
Profiling Dopamine-Induced Oxidized Proteoforms of β-synuclein by Top-Down Mass Spectrometry

 ,
,  , , , ,
, , , ,
Abstract
:1. Introduction
2. Materials and Methods
2.1. Chemicals and Proteins
2.2. BS Incubation and MS Analysis
2.3. Relative Quantitation of Oxidized Proteoforms
3. Results
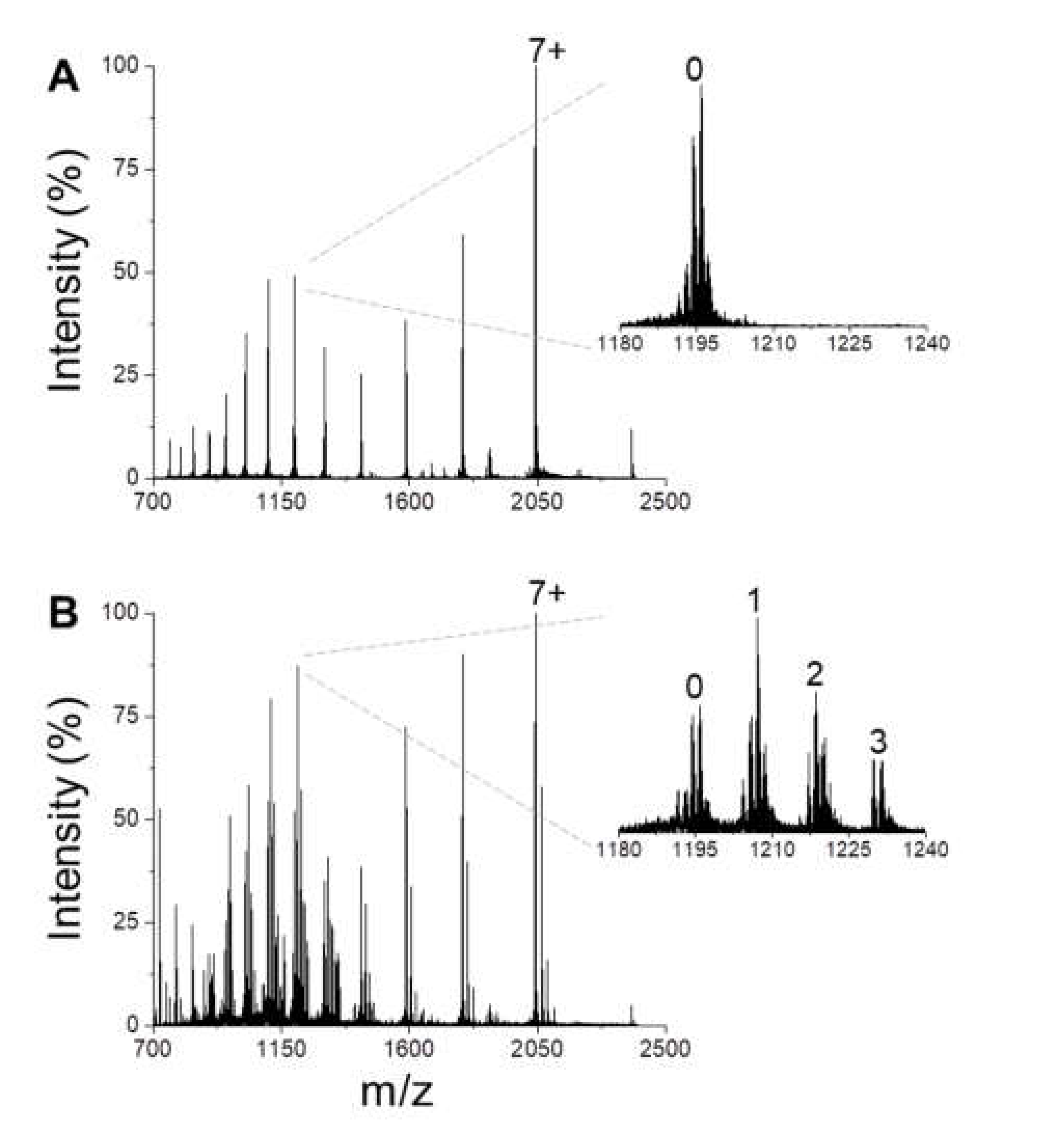
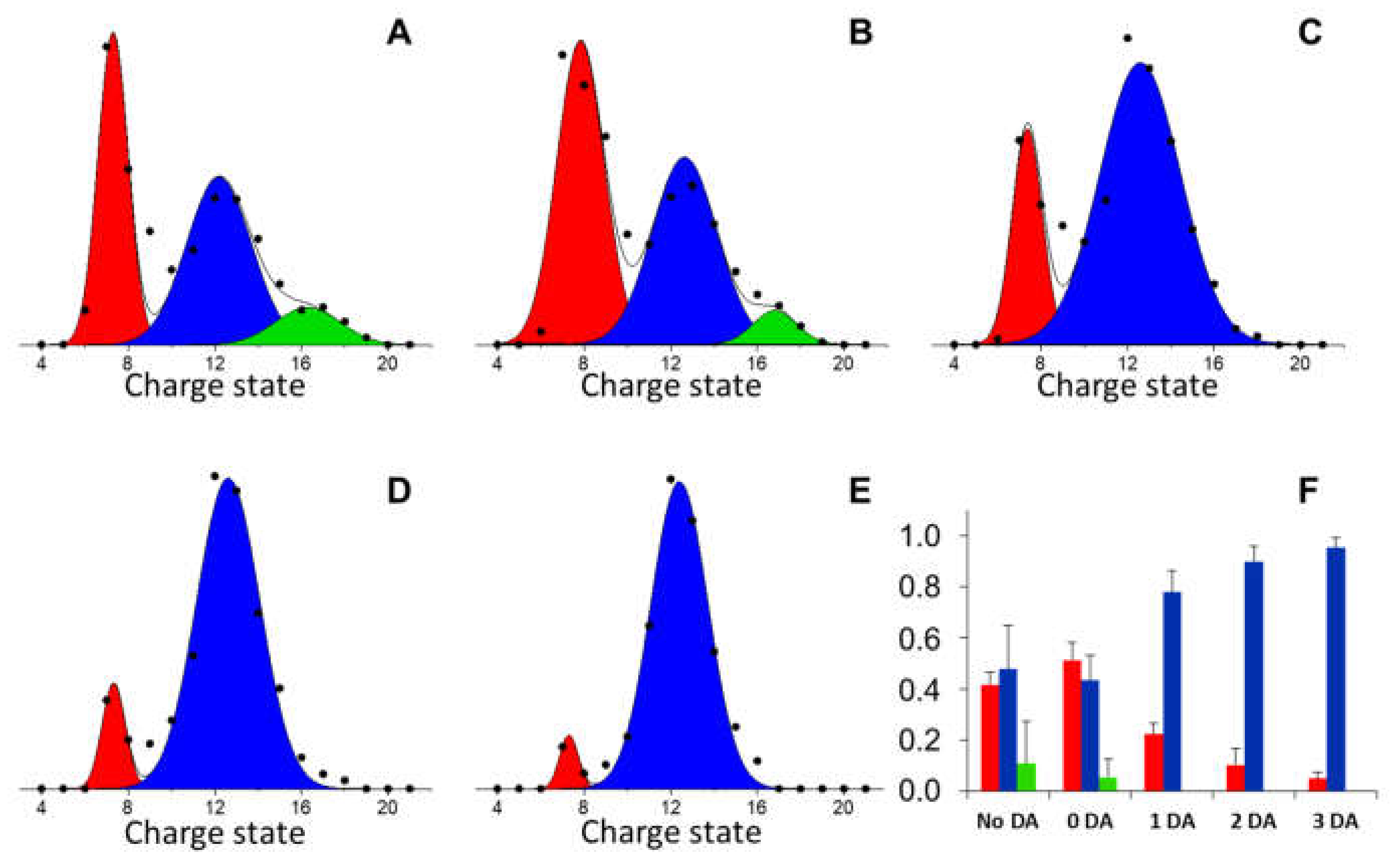
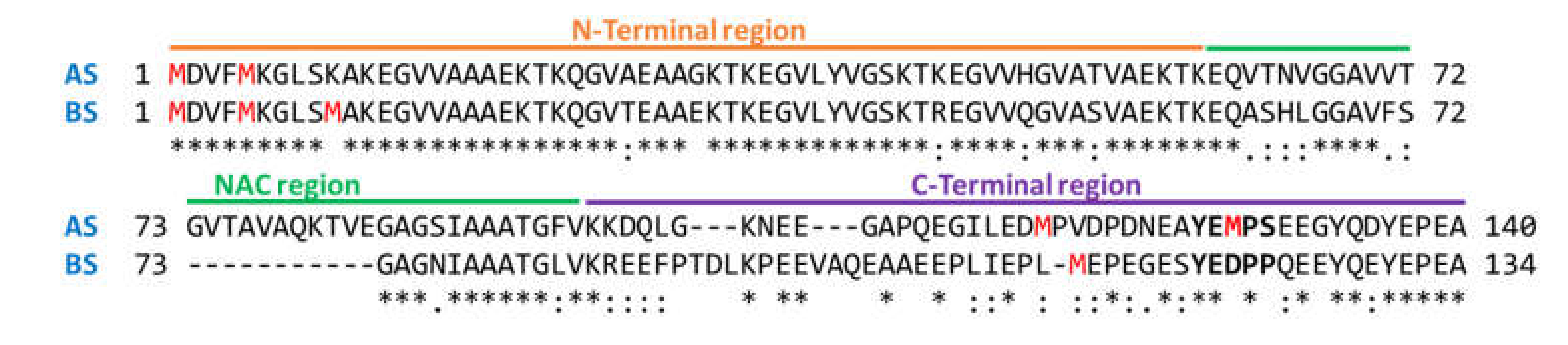
3.1. BS Conformational Ensemble and DA-Interaction
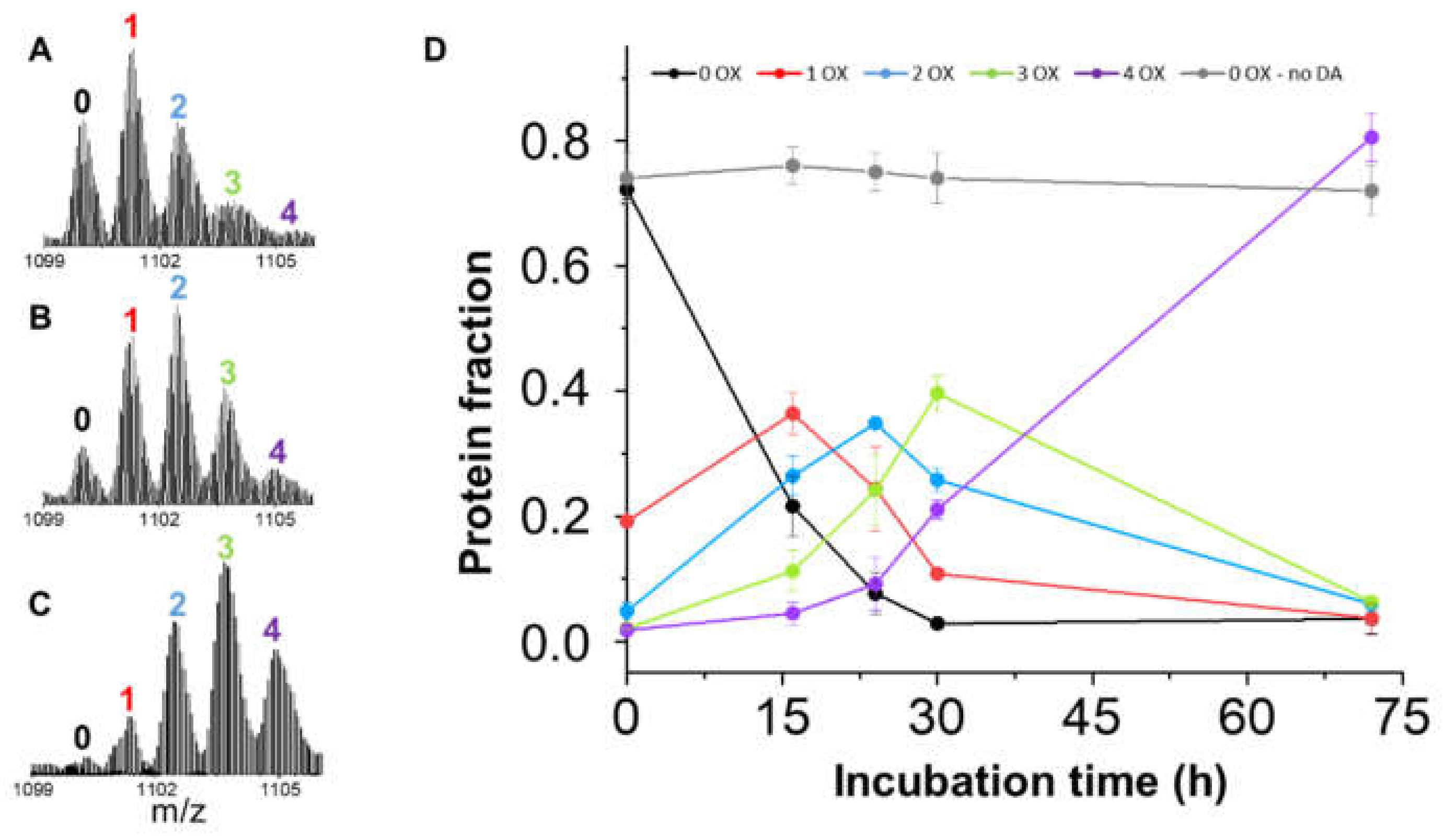
3.2. BS Oxidation Kinetics Induced by DA
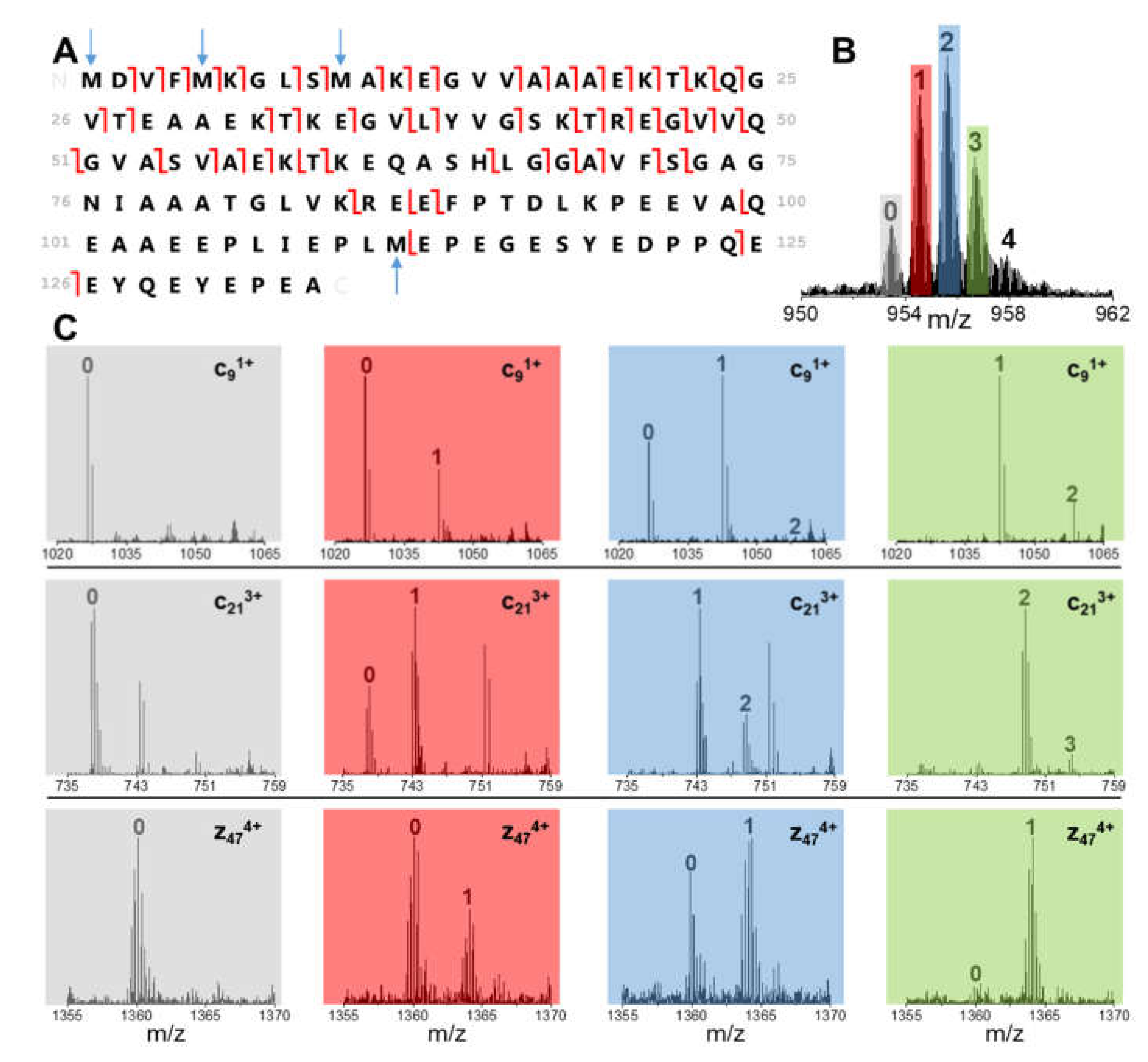
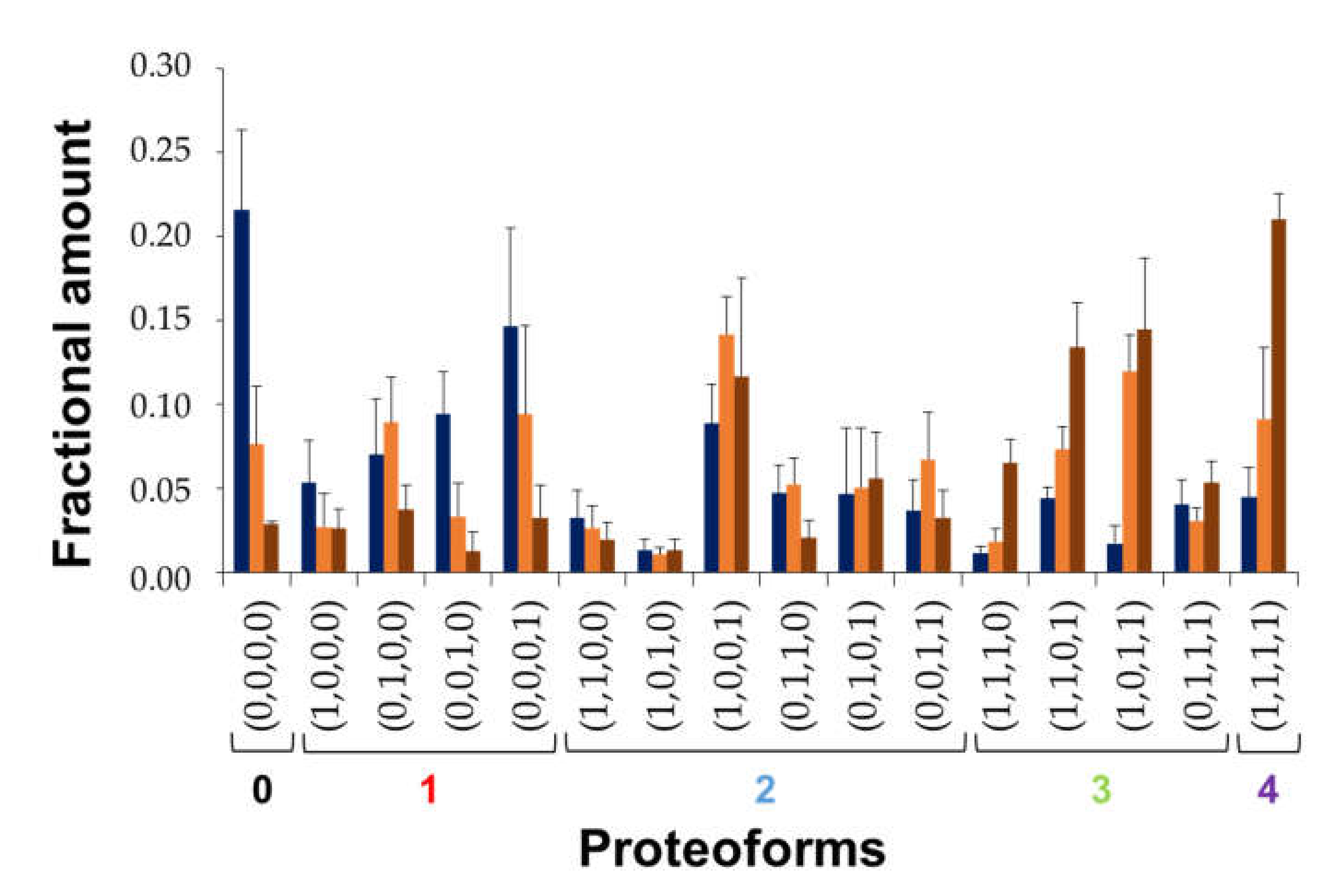
3.3. Quantitation of BS Oxidized Proteoforms
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
Appendix A
- OE = 0
- OE = 1
- OE = 2
- OE = 3
- OE = 4
References
- Nicklow, E.E.; Sevier, C.S. Activity of the Yeast Cytoplasmic Hsp70 Nucleotide-Exchange Factor Fes1 Is Regulated by Reversible Methionine Oxidation. J. Biol. Chem. 2020, 295, 552–569. [Google Scholar] [CrossRef] [PubMed]
- Kaya, A.; Lee, B.C.; Gladyshev, V.N. Regulation of Protein Function by Reversible Methionine Oxidation and the Role of Selenoprotein MsrB1. Antioxid. Redox. Signal. 2015, 23, 814–822. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.C.; Péterfi, Z.; Hoffmann, F.W.; Moore, R.E.; Kaya, A.; Avanesov, A.; Tarrago, L.; Zhou, Y.; Weerapana, E.; Fomenko, D.E.; et al. MsrB1 and MICALs Regulate Actin Assembly and Macrophage Function via Reversible Stereoselective Methionine Oxidation. Mol. Cell 2013, 51, 397–404. [Google Scholar] [CrossRef] [Green Version]
- Lee, H.-J.; Park, J.S.; Yoo, H.J.; Lee, H.M.; Lee, B.C.; Kim, J.H. The Selenoprotein MsrB1 Instructs Dendritic Cells to Induce T-Helper 1 Immune Responses. Antioxidants 2020, 9, 1021. [Google Scholar] [CrossRef]
- Martínez, Y.; Li, X.; Liu, G.; Bin, P.; Yan, W.; Más, D.; Valdivié, M.; Hu, C.-A.A.; Ren, W.; Yin, Y. The Role of Methionine on Metabolism, Oxidative Stress, and Diseases. Amino Acids 2017, 49, 2091–2098. [Google Scholar] [CrossRef]
- Schöneich, C. Methionine Oxidation by Reactive Oxygen Species: Reaction Mechanisms and Relevance to Alzheimer’s Disease. Biochim. Biophys. Acta 2005, 1703, 111–119. [Google Scholar] [CrossRef]
- Reiterer, M.; Schmidt-Kastner, R.; Milton, S.L. Methionine Sulfoxide Reductase (Msr) Dysfunction in Human Brain Disease. Free Radic. Res. 2019, 53, 1144–1154. [Google Scholar] [CrossRef] [PubMed]
- Bettinger, J.; Ghaemmaghami, S. Methionine Oxidation within the Prion Protein. Prion 2020, 14, 193–205. [Google Scholar] [CrossRef]
- Brás, I.C.; Dominguez-Meijide, A.; Gerhardt, E.; Koss, D.; Lázaro, D.F.; Santos, P.I.; Vasili, E.; Xylaki, M.; Outeiro, T.F. Synucleinopathies: Where We Are and Where We Need to Go. J. Neurochem. 2020, 153, 433–454. [Google Scholar] [CrossRef] [Green Version]
- Jain, M.K.; Singh, P.; Roy, S.; Bhat, R. Comparative Analysis of the Conformation, Aggregation, Interaction, and Fibril Morphologies of Human α-, β-, and γ-Synuclein Proteins. Biochemistry 2018, 57, 3830–3848. [Google Scholar] [CrossRef]
- Leong, S.L.; Pham, C.L.L.; Galatis, D.; Fodero-Tavoletti, M.T.; Perez, K.; Hill, A.F.; Masters, C.L.; Ali, F.E.; Barnham, K.J.; Cappai, R. Formation of Dopamine-Mediated Alpha-Synuclein-Soluble Oligomers Requires Methionine Oxidation. Free Radic. Biol. Med. 2009, 46, 1328–1337. [Google Scholar] [CrossRef] [PubMed]
- Surgucheva, I.; Sharov, V.S.; Surguchov, A. γ-Synuclein: Seeding of α-Synuclein Aggregation and Transmission between Cells. Biochemistry 2012, 51, 4743–4754. [Google Scholar] [CrossRef]
- Nakaso, K.; Tajima, N.; Ito, S.; Teraoka, M.; Yamashita, A.; Horikoshi, Y.; Kikuchi, D.; Mochida, S.; Nakashima, K.; Matsura, T. Dopamine-Mediated Oxidation of Methionine 127 in α-Synuclein Causes Cytotoxicity and Oligomerization of α-Synuclein. PLoS ONE 2013, 8, e55068. [Google Scholar] [CrossRef] [PubMed]
- Surgucheva, I.; Newell, K.L.; Burns, J.; Surguchov, A. New α- and γ-Synuclein Immunopathological Lesions in Human Brain. Acta Neuropathol. Commun. 2014, 2, 132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Alam, P.; Bousset, L.; Melki, R.; Otzen, D.E. α-Synuclein Oligomers and Fibrils: A Spectrum of Species, a Spectrum of Toxicities. J. Neurochem. 2019, 150, 522–534. [Google Scholar] [CrossRef] [Green Version]
- Konijnenberg, A.; Ranica, S.; Narkiewicz, J.; Legname, G.; Grandori, R.; Sobott, F.; Natalello, A. Opposite Structural Effects of Epigallocatechin-3-Gallate and Dopamine Binding to α-Synuclein. Anal. Chem. 2016, 88, 8468–8475. [Google Scholar] [CrossRef]
- Ponzini, E.; De Palma, A.; Cerboni, L.; Natalello, A.; Rossi, R.; Moons, R.; Konijnenberg, A.; Narkiewicz, J.; Legname, G.; Sobott, F.; et al. Methionine Oxidation in α-Synuclein Inhibits Its Propensity for Ordered Secondary Structure. J. Biol. Chem. 2019, 294, 5657–5665. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Park, J.-Y.; Lansbury, P.T. Beta-Synuclein Inhibits Formation of Alpha-Synuclein Protofibrils: A Possible Therapeutic Strategy against Parkinson’s Disease. Biochemistry 2003, 42, 3696–3700. [Google Scholar] [CrossRef]
- Snyder, H.; Mensah, K.; Hsu, C.; Hashimoto, M.; Surgucheva, I.G.; Festoff, B.; Surguchov, A.; Masliah, E.; Matouschek, A.; Wolozin, B. Beta-Synuclein Reduces Proteasomal Inhibition by Alpha-Synuclein but Not Gamma-Synuclein. J. Biol. Chem. 2005, 280, 7562–7569. [Google Scholar] [CrossRef] [Green Version]
- Tsigelny, I.F.; Bar-On, P.; Sharikov, Y.; Crews, L.; Hashimoto, M.; Miller, M.A.; Keller, S.H.; Platoshyn, O.; Yuan, J.X.-J.; Masliah, E. Dynamics of Alpha-Synuclein Aggregation and Inhibition of Pore-like Oligomer Development by Beta-Synuclein. FEBS J. 2007, 274, 1862–1877. [Google Scholar] [CrossRef]
- Taschenberger, G.; Toloe, J.; Tereshchenko, J.; Akerboom, J.; Wales, P.; Benz, R.; Becker, S.; Outeiro, T.F.; Looger, L.L.; Bähr, M.; et al. β-Synuclein Aggregates and Induces Neurodegeneration in Dopaminergic Neurons. Ann. Neurol. 2013, 74, 109–118. [Google Scholar] [CrossRef]
- Tolö, J.; Taschenberger, G.; Leite, K.; Stahlberg, M.A.; Spehlbrink, G.; Kues, J.; Munari, F.; Capaldi, S.; Becker, S.; Zweckstetter, M.; et al. Pathophysiological Consequences of Neuronal α-Synuclein Overexpression: Impacts on Ion Homeostasis, Stress Signaling, Mitochondrial Integrity, and Electrical Activity. Front. Mol. Neurosci. 2018, 11, 49. [Google Scholar] [CrossRef] [Green Version]
- Raina, A.; Leite, K.; Guerin, S.; Mahajani, S.U.; Chakrabarti, K.S.; Voll, D.; Becker, S.; Griesinger, C.; Bähr, M.; Kügler, S. Dopamine Promotes the Neurodegenerative Potential of β-Synuclein. J. Neurochem. 2021, 156, 674–691. [Google Scholar] [CrossRef]
- Ohtake, H.; Limprasert, P.; Fan, Y.; Onodera, O.; Kakita, A.; Takahashi, H.; Bonner, L.T.; Tsuang, D.W.; Murray, I.V.J.; Lee, V.M.-Y.; et al. Beta-Synuclein Gene Alterations in Dementia with Lewy Bodies. Neurology 2004, 63, 805–811. [Google Scholar] [CrossRef]
- Sharma, K.; Mehra, S.; Sawner, A.S.; Markam, P.S.; Panigrahi, R.; Navalkar, A.; Chatterjee, D.; Kumar, R.; Kadu, P.; Patel, K.; et al. Effect of Disease-Associated P123H and V70M Mutations on β-Synuclein Fibrillation. ACS Chem. Neurosci. 2020, 11, 2836–2848. [Google Scholar] [CrossRef]
- Ghesquière, B.; Gevaert, K. Proteomics Methods to Study Methionine Oxidation. Mass Spectrom. Rev. 2014, 33, 147–156. [Google Scholar] [CrossRef]
- Kassmann, M.; Hansel, A.; Leipold, E.; Birkenbeil, J.; Lu, S.-Q.; Hoshi, T.; Heinemann, S.H. Oxidation of Multiple Methionine Residues Impairs Rapid Sodium Channel Inactivation. Pflug. Arch. 2008, 456, 1085–1095. [Google Scholar] [CrossRef] [Green Version]
- Fricke, T.C.; Echtermeyer, F.; Zielke, J.; De la Roche, J.; Filipovic, M.R.; Claverol, S.; Herzog, C.; Tominaga, M.; Pumroy, R.A.; Moiseenkova-Bell, V.Y.; et al. Oxidation of Methionine Residues Activates the High-Threshold Heat-Sensitive Ion Channel TRPV2. Proc. Natl. Acad. Sci. USA 2019, 116, 24359–24365. [Google Scholar] [CrossRef]
- Veredas, F.J.; Cantón, F.R.; Aledo, J.C. Methionine Residues around Phosphorylation Sites Are Preferentially Oxidized in Vivo under Stress Conditions. Sci. Rep. 2017, 7, 40403. [Google Scholar] [CrossRef]
- Hardin, S.C.; Larue, C.T.; Oh, M.-H.; Jain, V.; Huber, S.C. Coupling Oxidative Signals to Protein Phosphorylation via Methionine Oxidation in Arabidopsis. Biochem. J. 2009, 422, 305–312. [Google Scholar] [CrossRef] [Green Version]
- Rao, R.S.P.; Møller, I.M.; Thelen, J.J.; Miernyk, J.A. Convergent Signaling Pathways--Interaction between Methionine Oxidation and Serine/Threonine/Tyrosine O-Phosphorylation. Cell Stress Chaperones 2015, 20, 15–21. [Google Scholar] [CrossRef] [Green Version]
- Lanucara, F.; Eyers, C.E. Top-down Mass Spectrometry for the Analysis of Combinatorial Post-Translational Modifications. Mass Spectrom. Rev. 2013, 32, 27–42. [Google Scholar] [CrossRef]
- Olsen, J.V.; Mann, M. Status of Large-Scale Analysis of Post-Translational Modifications by Mass Spectrometry. Mol. Cell. Proteom. 2013, 12, 3444–3452. [Google Scholar] [CrossRef] [Green Version]
- Dang, X.; Singh, A.; Spetman, B.D.; Nolan, K.D.; Isaacs, J.S.; Dennis, J.H.; Dalton, S.; Marshall, A.G.; Young, N.L. Label-Free Relative Quantitation of Isobaric and Isomeric Human Histone H2A and H2B Variants by Fourier Transform Ion Cyclotron Resonance Top-Down MS/MS. J. Proteome Res. 2016, 15, 3196–3203. [Google Scholar] [CrossRef]
- Zheng, Y.; Fornelli, L.; Compton, P.D.; Sharma, S.; Canterbury, J.; Mullen, C.; Zabrouskov, V.; Fellers, R.T.; Thomas, P.M.; Licht, J.D.; et al. Unabridged Analysis of Human Histone H3 by Differential Top-Down Mass Spectrometry Reveals Hypermethylated Proteoforms from MMSET/NSD2 Overexpression. Mol. Cell. Proteom. 2016, 15, 776–790. [Google Scholar] [CrossRef] [Green Version]
- Tucholski, T.; Ge, Y. Fourier-Transform Ion Cyclotron Resonance Mass Spectrometry for Characterizing Proteoforms. Mass Spectrom. Rev. 2020. [Google Scholar] [CrossRef]
- Schaffer, L.V.; Millikin, R.J.; Shortreed, M.R.; Scalf, M.; Smith, L.M. Improving Proteoform Identifications in Complex Systems Through Integration of Bottom-Up and Top-Down Data. J. Proteome Res. 2020, 19, 3510–3517. [Google Scholar] [CrossRef]
- Tadi, S.; Sharp, J.S. Top-Down ETD-MS Provides Unreliable Quantitation of Methionine Oxidation. J. Biomol. Tech. 2019, 30, 50–57. [Google Scholar] [CrossRef]
- Compton, P.D.; Kelleher, N.L.; Gunawardena, J. Estimating the Distribution of Protein Post-Translational Modification States by Mass Spectrometry. J. Proteome Res. 2018, 17, 2727–2734. [Google Scholar] [CrossRef]
- Latawiec, D.; Herrera, F.; Bek, A.; Losasso, V.; Candotti, M.; Benetti, F.; Carlino, E.; Kranjc, A.; Lazzarino, M.; Gustincich, S.; et al. Modulation of Alpha-Synuclein Aggregation by Dopamine Analogs. PLoS ONE 2010, 5, e9234. [Google Scholar] [CrossRef] [Green Version]
- Fellers, R.T.; Greer, J.B.; Early, B.P.; Yu, X.; LeDuc, R.D.; Kelleher, N.L.; Thomas, P.M. ProSight Lite: Graphical Software to Analyze Top-down Mass Spectrometry Data. Proteomics 2015, 15, 1235–1238. [Google Scholar] [CrossRef]
- Santambrogio, C.; Natalello, A.; Brocca, S.; Ponzini, E.; Grandori, R. Conformational Characterization and Classification of Intrinsically Disordered Proteins by Native Mass Spectrometry and Charge-State Distribution Analysis. Proteomics 2019, 19, e1800060. [Google Scholar] [CrossRef]
- Natalello, A.; Santambrogio, C.; Grandori, R. Are Charge-State Distributions a Reliable Tool Describing Molecular Ensembles of Intrinsically Disordered Proteins by Native MS? J. Am. Soc. Mass Spectrom. 2017, 28, 21–28. [Google Scholar] [CrossRef] [PubMed]
- Boys, B.L.; Kuprowski, M.C.; Noël, J.J.; Konermann, L. Protein Oxidative Modifications during Electrospray Ionization: Solution Phase Electrochemistry or Corona Discharge-Induced Radical Attack? Anal. Chem. 2009, 81, 4027–4034. [Google Scholar] [CrossRef] [PubMed]
- Schildknecht, S.; Gerding, H.R.; Karreman, C.; Drescher, M.; Lashuel, H.A.; Outeiro, T.F.; Di Monte, D.A.; Leist, M. Oxidative and Nitrative Alpha-Synuclein Modifications and Proteostatic Stress: Implications for Disease Mechanisms and Interventions in Synucleinopathies. J. Neurochem 2013, 125, 491–511. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Mor, D.E.; Daniels, M.J.; Ischiropoulos, H. The Usual Suspects, Dopamine and Alpha-Synuclein, Conspire to Cause Neurodegeneration. Mov. Disord. 2019, 34, 167–179. [Google Scholar] [CrossRef] [PubMed]
- Herrera, F.E.; Chesi, A.; Paleologou, K.E.; Schmid, A.; Munoz, A.; Vendruscolo, M.; Gustincich, S.; Lashuel, H.A.; Carloni, P. Inhibition of Alpha-Synuclein Fibrillization by Dopamine Is Mediated by Interactions with Five C-Terminal Residues and with E83 in the NAC Region. PLoS ONE 2008, 3, e3394. [Google Scholar] [CrossRef]
- Sievers, F.; Higgins, D.G. Clustal Omega for Making Accurate Alignments of Many Protein Sequences. Protein Sci. 2018, 27, 135–145. [Google Scholar] [CrossRef] [Green Version]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Oxidation Extent (OE) | Nomenclature | Met Residues | MetO Residues |
|---|---|---|---|
| 0 | (0,0,0,0) | 1, 5, 10, 112 | - |
| 1 | (1,0,0,0) | 5, 10, 112 | 1 |
| (0,1,0,0) | 1, 10, 112 | 5 | |
| (0,0,1,0) | 1, 5, 112 | 10 | |
| (0,0,0,1) | 1, 5, 10 | 112 | |
| 2 | (1,1,0,0) | 10, 112 | 1, 5 |
| (1,0,1,0) | 5, 112 | 1, 10 | |
| (1,0,0,1) | 5, 10 | 1, 112 | |
| (0,1,1,0) | 1, 112 | 5, 10 | |
| (0,1,0,1) | 1, 10 | 5, 112 | |
| (0,0,1,1) | 1, 5 | 10, 112 | |
| 3 | (1,1,1,0) | 112 | 1, 5, 10 |
| (1,1,0,1) | 10 | 1, 5, 112 | |
| (1,0,1,1) | 5 | 1, 10, 112 | |
| (0,1,1,1) | 1 | 5, 10, 112 | |
| 4 | (1,1,1,1) | - | 1, 5, 10, 112 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Luise, A.; De Cecco, E.; Ponzini, E.; Sollazzo, M.; Mauri, P.; Sobott, F.; Legname, G.; Grandori, R.; Santambrogio, C. Profiling Dopamine-Induced Oxidized Proteoforms of β-synuclein by Top-Down Mass Spectrometry. Antioxidants 2021, 10, 893. https://doi.org/10.3390/antiox10060893
Luise A, De Cecco E, Ponzini E, Sollazzo M, Mauri P, Sobott F, Legname G, Grandori R, Santambrogio C. Profiling Dopamine-Induced Oxidized Proteoforms of β-synuclein by Top-Down Mass Spectrometry. Antioxidants. 2021; 10(6):893. https://doi.org/10.3390/antiox10060893
Chicago/Turabian StyleLuise, Arianna, Elena De Cecco, Erika Ponzini, Martina Sollazzo, PierLuigi Mauri, Frank Sobott, Giuseppe Legname, Rita Grandori, and Carlo Santambrogio. 2021. "Profiling Dopamine-Induced Oxidized Proteoforms of β-synuclein by Top-Down Mass Spectrometry" Antioxidants 10, no. 6: 893. https://doi.org/10.3390/antiox10060893