Yu, Z.; Yin, S.; Wang, J. Ribbon Synapse Plasticity in the Cochleae of Guinea Pigs after Noise-Induced Silent Damage. PLoS ONE 2013, 8, e81566. [Google Scholar] [CrossRef]
- Shi, L.; Guo, X.; Shen, P.; Liu, L.; Tao, S.; Li, X.-W.; Song, Q.; Yu, Z.; Yin, S.; Wang, J. Noise-induced damage to ribbon synapses without permanent threshold shifts in neonatal mice. NeuroReport 2015, 304, 368–377. [Google Scholar] [CrossRef] [Green Version]
- Puel, J.-L.; Ruel, J.; D’aldin, C.G.; Pujol, R. Excitotoxicity and repair of cochlear synapses after noise-trauma induced hearing loss. NeuroReport 1998, 9, 2109–2114. [Google Scholar] [CrossRef] [PubMed]
- Chen, H.; Shi, L.; Liu, L.; Yin, S.; Aiken, S.; Wang, J. Noise-induced Cochlear Synaptopathy and Signal Processing Disorders. Neuroscience 2018, 407, 41–52. [Google Scholar] [CrossRef] [PubMed]
- Michanski, S.; Smaluch, K.; Steyer, A.; Chakrabarti, R.; Setz, C.; Oestreicher, D.; Fischer, C.; Möbius, W.; Moser, T.; Vogl, C.; et al. Mapping developmental maturation of inner hair cell ribbon synapses in the apical mouse cochlea. Proc. Natl. Acad. Sci. USA 2019, 116, 6415–6424. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Fetoni, A.R.; Paciello, F.; Rolesi, R.; Paludetti, G.; Troiani, D. Targeting dysregulation of redox homeostasis in noise-induced hearing loss: Oxidative stress and ROS signaling. Free. Radic. Biol. Med. 2019, 135, 46–59. [Google Scholar] [CrossRef]
- Jahrsdoerfer, R. The Effects of Impulse Noise on the Eardrum and Middle Ear. Otolaryngol. Clin. N. Am. 1979, 12, 515–520. [Google Scholar] [CrossRef]
- Hamernik, R.P.; Turrentine, G.; Roberto, M.; Salvi, R.; Henderson, D. Anatomical correlates of impulse noise-induced mechanical damage in the cochlea. Hear. Res. 1984, 13, 229–247. [Google Scholar] [CrossRef]
- Hu, B.H.; Henderson, D.; Nicotera, T.M. Extremely rapid induction of outer hair cell apoptosis in the chinchilla cochlea following exposure to impulse noise. Hear. Res. 2006, 211, 16–25. [Google Scholar] [CrossRef]
- Nuttall, A.L. Sound-Induced Cochlear Ischemia/Hypoxia as a Mechanism of Hearing Loss. Noise Health 1999, 2, 17–32. [Google Scholar]
- Xiong, M.; He, Q.; Lai, H.; Wang, J. Oxidative stress in spiral ganglion cells of pigmented and albino guinea pigs exposed to impulse noise. Acta Oto-Laryngol. 2011, 131, 914–920. [Google Scholar] [CrossRef]
- Garner, M.J.; Brett, S.J. Mechanisms of Injury by Explosive Devices. Anesthesiol. Clin. 2007, 25, 147–160. [Google Scholar] [CrossRef] [PubMed]
- Cho, S.-I.; Gao, S.; Xia, A.; Wang, R.; Salles, F.T.; Raphael, P.D.; Abaya, H.; Wachtel, J.; Baek, J.; Jacobs, D.; et al. Mechanisms of Hearing Loss after Blast Injury to the Ear. PLoS ONE 2013, 8, e67618. [Google Scholar] [CrossRef] [PubMed]
- Niwa, K.; Mizutari, K.; Matsui, T.; Kurioka, T.; Matsunobu, T.; Kawauchi, S.; Satoh, Y.; Sato, S.; Shiotani, A.; Kobayashi, Y. Pathophysiology of the inner ear after blast injury caused by laser-induced shock wave. Sci. Rep. 2016, 6, 31754. [Google Scholar] [CrossRef] [PubMed]
- Henderson, N. Audiometric and histological correlates of exposure to 1-msec noise impulses in the chinchilla. J. Acoust. Soc. Am. 1974, 56, 1210–1221. [Google Scholar] [CrossRef]
- Song, Q.; Shen, P.; Lijuan, S.; Shi, L.; Liu, L.; Wang, J.; Yu, Z.; Stephen, K.; Aiken, S.; Yin, S.; et al. Coding deficits in hidden hearing loss induced by noise: The nature and impacts. Sci. Rep. 2016, 6, 25200. [Google Scholar] [CrossRef]
- Pacher, P.; Beckman, J.S.; Liaudet, L. Nitric Oxide and Peroxynitrite in Health and Disease. Physiol. Rev. 2007, 87, 315–424. [Google Scholar] [CrossRef] [Green Version]
- Elsayed, N.M.; Armstrong, K.L.; William, M.T.; Cooper, M.F. Antioxidant loading reduces oxidative stress induced by high-energy impulse noise (blast) exposure. Toxicology 2000, 155, 91–99. [Google Scholar] [CrossRef]
- Duan, M.; Qiu, J.; Laurell, G.; Olofsson, Å.; Counter, S.A.; Borg, E. Dose and time-dependent protection of the antioxidant N-l-acetylcysteine against impulse noise trauma. Hear. Res. 2004, 192, 1–9. [Google Scholar] [CrossRef]
- Bielefeld, E.C. Reduction in Impulse Noise-Induced Permanent Threshold Shift with Intracochlear Application of an NADPH Oxidase Inhibitor. J. Am. Acad. Audiol. 2013, 24, 461–473. [Google Scholar] [CrossRef]
- Sharma, P.; Jha, A.B.; Dubey, R.S.; Pessarakli, M. Reactive Oxygen Species, Oxidative Damage, and Antioxidative Defense Mechanism in Plants under Stressful Conditions. J. Bot. 2012, 2012, 217037. [Google Scholar] [CrossRef] [Green Version]






Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Gratias, P.; Nasr, J.; Affortit, C.; Ceccato, J.-C.; François, F.; Casas, F.; Pujol, R.; Pucheu, S.; Puel, J.-L.; Wang, J. Impulse Noise Induced Hidden Hearing Loss, Hair Cell Ciliary Changes and Oxidative Stress in Mice. Antioxidants 2021, 10, 1880. https://doi.org/10.3390/antiox10121880
Gratias P, Nasr J, Affortit C, Ceccato J-C, François F, Casas F, Pujol R, Pucheu S, Puel J-L, Wang J. Impulse Noise Induced Hidden Hearing Loss, Hair Cell Ciliary Changes and Oxidative Stress in Mice. Antioxidants. 2021; 10(12):1880. https://doi.org/10.3390/antiox10121880
Chicago/Turabian StyleGratias, Paul, Jamal Nasr, Corentin Affortit, Jean-Charles Ceccato, Florence François, François Casas, Rémy Pujol, Sylvie Pucheu, Jean-Luc Puel, and Jing Wang. 2021. "Impulse Noise Induced Hidden Hearing Loss, Hair Cell Ciliary Changes and Oxidative Stress in Mice" Antioxidants 10, no. 12: 1880. https://doi.org/10.3390/antiox10121880