
Aminoguanidine Prevents the Oxidative Stress, Inhibiting Elements of Inflammation, Endothelial Activation, Mesenchymal Markers, and Confers a Renoprotective Effect in Renal Ischemia and Reperfusion Injury

 , ,
, , {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:1. Introduction
2. Materials and Methods
2.1. Animals
2.2. Ischemia-Reperfusion (IR) Experiments
2.3. Oxidative Stress Experiments
2.4. Quantification of Neutrophil Gelatinase-Associated Lipocalin
2.5. Glomerular Filtration Rate (GFR) Measurements
2.6. Real-Time PCR
2.7. Western Blot Assay
2.8. Histochemical Analysis and Tissue Damage Determination
2.9. Statistical Analysis
3. Results
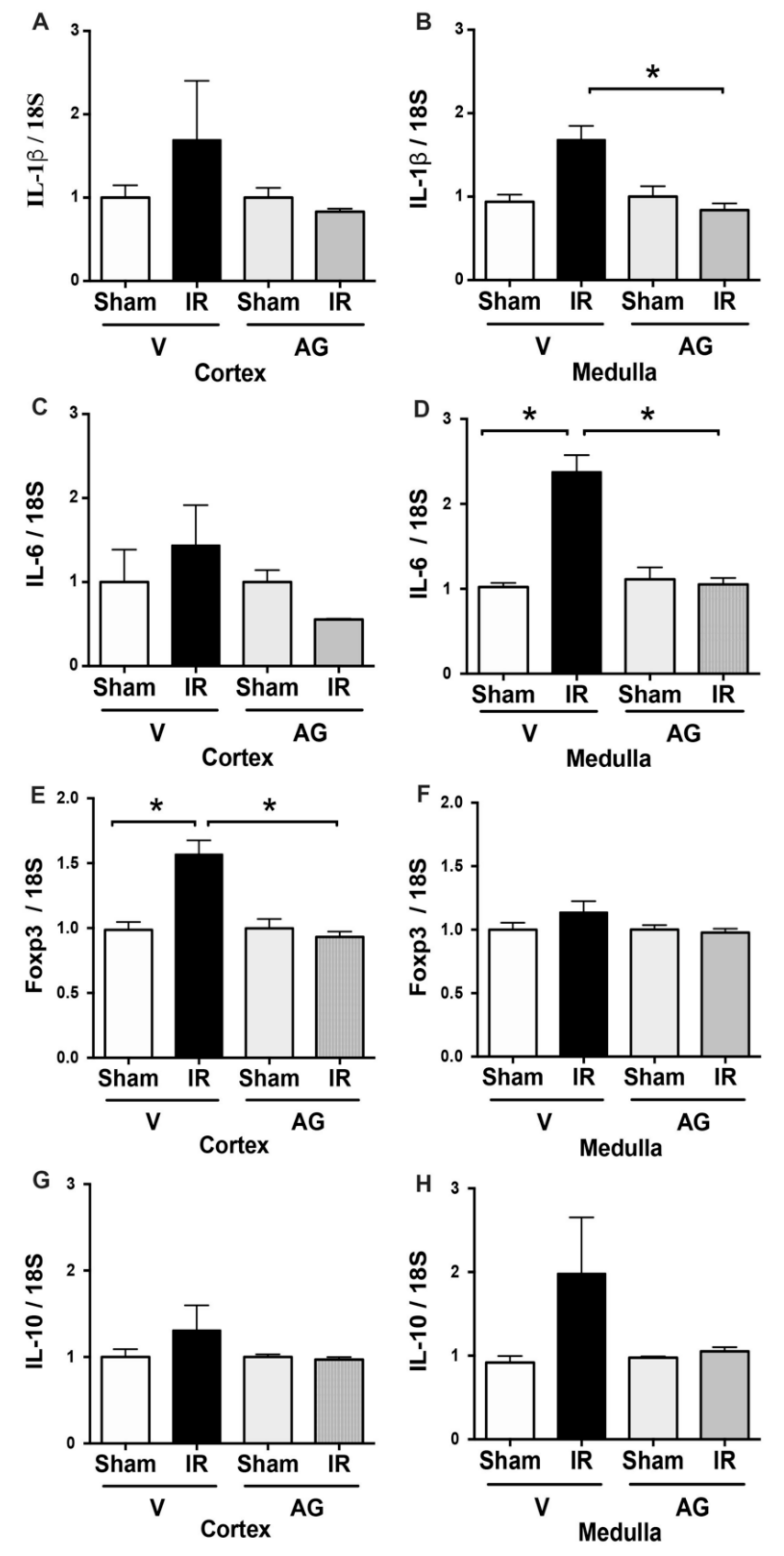
3.1. AG Prevented Kidney Injury due to Renal Ischemia and Reperfusion (IR)
3.2. AG Prevented Oxidative Stress
3.3. Molecular Sensors of AG Antioxidant Effect
3.4. Effect of AG on Endothelial Markers Expression
3.5. Effect of AG on Mesenchymal Markers Expression: Vimentin, Fascin, and Hsp47
4. Discussion
4.1. Antioxidant Effect of AG and Prosurvival Genes
4.2. Antioxidant Effect of AG and Endothelial Injury
4.3. AG and Oxidative Stress in the Kidney
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Itakura, M.; Yoshikawa, H.; Bannai, C.; Kato, M.; Kunika, K.; Kawakami, Y.; Yamaoka, T.; Yamashita, K. Aminoguanidine decreases urinary albumin and high-molecular-weight proteins in diabetic rats. Life Sci. 1991, 49, 889–897. [Google Scholar] [CrossRef]
- Luo, D.; Fan, Y.; Xu, X. The effects of aminoguanidine on retinopathy in STZ-induced diabetic rats. Bioorg. Med. Chem. Lett. 2012, 22, 4386–4390. [Google Scholar] [CrossRef]
- Yagihashi, S.; Kamijo, M.; Baba, M.; Yagihashi, N.; Nagai, K. Effect of aminoguanidine on functional and structural abnormalities in peripheral nerve of STZ-induced diabetic rats. Diabetes 1992, 41, 47–52. [Google Scholar] [CrossRef] [PubMed]
- Yamagishi, S.; Nakamura, N.; Suematsu, M.; Kaseda, K.; Matsui, T. Advanced Glycation End Products: A Molecular Target for Vascular Complications in Diabetes. Mol. Med. 2015, 21, S32–S40. [Google Scholar] [CrossRef] [PubMed]
- Schalkwijk, C.G.; Miyata, T. Early- and advanced non-enzymatic glycation in diabetic vascular complications: The search for therapeutics. Amino Acids 2012, 42, 1193–1204. [Google Scholar] [CrossRef] [Green Version]
- Dobšák, P.; Courderot-Masuyer, C.; Siegelová, J.; Svačinová, H.; Jančík, J.; Vergely-Vanriessen, C.; Rochette, L. Antioxidant properties of aminoguanidine: A Paramagnetic Resonance test. Scr. Med. Fac. Med. Univ. Brun. Masaryk. 2001, 74, 41–50. [Google Scholar]
- Colzani, M.; De Maddis, D.; Casali, G.; Carini, M.; Vistoli, G.; Aldini, G. Reactivity, Selectivity, and Reaction Mechanisms of Aminoguanidine, Hydralazine, Pyridoxamine, and Carnosine as Sequestering Agents of Reactive Carbonyl Species: A Comparative Study. ChemMedChem 2016, 11, 1778–1789. [Google Scholar] [CrossRef] [PubMed]
- Ihm, S.H.; Yoo, H.J.; Park, S.W.; Ihm, J. Effect of aminoguanidine on lipid peroxidation in streptozotocin-induced diabetic rats. Metabolism 1999, 48, 1141–1145. [Google Scholar] [CrossRef]
- Abd El-Gawad, H.M.; El-Sawalhi, M.M. Nitric oxide and oxidative stress in brain and heart of normal rats treated with doxorubicin: Role of aminoguanidine. J. Biochem. Mol. Toxicol. 2004, 18, 699–777. [Google Scholar] [CrossRef]
- Tunc, T.; Kesik, V.; Demirin, H.; Ersoz, N.; Vurucu, S.; Kul, M.; Uysal, B.; Sadir, S.; Guven, A.; Oztas, E. Effects of aminoguanidine and melatonin on intestinal ischemia/reperfusion injury in rats: An assessor-blinded, controlled experimental study. Curr. Ther. Res. Clin. Exp. 2009, 6, 449–459. [Google Scholar] [CrossRef] [Green Version]
- Esson, M.L.; Schrier, R.W. Diagnosis and treatment of acute tubular necrosis. Ann. Intern. Med. 2002, 9, 744–752. [Google Scholar] [CrossRef]
- Noiri, E.; Nakao, A.; Uchida, K.; Tsukahara, H.; Ohno, M.; Fujita, T.; Brodsky, S.; Goligorsky, M.S. Oxidative and nitrosative stress in acute renal ischemia. AJP-Ren. Physiol. 2001, 281, F948–F957. [Google Scholar] [CrossRef] [PubMed]
- Paller, M.S.; Hoidal, J.R.; Ferris, T.F. Oxygen free radicals in ischemic acute renal failure in the rat. J. Clin. Investig. 1984, 74, 1156–1164. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Goligorsky, M.S.; Brodsky, S.V.; Noiri, E. Nitric oxide in acute renal failure: NOS versus NOS. Kidney Int. 2002, 61, 855–861. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ratliff, B.B.; Abdulmahdi, W.; Pawar, R.; Wolin, M.S. Oxidant mechanisms in renal injury and disease. Antioxid. Redox Signal. 2016, 25, 119–146. [Google Scholar] [CrossRef] [Green Version]
- Ozbek, E. Induction of oxidative stress in kidney. Int. J. Nephrol. 2012, 2012, 465897. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dounousi, E.; Papavasiliou, E.; Makedou, A.; Ioannou, K.; Katopodis, K.P.; Tselepis, A.; Siamopoulos, K.C.; Tsakiris, D. Oxidative Stress Is Progressively Enhanced with Advancing Stages of CKD. Am. J. Kidney Dis. 2006, 48, 752–760. [Google Scholar] [CrossRef] [Green Version]
- Pasten, C.; Alvarado, C.; Rocco, J.; Contreras, L.; Aracena, P.; Liberona, J.; Suazo, C.; Michea, L.F.; Irarrázabal, C.E. L-NIL prevents the ischemia and reperfusion injury involving TRL4, GST, clusterin and NFAT5 in mice. AJP Ren. Physiol. 2018, 316, F624–F634. [Google Scholar] [CrossRef]
- Fatemikia, H.; Ketabchi, F.; Karimi, Z.; Moosavi, S.M. Distant effects of unilateral renal ischemia/reperfusion on contralateral kidney but not lung in rats: The roles of ROS and iNOS. Can. J. Physiol. Pharmacol. 2016, 94, 447–487. [Google Scholar] [CrossRef]
- Sahna, E.; Parlakpinar, H.; Cihan, O.F.; Turkoz, Y.; Acet, A. Effects of aminoguanidine against renal ischaemia-reperfusion injury in rats. Cell Biochem. Funct. 2006, 24, 137–141. [Google Scholar] [CrossRef]
- Onem, Y.; Ipcioglu, O.M.; Haholu, A.; Sen, H.; Aydinoz, S.; Suleymanoglu, S.; Bilgi, O.; Akyol, I. Posttreatment with aminoguanidine attenuates renal ischemia/reperfusion injury in rats. Ren. Fail. 2009, 31, 50–53. [Google Scholar] [CrossRef] [PubMed]
- Tsuji, M.; Higuchi, Y.; Shiraishi, K.; Kume, T.; Akaike, A.; Hattori, H. Protective effect of aminoguanidine on hypoxic-ischemic brain damage and temporal profile of brain nitric oxide in neonatal rat. Pediatric Res. 2000, 47, 79–83. [Google Scholar] [CrossRef] [Green Version]
- Parlakpinar, H.; Ozer, M.K.; Acet, A. Effect of aminoguanidine on ischemia-reperfusion induced myocardial injury in rats. Mol. Cell Biochem. 2005, 277, 137–142. [Google Scholar] [CrossRef] [PubMed]
- Sugimoto, K.; Iadecola, C. Effects of aminoguanidine on cerebral ischemia in mice: Comparison between mice with and without inducible nitric oxide synthase gene. Neurosci. Lett. 2002, 331, 25–28. [Google Scholar] [CrossRef]
- Mattson, D.L.; Wu, F. Nitric oxide synthase activity and isoforms in rat renal vasculature. Hypertension 2000, 35, 337–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Guo, Q.; Du, X.; Zhao, Y.; Zhang, D.; Yue, L.; Wang, Z. Ischemic postconditioning prevents renal ischemia reperfusion injury through the induction of heat shock proteins in rats. Mol. Med. Rep. 2014, 10, 2875–2881. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chebotareva, N.; Bobkova, I.; Shilov, E. Heat shock proteins and kidney disease: Perspectives of HSP therapy. Cell Stress Chaperones 2017, 22, 319–343. [Google Scholar] [CrossRef]
- Kim, M.; Park, S.W.; Kim, M.; Chen, S.W.; Gerthoffer, W.T.; D’Agati, V.D.; Lee, H.T. Selective renal overexpression of human heat shock protein 27 reduces renal ischemia-reperfusion injury in mice. Am. J. Physiol. Ren. Physiol. 2010, 299, F347–F358. [Google Scholar] [CrossRef] [Green Version]
- Wetzel, M.D.; Stanley, K.; Wang, W.W.; Maity, S.; Madesh, M.; Reeves, W.B.; Awad, A.S. Selective inhibition of arginase-2 in endothelial cells but not proximal tubules reduces renal fibrosis. JCI Insight 2020, 5, e142187. [Google Scholar] [CrossRef]
- Chung, S.; Overstreet, J.M.; Li, Y.; Wang, Y.; Niu, A.; Wang, S.; Fan, X.; Sasaki, K.; Jin, G.N.; Khodo, S.N.; et al. TGF-β promotes fibrosis after severe acute kidney injury by enhancing renal macrophage infiltration. JCI Insight 2018, 3, e123563. [Google Scholar] [CrossRef]
- Cruz-Solbes, A.S.; Youker, K. Epithelial to Mesenchymal Transition (EMT) and Endothelial to Mesenchymal Transition (EndMT): Role and Implications in Kidney Fibrosis. Results Probl. Cell Differ. 2017, 60, 345–372. [Google Scholar] [CrossRef] [PubMed]
- Kovacic, J.C.; Mercader, N.; Torres, M.; Boehm, M.; Fuster, V. Epithelial-to-mesenchymal and endothelial-to-mesenchymal transition from cardiovascular development to disease. Circulation 2012, 125, 1795–1808. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lamouille, S.; Xu, J.; Derynck, R. Molecular mechanisms of epithelial-mesenchymal transition. Nat. Rev. Mol. Cell Biol. 2014, 15, 178–196. [Google Scholar] [CrossRef] [Green Version]
- Ranchoux, B.; Antigny, F.; Rucker-Martin, C.; Hautefort, A.; Péchoux, C.; Bogaard, H.J.; Dorfmüller, P.; Remy, S.; Lecerf, F.; Planté, S.; et al. Endothelial-to-mesenchymal transition in pulmonary hypertension. Circulation 2015, 131, 1006–1018. [Google Scholar] [CrossRef] [Green Version]
- LeBleu, V.S.; Taduri, G.; O’Connell, J.; Teng, Y.; Cooke, V.G.; Woda, C.; Sugimoto, H.; Kalluri, R. Origin and function of myofibroblasts in kidney fibrosis. Nat. Med. 2013, 19, 1047–1053. [Google Scholar] [CrossRef]
- Xu-Dubois, Y.C.; Ahmadpoor, P.; Brocheriou, I.; Kevin, L.; Arzouk Snanoudj, N.; Rouvier, P.; Taupin, J.-L.; Corchia, A.; Galichon, P.; Barrou, B.; et al. Microvasculature partial endothelial mesenchymal transition in early posttransplant biopsy with acute tubular necrosis identifies poor recovery renal allografts. Am. J. Transplant. 2020, 20, 2400–2412. [Google Scholar] [CrossRef]
- Wei, Q.; Dong, Z. Mouse model of ischemic acute kidney injury: Technical notes and tricks. Am. J. Physiol. Renal. Physiol. 2012, 303, F1487–F1494. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lowry, O.H.; Rosebrough, N.J.; Farr, A.L.; Randall, R.J. Protein measurement with the Folin phenol reagent. J. Biol. Chem. 1951, 193, 265. [Google Scholar] [CrossRef]
- Ohkawa, H.; Ohishi, N.; Yagi, K. Assay for lipid peroxides in animal tissues by thiobarbituric acid reaction. Anal. Biochem. 1979, 95, 351–358. [Google Scholar] [CrossRef]
- Tietze, F. Enzymic method for quantitative determination of nanogram amounts of total and oxidized glutathione: Applications to mammalian blood and other tissues. Anal. Biochem. 1969, 27, 502–522. [Google Scholar] [CrossRef]
- Giani, J.F.; Bernstein, K.E.; Janjulia, T.; Han, J.; Toblli, J.E.; Shen, X.Z.; Rodriguez-Iturbe, B.; McDonough, A.A.; Gonzalez-Villaloboset, G.A. Salt Sensitivity in Response to Renal Injury Requires Renal Angiotensin-Converting Enzyme. Hypertension 2015, 66, 534. [Google Scholar] [CrossRef] [Green Version]
- Lieberthal, W. Biology of ischemic and toxic renal tubular cell injury: Role of nitric oxide and the inflammatory response. Curr. Opin. Nephrol. Hypertens. 1998, 7, 289–295. [Google Scholar] [CrossRef]
- Taylor, E.L.; Megson, I.L.; Haslett, C.; Rossi, A.G. Nitric oxide: A key regulator of myeloid inflammatory cell apoptosis. Cell Death Differ. 2003, 10, 418–430. [Google Scholar] [CrossRef] [Green Version]
- Mark, L.A.; Robinson, A.V.; Schulak, J.A. Inhibition of nitric oxide synthase reduces renal ischemia/reperfusion injury. J. Surg. Res. 2005, 129, 236–241. [Google Scholar] [CrossRef]
- Vidyasagar, A.; Reese, S.; Acun, Z.; Hullett, D.; Djamali, A. HSP27 is involved in the pathogenesis of kidney tubulointerstitial fibrosis. Am. J. Physiol. Ren. Physiol. 2008, 295, F707–F716. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, S.W.; Chen, S.W.; Kim, M.; D’Agati, V.D.; Lee, H.T. Human heat shock protein 27-overexpressing mice are protected against acute kidney injury after hepatic ischemia and reperfusion. Am. J. Physiol. Ren. Physiol. 2009, 297, F885–F894. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, A.; Gupta, S. The multifaceted role of glutathione S-transferases in cancer. Cancer Lett. 2018, 433, 33–42. [Google Scholar] [CrossRef]
- Arrigo, A.P.; Virot, S.; Chaufour, S.; Firdaus, W.; Kretz-Remy, C.; Diaz-Latoud, C. Hsp27 consolidates intracellular redox homeostasis by upholding glutathione in its reduced form and by decreasing iron intracellular levels. Antioxid. Redox Signal. 2005, 7, 414–422. [Google Scholar] [CrossRef] [PubMed]
- Jin, C.; Cleveland, J.C.; Ao, L.; Li, J.; Zeng, Q.; Fullerton, D.A.; Meng, X. Human myocardium releases heat shock protein 27 (HSP27) after global ischemia: The proinflammatory effect of extracellular HSP27 through toll-like receptor (TLR)-2 and TLR4. Mol. Med. 2014, 20, 280–289. [Google Scholar] [CrossRef] [PubMed]
- Stetler, R.A.; Gao, Y.; Signore, A.P.; Cao, G.; Chen, J. HSP27: Mechanisms of cellular protection against neuronal injury. Curr. Mol. Med. 2007, 9, 863–872. [Google Scholar] [CrossRef] [Green Version]
- Efthymiou, C.A.; Mocanu, M.M.; de Belleroche, J.; Wells, D.J.; Latchmann, D.S.; Yellon, D.M. Heat shock protein 27 protects the heart against myocardial infarction. Basic Res. Cardiol. 2004, 99, 392–394. [Google Scholar] [CrossRef]
- Xiong, B.; Li, M.; Xiang, S.; Han, L. A1AR-mediated renal protection against ischemia/reperfusion injury is dependent on HSP27 induction. Int. Urol. Nephrol. 2018, 50, 1355–1363. [Google Scholar] [CrossRef] [PubMed]
- Trougakos, I.P. The molecular chaperone apolipoprotein J/clusterin as a sensor of oxidative stress: Implications in therapeutic approaches—A mini-review. Gerontology 2013, 59, 514–523. [Google Scholar] [CrossRef]
- He, J.; Dijkstra, K.L.; Bakker, K.; Bus, P.; Bruijn, J.A.; Scharpfenecker, M.; Baelde, H.J. Glomerular clusterin expression is increased in diabetic nephropathy and protects against oxidative stress-induced apoptosis in podocytes. Sci. Rep. 2020, 10, 14888. [Google Scholar] [CrossRef]
- Guan, X.; Qian, Y.; Shen, Y.; Zhang, L.; Du, Y.; Dai, H.; Qian, J.; Yan, Y. Autophagy protects renal tubular cells against ischemia/reperfusion injury in a time-dependent manner. Cell Physiol. Biochem. 2015, 36, 285–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kaushal, G.P. Autophagy protects proximal tubular cells from injury and apoptosis. Kidney Int. 2012, 82, 1250–1253. [Google Scholar] [CrossRef] [Green Version]
- Paulus, P.; Rupprecht, K.; Baer, P.; Obermüller, N.; Penzkofer, D.; Reissig, C.; Scheller, B.; Holfeld, J.; Zacharowski, K.; Dimmeler, S.; et al. The early activation of toll-like receptor (TLR)-3 initiates kidney injury after ischemia and reperfusion. PLoS ONE 2014, 9, e94366. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Newman, P.J.; Newman, D.K. Signal transduction pathways mediated by PECAM-1: New roles for an old molecule in platelet and vascular cell biology. Arterioscler. Thromb. Vasc. Biol. 2003, 23, 953–964. [Google Scholar] [CrossRef] [PubMed]
- Eleftheriadis, T.; Pissas, G.; Filippidis, G.; Liakopoulos, V.; Stefanidis, I. Reoxygenation induces reactive oxygen species production and ferroptosis in renal tubular epithelial cells by activating aryl hydrocarbon receptor. Mol. Med. Rep. 2021, 23, 41. [Google Scholar] [CrossRef]
- Chatterjee, P.K.; Patel, N.S.A.; Kvale, E.O.; Cuzzocrea, S.; Brown, P.A.J.; Stewart, K.N.; Mota-Filipe, H.; Thiemermann, C. Inhibition of inducible nitric oxide synthase reduces renal ischemia/reperfusion injury. Kidney Int. 2002, 61, 862–871. [Google Scholar] [CrossRef] [Green Version]
- Radi, R.; Beckman, J.S.; Bush, K.M.; Freeman, B.A. Peroxynitrite-induced membrane lipid peroxidation: The cytotoxic potential of superoxide and nitric oxide. Arch. Biochem. Biophys. 1991, 288, 481–487. [Google Scholar] [CrossRef]
- Qian, J.; You, H.; Zhu, Q.; Ma, S.; Zhou, Y.; Zheng, Y.; Liu, J.; Kuang, D.; Gu, Y.; Hao, C.; et al. Nitrotyrosine level was associated with mortality in patients with acute kidney injury. PLoS ONE 2013, 8, E7996. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, J.U. Nitric oxide in the kidney: Its physiological role and pathophysiological implications. Electrolyte Blood Press 2008, 6, 27–34. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Legrand, M.; Almac, E.; Mik, E.G.; Johannes, T.; Kandil, A.; Bezemer, R.; Payen, D.; Ince, C. L-NIL prevents renal microvascular hypoxia and increase of renal oxygen consumption after ischemia-reperfusion in rats. AJP Ren. Physiol. 2009, 296, F1109–F1117. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Abraham, P.; Rabi, S. Protective effect of aminoguanidine against cyclophosphamide-induced oxidative stress and renal damage in rats. Redox Rep. 2011, 16, 8–14. [Google Scholar] [CrossRef] [PubMed]
- Polat, A.; Parlakpinar, H.; Tasdemir, S.; Colak, C.; Vardi, N.; Ucar, M.; Emre, M.H.; Acet, A. Protective role of aminoguanidine on gentamicin-induced acute renal failure in rats. Acta Histochem. 2006, 108, 365–371. [Google Scholar] [CrossRef]








Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pasten, C.; Lozano, M.; Rocco, J.; Carrión, F.; Alvarado, C.; Liberona, J.; Michea, L.; Irarrázabal, C.E. Aminoguanidine Prevents the Oxidative Stress, Inhibiting Elements of Inflammation, Endothelial Activation, Mesenchymal Markers, and Confers a Renoprotective Effect in Renal Ischemia and Reperfusion Injury. Antioxidants 2021, 10, 1724. https://doi.org/10.3390/antiox10111724
Pasten C, Lozano M, Rocco J, Carrión F, Alvarado C, Liberona J, Michea L, Irarrázabal CE. Aminoguanidine Prevents the Oxidative Stress, Inhibiting Elements of Inflammation, Endothelial Activation, Mesenchymal Markers, and Confers a Renoprotective Effect in Renal Ischemia and Reperfusion Injury. Antioxidants. 2021; 10(11):1724. https://doi.org/10.3390/antiox10111724
Chicago/Turabian StylePasten, Consuelo, Mauricio Lozano, Jocelyn Rocco, Flavio Carrión, Cristobal Alvarado, Jéssica Liberona, Luis Michea, and Carlos E. Irarrázabal. 2021. "Aminoguanidine Prevents the Oxidative Stress, Inhibiting Elements of Inflammation, Endothelial Activation, Mesenchymal Markers, and Confers a Renoprotective Effect in Renal Ischemia and Reperfusion Injury" Antioxidants 10, no. 11: 1724. https://doi.org/10.3390/antiox10111724