
Airway Thiol-NO Adducts as Determinants of Exhaled NO

 ,
,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Regulatory Information
2.1.1. Human Subjects Research
2.1.2. Animal Studies
2.2. Methods for Study 1
2.3. Methods for Study 2
2.4. Methods for Study 3
2.5. Statistical Analysis
3. Results
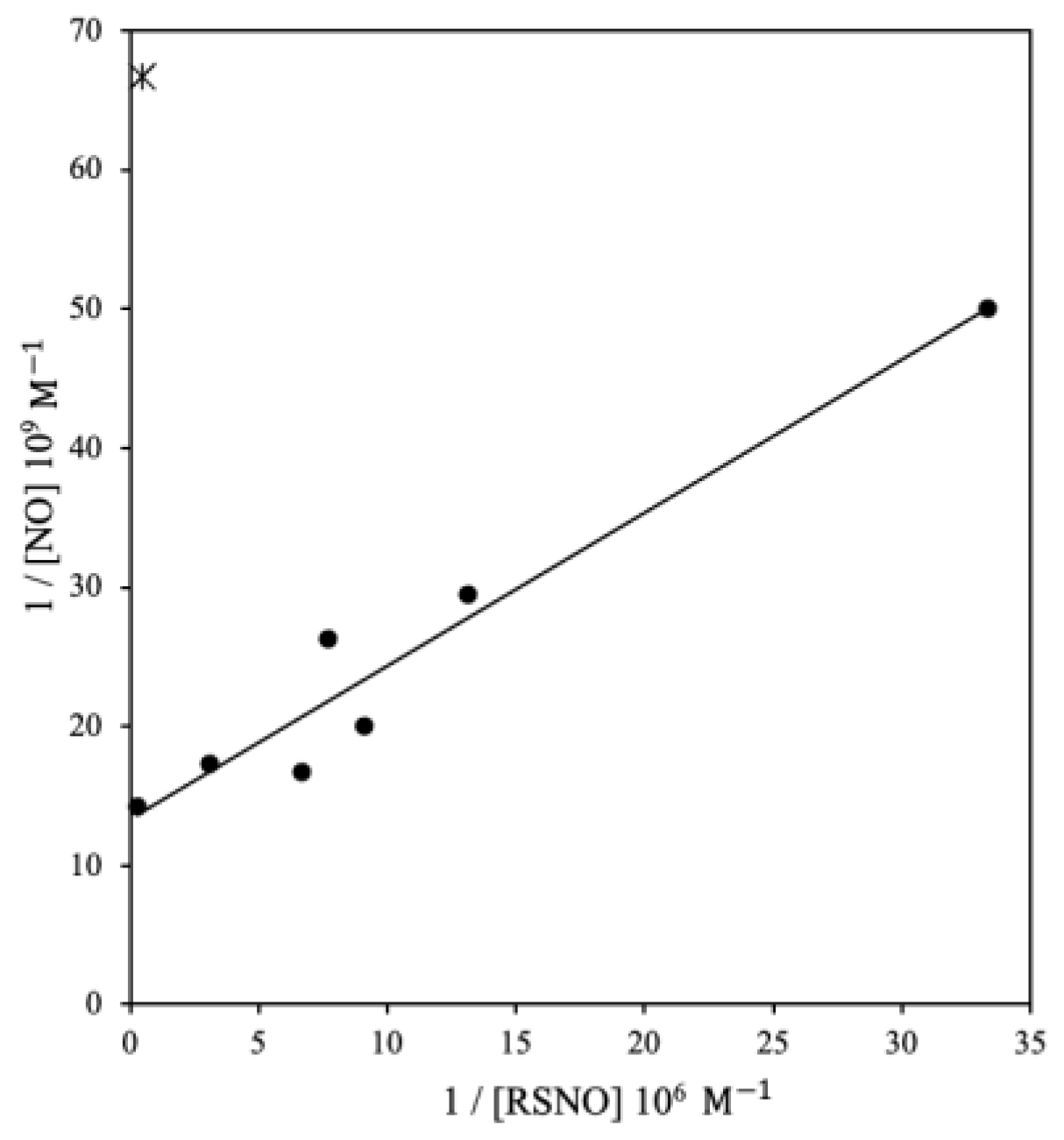
3.1. Study 1. Data from Children with Respiratory Failure
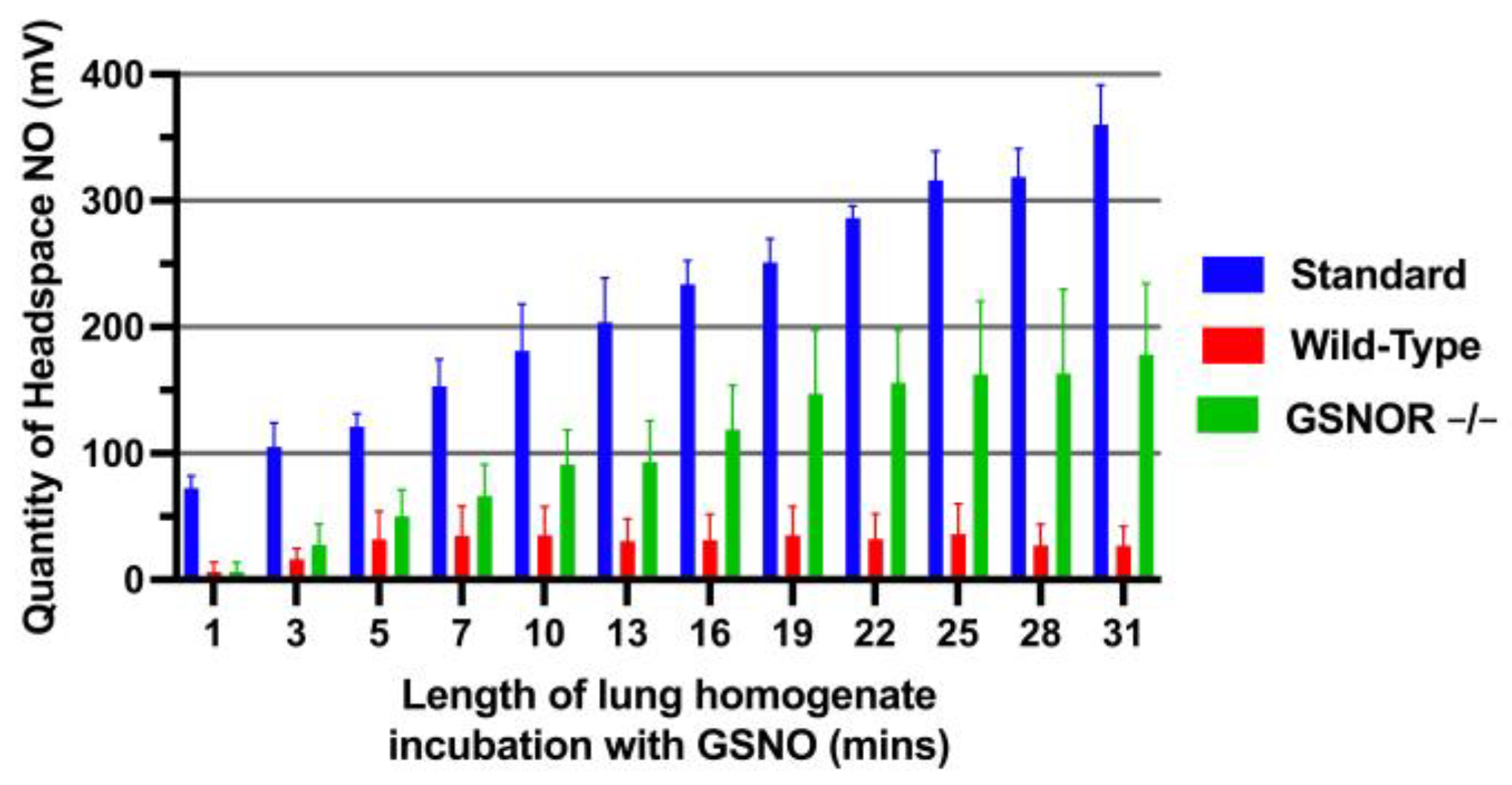
3.2. Study 2. Headspace Nitric Oxide Assay in Murine Lungs
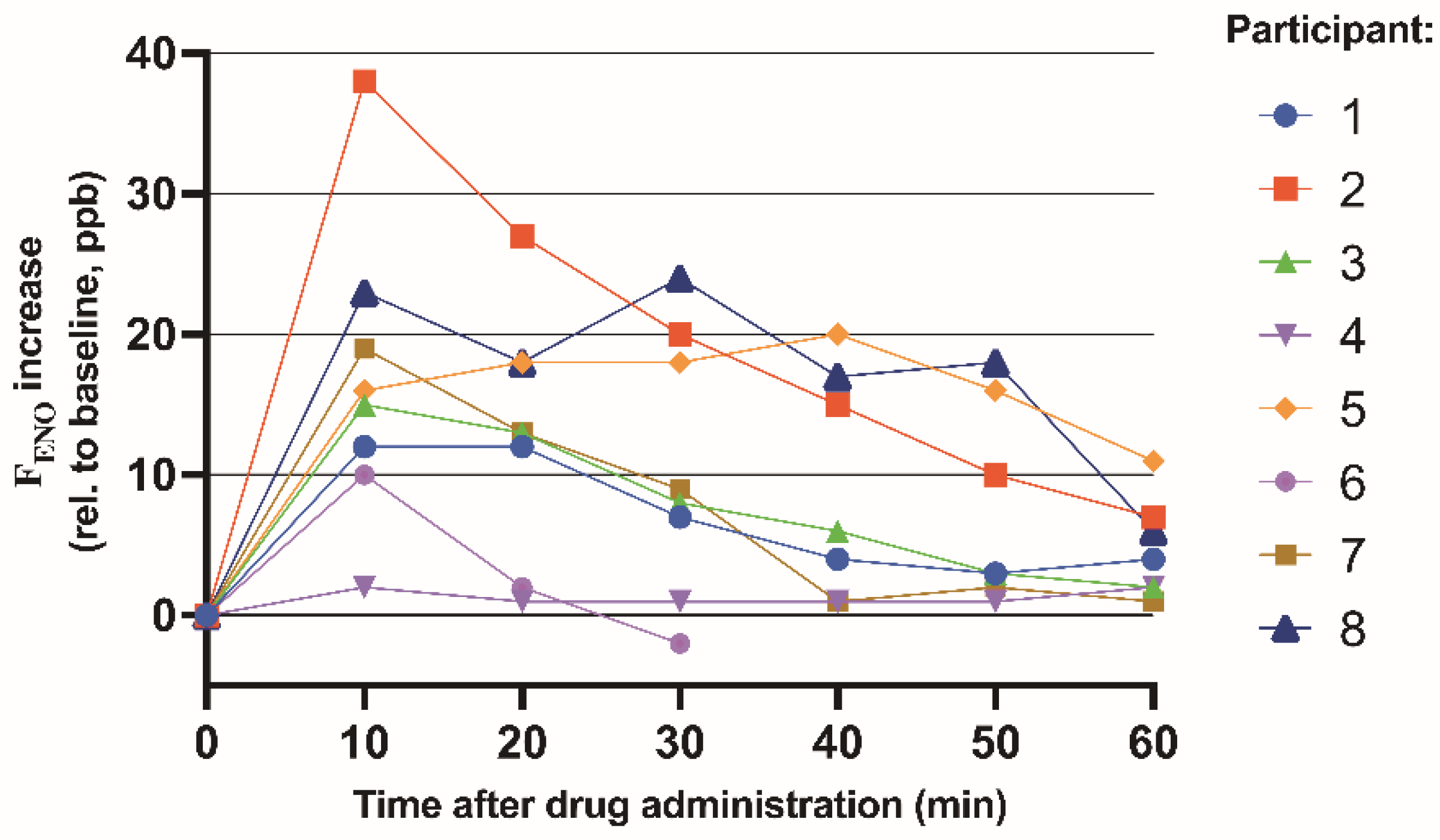
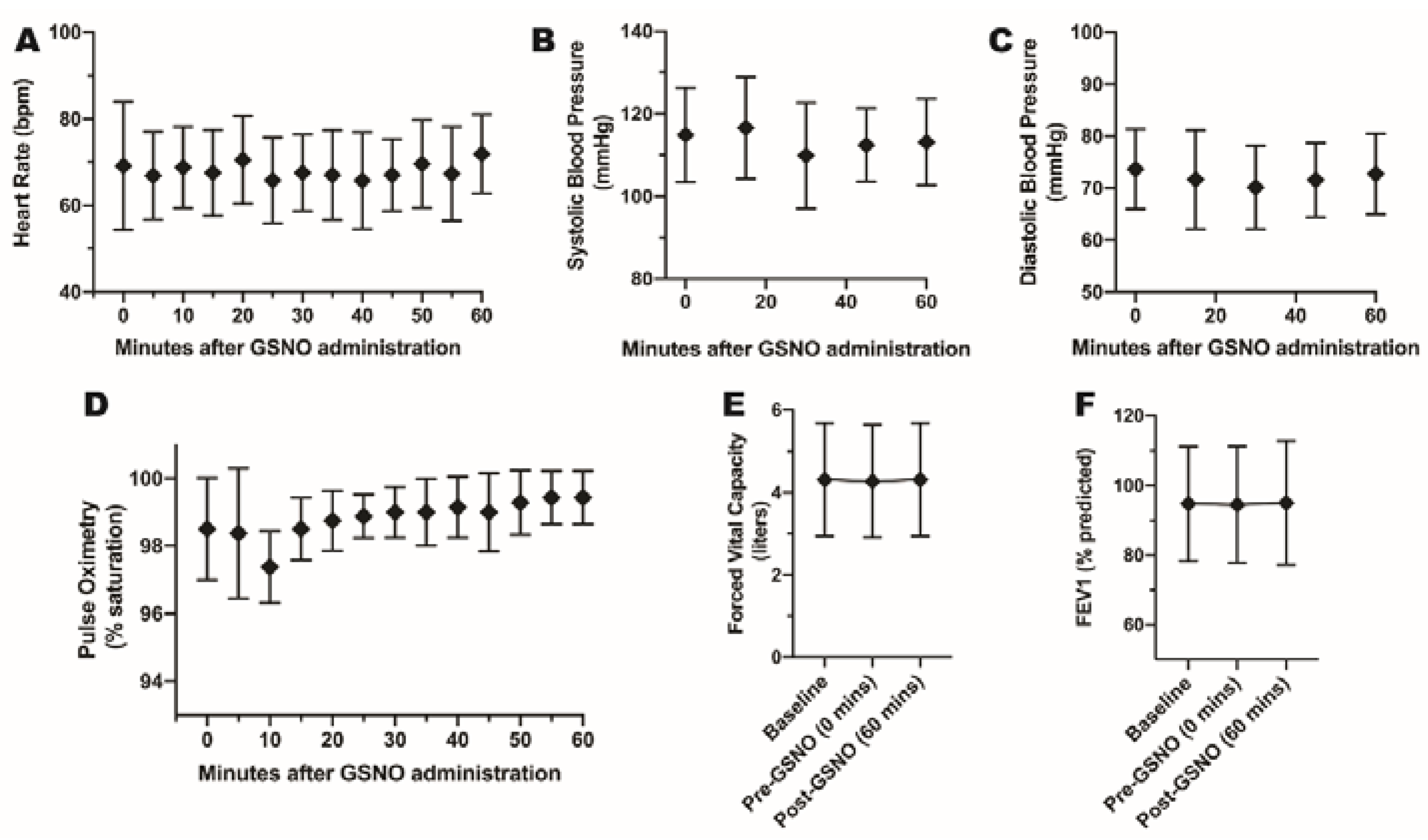
3.3. Study 3. Healthy Human FENO Response to Inhaled GSNO
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Gaston, B.; Drazen, J.M.; Jansen, A.; Sugarbaker, D.A.; Loscalzo, J.; Stamler, J.S. Relaxation of human bronchial smooth muscle by S-nitrosothiols in vitro. J. Pharmacol. Exp. Ther. 1994, 268, 978–984. [Google Scholar]
- Perkins, W.J.; Pabelick, C.; Warner, D.O.; Jones, K.A. cGMP-independent mechanism of airway smooth muscle relaxation induced by S-nitrosoglutathione. Am. J. Physiol. 1998, 275, C468–C474. [Google Scholar] [CrossRef]
- Bannenberg, G.; Xue, J.; Engman, L.; Cotgreave, I.; Moldéus, P.; Ryrfeldt, A. Characterization of bronchodilator effects and fate of S-nitrosothiols in the isolated perfused and ventilated guinea pig lung. J. Pharmacol. Exp. Ther. 1995, 272, 1238–1245. [Google Scholar]
- Gaston, B.; Sears, S.; Woods, J.; Hunt, J.; Ponaman, M.; McMahon, T.; Stamler, J. Bronchodilator S-nitrosothiol deficiency in asthmatic respiratory failure. Lancet 1998, 351, 1317–1319. [Google Scholar] [CrossRef]
- Marozkina, N.V.; Gaston, B. Nitrogen chemistry and lung physiology. Ann. Rev. Physiol. 2015, 77, 431–452. [Google Scholar] [CrossRef] [PubMed]
- Gow, A.J.; Stamler, J.S. Reactions between nitric oxide and haemoglobin under physiological conditions. Nature 1998, 391, 169–173. [Google Scholar] [CrossRef] [PubMed]
- Gow, A.J.; Chen, Q.; Hess, D.T.; Day, B.J.; Ischiropoulos, H.; Stamler, J.S. Basal and stimulated protein S-nitrosylation in multiple cell types and tissues. J. Biol. Chem. 2002, 277, 9637–9640. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Smith, B.C.; Marletta, M.A. Mechanisms of S-nitrosothiol formation and selectivity in nitric oxide signaling. Curr. Opin. Chem. Biol. 2012, 16, 498–506. [Google Scholar] [CrossRef] [Green Version]
- Schmidt, H.H.; Walter, U. NO at work. Cell 1994, 78, 919–925. [Google Scholar] [CrossRef]
- Inoue, K.; Akaike, T.; Miyamoto, Y.; Okamoto, T.; Sawa, T.; Otagiri, M.; Suzuki, S.; Yoshimura, T.; Maeda, H. Nitrosothiol formation catalyzed by ceruloplasmin. Implication for cytoprotective mechanism in vivo. J. Biol. Chem. 1999, 274, 27069–27075. [Google Scholar] [CrossRef] [Green Version]
- Gaston, B.; Reilly, J.; Drazen, J.M.; Fackler, J.; Ramdev, P.; Arnelle, D.; Mullins, M.E.; Sugarbaker, D.J.; Chee, C.; Singel, D.J.; et al. Endogenous nitrogen oxides and bronchodilator S-nitrosothiols in human airways. Proc. Natl. Acad. Sci. USA 1993, 90, 10957–10961. [Google Scholar] [CrossRef] [Green Version]
- Arnelle, D.R.; Stamler, J.S. NO+, NO, and NO- donation by S-nitrosothiols: Implications for regulation of physiological functions by S-nitrosylation and acceleration of disulfide formation. Arch. Biochem. Biophys. 1995, 318, 279–285. [Google Scholar] [CrossRef] [PubMed]
- Hogg, N.; Singh, R.J.; Kalyanaraman, B. The role of glutathione in the transport and catabolism of nitric oxide. FEBS Lett. 1996, 382, 223–228. [Google Scholar] [CrossRef] [Green Version]
- Wong, P.S.; Hyun, J.; Fukuto, J.M.; Shirota, F.N.; DeMaster, E.G.; Shoeman, D.W.; Nagasawa, H.T. Reaction between S-nitrosothiols and thiols: Generation of nitroxyl (HNO) and subsequent chemistry. Biochemistry 1998, 37, 5362–5371. [Google Scholar] [CrossRef]
- GSNOR Phenotyping/GSNO Challenge in Severe Asthma (NCT03926741). Available online: https://clinical-trials.gov/ct2/show/NCT0392674 (accessed on 17 June 2020).
- Barnett, S.D.; Buxton, I.L. The role of S-nitrosoglutathione reductase (GSNOR) in human disease and therapy. Crit. Rev. Biochem. Mol. Biol. 2017, 52, 340–354. [Google Scholar] [CrossRef] [PubMed]
- Zhou, H.L.; Zhang, R.; Anand, P.; Stomberski, C.T.; Qian, Z.; Hausladen, A.; Wang, L.; Rhee, P.; Parikh, S.M.; Karumanchi, A.; et al. Metabolic reprogramming by the S-nitroso-CoA reductase system protects against kidney injury. Nature 2019, 565, 96–100. [Google Scholar] [CrossRef] [PubMed]
- Nelson, B.; Sears, S.; Woods, J.; Ling, C.Y.; Hunt, J.; Clapper, L.M.; Gaston, B. Expired nitric oxide as a marker for childhood asthma. J. Pediatr. 1997, 130, 423–427. [Google Scholar] [CrossRef]
- Buchvald, F.; Baraldi, E.; Carraro, S.; Gaston, B.; De Jongste, J.; Pijnenburg, M.W.; Silkoff, P.E.; Bisgaard, H. Measurements of exhaled nitric oxide in healthy subjects age 4 to 17 years. J. Allergy Clin. Immunol. 2005, 115, 1130–1136. [Google Scholar] [CrossRef] [PubMed]
- Gaston, B.; Massaro, A.; Drazen, J.; Chee, C.B.E.; Wohl, M.E.B.; Stamler, J.S. Expired nitric oxide levels are elevated in patients with asthma. Nitric Oxide 1994, 3, 497–499. [Google Scholar]
- Dweik, R.A.; Sorkness, R.; Wenzel, S.; Hammel, J.; Curran-Everett, D.; Comhair, S.A.A.; Bleecker, E.; Busse, W.; Calhoun, W.; Castro, M.; et al. Use of exhaled nitric oxide measurement to identify a reactive, at-risk phenotype among patients with asthma. Am. J. Respir. Crit. Care Med. 2010, 181, 1033–1041. [Google Scholar] [CrossRef] [Green Version]
- Alving, K.; Weitzberg, E.; Lundberg, J.M. Increased amount of nitric oxide in exhaled air of asthmatics. Eur. Respir. J. 1993, 6, 1368. [Google Scholar]
- Hatziagorou, E.; Tsanakas, J. Assessment of airway inflammation with exhaled NO measurement. Hippokratia 2007, 11, 51–62. [Google Scholar]
- Gaston, B.; Ratjen, F.; Vaughan, J.W.; Malhotra, N.R.; Canady, R.G.; Snyder, A.H.; Hunt, J.F.; Gaertig, S.; Goldberg, J.B. Nitrogen redox balance in the cystic fibrosis airway: Effects of anti-pseudomonal therapy. Am. J. Respir. Crit. Care Med. 2002, 165, 387–390. [Google Scholar] [CrossRef] [PubMed]
- Shin, H.W.; Shelley, D.A.; Fitzpatrick, A.; Gaston, B.; George, S.C. Airway nitric oxide is reduced following phosphate buffered saline inhalation in asthma. J. Appl. Physiol. 2007, 102, 1028–1033. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Grasemann, H.; Gaston, B.; Fang, K.; Paul, K.; Ratjen, F. Decreased levels of nitrosothiols in the lower airways of patients with cystic fibrosis and normal pulmonary function. J. Pediatr. 1999, 135, 770. [Google Scholar] [CrossRef]
- Tsoukias, N.M.; Shin, H.W.; Wilson, A.F.; George, S.C. A single-breath technique with variable flow rate to characterize nitric oxide exchange dynamics in the lungs. J. Appl. Physiol. 2001, 91, 477–487. [Google Scholar] [CrossRef] [PubMed]
- Dweik, R.; Comhair, S.; Gaston, B.; Thunnissen, F.; Farver, C.; Thomassen, M.; Kavuru, M.; Hammel, J.; Abu-Soud, H.; Erzurum, S. NO chemical events in the human airway during the immediate and late antigen-induced asthmatic response. Proc. Natl. Acad. Sci. USA 2001, 98, 2622–2627. [Google Scholar] [CrossRef] [Green Version]
- Snyder, A.H.; McPherson, M.E.; Hunt, J.F.; Johnson, M.; Stamler, J.S.; Gaston, B. Acute effects of aerosolized S-nitrosoglutathione in cystic fibrosis. Am. J. Respir. Crit. Care Med. 2002, 165, 922–926. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Davis, M.D.; Walsh, B.K.; Dwyer, S.T.; Combs, C.; Vehse, N.; Paget-Brown, A.; Pajewski, T.; Hunt, J.F. Safety of an alkalinizing buffer designed for inhaled medications in humans. Respir. Care 2013, 58, 1226–1232. [Google Scholar] [CrossRef]
- Shoemark, A.; Wilson, R. Bronchial and peripheral airway nitric oxide in primary ciliary dyskinesia and bronchiectasis. Respir. Med. 2009, 103, 700–706. [Google Scholar] [CrossRef] [Green Version]
- Ho, L.P.; Innes, J.A.; Greening, A.P. Exhaled nitric oxide is not elevated in the inflammatory airways diseases of cystic fibrosis and bronchiectasis. Eur. Respir. J. 1998, 12, 1290–1294. [Google Scholar] [CrossRef] [Green Version]
- DeSanctis, G.T.; MacLean, J.A.; Hamada, K.; Mehta, S.; Scott, J.A.; Jiao, A.; Yandava, C.N.; Kobzik, L.; Wolyniec, W.W.; Fabia, A.J.; et al. Contribution of nitric oxide synthases 1, 2, and 3 to airway hyperresponsiveness and inflammation in a murine model of asthma. J. Exp. Med. 1999, 189, 1621–1630. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Steudel, W.; Ichinose, F.; Huang, P.L.; Hurford, W.E.; Jones, R.C.; Bevan, J.A.; Fishman, M.C.; Zapol, W.M. Pulmonary vasoconstriction and hypertension in mice with targeted disruption of the endothelial nitric oxide synthase (NOS 3) gene. Circ. Res. 1997, 81, 34–41. [Google Scholar] [CrossRef] [PubMed]
- Marozkina, N.; Gaston, B. An Update on Thiol Signaling: S-Nitrosothiols, Hydrogen Sulfide and a Putative Role for Thionitrous Acid. Antioxidants 2020, 9, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Liu, L.; Hausladen, A.; Zeng, M.; Que, L.; Heitman, J.; Stamler, J.S. A metabolic enzyme for S-nitrosothiol conserved from bacteria to humans. Nature 2001, 410, 490–494. [Google Scholar] [CrossRef]
- Gaston, B.; Kelly, R.; Urban, P.; Liu, L.; Henderson, E.M.; Doctor, A.; Teague, W.G.; Fitzpatrick, A.; Erzurum, S.; Hunt, J.F. Buffering airway acid decreases exhaled nitric oxide in asthma. J. Allergy Clin. Immunol. 2006, 118, 817–822. [Google Scholar] [CrossRef]
- American Thoracic Society. ATS/ERS recommendations for standardized procedures for the online and offline measurement of exhaled lower respiratory nitric oxide and nasal nitric oxide. Am. J. Respir. Crit. Care Med. 2005, 171, 912–930. [Google Scholar] [CrossRef]
- Que, L.G.; Liu, L.; Yan, Y.; Whitehead, G.S.; Gavett, S.H.; Schwartz, D.A.; Stamler, J.S. Protection from experimental asthma by an endogenous bronchodilator. Science 2005, 308, 1618–1621. [Google Scholar] [CrossRef] [Green Version]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Pediatric Participant | Age | Sex | Underlying Diagnosis/es | [NO] (nM) | [SNO] (uM) |
|---|---|---|---|---|---|
| 1 | 2 y/o | Female | Urosepsis | 0.07 | 3.4 |
| 2 | 6 m/o | Male | Enterococcal sepsis | 0.05 | 0.11 |
| 3 | 3 y/o | Male | Aspiration pneumonia | 0.058 | 0.32 |
| 4 | 9 m/o | Male | Pneumocystis jirovecii pneumonia | 0.06 | 0.15 |
| 5 | 12 y/o | Female | Pseudomonas aeruginosa sepsis | 0.015 | 2.1 |
| 6 | 5 m/o | Female | Pseudomembranous colitis, C. difficile, Klebsiella sepsis | 0.02 | 0.03 |
| 7 | 5 y/o | Female | Candida sepsis, Acute Lymphoblastic leukaemia), Pulmonary hemorrhage | 0.034 | 0.076 |
| 8 | 1 y/o | Male | Community acquired pneumonia/ARDS | 0.038 | 0.13 |
| Adult Participant | Gender | Age (Years) | Smoking Status | Passive Smoke Exposure during Childhood | Current Exposure to Environmental Smoke | Baseline Pulse Oximetry (% sat) | Baseline FENO (ppb) | Peak FENO (Post- GSNO, ppb) |
|---|---|---|---|---|---|---|---|---|
| 1 | Female | 26 | Non-smoker | No | None | 98 | 16 | 28 |
| 2 | Female | 27 | Non-smoker | Yes | None | 99 | 5 | 43 |
| 3 | Female | 19 | Non-smoker | No | None | 97 | 5 | 20 |
| 4 | Female | 19 | Non-smoker | Yes | None | 100 | 17 | 19 |
| 5 | Female | 21 | Non-smoker | No | None | 100 | 11 | 31 |
| 6 | Female | 26 | Non-smoker | No | None | 96 | 50 | 60 |
| 7 | Male | 23 | Non-smoker | No | None | 98 | 19 | 38 |
| 8 | Male | 19 | Non-smoker | No | None | 100 | 59 | 83 |
| Participant | AUC (ppb*min−1) | t1/2 (min) |
|---|---|---|
| 1 | 400 | 34 |
| 2 | 1135 | 32 |
| 3 | 460 | 33 |
| 4 | 70 | 20 |
| 5 | 935 | >60 |
| 6 | 110 | 16 |
| 7 | 445 | 29 |
| 8 | 1030 | 58 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Pophal, M.; Grimmett, Z.W.; Chu, C.; Margevicius, S.; Raffay, T.; Ross, K.; Jafri, A.; Giddings, O.; Stamler, J.S.; Gaston, B.; et al. Airway Thiol-NO Adducts as Determinants of Exhaled NO. Antioxidants 2021, 10, 1527. https://doi.org/10.3390/antiox10101527
Pophal M, Grimmett ZW, Chu C, Margevicius S, Raffay T, Ross K, Jafri A, Giddings O, Stamler JS, Gaston B, et al. Airway Thiol-NO Adducts as Determinants of Exhaled NO. Antioxidants. 2021; 10(10):1527. https://doi.org/10.3390/antiox10101527
Chicago/Turabian StylePophal, Megan, Zachary W. Grimmett, Clara Chu, Seunghee Margevicius, Thomas Raffay, Kristie Ross, Anjum Jafri, Olivia Giddings, Jonathan S. Stamler, Benjamin Gaston, and et al. 2021. "Airway Thiol-NO Adducts as Determinants of Exhaled NO" Antioxidants 10, no. 10: 1527. https://doi.org/10.3390/antiox10101527