
Insulin–Mimetic Dihydroxanthyletin-Type Coumarins from Angelica decursiva with Protein Tyrosine Phosphatase 1B and α-Glucosidase Inhibitory Activities and Docking Studies of Their Molecular Mechanisms


Abstract
:1. Introduction
2. Materials and Methods
2.1. General Experimental Procedures
2.2. Chemicals and Reagents
2.3. Plant Materials
2.4. Extraction, Fractionation, and Isolation
2.5. Assay for PTP1B Inhibitory Assay
2.6. α-Glucosidase Inhibitory Assay
2.7. Inhibition of ONOO−-Mediated Protein Tyrosine Nitration
2.8. Assay for ONOO− Scavenging Activity
2.9. Total ROS Generation Inhibitory Activity Assay
2.10. Measurement of Intracellular Reactive Oxygen Species Level
2.11. Kinetic Parameters of Coumarins in Lineweaver-Burk and Dixon Plots for Both PTP1B and α-Glucosidase Inhibition
2.12. Molecular Docking Simulation of PTP1B and α-Glucosidase Inhibition
2.13. Assay for Cell Viability
2.14. Induction of Insulin Resistant HepG2 Cells
2.15. 2-NBDG Glucose Uptake Assay
2.16. Cell Lysates Preparation and Analysis of Western Blot
2.17. Statistical Analysis
3. Results
3.1. Coumarins’ Activity in Inhibiting PTP1B and α-glucosidase
3.2. PTP1B and α-Glucosidase Inhibitory Enzyme Kinetics
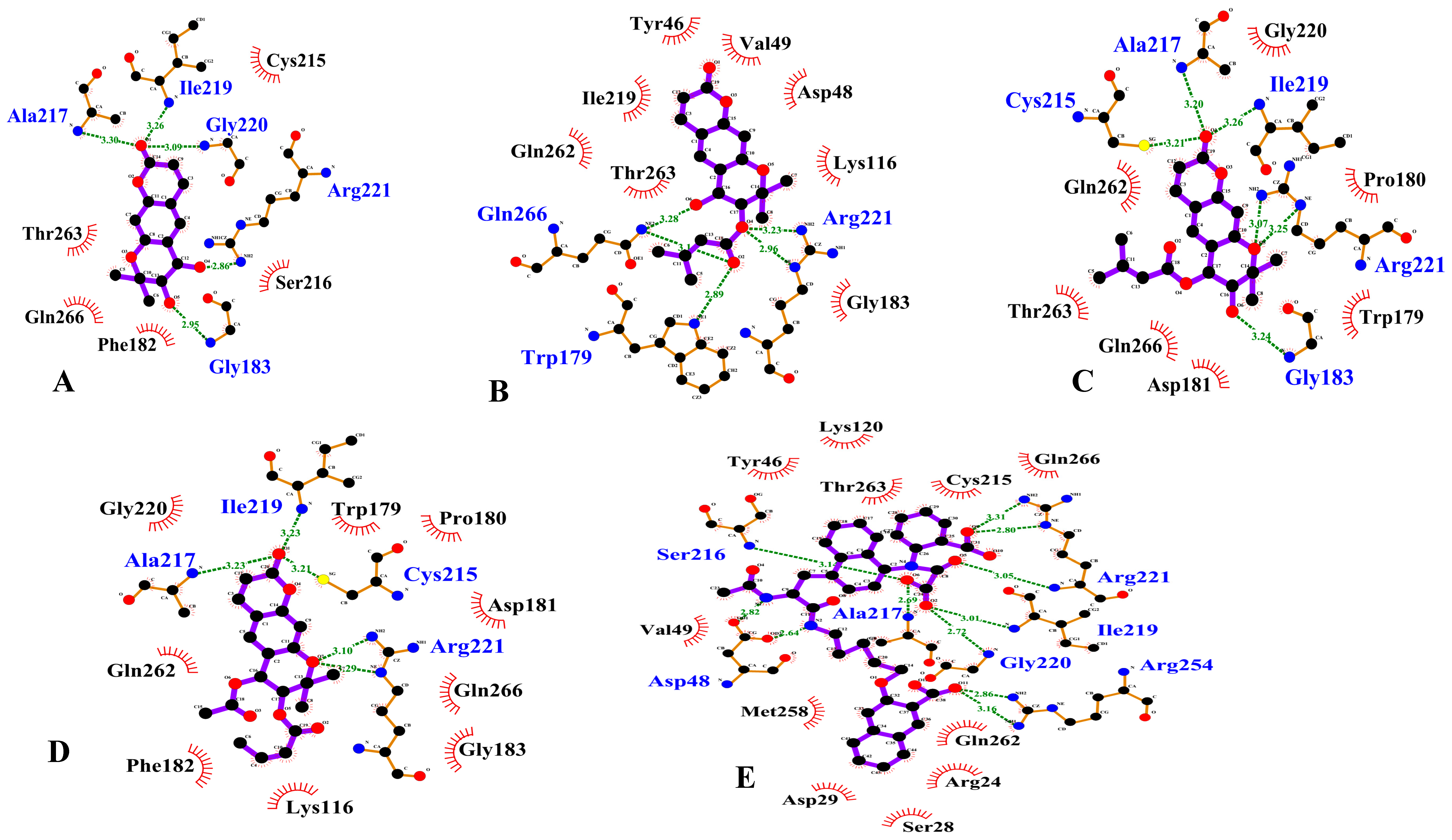
3.3. PTP1B Inhibition Molecular Docking Simulation
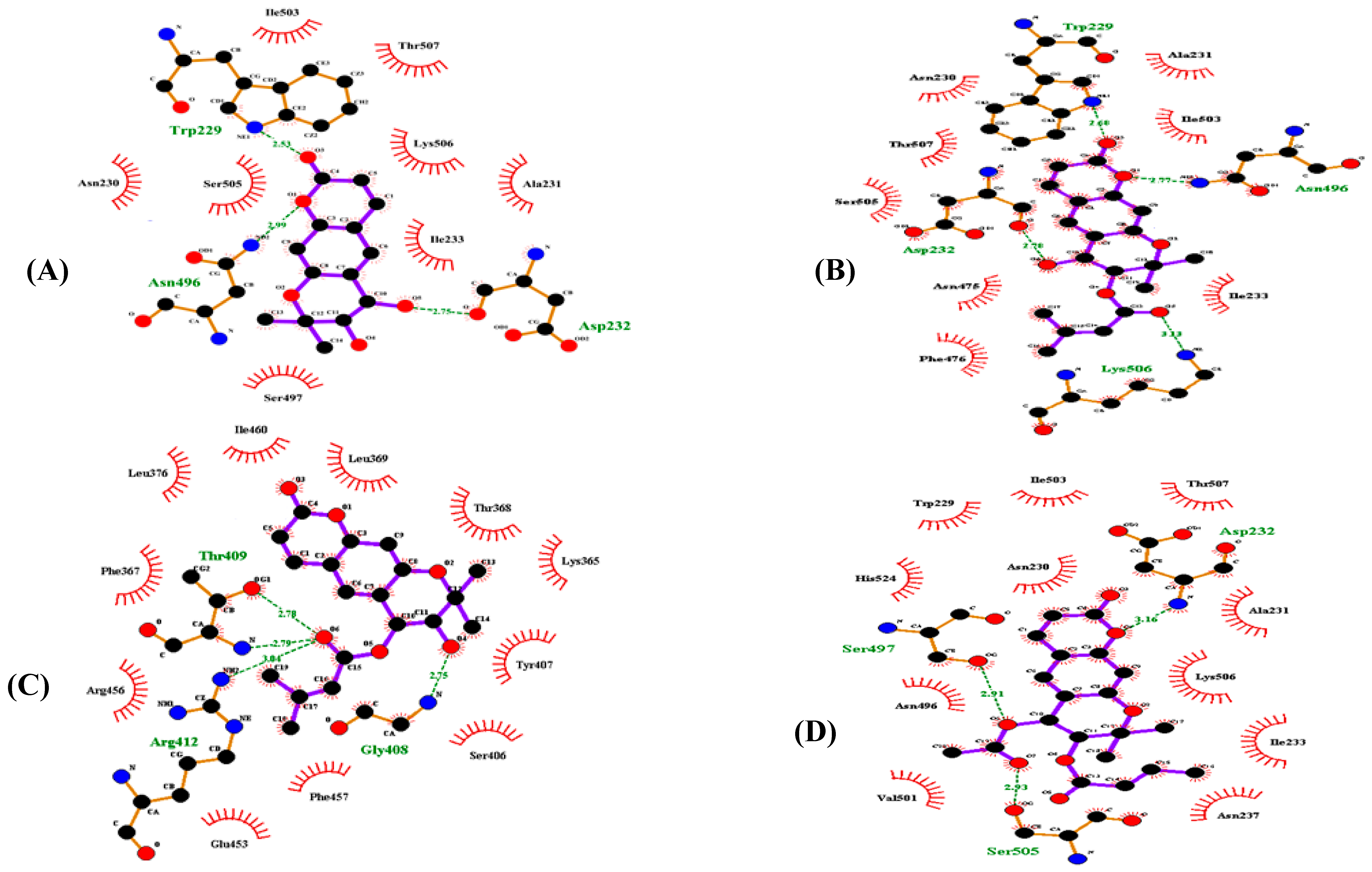
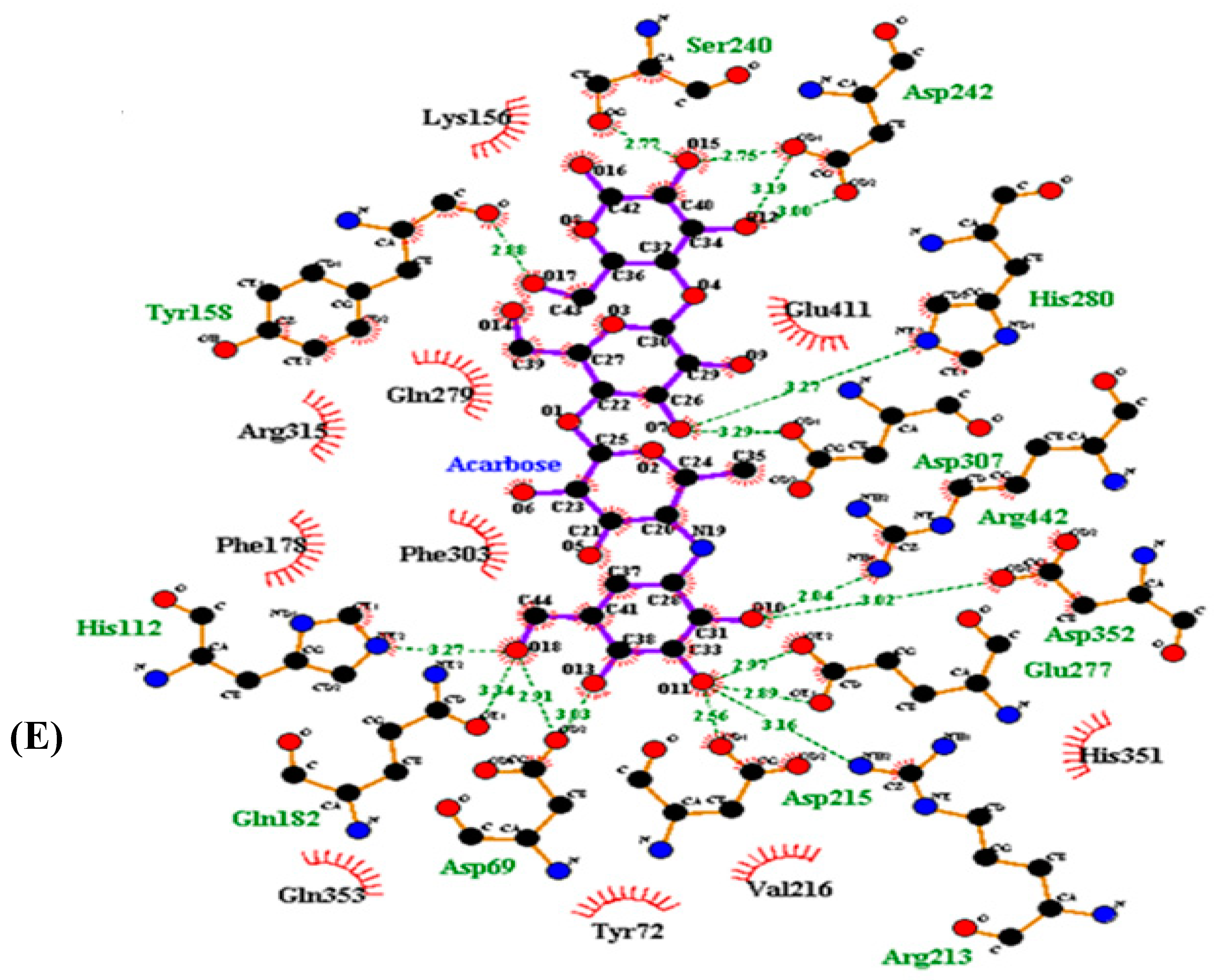
3.4. Molecular Docking Simulation of α-Glucosidase Inhibition
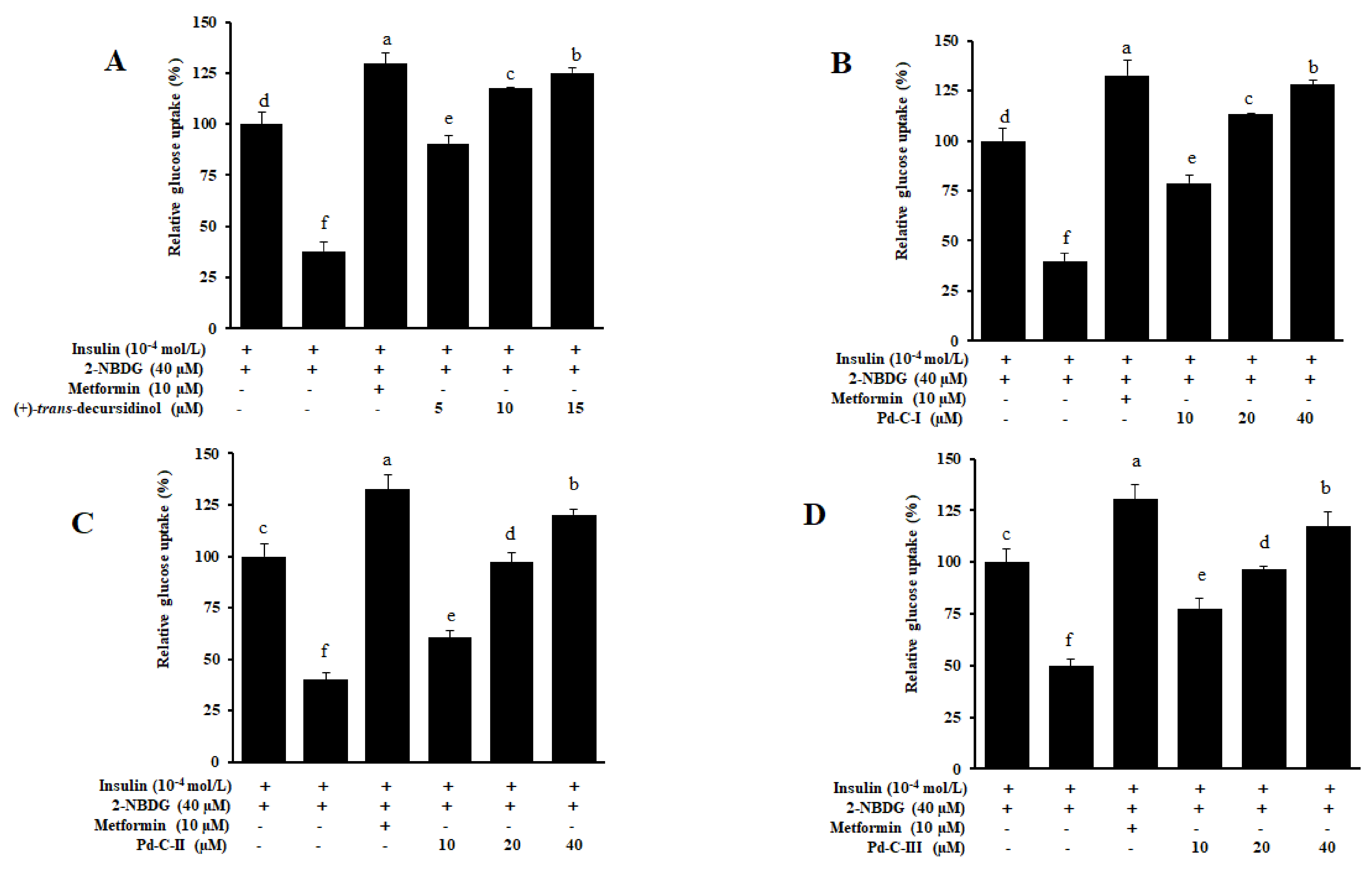
3.5. Coumarins’ Effect on Glucose Uptake in HepG2 Insulin-Resistant Cells
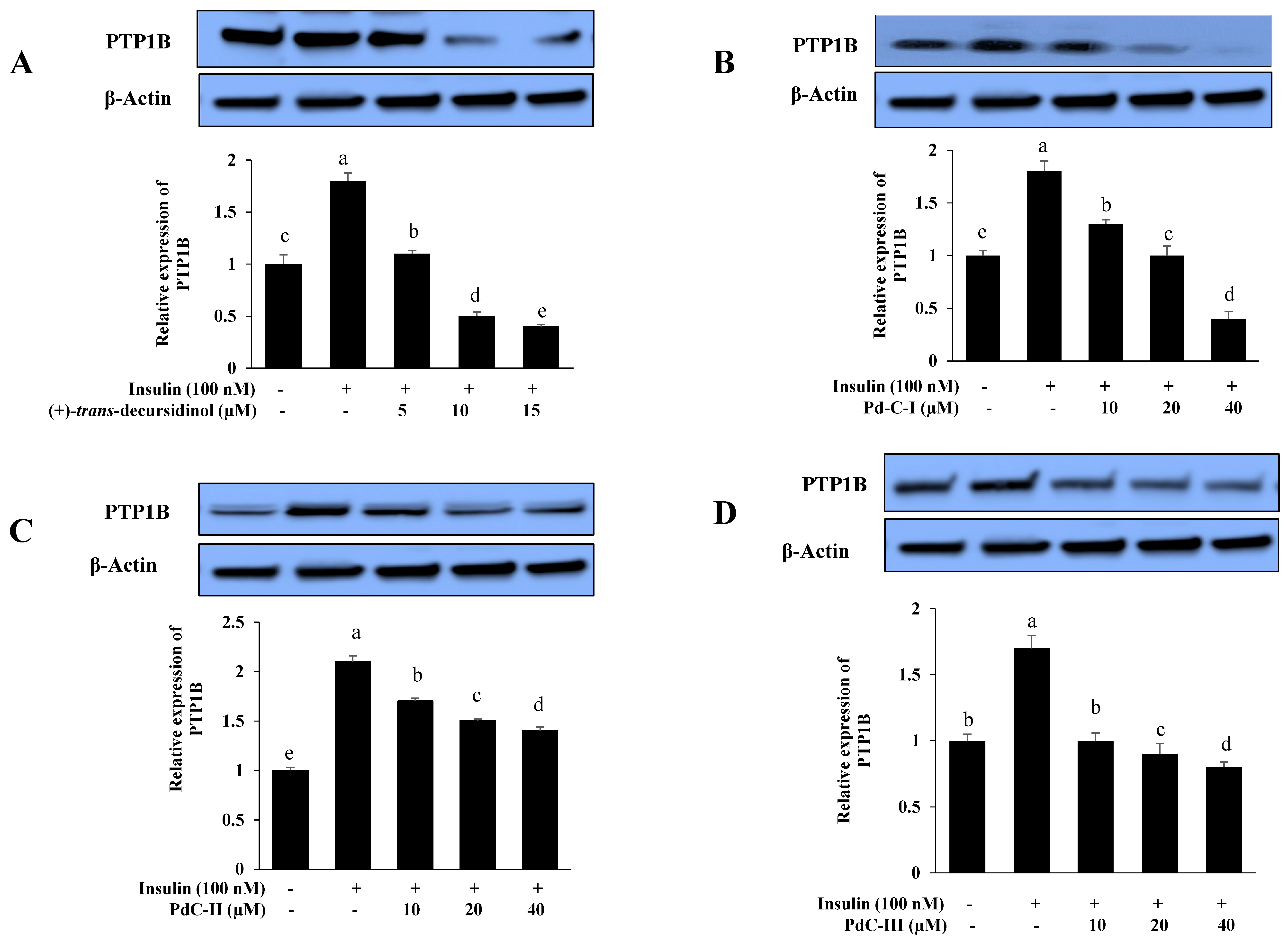
3.6. Effects on PTP1B Expression Level in Insulin-Resistant HepG2 Cells
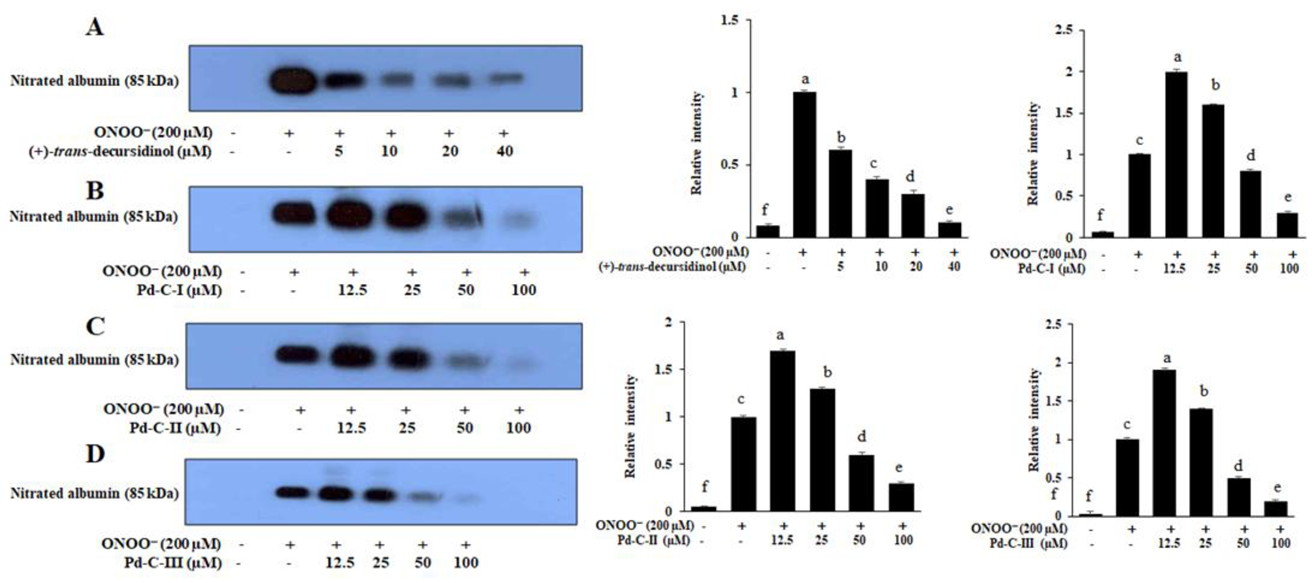
3.7. The Coumarins’ Inhibitory Effect on ONOO−-Mediated Albumin Nitration
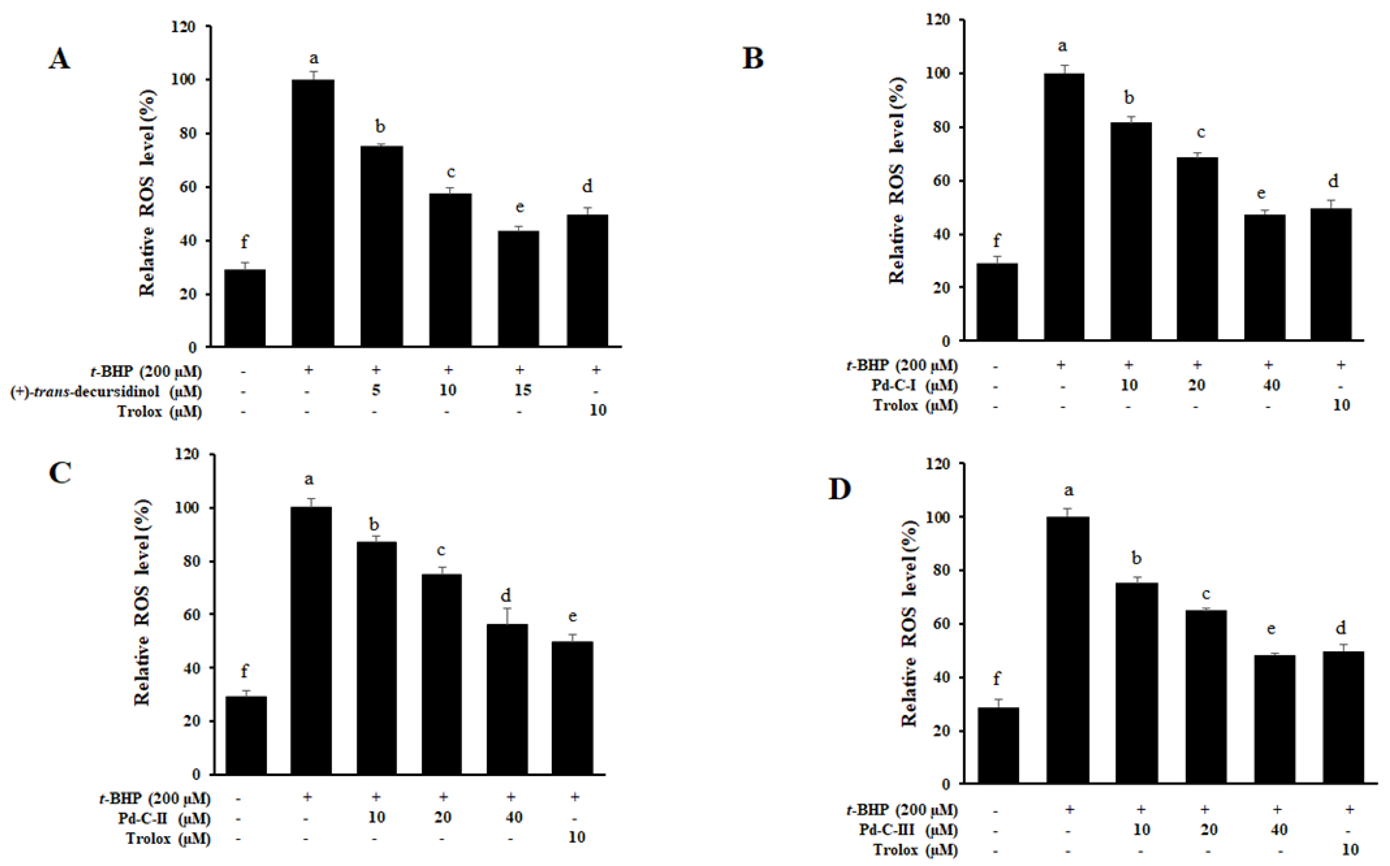
3.8. The Effect of Coumarins’ on the Levels of Intracellular ROS in t-BHP-Treated HepG2 Cells
3.9. The Coumarins’ Inhibitory Effect on ROS and ONOO−
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Kong, W.; Wu, D.; Hu, N.; Li, N.; Dai, C.; Chen, X.; Suo, Y.; Li, G.; Wu, Y. Robust hybrid enzyme nanoreactor mediated plasmonic sensing strategy for ultrasensitive screening of anti-diabetic drug. Biosens. Bioelectron. 2018, 99, 653–659. [Google Scholar] [CrossRef]
- Fox, D.A.; Islam, N.; Sutherland, J.; Reimer, K.; Amed, S. Type 1 diabetes incidence and prevalence trends in a cohort of Canadian children and youth. Pediatr. Diabetes 2018, 19, 501–505. [Google Scholar] [CrossRef] [PubMed]
- Jung, H.A.; Ali, M.Y.; Bhakta, H.K.; Min, B.S.; Choi, J.S. Prunin is a highly potent flavonoid from Prunus davidiana stems that inhibits protein tyrosine phosphatase 1B and stimulates glucose uptake in insulin-resistant HepG2 cells. Arch. Pharm. Res. 2017, 40, 37–48. [Google Scholar] [CrossRef] [PubMed]
- Lebovitz, H.E. Type 2 diabetes mellitus—Current therapies and the emergence of surgical options. Nat. Rev. Endocrinol. 2011, 7, 408. [Google Scholar] [CrossRef] [PubMed]
- Dang, F.; Jiang, Y.; Pan, R.; Zhou, Y.; Wu, S.; Wang, R.; Zhuang, K.; Zhang, W.; Li, T.; Man, C. Administration of Lactobacillus paracasei ameliorates type 2 diabetes in mice. Food Funct. 2018, 9, 3630–3639. [Google Scholar] [CrossRef]
- Paudel, P.; Yu, T.; Seong, S.H.; Kuk, E.B.; Jung, H.A.; Choi, J.S. Protein tyrosine phosphatase 1B inhibition and glucose uptake potentials of mulberrofuran G, albanol B, and kuwanon G from root bark of Morus alba L. in insulin resistant HepG2 cells: An in vitro and in silico study. Int. J. Mol. Sci. 2018, 19, 1542. [Google Scholar] [CrossRef] [Green Version]
- Johnson, T.O.; Ermolieff, J.; Jirousek, M.R. Protein tyrosine phosphatase 1B inhibitors for diabetes. Nat. Rev. Drug Discov. 2002, 1, 696–709. [Google Scholar] [CrossRef]
- Haque, A.; Andersen, J.N.; Salmeen, A.; Barford, D.; Tonks, N.K. Conformation-sensing antibodies stabilize the oxidized form of PTP1B and inhibit its phosphatase activity. Cell 2011, 147, 185–198. [Google Scholar] [CrossRef] [Green Version]
- Goldstein, B.J.; Bitter-Kowalczyk, A.; White, M.F.; Harbeck, M.J. Tyrosine dephosphorylation and deactivation of insulin receptor substrate-1 by protein tyrosine phosphatase 1B. Possible facilitation by the formation of a ternary complex with the GRB2 adaptor protein. Biol. Chem. 2000, 275, 4283–4289. [Google Scholar] [CrossRef] [Green Version]
- Elchebly, M.; Payette, P.; Michaliszyn, E.; Cromlish, W.; Collins, S.; Loy, A.L.; Normandin, D.; Cheng, A.; Himms-Hagen, J.; Chan, C.C.; et al. Increased insulin sensitivity and obesity resistance in mice lacking the protein tyrosine phosphatase-1B gene. Science 1999, 283, 1544–1548. [Google Scholar] [CrossRef]
- Klaman, L.D.; Boss, O.; Peroni, O.D.; Kim, J.K.; Martino, J.L.; Zabolotny, J.M.; Moghal, N.; Lubkin, M.; Kim, Y.B.; Sharpe, A.H.; et al. Increased energy expenditure, decreased adiposity, and tissue- specific insulin sensitivity in protein-tyrosine phosphatase1B-deficient mice. Mol. Cell. Biol. 2000, 20, 5479–5489. [Google Scholar] [CrossRef] [Green Version]
- Rasouli, H.; Hosseini-Ghazvini, S.M.B.; Adibi, H.; Khodarahmi, R. Differential α-amylase/α-glucosidase inhibitory activities of plant-derived phenolic compounds: A virtual screening perspective for the treatment of obesity and diabetes. Food Funct. 2017, 8, 1942–1954. [Google Scholar] [CrossRef]
- Han, L.; Fang, C.; Zhu, R.; Peng, Q.; Li, D.; Wang, M. Inhibitory effect of phloretin on α-glucosidase: Kinetics, interaction mechanism and molecular docking. Int. J. Biol. Macromol. 2017, 95, 520–527. [Google Scholar] [CrossRef]
- Lee, B.H.; Rose, D.R.; Lin, A.H.-M.; Quezada-Calvillo, R.; Nichols, B.L.; Hamaker, B.R. Contribution of the individual small intestinal α-glucosidases to digestion of unusual α-linked glycemic disaccharides. J. Agric. Food Chem. 2016, 64, 6487–6494. [Google Scholar] [CrossRef] [PubMed]
- Wen, H.; Tang, B.; Stewart, A.J.; Tao, Y.; Shao, Y.; Cui, Y.; Yue, H.; Pei, J.; Liu, Z.; Mei, L.; et al. Erythritol attenuates postprandial blood glucose by inhibiting α-glucosidase. J. Agric. Food Chem. 2018, 66, 1401–1407. [Google Scholar] [CrossRef] [PubMed]
- Wilson, R.L. Free radicals and tissue damage, mechanistic evidence from radiation studies. In Biochemical Mechanisms of Liver Injury; Academic Press: New York, NY, USA, 1998; pp. 123–125. [Google Scholar]
- Evans, J.L.; Goldfine, I.D.; Maddux, B.A.; Grodsky, G.M. Are oxidative stress activated signaling pathways mediators of insulin resistance and beta-cell dysfunction? Diabetes 2003, 52, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Drews, G.; Krippeit-Drews, P.; Dufer, M. Oxidative stress and beta-cell dysfunction. Pflugers Arch. 2010, 460, 703–718. [Google Scholar] [CrossRef]
- Bonnefont-Rousselot, D.; Bastard, J.P.; Jaudon, M.C.; Delattre, J. Consequences of the diabetic status on the oxidant/antioxidant balance. Diabetes Metab. 2000, 26, 163–176. [Google Scholar]
- Ceriello, A. Oxidative stress and diabetes-associated complications. Endocr. Pract. 2006, 12, 60–62. [Google Scholar] [CrossRef] [PubMed]
- Rains, J.L.; Jain, S.K. Oxidative stress, insulin signaling and diabetes. Free Radic. Biol. Med. 2011, 50, 567–575. [Google Scholar] [CrossRef] [Green Version]
- Ischiropoulos, H. Biological tyrosine nitration: A pathophysiological function of nitric oxide and reactive oxygen species. Arch. Biochem. Biophys. 1998, 356, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Ceriello, A.; Mercuri, F.; Quagliaro, L.; Assaloni, R.; Motz, E.; Tonutti, L.; Tabaga, C. Detection of nitrotyrosine in the diabetic plasma: Evidence of oxidative stress. Diabetologia 2001, 44, 834–838. [Google Scholar]
- Chi, Y.S.; Cheon, B.S.; Kim, H.P. Effect of wogonin, a plant flavone from Scutellaria radix, on the suppression of cyclooxygenase and the induction of inducible nitric oxide synthase in lipolpolysaccharide-treated RAW 264.7 cells. Biochem. Pharmacol. 2001, 61, 1195–1203. [Google Scholar] [PubMed]
- Kong, L.Y.; Li, Y.; Min, Z.D.; Li, X.; Zhu, T.R. Coumarins from Peucedanum praeruptorum. Phytochemistry 1996, 41, 1423–1426. [Google Scholar] [CrossRef]
- Yi, J.H.; Park, I.K.; Choi, K.S.; Shin, S.C.; Ahn, Y.J. Toxicity of medicinal plant extracts to Lycoriella ingenua (Diptera: Sciaridae) and Coboldia fuscipes (Diptera: Scatopsidae). J. Asia Pac. Entomol. 2008, 11, 221–223. [Google Scholar] [CrossRef]
- Zhao, D.; Islam, M.N.; Ahn, B.R.; Jung, H.A.; Kim, B.W.; Choi, J.S. In vitro antioxidant and anti-inflammatory activities of Angelica decursiva. Arch. Pharm. Res. 2012, 35, 179–192. [Google Scholar] [CrossRef]
- Ishita, I.J.; Islam, M.N.; Kim, Y.S.; Choi, R.J.; Sohn, H.S.; Jung, H.A.; Choi, J.S. Coumarins from Angelica decursiva inhibit lipopolysacharide-induced nitric oxide production in RAW 264.7 cells. Arch. Pharm. Res. 2016, 39, 115–126. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.Y.; Jung, H.A.; Choi, J.S. Anti-diabetic and anti-Alzheimer’s disease activities of Angelica decursiva. Arch. Pharm. Res. 2015, 38, 2216–2227. [Google Scholar]
- Ali, M.Y.; Jannat, S.; Jung, H.A.; Choi, R.J.; Roy, A.; Choi, J.S. Anti-Alzheimer potential of coumarins from Angelica decursiva and Artemisia capillaris and structure-activity analysis. Asian Pac. J. Trop. Med. 2016, 9, 103–111. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.Y.; Jannat, S.; Jung, H.A.; Jeong, H.O.; Chung, H.Y.; Choi, J.S. Coumarins from Angelica decursiva inhibit α-glucosidase activity and protein tyrosine phosphatase 1B. Chem. Biol. Interact. 2016, 252, 93–101. [Google Scholar] [CrossRef]
- Ali, M.Y.; Seong, S.H.; Reddy, M.R.; Seo, S.Y.; Choi, J.S.; Jung, H.A. Kinetics and molecular docking studies of 6-formyl umbelliferone isolated from Angelica decursiva as an inhibitor of cholinesterase and BACE1. Molecules 2017, 22, 1604. [Google Scholar] [CrossRef] [Green Version]
- Ali, M.Y.; Seong, S.H.; Jung, H.A.; Jannat, S.; Choi, J.S. Kinetics and molecular docking of dihydroxanthyletin-type coumarins from Angelica decursiva that inhibit cholinesterase and BACE1. Arch. Pharm. Res. 2018, 41, 753–764. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.Y.; Jung, H.A.; Jannat, S.; Choi, J.S. Dihydroxanthyletin-type coumarins from Angelica decursiva that inhibits the formation of advanced glycation end products and human recombinant aldose reductase. Arch. Pharm. Res. 2018, 41, 196–207. [Google Scholar] [CrossRef] [PubMed]
- Ali, M.Y.; Seong, S.H.; Jung, H.A.; Choi, J.S. Ethanobotany, phytochemistry, and pharmacology of Angelica decursiva Fr. et Sav. Nat. Prod. Sci. 2019, 25, 181–199. [Google Scholar] [CrossRef]
- Ali, M.Y.; Seong, S.H.; Jung, H.A.; Choi, J.S. Angiotensin-I-converting enzyme inhibitory activity of coumarins from Angelica decursiva. Molecules 2019, 24, 3937. [Google Scholar] [CrossRef] [Green Version]
- Chen, Y.C.; Chen, P.Y.; Wu, C.C.; Tsai, I.L.; Chen, I.S. Chemical constituents and anti-platelet aggregation activity from the root of Peucedanum formosanum. J. Food Drug Anal. 2008, 16, 15–25. [Google Scholar]
- Sarkhail, P. Traditional uses phytochemistry and pharmacological properties of the genus Peucedanum: A review. J. Ethnopharmacol. 2014, 156, 235–270. [Google Scholar] [CrossRef]
- Ahn, K.S.; Sim, W.S.; Lee, I.K.; Seu, Y.B.; Kim, I.H. Decursinol angelate: A cytotoxic and protein kinase C activating agent from the root of Angelica gigas. Planta Med. 1997, 63, 360–361. [Google Scholar] [CrossRef]
- Lee, Y.Y.; Lee, S.; Jin, J.L.; Yun-Choi, H.S. Platelet antiaggregatory effects of coumarins from the roots of Angelica genuflexa and Angelica gigas. Arch. Pharm. Res. 2003, 26, 723–726. [Google Scholar] [CrossRef] [PubMed]
- Gao, Q.; Jeon, S.J.; Jung, H.A.; Lee, H.E.; Park, S.J.; Lee, Y.; Lee, Y.; Ko, S.Y.; Kim, B.; Choi, J.S.; et al. Nodakenin enhances cognitive function and adult hippocampal neurogenesis in mice. Neurochem. Res. 2015, 40, 1438–1447. [Google Scholar] [CrossRef]
- Kang, S.Y.; Lee, K.Y.; Sung, S.H.; Kim, Y.C. Four new neuroprotective dihydropyranocoumarins from Angelica gigas. J. Nat. Prod. 2005, 68, 56–59. [Google Scholar] [CrossRef]
- Kim, D.H.; Kim, D.Y.; Kim, Y.C.; Jung, J.W.; Lee, S.J.; Yoon, B.H.; Cheong, J.H.; Kim, Y.S.; Kang, S.S.; Ko, K.H.; et al. Nodakenin, a coumarin compound, ameliorates scopolamine-induced memory disruption in mice. Life Sci. 2007, 80, 1944–1950. [Google Scholar] [CrossRef]
- Islam, M.N.; Choi, R.J.; Jin, S.E.; Kim, Y.S.; Ahn, B.R.; Zhao, D.; Jung, H.A.; Choi, J.S. Mechanism of anti-inflammatory activity of umbelliferone 6-carboxylic acid isolated from Angelica decursiva. J. Ethnopharmacol. 2012, 144, 175–181. [Google Scholar] [CrossRef]
- Sakakibara, I.; Okuyama, T.; Shibata, S. Studies on coumarins of a Chinese drug “Qian-Hu”. Planta Med. 1982, 44, 199–203. [Google Scholar] [CrossRef]
- Kooy, N.W.; Royall, J.A.; Ischiropoulos, H.; Beckman, J.S. Peroxynitrite-mediated oxidation of dihydrorhodamine 123. Free Radic. Biol. Med. 1994, 2, 149–156. [Google Scholar] [CrossRef]
- Lebel, C.P.; Bondy, S.C. Sensitive and rapid quantitation of oxygen reactive species formation in rat synaptosome. Neurochem. Int. 1990, 17, 435–440. [Google Scholar] [CrossRef] [Green Version]
- Lineweaver, H.; Burk, D. The determination of enzyme dissociation constants. J. Am. Chem. Soc. 1934, 56, 658–666. [Google Scholar] [CrossRef]
- Dixon, M. The determination of enzyme inhibitor constant. Biochem. J. 1953, 55, 170–171. [Google Scholar] [CrossRef]
- Cornish-Bowden, A. A simple graphical method for determining the inhibition constants of mixed, uncompetitive and non- competitive inhibitors. Biochem. J. 1974, 137, 143–144. [Google Scholar] [CrossRef] [PubMed]
- Wiesmann, C.; Barr, K.J.; Kung, J.; Zhu, J.; Erlanson, D.A.; Shen, W.; Fahr, B.J.; Zhong, M.; Taylor, L.; Randal, M.; et al. Allosteric inhibition of protein tyrosine phosphatase 1B. Nat. Struct. Mol. Biol. 2004, 11, 730–737. [Google Scholar] [CrossRef]
- Ali, M.Y.; Kim, D.H.; Seong, S.H.; Kim, H.R.; Jung, H.A.; Choi, J.S. α-Glucosidase and protein tyrosine phosphatase 1B inhibitory activity of plastoquinones from marine brown alga Sargassum serratifolium. Mar. Drugs. 2017, 15, 368. [Google Scholar] [CrossRef] [Green Version]
- Verma, M.; Gupta, S.J.; Chaudhary, A.; Garg, V.K. Protein tyrosine phosphatase 1B inhibitors as antidiabetic agents –A brief review. Bioorganic Chem. 2017, 70, 267–283. [Google Scholar] [CrossRef]
- Wang, L.J.; Jiang, B.; Wu, N.; Wang, S.Y.; Shi, D.Y. Natural and semisynthetic protein tyrosine phosphatase 1B (PTP1B) inhibitors as anti-diabetic agents. RSC. Adv. 2015, 5, 48822–48834. [Google Scholar] [CrossRef]
- Israili, Z.H. Advances in the treatment of type 2 diabetes mellitus. Am. J. Ther. 2011, 18, 117–152. [Google Scholar] [CrossRef]
- Wu, X.; Hardy, V.E.; Joseph, J.I. Protein-tyrosine phosphatase activity in human adipocytes is strongly correlated with insulin- stimulated glucose uptake and is a target of insulin induced oxidative inhibition. Metabolism 2003, 52, 705–712. [Google Scholar] [CrossRef]
- Panzhinskiy, E.; Ren, J.; Nair, S. Pharmacological inhibition of protein tyrosine phosphatase 1B: A promising strategy for the treatment of obesity and type 2 diabetes mellitus. Curr. Med. Chem. 2013, 20, 2609–2625. [Google Scholar] [CrossRef] [PubMed]
- Derosa, G.; Maffioli, P. α-Glucosidase inhibitors and their use in clinical practice. Arch. Med. Sci. 2012, 8, 899–906. [Google Scholar] [CrossRef] [PubMed]
- Barford, D.; Flint, A.J.; Tonks, N.K. Crystal structure of human protein tyrosine phosphatase 1B. Science 1994, 263, 1397–1404. [Google Scholar] [CrossRef] [PubMed]
- Shen, K.; Keng, Y.F.; Wu, L.; Guo, X.L.; Lawrence, D.S.; Zhang, Z.Y. Acquisition of a specific and potent PTP1B inhibitor from a novel combinatorial library and screening procedure. J. Biol. Chem. 2001, 276, 47311–47319. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Maeda, A.; Kai, K.; Ishii, M.; Ishii, T.; Akagawa, M. Safranal, a novel protein tyrosine phosphatase 1B inhibitor, activates insulin signaling in C2C12 myotubes and improves glucose tolerance in diabetic KK-Ay mice. Mol. Nutr. Food Res. 2014, 58, 1177–1189. [Google Scholar] [CrossRef]
- Takada, M.; Sumi, M.; Maeda, A.; Watanabe, F.; Kamiya, T.; Ishii, T.; Nakano, M.; Akagawa, M. Pyrroloquinoline quinone, a novel protein tyrosine phosphatase 1B inhibitor, activates insulin signaling in C2C12myotubes and improves impaired glucose tolerance in diabetic KK-Ay mice. Biochem. Biophys. Res. Commun. 2012, 428, 315–320. [Google Scholar] [CrossRef] [PubMed]
- Liu, Z.Q.; Liu, T.; Chen, C.; Li, M.Y.; Wang, Z.Y.; Chen, R.S.; Wei, G.X.; Wang, X.Y.; Luo, D.Q. Fumosorinone, a novel PTP1B inhibitor, activates insulin signaling in insulin-resistance HepG2 cells and shows anti-diabetic effect in diabetic KKAy mice. Toxicol. Appl. Pharmacol. 2015, 285, 61–70. [Google Scholar] [CrossRef] [PubMed]
- Ross, J.A.; Kasum, C.M. Dietary flavonoids: Bioavailability, metabolic effects, and safety. Annu. Rev. Nutr. 2002, 22, 19–34. [Google Scholar] [CrossRef]
- Vitaglione, P.; Morisco, F.; Caporaso, N.; Fogliano, V. Dietary antioxidant compounds and liver health. Crit. Rev. Food Sci. Nutr. 2004, 44, 575–586. [Google Scholar] [CrossRef] [Green Version]
- Johansen, J.S.; Harris, A.K.; Rychly, D.J.; Ergu, A. Oxidative stress and the use of antioxidants in diabetes: Linking basic science to clinical practice. Cardiovasc. Diabetol. 2005, 4, 5. [Google Scholar] [CrossRef] [Green Version]









{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Compounds | Protein Tyrosine Phosphatase 1B | α-Glucosidase | ||||
|---|---|---|---|---|---|---|
| IC50 (µM) a | Ki Value b | Inhibition Type c | IC50 (µM) a | Ki Value b | Inhibition Type c | |
| (+)-trans-decursidinol | 2.33 ± 0.07 | 2.84 | Competitive | 11.32 ± 0.56 | 16.02 | Competitive |
| Pd-C-I | 4.32 ± 0.12 | 6.21 | Competitive | 17.40 ± 0.33 | 22.98 | Mixed |
| Pd-C-II | 6.17 ± 0.31 | 9.72 | Competitive | 24.74 ± 0.89 | 30.83 | Mixed |
| Pd-C-III | 11.98 ± 0.43 | 16.04 | Mixed | 36.77 ± 1.04 | 55.59 | Noncompetitive |
| Ursolic acid d | 6.87 ± 0.19 | - | - | |||
| Acarbose e | 169.49 ± 3.25 | - | - | |||
| Compounds | Binding Energies (Kcal/mol) a | No. of H Bond b | H-bonding Interacting Residues c | Van der Waals Interacting Residues d |
|---|---|---|---|---|
| (+)-trans-decursidinol | −7.9 | 5 | Ala27, Ile219, Gly220, Arg221, Gly183 | Cys215, Gln266, Phe182, Thr263, Ser216 |
| Pd-C-I | −7.8 | 6 | Gln266, Arg221, Trp179 | Asp48, Gln262, Gly163, Ile219, Lys116, Thr263, Tyr46, Val49 |
| Pd-C-II | −7.5 | 6 | Cys215, Ala217, Ile219, Gly183, Arg221 | Asp181, Gln266, Gly220, Gln262, Pro180, Trp179, Thr263 |
| Pd-C-III | −7.7 | 5 | Ile219, Ala217, Cys215, Arg221 | Asp181, Gly183, Gln266, Gln262, Gly220, Lys116, Phe182, Pro180, Trp179 |
| Compound 23 | −11.23 | 11 | Asp48, Ala217, Arg221, Arg254, Gly220, Ile219, Ser216 | Arg24, Asp29, Cys215, Gln266, Gln262, Lys120, Met258, Ser28, Thr263, Tyr46, Val49 |
| Compounds. | Binding Energies (Kcal/mol) a | No. of H Bond b | H-bonding Interacting Residues c | Van der Waals Interacting Residues d |
|---|---|---|---|---|
| (+)-trans-decursidinol | −7.09 | 3 | Trp229, Ans496, Asp232 | Ile503, Thr507, Lys506, Ala231, Ile233, Ser497, Asn230, Ser505 |
| Pd-C-I | −8.09 | 4 | Trp229, Asp232, Ans496, Lys506 | Asn230, Ala231, Thr507, Ile503, Ser505, Asn475, Ile233, Phe476 |
| Pd-C-II | −7.6 | 4 | Thr409, Arg412, Gly408 | Ile460, Leu369, Thr368, Lys365, Tyr407, Ser406, Phe457, Glu453, Arg456, Phe367, Leu376 |
| Pd-C-III | −7.24 | 3 | Asp232, Ser497, Ser505 | Ile503, Thr507, Ala231, Lys506, Ile233, Asn237, Val501, Asn496, His524, Asn230, Trp229 |
| Acarbose | −10.20 | 17 | Arg442, Asp307, Asp215, Arg213, Asp352, Asp69, Asp242, His280, Tyr158, His112, Gln182, Glu277, Ser240, | Arg315, Gln353, His351, Glu411, Tyr72, Lys156, Gln279, Phe178, Phe303, Val216 |
| Test Sample | IC50 (µM) a | |
|---|---|---|
| ONOO− | ROS | |
| (+)-trans-decursidinol | 4.57 ± 0.21 | 9.77 ± 0.32 |
| Pd-C-I | 1.82 ± 0.44 | 6.32 ± 0.41 |
| Pd-C-II | 3.77 ± 0.15 | 10.76 ± 0.73 |
| Pd-C-III | 1.44 ± 0.22 | 9.69 ± 0.21 |
| l-Penicillamine b | 1.86 ± 0.11 | - |
| Trolox c | - | 3.29 ± 0.26 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Ali, M.Y.; Jannat, S.; Jung, H.A.; Choi, J.S. Insulin–Mimetic Dihydroxanthyletin-Type Coumarins from Angelica decursiva with Protein Tyrosine Phosphatase 1B and α-Glucosidase Inhibitory Activities and Docking Studies of Their Molecular Mechanisms. Antioxidants 2021, 10, 292. https://doi.org/10.3390/antiox10020292
Ali MY, Jannat S, Jung HA, Choi JS. Insulin–Mimetic Dihydroxanthyletin-Type Coumarins from Angelica decursiva with Protein Tyrosine Phosphatase 1B and α-Glucosidase Inhibitory Activities and Docking Studies of Their Molecular Mechanisms. Antioxidants. 2021; 10(2):292. https://doi.org/10.3390/antiox10020292
Chicago/Turabian StyleAli, Md Yousof, Susoma Jannat, Hyun Ah Jung, and Jae Sue Choi. 2021. "Insulin–Mimetic Dihydroxanthyletin-Type Coumarins from Angelica decursiva with Protein Tyrosine Phosphatase 1B and α-Glucosidase Inhibitory Activities and Docking Studies of Their Molecular Mechanisms" Antioxidants 10, no. 2: 292. https://doi.org/10.3390/antiox10020292