
Assessment of Bioactive Compounds under Simulated Gastrointestinal Digestion of Bee Pollen and Bee Bread: Bioaccessibility and Antioxidant Activity
 ,
,  ,
, 

Abstract
:1. Introduction
2. Materials and Methods
2.1. Standards and Reagents
2.2. Sample Collection and Preparation
2.3. Palynological Analysis
2.4. Phenolic Compounds Extraction
2.5. Phenolic Content
2.5.1. Total Phenolic Content
2.5.2. Total Flavonoid Content
2.6. LC/DAD/ESI-MSn Bioactive Compounds Analysis
2.7. Antioxidant Activity
2.7.1. DPPH Radical Scavenging Assay
2.7.2. Reducing Power Assay
2.8. In Vitro Gastrointestinal Digestion
2.9. Bioaccessibility
2.10. Data Analysis
3. Results
3.1. Botanical Origin of Bee Pollen and Bee Bread
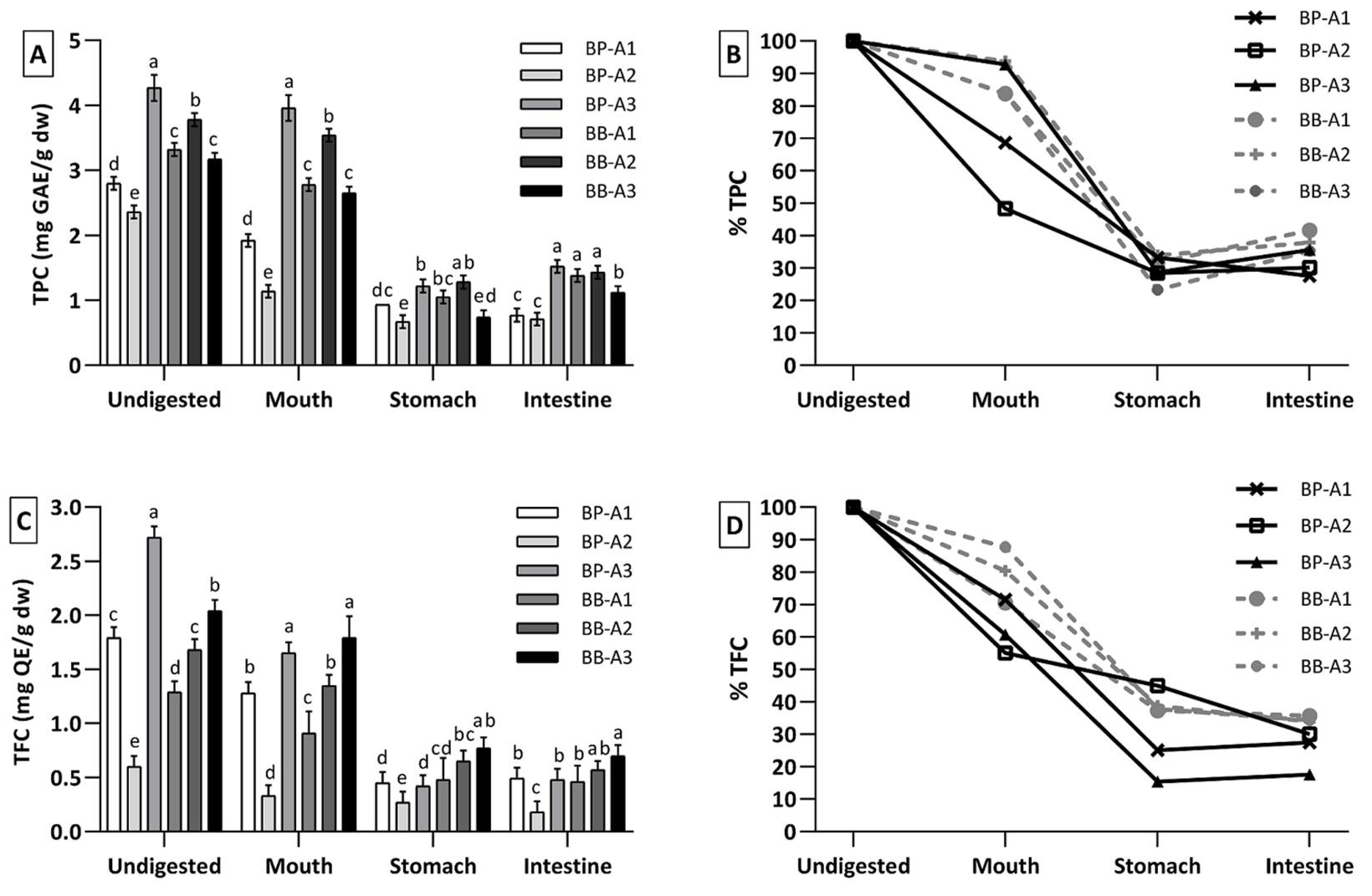
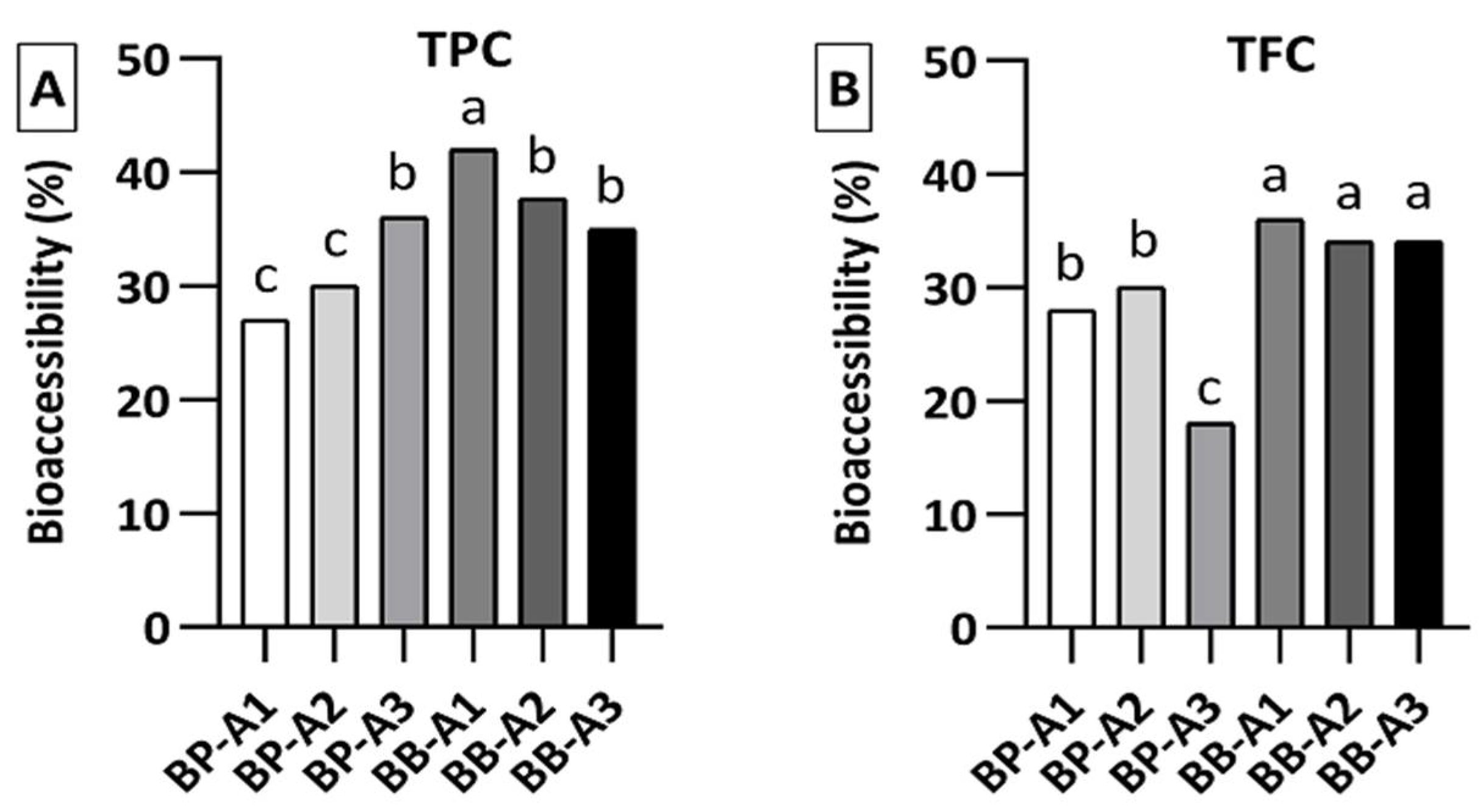
3.2. Total Phenolic and Flavonoid Content, and Bioaccessibility Level
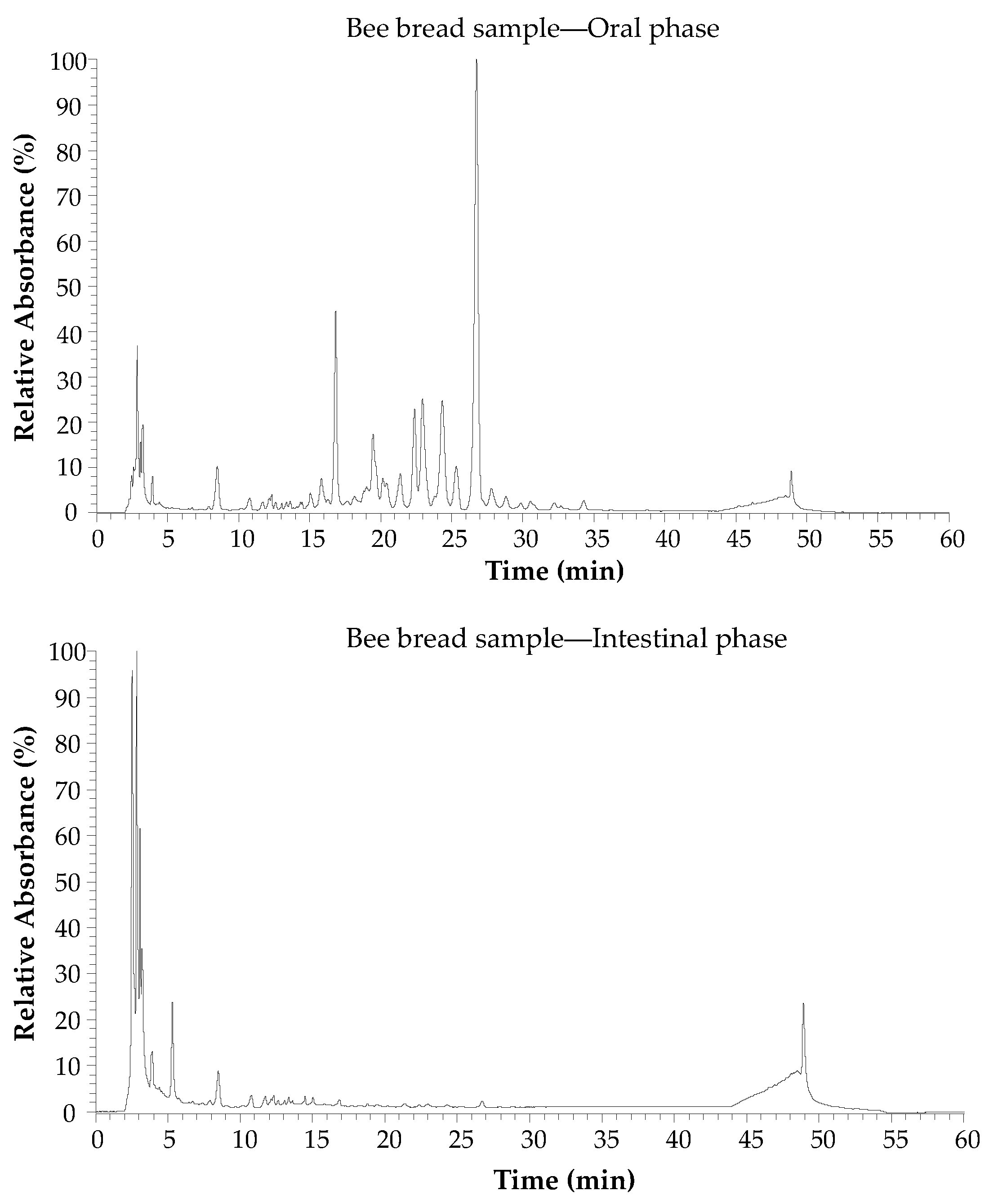
3.3. Phytochemical Profile and Bioaccessibility Level
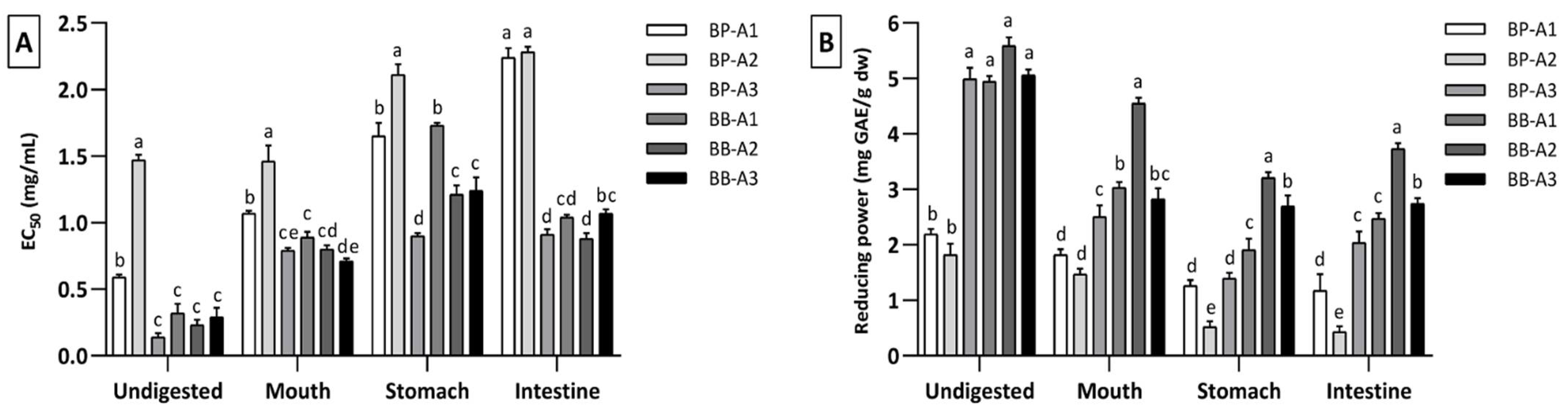
3.4. Antioxidant Capacity
4. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Conflicts of Interest
References
- Kieliszek, M.; Piwowarek, K.; Kot, A.M.; Błażejak, S.; Chlebowska-Śmigiel, A.; Wolska, I. Pollen and bee bread as new health-oriented products: A review. Trends Food Sci. Technol. 2018, 71, 170–180. [Google Scholar] [CrossRef]
- Campos, M.G.; Bogdanov, S.; de Almeida-Muradian, L.B.; Szczesna, T.; Mancebo, Y.; Frigerio, C.; Ferreira, F. Pollen composition and standardisation of analytical methods. J. Apic. Res. 2008, 47, 154–161. [Google Scholar] [CrossRef]
- Feás, X.; Vázquez-Tato, M.P.; Estevinho, L.; Seijas, J.A.; Iglesias, A. Organic bee pollen: Botanical origin, nutritional value, bioactive compounds, antioxidant activity and microbiological quality. Molecules 2012, 17, 8359–8377. [Google Scholar] [CrossRef] [PubMed]
- Tomás, A.; Falcão, S.I.; Russo-Almeida, P.; Vilas-Boas, M. Potentialities of beebread as a food supplement and source of nutraceuticals: Botanical origin, nutritional composition and antioxidant activity. J. Apic. Res. 2017, 56, 219–230. [Google Scholar] [CrossRef]
- Bakour, M.; Fernandes, Â.; Barros, L.; Sokovic, M.; Ferreira, I.C. Bee bread as a functional product: Chemical composition and bioactive properties. LWT 2019, 109, 276–282. [Google Scholar] [CrossRef] [Green Version]
- Negri, G.; Barreto, L.M.R.C.; Sper, F.L.; Carvalho, C.D.; Campos, M.D.G.R. Phytochemical analysis and botanical origin of Apis mellifera bee pollen from the municipality of Canavieiras, Bahia State, Brazil. Braz. J. Food Technol. 2018, 21, e2016176. [Google Scholar] [CrossRef] [Green Version]
- Estevinho, L.M.; Rodrigues, S.; Pereira, A.P.; Feás, X. Portuguese bee pollen: Palynological study, nutritional and microbiological evaluation. Int. J. Food Sci. Technol. 2012, 47, 429–435. [Google Scholar] [CrossRef]
- Kaškonienė, V.; Adaškevičiūtė, V.; Kaškonas, P.; Mickienė, R.; Maruška, A. Antimicrobial and antioxidant activities of natural and fermented bee pollen. Food Biosci. 2020, 34, 100532. [Google Scholar] [CrossRef]
- El Ghouizi, A.; El Menyiy, N.; Falcão, S.I.; Vilas-Boas, M.; Lyoussi, B. Chemical composition, antioxidant activity, and diuretic effect of Moroccan fresh bee pollen in rats. Vet. World 2020, 13, 1251–1261. [Google Scholar] [CrossRef]
- Sobral, F.; Calhelha, R.C.; Barros, L.; Dueñas, M.; Tomás, A.; Santos-Buelga, C.; Vilas-Boas, M.; Ferreira, I.C. Flavonoid composition and antitumor activity of bee bread collected in northeast Portugal. Molecules 2017, 22, 248. [Google Scholar] [CrossRef] [Green Version]
- Bartkiene, E.; Lele, V.; Sakiene, V.; Zavistanaviciute, P.; Zokaityte, E.; Dauksiene, A.; Jagminas, P.; Klupsaite, D.; Bliznikas, S.; Ruzauskas, M. Variations of the antimicrobial, antioxidant, sensory attributes and biogenic amines content in Lithuania-derived bee products. LWT 2020, 118, 108793. [Google Scholar] [CrossRef]
- Mărgăoan, R.; Stranț, M.; Varadi, A.; Topal, E.; Yücel, B.; Cornea-Cipcigan, M.; Campos, M.G.; Vodnar, D.C. Bee collected pollen and bee bread: Bioactive constituents and health benefits. Antioxidants 2019, 8, 568. [Google Scholar] [CrossRef] [Green Version]
- Yesiltas, B.; Capanoglu, E.; Firatligil-Durmus, E.; Sunay, A.E.; Samanci, T.; Boyacioglu, D. Investigating the in-vitro bioaccessibility of propolis and pollen using a simulated gastrointestinal digestion System. J. Apic. Res. 2014, 53, 101–108. [Google Scholar] [CrossRef] [Green Version]
- Benavides-Guevara, R.M.; Quicazan, M.C.; Ramírez-Toro, C. Digestibility and availability of nutrients in bee pollen applying different pretreatments. Ing. Compet. 2017, 19, 119–128. [Google Scholar]
- Louveaux, J.; Maurizio, A.; Vorwohl, G. Methods of Melissopalynology. Bee World 1978, 59, 139–157. [Google Scholar] [CrossRef]
- Vergeron, P. Interprétation statistique des résultats en matière d’analyse pollinique des miels. Ann. l’Abeille 1964, 7, 349–364. [Google Scholar] [CrossRef]
- Falcão, S.I.; Freire, C.; Vilas-Boas, M. A proposal for physicochemical standards and antioxidant activity of Portuguese propolis. J. Am. Oil Chem. Soc. 2013, 90, 1729–1741. [Google Scholar] [CrossRef]
- Minekus, M.; Alminger, M.; Alvito, P.; Ballance, S.; Bohn, T.; Bourlieu, C.; Carriere, F.; Boutrou, R.; Corredig, M.; Dupont, D. A standardised static in vitro digestion method suitable for food–an international consensus. Food Funct. 2014, 5, 1113–1124. [Google Scholar] [CrossRef] [Green Version]
- Schulz, M.; Biluca, F.C.; Gonzaga, L.V.; Borges, G.D.S.C.; Vitali, L.; Micke, G.A.; de Gois, J.S.; de Almeida, T.S.; Borges, D.L.G.; Miller, P.R.M. Bioaccessibility of bioactive compounds and antioxidant potential of juçara fruits (Euterpe edulis Martius) subjected to in vitro gastrointestinal digestion. Food Chem. 2017, 228, 447–454. [Google Scholar] [CrossRef]
- Bouayed, J.; Hoffmann, L.; Bohn, T. Total phenolics, flavonoids, anthocyanins and antioxidant activity following simulated gastro-intestinal digestion and dialysis of apple varieties: Bioaccessibility and potential uptake. Food Chem. 2011, 128, 14–21. [Google Scholar] [CrossRef]
- Ng, Z.X.; See, A.N. Effect of in vitro digestion on the total polyphenol and flavonoid, antioxidant activity and carbohydrate hydrolyzing enzymes inhibitory potential of selected functional plant-based foods. J. Food Process. Preserv. 2019, 43, e13903. [Google Scholar] [CrossRef]
- Kamiloglu, S.; Pasli, A.A.; Ozcelik, B.; van Camp, J.; Capanoglu, E. Influence of different processing and storage conditions on in vitro bioaccessibility of polyphenols in black carrot jams and marmalades. Food Chem. 2015, 186, 74–82. [Google Scholar] [CrossRef]
- Quan, W.; Qie, X.; Chen, Y.; Zeng, M.; Qin, F.; Chen, J.; He, Z. Effect of milk addition and processing on the antioxidant capacity and phenolic bioaccessibility of coffee by using an in vitro gastrointestinal digestion model. Food Chem. 2020, 308, 125598. [Google Scholar] [CrossRef] [PubMed]
- Pinto, J.; Spínola, V.; Llorent-Martínez, E.J.; Fernández-de Córdova, M.L.; Molina-García, L.; Castilho, P.C. Polyphenolic profile and antioxidant activities of Madeiran elderberry (Sambucus lanceolata) as affected by simulated in vitro digestion. Food Res. Int. 2017, 100, 404–410. [Google Scholar] [CrossRef]
- Ng, Z.X.; Rosman, N.F. In vitro digestion and domestic cooking improved the total antioxidant activity and carbohydrate-digestive enzymes inhibitory potential of selected edible mushrooms. J. Food Sci. Technol. 2019, 56, 865–877. [Google Scholar] [CrossRef]
- Ortega, N.; Macià, A.; Romero, M.-P.; Reguant, J.; Motilva, M.-J. Matrix composition effect on the digestibility of carob flour phenols by an in-vitro digestion model. Food Chem. 2011, 124, 65–71. [Google Scholar] [CrossRef]
- Falcão, S.I.; Vale, N.; Gomes, P.; Domingues, M.R.; Freire, C.; Cardoso, S.M.; Vilas-Boas, M. Phenolic profiling of Portuguese propolis by LC–MS spectrometry: Uncommon propolis rich in flavonoid glycosides. Phytochem. Anal. 2013, 24, 309–318. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Su, J.; Yang, X.; Lu, Q.; Liu, R. Antioxidant and anti-tyrosinase activities of bee pollen and identification of active components. J. Apic. Res. 2020, 1–11. [Google Scholar] [CrossRef]
- Mihajlovic, L.; Radosavljevic, J.; Burazer, L.; Smiljanic, K.; Velickovic, T.C. Composition of polyphenol and polyamide compounds in common ragweed (Ambrosia artemisiifolia L.) pollen and sub-pollen particles. Phytochemistry 2015, 109, 125–132. [Google Scholar] [CrossRef] [PubMed]
- Sobolev, V.S.; Sy, A.A.; Gloer, J.B. Spermidine and flavonoid conjugates from peanut (Arachis hypogaea) flowers. J. Agric. Food Chem. 2008, 56, 2960–2969. [Google Scholar] [CrossRef] [PubMed]
- Cuyckens, F.; Claeys, M. Mass spectrometry in the structural analysis of flavonoids. J. Mass Spectrom. 2004, 39, 1–15. [Google Scholar] [CrossRef] [PubMed]
- Barros, L.; Dueñas, M.; Carvalho, A.M.; Ferreira, I.C.; Santos-Buelga, C. Characterization of phenolic compounds in flowers of wild medicinal plants from Northeastern Portugal. Food Chem. Toxicol. 2012, 50, 1576–1582. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Paupière, M.J.; Müller, F.; Li, H.; Rieu, I.; Tikunov, Y.M.; Visser, R.G.; Bovy, A.G. Untargeted metabolomic analysis of tomato pollen development and heat stress response. Plant Reprod. 2017, 30, 81–94. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Anjos, O.; Fernandes, R.; Cardoso, S.M.; Delgado, T.; Farinha, N.; Paula, V.; Estevinho, L.M.; Carpes, S.T. Bee pollen as a natural antioxidant source to prevent lipid oxidation in black pudding. LWT 2019, 111, 869–875. [Google Scholar] [CrossRef] [Green Version]
- Urcan, A.C.; Criste, A.D.; Dezmirean, D.S.; Mărgăoan, R.; Caeiro, A.; Graça Campos, M. Similarity of data from bee bread with the same taxa collected in India and Romania. Molecules 2018, 23, 2491. [Google Scholar] [CrossRef] [Green Version]
- Edreva, A.; Velikova, V.; Tsonev, T. Phenylamides in plants. Russ. J. Plant Physiol. 2007, 54, 287–301. [Google Scholar] [CrossRef]
- Eliašová, A.; Poracká, V.; Pal’ove-Balang, P.; Imrich, J.; Repčák, M. Accumulation of tetracoumaroyl spermine in Matricaria chamomilla during floral development and nitrogen deficiency. Z. Nat. C. J. Biosci. 2012, 67, 58–64. [Google Scholar] [CrossRef]
- Lucas-Gonzalez, R.; Navarro-Coves, S.; Pérez-Álvarez, J.A.; Fernández-López, J.; Muñoz, L.A.; Viuda-Martos, M. Assessment of polyphenolic profile stability and changes in the antioxidant potential of maqui berry (Aristotelia chilensis (Molina) Stuntz) during in vitro gastrointestinal digestion. Ind. Crops Prod. 2016, 94, 774–782. [Google Scholar] [CrossRef]
- Toro-Funes, N.; Bosch-Fusté, J.; Veciana-Nogués, M.T.; Izquierdo-Pulido, M.; Vidal-Carou, M.C. In vitro antioxidant activity of dietary polyamines. Food Res. Int. 2013, 51, 141–147. [Google Scholar] [CrossRef]
- De-Melo, A.A.M.; Estevinho, L.M.; Moreira, M.M.; Delerue-Matos, C.; de Freitas, A.D.S.; Barth, O.M.; de Almeida-Muradian, L.B. A multivariate approach based on physicochemical parameters and biological potential for the botanical and geographical discrimination of Brazilian bee pollen. Food Biosci. 2018, 25, 91–110. [Google Scholar] [CrossRef] [Green Version]






{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Relative Frequency (%) of Pollen Types | |||||||
|---|---|---|---|---|---|---|---|
| Family | Pollen Types | BP-A1 a | BP-A2 b | BP-A3 | BB-A1 a | BB-A2 b | BB-A3 |
| Amaranthaceae | Chenopodium sp. | — | 1.60 | — | — | — | 1.36 |
| Apiaceae | Eryngium | 1.02 | — | — | — | — | 0.14 |
| Thapsia vilosa | 0.11 | 6.75 | — | — | — | — | |
| Asteraceae | Centaurea sp. | 6.21 | 1.60 | — | 1.90 | — | — |
| Crepis capillaris | 11.17 | 59.84 | — | 0.76 | 0.80 | 1.64 | |
| Boraginaceae | Echium sp. | 8.80 | 0.57 | 0.72 | 4.19 | 1.50 | — |
| Pentaglotis sempervirens | — | — | — | 1.02 | — | — | |
| Brassicaceae | Raphanus raphanistrum | — | — | 3.79 | 0.38 | 0.10 | 1.91 |
| Campanulaceae | Jasione montana | — | — | — | 0.13 | 3.40 | 21.96 |
| Crassulaceae | Sedum sp. | — | — | 0.21 | 0.51 | 3.30 | 0.82 |
| Ericaceae | Erica sp. | — | — | 9.03 | — | — | — |
| Fabaceae | Lotus sp. | 0.11 | — | — | 1.14 | — | — |
| Cytisus striatus | — | 0.11 | 47.69 | 6.22 | 6.50 | 19.65 | |
| Trifolium sp. | 5.98 | 0.23 | 1.74 | 12.18 | 6.50 | — | |
| Fagaceae | Castanea sativa | — | — | — | 25.38 | 47.50 | 25.51 |
| Quercus sp. | — | 0.34 | — | — | — | 6.96 | |
| Lamiaceae | Lavandula sp. | — | — | — | 0.38 | 1.10 | 3.27 |
| Myrtaceae | Eucalyptus sp. | — | — | 24.10 | — | — | — |
| Papaveraceae | — | 0.34 | 0.23 | 3.49 | 0.25 | 2.20 | 0.55 |
| Plantaginaceae | Plantago sp. | 47.18 | 20.14 | 0.10 | 2.79 | 1.20 | 1.91 |
| Poaceae | Zea mays | 1.81 | — | — | — | — | — |
| Resedaceae | Sesamoides sp. or Reseda sp. | — | — | 1.85 | 0.13 | 0.20 | — |
| Rhamnus alaternus | — | — | 0.10 | 1.02 | — | — | |
| Rosaceae | — | — | — | 1.33 | — | — | 0.68 |
| Rubus sp. | 0.56 | — | 2.67 | 37.31 | 22.30 | 10.23 | |
| Salicaceae | Salix sp. | — | — | 1.33 | 0.51 | 0.80 | — |
| Solanaceae | — | — | 2.06 | — | — | — | — |
| Classification | Multifloral | ||||||
| tR (min) | λmax (nm) | [M-H]- m/z | MSn (% Base Peak) | Proposed Compound | mg/g Raw Sample | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| BP-A1 | BP-A2 | BP-A3 | BB-A1 | BB-A2 | BB-A3 | |||||
| 6.58 | 255, 349 | 771 | MS2: 609 (100); MS3: 301 (100) | Quercetin-O-hexosyl-O-rutinoside a,c | 0.08 ± 0.0 | ND | ND | ND | ND | ND |
| 7.48 | 257, 353 | 625 | MS2: 301 (100), 300 (99), 445 (85), 271 (18) | Quercetin-diglucoside a,e,h, | ND | ND | ND | ND | ND | ND |
| 8.43 | 272, 326sh, 353sh | 639 | MS2:271 (10), 300 (34), 315 (91), 459 (100), 477 (11), 624 (20) | Methyl herbacetin-O-dihexoside a,c,d | 0.04 ± 0.0 | ND | 0.31 ± 0.0 | 0.43 ± 0.0 | 0.33 ± 0.0 | 0.14 ± 0.0 |
| 9.94 | 265, 348 | 609 | MS2: 285 (100), 429 (49) | Kaempferol-O-dihexoside a,d | 0.08 ± 0.0 | ND | ND | ND | ND | ND |
| 10.69 | 272, 326sh, 353sh | 623 | MS2: 299 (61), 300 (38), 314 (100), 315 (69), 459 (86), 477 (19) | Methyl herbacetin-O-rutinoside a,c | ND | ND | ND | 0.09 ± 0.0 | 0.08 ± 0.0 | 0.02 ± 0.0 |
| 11.69 | 255, 353 | 609 | MS2: 315 (100), 314 (47), 459 (51), 300 (20) | Isorhamnetin-O-pentosyl-hexoside a,e | 0.22 ± 0.0 | 0.07 ± 0.0 | ND | 0.06 ± 0.0 | 0.05 ± 0.0 | 0.06 ± 0.0 |
| 12.09 | 265, 348 | 593 | MS2: 284 (100), 285 (73), 429 (93) | Kaempferol-3-O-rutinoside a,b,f | 0.02 ± 0.0 | 0.00 ± 0.0 | 0.03 ± 0.0 | 0.01 ± 0.0 | 0.02 ± 0.0 | 0.01 ± 0.0 |
| 12.62 | 256, 354 | 463 | MS2: 301 (100) | Quercetin-3-O-glucoside a,b,f | 0.01 ± 0.0 | ND | ND | ND | ND | 0.02 ± 0.0 |
| 13.31 | 256, 353 | 549 | MS2: 505 (100); MS3: 301 (100), 300 (28), 463 (26) | Quercetin-O-malonyl hexoside a,j | ND | ND | ND | ND | 0.01 ± 0.0 | 0.07 ± 0.0 |
| 13.61 | 270 | 477 | MS2: 315 (100), 462 (42), 300 (14); MS3: 300 (100) | Methyl herbacetin-3-O-hexoside a,c | ND | ND | ND | 0.04 ± 0.0 | ND | ND |
| 14.06 | 265, 347 | 447 | MS2: 285 (100), 284 (80) | Kaempferol-O-hexoside a,k | ND | ND | ND | ND | 0.03 ± 0.0 | 0.16 ± 0.0 |
| 14.18 | 254, 347 | 447 | MS2: 301 (100) | Quercetin-3-O-rhamnosidea,b,e | ND | ND | 0.81 ± 0.0 | ND | ND | ND |
| 14.31 | 254, 355 | 477 | MS2: 314 (100), 315 (45) | Isorhamnetin-O-hexoside a,k | ND | ND | ND | ND | 0.01 ± 0.0 | 0.01 ± 0.0 |
| 14.44 | 255, 354 | 563 | MS2: 519 (100); MS3: 315 (100) | Isorhamnetin-3-O-malonyl glucoside a,h | ND | ND | 0.06 ± 0.0 | ND | ND | ND |
| 14.73 | 277, 311 | 301 | MS2: 283 (100), 286 (40) | Hesperetin a,b | ND | ND | 0.05 ± 0.0 | ND | ND | ND |
| 14.99 | 265, 345 | 533 | MS2: 489 (100); MS3: 285 (100) | Kaempferol-O-malonyl hexoside a,h | ND | ND | ND | ND | 0.07 ± 0.0 | 0.09 ± 0.0 |
| 15.39 | 299, 308 | 436 | MS2: 316 (100) | Di-p-coumaroylspermidine a,i | 0.72 ± 0.0 | 0.20 ± 0.0 | ND | ND | ND | ND |
| 15.76 | 295, 315 | 630 | MS2: 468 (100), 494 (84), 358 (7); MS3: 332 (100) | N1, N5, N10-tri-caffeoylspermidine a,d,g | ND | ND | ND | 0.20 ± 0.0 | 0.50 ± 0.0 | ND |
| 16.18 | 264, 341 | 431 | MS2: 285 (100) | Kaempferol-3-O-rhamnoside a,c | ND | ND | 0.04 ± 0.0 | ND | ND | ND |
| 16.64 | 255, 354 | 461 | MS2: 314 (100), 315 (77), 299 (39) | Isorhamnetin-O-deoxyhexoside a,c | ND | ND | 0.31 ± 0.0 | ND | ND | ND |
| 16.75 | 296, 319 | 630 | MS2: 468 (100), 494 (86), 358 (7); MS3: 332 (100) | N1, N5, N10-tri-caffeoylspermidine a,d,g | 0.43 ± 0.0 | 0.08 ± 0.0 | ND | 1.53 ± 0.0 | 4.15 ± 0.0 | 0.51 ± 0.0 |
| 17.57 | 293, 314 | 644 | MS2: 358 (11), 482 (11), 508 (100); MS3: 332 (27), 358 (100), 372 (49) | N1-feruloyl-N5, N10-dicaffeoylspermidine a,d,e | ND | ND | ND | ND | 0.04 ± 0.0 | ND |
| 18.09 | 295, 311 | 614 | MS2: 494 (25), 478 (100), 452 (69), 358 (20) | N1-p-coumaroyl-N5, N10-dicaffeoylspermidine a,e | 0.21 ± 0.0 | ND | 0.38 ± 0.0 | 0.16 ± 0.0 | 0.13 ± 0.0 | 0.07 ± 0.0 |
| 18.18 | 299, 308 | 478 | MS2: 358 (100), 332 (12), 145 (5) | N1-acetyl-N5, N10-di-p-coumaroylspermidine a,f | 2.83 ± 0.1 | 0.69 ± 0.0 | ND | ND | ND | ND |
| 18.74 | 295, 311 | 614 | MS2: 478 (100), 468 (20), 452 (68), 342(5) | N1-p-coumaroyl-N5, N10-dicaffeoylspermidine (isomer) a,e | ND | ND | ND | 0.06 ± 0.0 | 0.20 ± 0.0 | ND |
| 19.34 | 295, 311 | 614 | MS2: 494 (24), 478 (100), 452 (76), 358 (22) | N1-p-coumaroyl-N5, N10-dicaffeoylspermidine (isomer) a,e | ND | ND | 1.36 ± 0.0 | 0.76 ± 0.0 | 1.98 ± 0.0 | 0.89 ± 0.0 |
| 20.02 | 295, 318 | 644 | MS2: 358 (8), 482 (75), 508 (100); MS3: 332 (27), 358 (100), 372 (49) | N1-feruloyl-N5, N10-dicaffeoylspermidine (isomer) a,d,g | ND | ND | ND | ND | 0.18 ± 0.0 | ND |
| 20.27 | 295, 310 | 598 | MS2: 478 (46), 462 (100), 452 (46), 342 (14) | N1, N5-di-p-coumaroyl-N10-caffeoylspermidine a,e | ND | ND | ND | 0.22 ± 0.0 | 0.08 ± 0.0 | ND |
| 21.23 | 295, 310 | 582 | MS2: 462 (100), 436 (9), 342 (7) | N1, N5, N10-tri-p-coumaroylspermidine a,e | ND | 0.13 ± 0.0 | ND | 0.69 ± 0.0 | 1.02 ± 0.0 | 0.42 ± 0.0 |
| 21.33 | 254, 268sh, 348 | 285 | MS2: 285 (100) | Luteolin a,b | ND | ND | 1.00 ± 0.0 | ND | ND | ND |
| 22.22 | 294, 309 | 598 | MS2: 462 (100), 478 (39), 452 (34), 342 (14) | N1, N5-di-p-coumaroyl-N10-caffeoylspermidine a,d,g | 0.21 ± 0.0 | 0.48 ± 0.0 | 1.16 ± 0.0 | 1.40 ± 0.0 | 2.12 ± 0.0 | 0.85 ± 0.0 |
| 22.73 | 295, 310 | 582 | MS2: 462 (100), 436 (9), 342 (7) | N1, N5, N10-tri-p-coumaroylspermidine a,e | ND | 0.16 ± 0.0 | ND | 1.82 ± 0.0 | 2.84 ± 0.0 | 1.68 ± 0.0 |
| 22.77 | 295, 310 | 598 | MS2: 342 (13), 452 (32), 462 (100), 478 (37) | N1, N5-di-p-coumaroyl-N10-caffeoylspermidine (isomer) a,e | ND | ND | 2.61 ± 0.1 | ND | ND | ND |
| 24.12 | 295, 310 | 582 | MS2: 462 (100), 436 (9), 342 (6) | N1, N5, N10-tri-p-coumaroylspermidine a,e | 0.14 ± 0.0 | 0.18 ± 0.0 | 0.61 ± 0.0 | 2.13 ± 0.0 | 3.20 ± 0.0 | 0.93 ± 0.0 |
| 25.08 | 295, 310 | 582 | MS2: 462 (100), 436 (9), 342 (7) | N1, N5, N10-tri-p-coumaroylspermidine a,e | ND | ND | 0.20 ± 0.0 | 0.77 ± 0.0 | 1.12 ± 0.1 | 0.34 ± 0.0 |
| 26.47 | 295, 305 | 582 | MS2: 342 (100), 436 (9), 462 (100) | N1, N5, N10-tri-p-coumaroylspermidine (isomer) a,e | 0.55 ± 0.0 | 0.29 ± 0.0 | 6.28 ± 0.0 | 8.47 ± 0.0 | 15.23 ± 0.1 | 5.33 ± 0.0 |
| 26.92 | 270 | 785 | MS2: 665 (100), 545 (14), 639 (13); MS3: 545 (100) | Tetracoumaroyl spermine a,l | ND | 3.34 ± 0.0 | ND | ND | ND | ND |
| 27.52 | 295, 308 | 612 | MS2: 492 (100); MS3: 372 (100), 449 (24) | Feruloyl dicoumaroyl spermidine a,l | ND | ND | 0.93 ± 0.0 | 0.32 ± 0.0 | 0.70 ± 0.0 | ND |
| 27.67 | 271 | 315 | MS2: 300 (100); MS3: 272 (100), 255 (54), 165 (26) | Herbacetin-methyl-ether a | ND | ND | ND | ND | ND | 0.11 ± 0.0 |
| 28.55 | 280, 307sh | 785 | MS2: 665 (100), 545 (13), 639 (13); MS3: 545 (100) | Tetracoumaroyl spermine (isomer) a,l | 0.90 ± 0.0 | 8.25 ± 0.1 | 0.76 ± 0.0 | 0.42 ± 0.0 | 0.31 ± 0.0 | ND |
| 28.77 | 266, 365 | 285 | MS2: 285 (100) | Kaempferol a,b | ND | ND | ND | ND | ND | 0.28 ± 0.0 |
| 29.28 | 277, 310sh | 785 | MS2: 665 (100), 545 (13), 639 (13); MS3: 545 (100) | Tetracoumaroyl spermine (isomer) a,l | ND | 0.83 ± 0.1 | ND | ND | ND | ND |
| 29.57 | 295, 318 | 672 | - | Polyamide derivative a,e | ND | ND | 1.70 ± 0.0 | ND | ND | ND |
| 30.28 | 289, 306sh | 785 | MS2: 665 (100), 545 (13), 639 (13); MS3: 545 (100) | Tetracoumaroyl spermine (isomer) a,l | 0.74 ± 0.0 | 9.11 ± 0.1 | ND | 0.50 ± 0.0 | ND | ND |
| 31.92 | 293, 310 | 785 | MS2: 665 (100), 545 (13), 639 (13); MS3: 545 (100) | Tetracoumaroyl spermine (isomer) a,l | 0.50 ± 0.0 | 6.51 ± 0.1 | ND | 0.47 ± 0.0 | ND | 0.36 ± 0.0 |
| 33.97 | 299, 310 | 785 | MS2: 665 (100), 545 (13), 639 (13); MS3: 545 (100) | Tetracoumaroyl spermine (isomer) a,l | 0.70 ± 0.0 | 8.49 ± 0.1 | ND | 0.86 ± 0.0 | 0.36 ± 0.1 | ND |
| Total amount of phenolic compounds | 0.5 | 0.1 | 2.6 | 0.6 | 0.6 | 1.0 | ||||
| Total amount of phenylamides | 7.9 | 38.7 | 15.9 | 20.8 | 34.2 | 11.4 | ||||
| Compound | Mouth | Stomach | Intestine | ||||||
|---|---|---|---|---|---|---|---|---|---|
| BP-A1 | BP-A2 | BP-A3 | BP-A1 | BP-A2 | BP-A3 | BP-A1 | BP-A2 | BP-A3 | |
| Quercetin-O-hexosyl-O-rutinoside | 0.03 ± 0.0 | ND | ND | ND | ND | ND | ND | ND | ND |
| Quercetin-diglucoside | ND | ND | 0.03 ± 0.0 | ND | ND | 0.04 ± 0.0 | ND | ND | 0.05 ± 0.0 |
| Methyl herbacetin-O-dihexoside | 0.03 ± 0.0 | ND | 0.32 ± 0.0 | ND | ND | 0.17 ± 0.0 | ND | ND | 0.22 ± 0.0 |
| Kaempferol-O-dihexoside | 0.05 ± 0.0 | ND | ND | ND | ND | ND | ND | ND | ND |
| Isorhamnetin-O-pentosyl-hexoside | 0.12 ± 0.0 | 0.06 ± 0.0 | ND | 0.05 ± 0.0 | 0.05 ± 0.0 | ND | 0.09 ± 0.0 | 0.04 ± 0.0 | ND |
| Kaempferol-3-O-rutinoside | 0.01 ± 0.0 | 0.00 ± 0.0 | 0.03 ± 0.0 | ND | ND | 0.01 ± 0.0 | 0.01 ± 0.0 | ND | 0.01 ± 0.0 |
| Quercetin-3-O-glucoside | 0.01 ± 0.0 | ND | ND | ND | ND | ND | 0.01 ± 0.0 | ND | ND |
| Quercetin-3-O-rhamnoside | ND | ND | 0.77 ± 0.0 | ND | ND | 0.15 ± 0.0 | ND | ND | 0.14 ± 0.0 |
| Isorhamnetin-3-O-malonyl glucoside | ND | ND | 0.03 ± 0.0 | ND | ND | 0.01 ± 0.0 | ND | ND | ND |
| Hesperetin | ND | ND | 0.05 ± 0.0 | ND | ND | ND | ND | ND | ND |
| Di-p-coumaroylspermidine | 0.77 ± 0.0 | 0.03 ± 0.0 | ND | ND | ND | ND | ND | ND | ND |
| Kaempferol-3-O-rhamnoside | ND | ND | 0.03 ± 0.0 | ND | ND | ND | ND | ND | ND |
| Isorhamnetin-O-deoxyhexoside | ND | ND | 0.31 ± 0.0 | ND | ND | 0.02 ± 0.0 | ND | ND | 0.02 ± 0.0 |
| N1, N5, N10-tri-caffeoylspermidine | 0.26 ± 0.0 | 0.04 ± 0.0 | ND | ND | ND | ND | ND | ND | ND |
| N1-acetyl-N5, N10-di-p-coumaroylspermidine | 2.01 ± 0.1 | 0.35 ± 0.0 | ND | ND | ND | ND | ND | ND | ND |
| N1-p-coumaroyl-N5, N10-dicaffeoylspermidine | 0.14 ± 0.0 | ND | 0.39 ± 0.0 | ND | ND | ND | ND | ND | ND |
| N1-p-coumaroyl-N5, N10-dicaffeoylspermidine (isomer) | ND | ND | 1.16 ± 0.0 | ND | ND | ND | ND | ND | ND |
| N1, N5-di-p-coumaroyl-N10-caffeoylspermidine | 0.10 ± 0.0 | 0.22 ± 0.0 | 1.17 ± 0.0 | ND | ND | ND | ND | ND | ND |
| N1, N5-di-p-coumaroyl-N10-caffeoylspermidine (isomer) | ND | ND | 1.97 ± 0.1 | ND | ND | ND | ND | ND | ND |
| Luteolin | ND | ND | 0.99 ± 0.0 | ND | ND | ND | ND | ND | ND |
| N1, N5, N10-tri-p-coumaroylspermidine | ND | 0.11 ± 0.0 | 0.56 ± 0.0 | ND | ND | ND | ND | ND | ND |
| N1, N5, N10-tri-p-coumaroylspermidine (isomer) | 0.43 ± 0.0 | 0.22 ± 0.0 | 6.31 ± 0.1 | ND | ND | ND | ND | ND | ND |
| Tetracoumaroyl spermine | ND | ND | ND | ND | 0.18 ± 0.0 | ND | ND | ND | ND |
| Feruloyl dicoumaroyl spermidine | ND | ND | 0.75 ± 0.0 | ND | ND | ND | ND | ND | ND |
| Tetracoumaroyl spermine (isomer) | ND | 0.95 ± 0.0 | ND | ND | ND | ND | ND | ND | ND |
| Diferuloyl coumarouyl spermidine | 0.75 ± 0.0 | ND | ND | ND | ND | ND | ND | ND | ND |
| Polyamide derivative | ND | ND | 3.12 ± 0.2 | ND | ND | ND | ND | ND | ND |
| Tetracoumaroyl spermine (isomer) | ND | ND | ND | ND | 0.36 ± 0.0 | ND | ND | ND | ND |
| Tetracoumaroyl spermine (isomer) | ND | ND | ND | ND | 0.03 ± 0.0 | ND | ND | ND | ND |
| Tetracoumaroyl spermine (isomer) | ND | 0.99 ± 0.0 | ND | ND | 0.36 ± 0.0 | ND | ND | ND | ND |
| Tetracoumaroyl spermine (isomer) | ND | ND | ND | ND | 0.23 ± 0.0 | ND | ND | ND | ND |
| Total amount of phenolic compounds | 0.2 | 0.1 | 2.6 | 0.1 | 0.0 | 0.4 | 0.1 | 0.0 | 0.4 |
| Total amount of phenylamides | 3.7 | 2.9 | 14.7 | 0.0 | 1.2 | 0.0 | 0.0 | 0.0 | 0.0 |
| Compound | Mouth | Stomach | Intestine | ||||||
|---|---|---|---|---|---|---|---|---|---|
| BB-A1 | BB-A2 | BB-A3 | BB-A1 | BB-A2 | BB-A3 | BB-A1 | BB-A2 | BB-A3 | |
| Methyl herbacetin-O-dihexoside | 0.41 ± 0.0 | 0.33 ± 0.0 | 0.07 ± 0.0 | 0.22 ± 0.0 | 0.16 ± 0.0 | 0.06 ± 0.0 | 0.16 ± 0.0 | 0.12 ± 0.0 | 0.04 ± 0.0 |
| Methyl herbacetin-O-rutinoside | 0.08 ± 0.0 | 0.08 ± 0.0 | 0.01 ± 0.0 | 0.05 ± 0.0 | 0.05 ± 0.0 | ND | 0.04 ± 0.0 | 0.04 ± 0.0 | ND |
| Isorhamnetin-O-pentosyl hexoside | 0.05 ± 0.0 | 0.06 ± 0.0 | 0.04 ± 0.0 | 0.02 ± 0.0 | ND | 0.03 ± 0.0 | 0.02 ± 0.0 | ND | 0.04 ± 0.0 |
| Kaempferol-3-O-rutinoside | 0.01 ± 0.0 | 0.01 ± 0.0 | 0.01 ± 0.0 | 0.00 ± 0.0 | 0.00 ± 0.0 | 0.01 ± 0.0 | ND | 0.00 ± 0.0 | 0.00 ± 0.0 |
| Quercetin-3-O-glucoside | ND | ND | 0.01 ± 0.0 | ND | ND | 0.00 ± 0.0 | ND | ND | ND |
| Quercetin-O-malonyl hexoside | ND | 0.02 ± 0.0 | 0.02 ± 0.0 | ND | ND | ND | ND | ND | ND |
| Methyl herbacetin-3-O-hexoside | 0.02 ± 0.0 | ND | ND | 0.01 ± 0.0 | ND | ND | ND | ND | ND |
| Kaempferol-O-hexoside | ND | 0.00 ± 0.0 | 0.01 ± 0.0 | ND | ND | ND | ND | ND | ND |
| Isorhamnetin-O-hexoside | ND | 0.01 ± 0.0 | ND | ND | ND | ND | ND | ND | ND |
| Kaempferol-O-malonyl hexoside | ND | 0.03 ± 0.0 | 0.03 ± 0.0 | ND | 0.01 ± 0.0 | 0.03 ± 0.0 | ND | 0.01 ± 0.0 | 0.02 ± 0.0 |
| N1, N5, N10-tri-caffeoylspermidine | 1.84 ± 0.1 | 4.14 ± 0.2 | 0.40 ± 0.0 | 0.07 ± 0.0 | 0.09 ± 0.0 | ND | ND | ND | ND |
| N1-feruloyl-N5, N10-dicaffeoylspermidine | ND | 0.06 ± 0.0 | ND | ND | ND | ND | ND | ND | ND |
| N1-feruloyl-N5, N10-dicaffeoylspermidine (isomer) | ND | 0.17 ± 0.0 | ND | ND | ND | ND | ND | ND | ND |
| N1-p-coumaroyl-N5, N10-dicaffeoylspermidine | 0.21 ± 0.0 | 0.13 ± 0.0 | 0.06 ± 0.0 | ND | ND | ND | ND | ND | ND |
| N1-p-coumaroyl-N5, N10-dicaffeoylspermidine (isomer) | 0.50 ± 0.0 | 1.48 ± 0.0 | 0.69 ± 0.0 | ND | 0.03 ± 0.0 | ND | ND | ND | ND |
| N1, N5-di-p-coumaroyl-N10-caffeoylspermidine | 0.30 ± 0.0 | 0.07 ± 0.0 | 0.66 ± 0.0 | ND | ND | ND | ND | ND | ND |
| N1, N5-di-p-coumaroyl-N10-caffeoylspermidine (isomer) | 1.23 ± 0.0 | 1.79 ± 0.1 | ND | 0.07 ± 0.0 | ND | ND | ND | ND | ND |
| N1, N5, N10-tri-p-coumaroylspermidine | 12.25 ± 0.1 | 19.78 ± 0.2 | 7.17 ± 0.1 | 0.42 ± 0.0 | 0.18 ± 0.0 | ND | ND | ND | ND |
| Methyl herbacetin | ND | ND | 0.09 ± 0.0 | ND | ND | ND | ND | ND | ND |
| Feruloyl dicoumaroyl spermidine | 0.54 ± 0.1 | 0.50 ± 0.1 | ND | ND | ND | ND | ND | ND | ND |
| Tetracoumaroyl spermine | 0.59 ± 0.0 | 0.23 ± 0.0 | ND | ND | ND | ND | ND | ND | ND |
| Tetracoumaroyl spermine (isomer) | 0.69 ± 0.0 | ND | ND | ND | ND | ND | ND | ND | ND |
| Tetracoumaroyl spermine (isomer) | 0.68 ± 0.1 | ND | ND | ND | ND | ND | ND | ND | ND |
| Tetracoumaroyl spermine (isomer) | 1.14 ± 0.1 | ND | ND | ND | ND | ND | ND | ND | ND |
| Total amount of phenolic compounds | 0.6 | 0.6 | 0.3 | 0.3 | 0.2 | 0.1 | 0.2 | 0.2 | 0.1 |
| Total amount of phenylamides | 19.9 | 28.4 | 8.9 | 0.5 | 0.3 | 0.0 | 0.0 | 0.0 | 0.0 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Aylanc, V.; Tomás, A.; Russo-Almeida, P.; Falcão, S.I.; Vilas-Boas, M. Assessment of Bioactive Compounds under Simulated Gastrointestinal Digestion of Bee Pollen and Bee Bread: Bioaccessibility and Antioxidant Activity. Antioxidants 2021, 10, 651. https://doi.org/10.3390/antiox10050651
Aylanc V, Tomás A, Russo-Almeida P, Falcão SI, Vilas-Boas M. Assessment of Bioactive Compounds under Simulated Gastrointestinal Digestion of Bee Pollen and Bee Bread: Bioaccessibility and Antioxidant Activity. Antioxidants. 2021; 10(5):651. https://doi.org/10.3390/antiox10050651
Chicago/Turabian StyleAylanc, Volkan, Andreia Tomás, Paulo Russo-Almeida, Soraia I. Falcão, and Miguel Vilas-Boas. 2021. "Assessment of Bioactive Compounds under Simulated Gastrointestinal Digestion of Bee Pollen and Bee Bread: Bioaccessibility and Antioxidant Activity" Antioxidants 10, no. 5: 651. https://doi.org/10.3390/antiox10050651