Prevalence of Antibiotic Resistance Genes in Multidrug-Resistant Enterobacteriaceae on Portuguese Livestock Manure



1
Environment Department, Research Centre for Natural Resources, Environment and Society (CERNAS), College of Agriculture, Polytechnic of Coimbra, 3045-601 Coimbra, Portugal
2
Department Chemical Sciences and Biomolecules, School Allied Health Sciences, Polytechnic of Porto, 4200-072 Porto, Portugal
*
Author to whom correspondence should be addressed.
Antibiotics 2019, 8(1), 23; https://doi.org/10.3390/antibiotics8010023
Submission received: 8 January 2019
/
Revised: 22 February 2019
/
Accepted: 23 February 2019
/
Published: 13 March 2019
(This article belongs to the Special Issue Antimicrobial Resistance in Gram-negative Bacteria)
Abstract
:The exposure of both crop fields and humans to antibiotic-resistant bacteria in animal excreta is an emergent concern of the One Health initiative. This study assessed the contamination of livestock manure from poultry, pig, dairy farms and slaughterhouses in Portugal with resistance determinants. The resistance profiles of 331 Enterobacteriaceae isolates to eight β-lactam (amoxicillin, cefoxitin, cefotaxime, cefpirome, aztreonam, ceftazidime, imipenem and meropenem) and to five non-β-lactam antibiotics (tetracycline (TET), trimethoprim/sulfamethoxazole (SXT), ciprofloxacin (CIP), chloramphenicol (CHL) and gentamicin) was investigated. Forty-nine integron and non-β-lactam resistance genes were also screened for. Rates of resistance to the 13 antibiotics ranged from 80.8% to 0.6%. Multidrug resistance (MDR) rates were highest in pig farm samples (79%). Thirty different integron and resistance genes were identified. These were mainly associated with resistance to CHL (catI and catII), CIP (mainly, qnrS, qnrB and oqx), TET (mainly tet(A) and tet(M)) and SXT (mostly dfrIa group and sul3). In MDR isolates, integron presence and non-β-lactam resistance to TET, SXT and CHL were positively correlated. Overall, a high prevalence of MDR Enterobacteriaceae was found in livestock manure. The high gene diversity for antibiotic resistance identified in this study highlights the risk of MDR spread within the environment through manure use.
1. Introduction
The active ingredients of antibiotics (AB) legally prescribed in Portugal for veterinarian use in livestock prophylactic and metaphylactic treatments, amounted to 179,832 tons in 2013 [1]. Up to 90% of the ingested doses may be excreted unmodified or partially metabolised through urine and faeces [2]. Consequently, the excreta of AB-treated livestock became important reservoirs of AB residues, antibiotic-resistant bacteria (ARB) and antibiotic-resistance genes (ARGs) [3], where horizontal gene transference plays an essential role in the acquisition, spreading and assembly of various ARGs. Although integrons are not considered genetic elements per se, their location on plasmids and transposons enables gene transmission in an inter- and intra-species manner in a single event. For this reason, they are increasingly reported worldwide, especially among Enterobacteriaceae [4,5]. Large volumes of excreta enriched with Enterobacteriaceae end up as manure, slurry and wastewater, potentially vectoring these emergent contaminants. As such, they pose significant environmental concerns.
The slurry and the manure of intensively reared animals, composted or raw, are generally spread into soil for fertilisation; however, animal manure and wastewater are regarded as important potential AB-resistance reservoirs. Therefore, this reuse practice can lead to an increase of those pollutants in soil [6,7] and in endophytic bacteria of crops grown in manured soil [8]. The excreta ARB and ARGs can be spread over longer distances through other routes, such as anemophily [9], entomophily [10] or water run-off from farms and rural settlements, posing serious health problems. Despite the awareness about these spread routes, there are no available cost-effective tools to control the contaminants in municipal wastewater treatment plants (WWTPs) and livestock manure [11].
After recognising the importance of these emergent concerning contaminants, countries are developing their own national strategies to implement the Global Action Plan on antimicrobial resistance [12,13]. Moreover, the recent awareness that the health of humans and animals are inseparably interconnected with their environment led to an integrated One Health approach, particularly focusing on food safety, zoonosis surveillance and ARB control [14]. Dependence on livestock animals is considered one of the most critical pathogens associated risk factors to human given the transmission of AB resistance that occurs through consumption of contaminated animal products. Enterobacteriaceae is the major family associated with AB resistance and this is particularly relevant to this issue [15].
Understanding the diversity and the distribution of ARB and ARGs at a farm level is of great importance in order to identify the genetic background of this problem and, thus, educate and inform farm management. This study aimed to assess the level of contamination of livestock enterprises and the meat industry, in the central region of Portugal, through identification of multidrug resistant (MDR) isolates in manure. The analysis focused on the ARB and ARGs in resistant Enterobacteriaceae.
2. Materials and Methods
2.1. Study Area
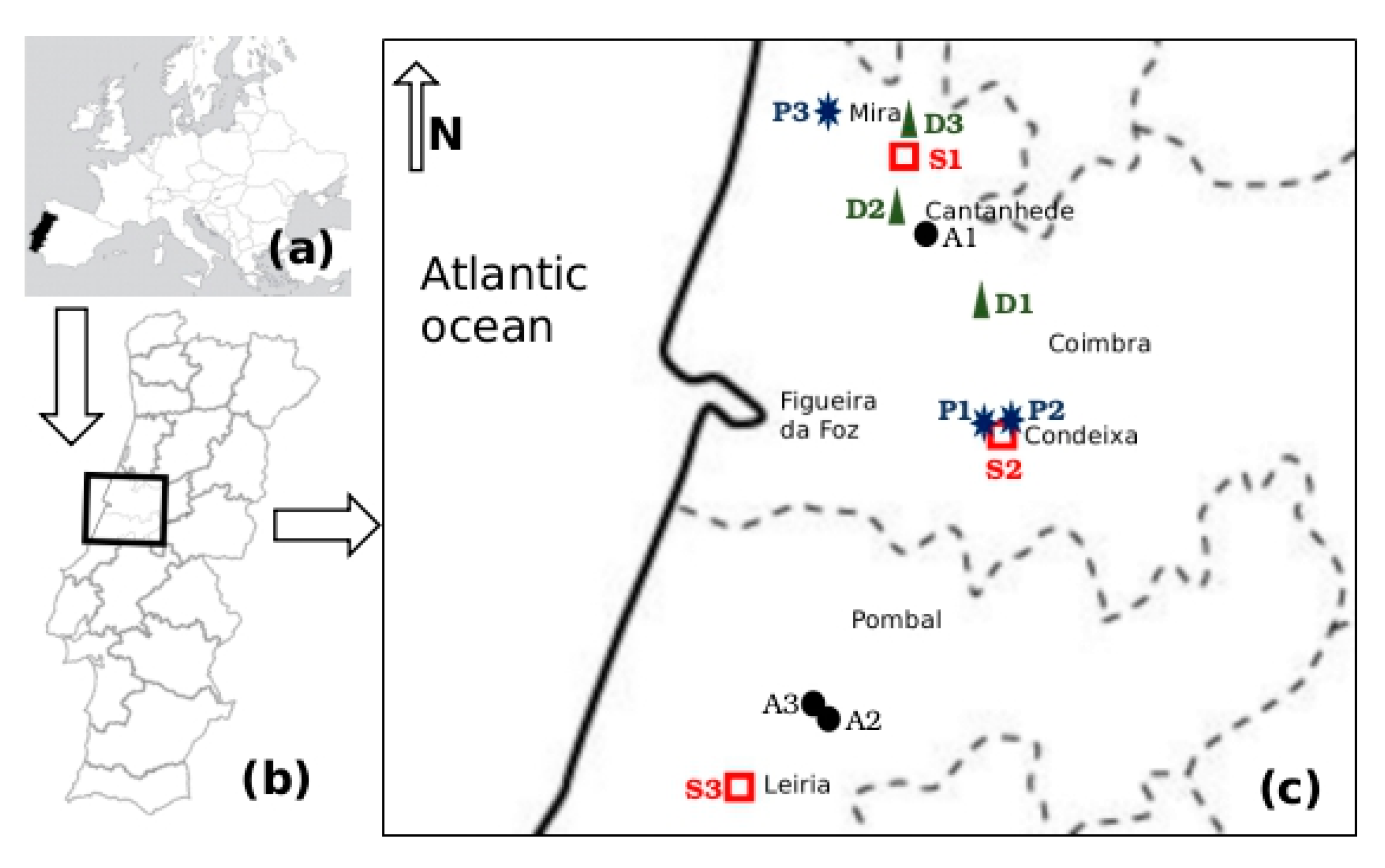
The study was conducted in three farm types and slaughterhouses representative of the livestock production sector in the central region of Portugal. The 12 selected small-scale enterprises included intensive raising of dairy cattle (D1, D2 and D3), poultry (A1, A2 and A3), pig (P1, P2 and P3) and three slaughterhouse companies (S1, S2 and S3). These sampling sites are located in the Coimbra and Leiria administrative regions, within Lis, Mondego and Vouga watersheds, at altitudes between 100 m and 200 m, higher than the flat alluvial river valleys (Figure 1).
As per information obtained from the farms’ personnel, all animals had adequate veterinary monitoring and a few, when necessary, received medical treatments. The manure and slurry generated in these farms were either composted or directly used as raw soil fertilizers in the animal feed crop fields of each enterprise. According to the slaughterhouse personnel, all animals were subjected to the required withdrawal time as per public health safety regulations and the defined holding time before slaughter was followed. Each slaughterhouse had its own protocol for treating wastewater.
2.2. Sampling
Between March 2016 and March 2017, three samples were collected per site. Dairy cows or swine manure samples were collected from open tanks with different maturation ages, whereas poultry manure from was collected from waste pools.
In the slaughterhouses, samples were collected from reservoirs holding the wastewaters generated by the daily slaughter of animals. The number of animals slaughtered daily is presented in Table 1.
Samples were collected in sterile plastic bottles or bags and maintained at 4 °C until the microbiological processing, which was always carried out within 4 h after sampling. In the laboratory, the three samples from each site were vigorously mixed and processed conjointly.
2.3. Microbiological Analysis and Phenotypic Characterisation
The enumeration, detection and identification of bacteria was carried out according to protocols previously described [16]. Decimal dilutions of the samples were prepared in sterile saline 0.9% NaCl. From each dilution, 100 mL were filtered through cellulose membranes of 0.45 µm (Millipore, Bedford, MA, USA) and the filters placed on the surface of selective medium for Enterobacteriaceae VRBG (Violet Red Bile Glucose) agar (Oxoid, Hampshire, England). The isolates were counted after overnight aerobic incubation at 37 °C. The colonies with different morphotypes were selected, picked out three times and their purity further confirmed microscopically. Species identification was performed according to the manufacturer’s recommendations by the standard API 20E galleries (BioMerieux, Marcy l’Etoile, Lyon, France).
Thirteen AB were selected for the phenotypic characterisation the isolates. These were chosen to represent the main AB classes used in human medicine and livestock production in Portugal, namely: amoxicillin/clavulanic acid combination (AMC) 30 μg/10 μg, respectively; ceftazidime (CAZ) 30 μg; cefotaxime (CTX) 30 μg; cefpirome (CPO) 30 μg; aztreonam (ATM) 30 μg; cefoxitin (FOX) 30 μg; imipenem (IPM) 10 μg; meropenem (MEM) 10 μg; chloramphenicol (CHL) 30 μg; gentamicin (GEN) 10 μg; ciprofloxacin (CIP) 5 μg; trimethoprim/sulfamethoxazol (SXT) combination (1:19) and tetracycline (TET) 30 μg. The disk diffusion Kirby-Bauer method was performed in agreement with the guidelines for antimicrobial susceptibility tests defined by the Clinical Laboratory Standards Institute [17], using Escherichia coli ATCC25922 (Liofilchem S.R.L., Roseto degli Abruzzi, Italy), E. coli J53-AzR (provided by Instituto de Ciências Biomédicas Abel Salazar, Portugal) and E. coli HB101-StpR–(Bio-Rad Laboratories Lda, Lisbon, Portugal) as quality controls, Mueller-Hinton agar and AB disks from Oxoid (Hampshire, England). The isolates with a resistance phenotype against three or more structurally unrelated antimicrobial agents were defined as multidrug resistant (MDR) [18].
2.4. Multiplex PCR (Polymerase Chain Reaction) for Genes Detection
The screening for resistance genes was focused on a subset of isolates chosen according to their phenotypic profile of resistance. The presence of the most frequent Enterobacteriaceae resistance genes in these isolates was determined by different multiplex PCR assays. Total DNA of these isolates was extracted as described by Amador et al [19] and 2 μL of each was subjected to multiplex PCR in a 25 μL reaction mixture containing 1× PCR buffer (200 mM Tris-HCl, pH 8.4, 500 mM KCl) and, according to the target gene, a variable concentration of primers (Table S1), MgCl2, dNTPs and 0.5 U of Taq DNA polymerase (Invitrogen, Thermo Fisher Scientific, Carlsbad, California, USA) (Table S2). The PCR conditions described by referenced authors in Table S1 were modified as specified in Table S2.
The chloramphenicol-resistance genes were detected by a single multiplex PCR with four forward primers and one reverse primer to target four genes, namely, catI, catII, catIII and catIV. The detection of sul1, sul2 and sul3 genes, that confer resistance to sulphonamide, was performed by two multiplex PCR assays given the different annealing temperature for sul3 gene amplification (Table S2).
The search for 14 genes involved in three tetracycline resistance mechanisms, namely efflux pump, ribosomal protection and tetracycline enzymatic alteration, was performed by two multiplex PCRs, one targeting genes tet(A), tet(E), tet(G), tet(K), tet(L), tet(M), tet(O) and tet(S) and the other tet(B), tet(C), tet(D), tet(Q), tetA(P) and tet(X).
For the detection of genes conferring resistance to trimethoprim, four distinct multiplex PCRs were performed according to the dfr gene group. The first PCR was used to detect genes belonging to groups Ia, (dfrA1, dfrA15, dfrA15b, dfrA16, dfrA16b, dfrA28), Ib (dfrA8) and Ic (dfrA12, dfrA13, dfrA21, dfrA22); the second for the group genes IIa (dfrA5, dfrA14, dfrA25, dfrA27), IIb (dfrA7, dfrA17) and IIc (dfrA3b); the third for groups III and IV, namely, IIIa (dfrA3), IIIb (dfrA10), IIIc (dfrA26), IVa (dfrA6), IVb (dfrA24), IVc (dfrA23); and the fourth for the groups Va (dfrB1, dfrB2, dfrB3, dfrB4, dfrB5, dfrB6), Vb (dfrA9), Vc (dfrA19) and Vd (dfrA20) [20].
Regarding plasmid-mediated quinolone resistance, nine genes responsible for conferring three different resistance mechanisms were analysed, namely, DNA gyrase protection from the action of the quinolones (qnrA, qnrB, qnrS, qnrC and qnrD), AB acetylation (aac(6’)-Ib-cr) and efflux pumps production (qepA, oqxA, oqxB). For this purpose, four different multiplex PCR assays were performed according to the annealing temperature (Table S2); one for the qnrA, qnrB and aac(6’)-Ib-cr; one for qnrD, qnrC and qepA; one for qnrS and oqxA; and a last one for oqxB gene.
To determine whether the ARGs were putatively disseminated via mobile genetic elements, three integrons classes, known to circulate in food-animal Enterobacteriaceae, were searched among the MDR isolates under study. Classes 1 and 2 of these genes were chosen for being most frequent among Enterobacteriaceae, whereas class 3 was chosen for being presently considered an emergent [21]. For this reason, a PCR was performed to detect intI1, intI2 and intI3 genes.
Multiplex PCRs were performed in a thermocycler (iCycler, Bio-Rad, Thermal Cycler, Hercules, CA, USA) and the amplification products obtained were separated by electrophoresis on a 1% agarose gel (BioRad), stained with ethidium bromide (125 µg/mL) and visualized under a UV transillumination (Vilber Lourmat, Marne-la-Vallé, France).
3. Results and Discussion
MDR information is crucial to support environmental and public health mitigation actions. Environmental systems have very complex microbiome communities, which are difficult to study due to the inability to artificially culture the vast majority of the species present in a given sample. Metagenomic approaches have overcome this difficulty, revealing the vast genetic diversity of the collective molecular signature of the samples’ resistomes. However, these molecular methods alone do not allow attributing bacterial ARG to a single organism and therefore cannot be used to recognise MDR. This study is based on an organismal approach, selecting the in vitro cultivable ARB, specifically with the Enterobacteriaceae family. Consequently, the isolates characterized are a slice of the sample bacteria: the AB resistant Enterobacteriaceae.
3.1. Microbiological Parameters
The higher enumeration of the Enterobacteriaceae was recorded in manure samples from poultry farms when compared to those of slaughterhouse, swine and dairy cows (four orders of magnitude smaller than poultry). The average temperature observed in the samples ranged between 9.4–24.0 °C. The pH values of poultry and pig manure samples were neutral, while dairy cows manure slightly alkaline. Slaughterhouse samples varied from neutral to acid (Table 1).
Out of the 331 isolates recovered from 12 sampling sites, grouped by four enterprise types, 90 were originated from poultry, 71 from dairy, 81 from pig and 89 from slaughterhouse samples. The Enterobacteriaceae species, detected, by decreasing order of prevalence, were: E. coli, unidentified isolates, Citrobacter freundii, Raoultella ornithinolytica, Salmonella enterica, C. koseri, Enterobacter cloacae, Morganella morganii, C. braakii, E. hermannii, E. vulneris, Klebsiella oxytoca, Kluyvera spp. A higher frequency of E. coli was registered in samples from slaughterhouses, pig and poultry farms, whereas C. freundii was more prevalent in dairy farm samples (Table 2).
3.2. Antibiotic Resistance Profile
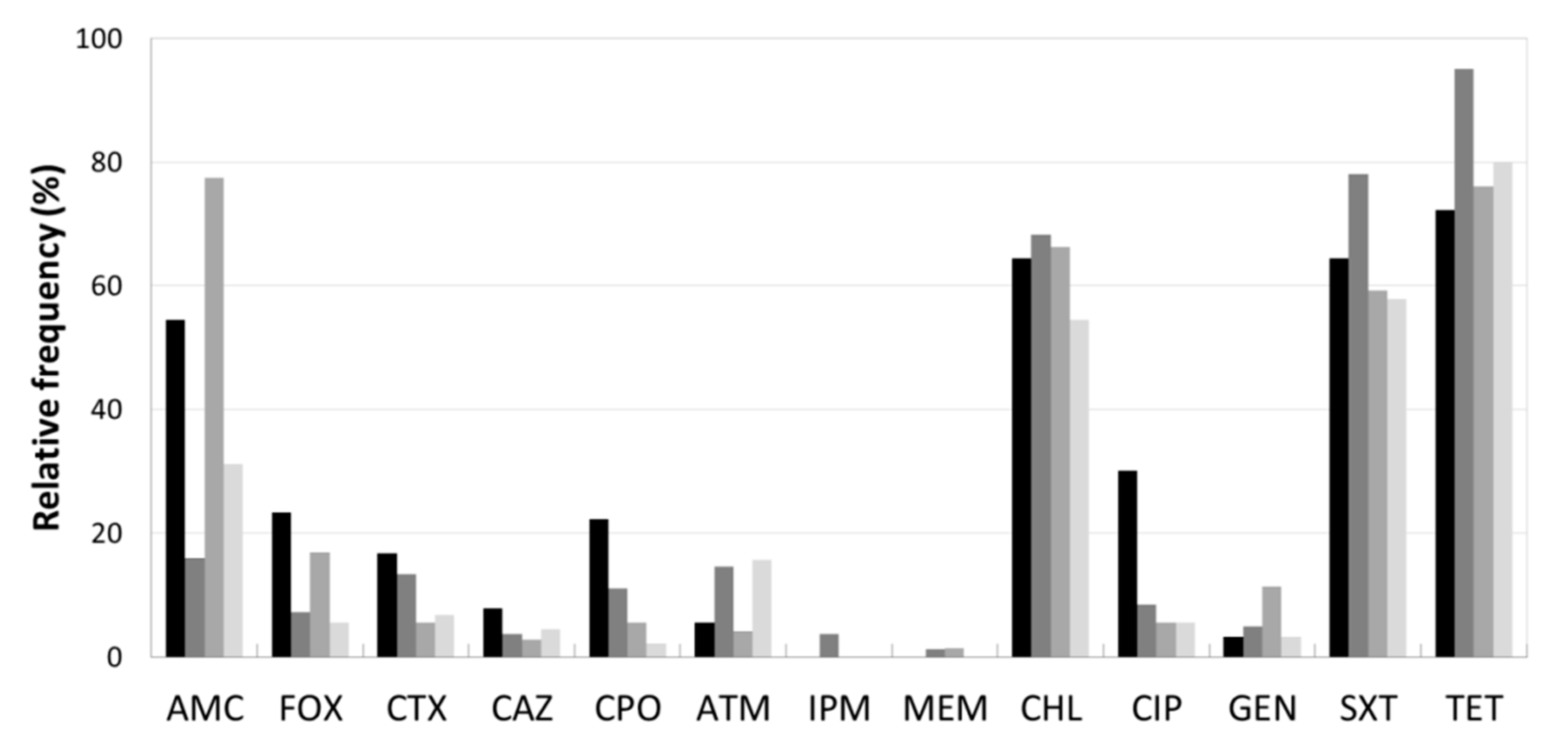
The analysis of AB resistance profiles of the isolates under study revealed by decreasing frequency order, among the β-lactam group, AMC, FOX, CPO, CTX, ATM and CAZ. The carbapenems, IPM and MEM, had the lowest frequencies. With regards to the non-β-lactam group, TET, SXT and CHL revealed to be less effective, whereas CIP and GEN were the most effectives (Figure 2). These findings can be explained by the intensive and recurrent use of tetracyclines, sulphonamides, penicillins and 2nd, 3rd and 4th cephalosporins generation in livestock production in Portugal [1] and all over the world [22,23]. The last report of the national control plan of drug use in Portugal referred the tetracyclines (38.9%), penicillins (18.8%), quinolones (4.5%) and sulphonamides (3.4%), as the AB classes for veterinary use with higher sales [1]. Curiously, as chloramphenicol (CHL) was banned in livestock according to the European legislation [24], the low sales of amphenicols (0.7%), apparently not justify the high frequency of CHL resistant isolates.
The AB resistance patterns per enterprise type revealed that the isolates from poultry farms had the highest resistance rates to TET, SXT, CHL and AMC (Figure 2). These results are in agreement with reports from: (i) Ghana, with high resistance frequencies to TET (88.9%), sulphonamide (75.0%), ampicillin (69.4%) and trimethoprim (66.7%) [25]; (ii) Madagascar broil farms with 97.6% to TET [26]; (iii) Chinese carcasses with ampicillin (98.9%), CHL (92.2%), TET (78.9%) [27]; and (iv) Portuguese healthy chickens with TET (70%), ampicillin (63%) and CIP (49%), however, with lower frequencies for SXT (33%), CHL (12%) and AMC (18%) [28]. It is noteworthy a higher frequency of poultry isolates resistant to CIP (30.0%) compared with those from the other enterprise types. An increasing resistance to this AB in poultry farming have been reported, for example from Canada [29], China [30], Brazil [31] and India [32]. This raise of resistance to CIP might be due to the increasing administration of quinolones to treat avian infections [21].
No isolate resisted to MEM and IMP (Figure 2), which agrees with many other studies carried out in livestock environments, showing only sporadic reports of isolates resistant to carbapenemases [28,33].
Regarding pig farms, the resistance rates of the isolates were higher for TET, SXT and CHL, followed by the β-lactams AMC, ATM and CTX (Figure 2). The lower resistance frequencies to carbapenems, IPM and MEM were not surprising, as previously mentioned. Similar results were published from China with over 60% for TET and CHL and no resistances to carbapenem (IMP) [34]. In the same country, another study revealed identical results towards carbapenems but all isolates were resistant to TET, ampicillin, CAZ and CTX. However, contrary to our results, a lower incidence for CHL (26.3%) and SXT (52.6%) were detected [35]. The association among the resistance phenotypes ampicillin–doxycycline–TET–SXT was reported as the predominant in poultry and swine production in China [36].
Among dairy farm isolates, higher resistance prevalence was observed to AMC, followed by TET, CHL and SXT. A single isolate resistant to carbapenem, MEM was found (Figure 2). These results are in agreement with many studies carried out in dairy farms, which usually report high TET and β-lactamic group resistances [37] and low or null carbapenem resistance. However, disagreeing with most published results, SXT resistances were lower than to CHL.
The AB resistance patterns (TET, SXT, CHL and AMC) of slaughterhouses isolates were similar to those of the other three enterprise types. These are unsurprising results, once the majority of the animals slaughtered in the abattoirs in this region, are locally raised cattle and pigs. Moreover, these results are consistent with many other studies carried out in agricultural environments. For instance, a Portuguese study on S. enterica and E. coli strains isolated from pork and beef products revealed similar resistances to TET, sulfamethoxazole and ampicillin [38]. In addition, a study in swine faecal samples obtained from Spanish slaughterhouses, showed the following resistance percentages: TET (90%), AMC (78%); SXT (67.5%); CHL (26%); GEN (6%) and CTX (0%) [39].
The comprehensive analysis of AB resistances rates by enterprise type revealed differences between poultry and pig isolates compared to those of dairy cattle. This seems consistent with other results [21] also reporting a lower prevalence of resistant E. coli in cattle than in pig and poultry. The highest rates of resistance to all the AB were found among the E. coli and the unidentified Enterobacteriaceae isolates. Worth noting was the high resistance to CTX in Citrobacter freundii and to MEM in Kluyvera spp. (Figure S1).
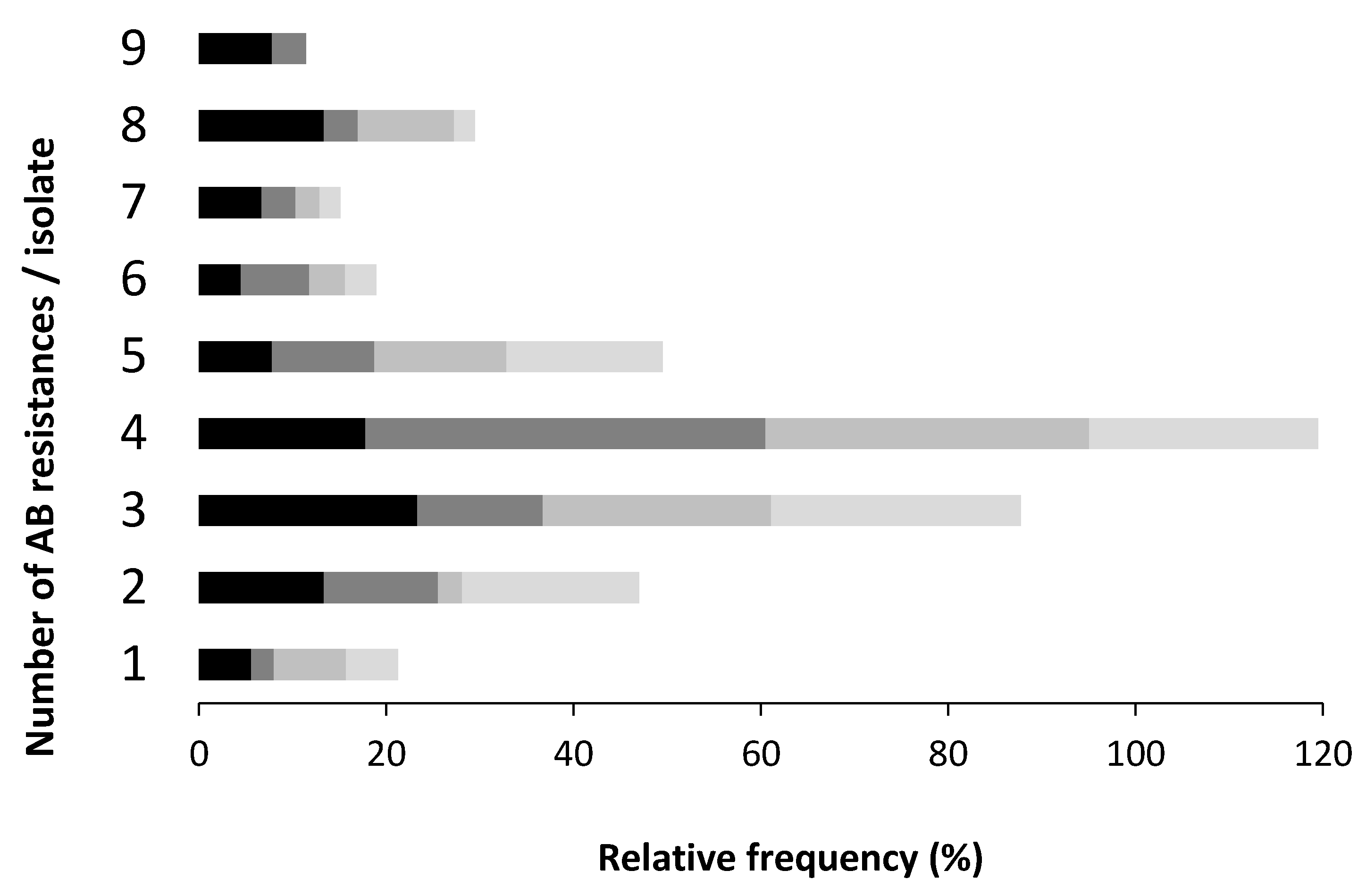
The majority of the isolates under study resisted simultaneously to three or four AB, regardless the sample origin. Poultry farm samples had the highest rate of resistant isolates to more than seven AB, when compared with slaughterhouse samples, where a higher rate of isolates simultaneously resistant to a lower number of AB was recorded (Figure 3).
The MDR rates observed in this study, poultry (71%), pig (79%), dairy cattle (69%) and slaughterhouse (63%), were higher if compared with other three Portuguese studies. One with pork samples, (60.7%) [40], other with chicken carcasses samples (56%) [28] and the third one with faecal and tissues samples of chickens and pigs (over 50%) [41]. The increasing trend of the AB co-resistant phenotypes numbers in humans and animals is a consequence over time of the intensive broad spectrum AB use and the introduction of newer compounds in human and veterinary medicine. This increase of MDR isolates, all over the world, can be explained by the mobile genetic elements in bacteria isolated from facilities farms, including WWTP, farm surrounding air, faeces, carcasses and organs of the animals, identified in numerous studies [21].
3.3. Screening of AB Resistance and Integrons Genes
Based on phenotypically characterised isolates, 70 MDR (22, 19, 13 and 16 isolated from poultry, pig, dairy and slaughterhouse samples, respectively) were selected for molecular characterisation (Table 2 and Table 3).
3.3.1. Chloramphenicol
All the isolates harbouring CHL resistance genes revealed an according phenotypic resistance profile (Table 2). However, in 61.4% (43/70) of those isolates no cat genes were detected. These results may be explained by the CHL resistance mechanisms, which are not exclusively determined by the cat genes family (I, II, III and IV). Although, the plasmid-mediated chloramphenicol acetyltransferase (cat) gene is the most common mechanism, other non-enzymatic resistance plasmidic genes, cmlA and floR, encoding efflux pumps, have also been identified in Enterobacteriaceae. The latter gene encodes a florfenicol/chloramphenicol transporter that confers resistance to both AB. A less common resistance mechanism involves mutations in 50S ribosomal subunit [42,43].
No CHL resistant isolates harboured genes catIII and catIV and those which had a gene, had either catI or catII (Table 3). The highest frequencies of cat genes were detected in samples from poultry and dairy cattle farms. The gene catI was more prevalent (21.4%) than catII (2.9%), the latter only represented by two M. morganii isolates from dairy samples (Table 3). Similarly, a study on MDR E. coli from Irish cattle and the farm environment, reported 9% CHL resistant isolates, mainly mediated by catI and floR [42]. Other reports also reveal that catI gene together with catIII gene is the most widely distributed, being catII less frequent [44]. However, a Korean study on fish pathogens isolates detected the catII gene as the most frequent, although catIV was also found in some isolates [45].
The high resistance to CHL found in this and other studies may have several explanatory hypotheses. Assuming that this AB is no longer used in human and veterinary medicine due to its proven chronic toxicity, the bacteria continue to resist because the determinant genes (often integrated in gene cassettes inserted in genetic elements, including integrons) could be transferred among them, even without high selective drug pressure. Another explanation is the licensed use of florfenicol for treatment of respiratory infections in cattle and pigs (Regulation (EEC) No 2377/90). This AB is a derivative of CHL, which shares one gene that determines the resistance mechanism [21].
3.3.2. Quinolones
The detection of the genes conferring resistance to the quinolones class, showed a higher incidence among isolates of poultry farms (Table 2). Quinolones, such as difloxacin, enrofloxacin, danofloxacin, marbofloxacin, pradofloxacin represent 4.5% of the AB sales, mainly for the avian sector in Portugal [1]. Likewise, ciprofloxacin (CIP), levofloxacin, norfloxacin, ofloxacin are widely used clinically to treat avian colibacillosis in many other countries [30]. The intensification of this AB class usage might explain the higher relative frequencies of CIP resistance in poultry isolates than others, sampled in other enterprise types (Figure 2). These quinolones class resistances might difficult the future therapy in this livestock sector.
No qnrA, oqxA, aac(6’)–Ib-cr and qepA genes were detected in any isolate. The qnrS gene was the most prevalent, mainly in the assayed poultry isolates (Table 3; Table 4), which is consistent with previous works carried out worldwide. An Italian work showed a higher frequency of the qnrS1 and qnrB19 genes for poultry isolates, respectively but no genes of qnrC, qnrD, qepA and aac(6’)-Ib-cr [46]. Other study involving E. coli isolates from chicken farms and turkeys at slaughterhouses in the Czech Republic, showed through screening of several genes, namely, qnrB1, qnrB4, qnrB8, qnrB10, qnrB19, qnrD, qnrS1, qnrS2, aac(6’)-Ib-cr and oqxAB, the same trend of results, a higher prevalence of qnrS1 followed by qnrB19 [47]. Another Chinese study, investigating the quinolones resistance in E. coli isolated from septicaemic broilers, namely, qnrA, qnrB, qnrS and aac(6’)-Ib-cr, recorded the highest prevalence of the gene aac(6’)-Ib-cr (36.0%), followed by qnrS (8.1%), qnrB (0.9%) and qnrA (0%) [30]. Conversely, in Nigerian poultry faecal samples no isolates containing the qnrS gene were registered, being oqxB the most prevalent [48].
Considering the pig farm isolates, our data revealed a higher prevalence for the qnrB gene, followed by the same prevalence for qnrS and oqxB, with 2 isolates each (Table 3). A study on sewage and soil adjacent to swine feedlots, showed a higher frequency for the qnrD, qepA and oqxB genes, whereas qnrS and oqxA were only present in the samples of residual waters [49].
3.3.3. Tetracycline
The screening of the 14 most frequent tet genes conferring resistance to TET revealed that genotypes are consistent with the phenotypes observed, excepting for three isolates, from pig and dairy farms (Table 2 and Table 3). The TET resistance in those isolates without tet genes, might be due to other resistance genes, not targeted in this study. There are at least 47 distinct genes identified, responsible for four main mechanisms by which the bacteria acquire resistance to tetracyclines [50].
Additionally, no isolates harboured the following genes: tet(D) and tet(G), responsible for the efflux pump resistance mechanism; tet(S) and tet(Q), responsible for the mechanism of ribosomal protection and tet(X), for the degradation of AB (Table 3). The most incident genes were tet(A) and tet(M) (Table 3), encoding an efflux pump and providing ribosomal protection, respectively. All the other surveyed genes had frequencies below or equal 10%. The gene tet(A) prevailed in isolates of all the four enterprise types; however, the second most frequent gene varied according to the sample source: tet(L) in poultry farms, tet(E) in dairy farms and tet(M) in pig farms and slaughterhouses. The prevalence of tet(A) gene in this study is in agreement with the dominant tet gene in different animal and environmental samples reported worldwide in a myriad of studies [42,51,52,53,54].
The isolates from poultry farms harboured the highest diversity of different tet genes (9/14), followed by slaughterhouses isolates with (7/14), conversely to those from dairy and pig farms, which showed lower genes diversity. The tet(C) gene was unique from poultry isolates group. Regarding the number of different tet genes per isolate, 14.3%, 45.7%, 27.1% and 7.1% of them contained, zero, one, two and three, respectively (Table 2). It should also be noted that one isolate of E. cloacae and one of E. coli from the poultry farms presented five different genes, while two other isolates, a S. enterica from laying hens samples and an E. coli, from the slaughterhouse, presented four genes. The analysis by sample source showed that the poultry farm isolates had the higher number of resistant isolates containing more tet gene types simultaneously. Identical results were reported in a study carried out in swine sewage ponds, where the isolates most often harbour multiple tet genes [55].
3.3.4. Sulphonamide
The most prevalent resistance genes to the sulphonamide class were sul3, followed by sul1 and at last sul2 (Table 3); however, this ranking varies according to the sample source. For instance, among the poultry farm isolates genes sul1 and sul3 were more frequent than sul2. Curiously, the isolates from dairy farms presented a higher incidence of sul1, whereas sul3 prevailed in slaughterhouses isolates and sul2 in those from pig farms. These genes prevalence can vary greatly with the matrixes, geographic region, as well as the species.
Our results are partially in accordance with two studies carried out in Denmark on isolates of E. coli resistant to sulphonamide obtained from swine faeces. The first work presented the highest frequency for the sul1 gene (55%), followed by sul2 (50%) and finally sul3 (11%) [56], whereas the second work reported sul2 gene, as the most prevalent (65%) [57]. On the other hand, a Portuguese study on resistances of E. coli isolated from carcasses and internal organs of healthy chickens from intensive farms detected 37 different resistance genes and the most common were tet(A) (72%) and sul1 (47%) [28]. A study in Cambodia on sulfamethoxazole-resistant isolates obtained from faecal samples of healthy chickens found a higher prevalence of the sul2 gene in E. coli (56%) and in Salmonella Corvallis (53%). However, among the Salmonella Albany the most prevalent gene was sul1 (54%) [58].
The analysis by isolates shows that 22.9% did not have a sul gene, 30.0% harboured one type of sul gene, 34.3% contained simultaneously two and 12.9% three sul genes (Table 2). It should be noted that the latter isolates were more prevalent in poultry and pig farm samples, which corroborates the idea that there is a higher AB selective pressure in these two livestock intensive raising systems.
3.3.5. Trimethoprim
Regarding the dfr genes detected in this study, a higher prevalence of the dfrIa group was found as a general trend in all enterprise type samples; however, being most prevalent in those of poultry farms (Table 3). Genes of dfrIc, dfrIIIb, dfrIIb groups were recorded but dfrIV and dfrV were not identified. Moreover, the dfrIIc group genes had the lowest detection rate, in only an isolate from a dairy farm sample. The isolates of poultry and dairy farms showed the highest diversity in this gene family. These results are in accordance with those of a Lithuanian study about the prevalence of dfr genes on E. coli isolates from human and animal clinical samples, concluding that 92% contained at least one of the six genes, dfrA1, dfrA5, dfrA8, dfrA12, dfrA14 and dfrA17 [20]. This gene nomenclature corresponds in our study to the genes of the group dfrIa, dfrIIa, dfrIb, dfrIc, dfrIIa, dfrIIb, respectively). The genes dfrA1 (dfrIa group) and dfrA17 (dfrIIb group) were more frequently found in clinical isolates, while the dfrA1 (dfrIa group) and dfrA14 (dfrIIa group) genes dominated in isolates of animal origin. The genes dfrA5 (dfrIIa group), dfrA12 (dfrIc group) and dfrA8 (dfrIb group) were detected at lower frequencies.
For the analysis of the genes involved in SXT resistance it is required to consider simultaneously the genes sul and dfr, since they act synergistically to confer resistance to an association of two antimicrobials, the sulfamethoxazole and the trimethoprim, both inhibitors of the biosynthetic pathway of folic acid. Resistance to both AB occurs by the acquisition of genes encoding enzymes that have no affinity for binding to these antimicrobial compounds, inactivating them [43].
All isolates listed in Table 2 with at least one type of gene from each family, sul or dfr exhibited SXT resistance, as for example some SXT resistant isolates without dfr genes but with one or more sul genes. This occurrence may be explained, as previously stated, by the presence of other genes that were not searched in this study, since there are more than 30 different known genes of trimethoprim resistance [20]. On the contrary, the phenotype of SXT resistant isolates lacking the sul genes might be explained by chromosomal mutations, resulting in molecular variants providing this resistance [43], even in absence of the resistance genes sul1, sul2 and sul3.
3.3.6. Integrons
The analysis of the AB resistance profiles reveals their clear MDR phenotype (Table 2). Those isolates with co-resistant phenotypes showed that the underlying resistance determinants of the non-β-lactams are based on three or more gene groups: tetracycline (tet(A) and tet(M)), sulfamethoxazole (sul3 and sul1), trimethoprim (dfrA1-like) and CHL (catI) (Table 4). Although it was not the scope of this work, the information on the Extended Spectrum Beta-Lactamases (ESBL) genotyping of these isolates would have been interesting, as these enzymes are often associated with the MDR [59]. Nevertheless, our results show the prevalence of a common resistance association pattern of several gene classes, which are known to be specially spread and persistent in livestock environments [21]. The widespread dissemination of the same MDR phenotypes mediating AB resistance was also reported by the European surveys on healthy food animals [22,60]. Other studies also show a predominant association among ampicillin-tetracycline-sulfamethoxazole/trimethoprim in Chinese poultry and pig farming [36].
The search for three integrons classes among the 65 MDR isolates under study, revealing 25 integrons (38.5%) of classes 1 and 2 and none of class 3. This latter, result is expected, as it is not prevalent in Enterobacteriaceae. The integrase gene IntI2 (class 2), initially present in Tn7 transposon and derivatives, is more frequent than the class 3 integrons [61]. Moreover, no isolate harboured simultaneously two integrons classes.
Class 1 and class 2, with 16 and 9 integrons, respectively were the most prevalent in isolates of all enterprise type samples (Table 3). These results are consistent with those of a Portuguese study conducted in poultry and pigs farms, having identified five isolates with class 1 of integrons and one isolate with class 2 [41]. Likewise, other US study on Enterobacteriaceae isolates obtained from fish, poultry, swine and cattle reported that 52% of them contained either class 1 or class 2 integrases, with the frequencies of 46% intI1, 6% intI2 and 5% intI1 plus intI2 [62]. Additionally, another study performed on MDR K. pneumoniae isolates from Iranian hospitals showed similar trends regarding the prevalence of the two integrons classes and a positive association between the class 1 integrons and MDR [63]. A Polish investigation on clinical samples from patients not undergoing AB therapy and from a WWTP detected 12.1% of the E. coli isolates with integrase genes, 10.9% of which with class 1, 1.4% with class 2 and 0% with class 3 [64].
Contrary to what was expected, the integrons were not prevalent among the isolates of poultry farm samples, which contradict the idea that the prevalence of integrons could reflect a strong AB pressure, selecting for those isolates with integrons [64]. In fact, the highest prevalence was recorded in pig farm isolates, with 11 integrons of both classes (Table 3). These frequency trends of the integrons classes are somewhat consistent with those found in E. coli isolated from samples of different livestock environments in Georgia reported for: (i) poultry isolates, intI1 (66%), intI2 (14%) and intI1 plus intI2 (11%); (ii) swine isolates, intI1 (86%), intI2 (0%) and intI1 plus intI2 (0%); and iii) beef cattle isolates, intI1 (75%), intI2 (23%) and intI1 plus intI2 (20%) [62].
Comparing the resistances to the non-β-lactamic AB with the presence of integrons in all the isolates under study, a positive relation between the SXT, CHL and TET resistant phenotype with the presence of integrons was observed. This relation is less evident with the quinolone CIP and aminoglycoside GEN (Table 4).
A Portuguese study on isolates from several polluted rivers, due to heavy domestic, industrial and agricultural influence, registered among the cefotaxime-resistant isolates, the prevalence of class 1 integrase (56.41% in those ESBL+) and the gene cassettes identified conferred resistance to β-lactams, trimethoprim and chloramphenicol [59]. The different sampling matrices may account to dissimilar results to ours, regarding CIP and GEN. However, several studies revealing the diversity of class 1 integrons and the gene cassettes in Enterobacteriaceae, indicate that the integrons generate new bindings of AB resistances, mainly in areas highly exposed to antimicrobial agents [65].
4. Conclusions
This study demonstrated a high prevalence of MDR Enterobacteriaceae and high resistance-gene diversity in livestock manure, possibly a consequence of intensive animal farming. Therefore, field amended with manure containing these biological contaminants can potentially contribute to an increased frequency of MDR bacteria in surrounding soil and aquatic ecosystems. As untreated manure is often used as a fertiliser of maize, rice and horticultural crops in agricultural fields of the Vouga, Lis and Mondego valleys, the common practices of nutrient and organic matter reuse for crop production has become a matter of concern, since the resistance of zoonotic bacteria should also be considered, as those contaminants may be transferred to livestock and humans through the consumption of raw crops fertilised with contaminated manure.
It should be important not only to reinforce the prudent, ethical and professional use of AB but also to improve appropriate treatments in the WWTP of the farms and slaughterhouses. Our findings provide an insight into the real impact of livestock farming manure to the environmental and public health problem of these contaminants in the Central Region of mainland Portugal.
Supplementary Materials
The following are available online at https://www.mdpi.com/2079-6382/8/1/23/s1, Figure S1: Relative frequency of antibiotics resistance by species. Table S1: Primers used for the identification of chloramphenicol (cat), trimethoprim (dfr), quinolones (qnr, aac(6’)-Ib, oqx, qep), sulphonamides (sul) and tetracyclines (tet) resistance genes and for integron class, Table S2: Multiplex PCR conditions for target genes.
Author Contributions
P.A. and I.D. conceived and design the study, carried out the sampling and the biological analysis, worked on the microbiological and molecular characterisation of antibiotic resistance bacteria. P.A. performed the data analysis and worked on the manuscript. C.P. and R.F. contributed with reagents and with I.D. worked on the manuscript.
Funding
This research received no external funding and the APC was funded by the authors.
Acknowledgments
This research was conducted within project Grupo Operacional para a Gestão da Água no Vale do Lis, PDR2020-1.0.1-FEADER-030911, funded by Program PDR2020, co-funded by FEDER, Innovation Measure, Portugal and by Research Centre for Natural Resources, Environment and Society—CERNAS—under project UID/AMB/00681/2019, funded by the Portuguese Science and Technology Foundation (FCT). The authors would like to thank Engineer Diogo Vilão for technical support and advice selecting the sampling sites.
Conflicts of Interest
The authors declare that there are no conflicts of interest.
References
- DGAV. Relatório Nacional de Monitorização do Consumo de Antimicrobianos Ano 2013—Portugal; Direcção-Geral de Alimentação e Veterinária/Direcção de Serviços de Medicamentos e Produtos de Uso Veterinário. Ministério da Agricultura, Mar, Ambiente e Ordenamento do Território; Governo de Portugal: Lisbon, Portugal, 2014; pp. 1–13.
- Jury, K.L.; Vancov, T.; Stuetz, R.M.; Khan, S.J. Antibiotic resistance dissemination and sewage treatment plants. In Current Research, Technology and Education Topics in Applied Microbiology and Microbial Biotechnology; Méndez-Vilas, A., Ed.; Formatex Research Center: Badajoz, Spain, 2010; pp. 509–519. ISBN 978-84-614-6194-3. [Google Scholar]
- Faldynova, M.; Videnska, P.; Havlickova, H.; Sisak, F.; Juricova, H.; Babak, V.; Steinhauser, L.; Rychlik, I. Prevalence of antibiotic resistance genes in faecal samples from cattle, pigs and poultry. Vet. Med. 2013, 58, 298–304. [Google Scholar] [CrossRef] [Green Version]
- Fluit, A.C.; Schmitz, F.J. Class 1 integrons, gene cassettes, mobility and epidemiology. Eur. J. Clin. Microbiol. Infect. Dis. 1999, 18, 761–770. [Google Scholar] [CrossRef] [PubMed]
- Kaushik, M.; Kumar, S.; Kapoor, R.K.; Virdi, J.S.; Gulati, P. Integrons in Enterobacteriaceae: Diversity, distribution and epidemiology. Int. J. Antimicrob. Agents 2017, 51, 167–176. [Google Scholar] [CrossRef]
- Martinez-Carballo, E.; Gonzalez-Barreiro, C.; Scharf, S.; Gans, O. Environmental monitoring study of selected veterinary antibiotics in animal manure and soils in Austria. Environ. Pollut. 2007, 148, 570–579. [Google Scholar] [CrossRef]
- Ji, X.L.; Shen, Q.H.; Liu, F.; Ma, J.; Xu, G.; Wang, Y.L.; Wu, M.H. Antibiotic resistance gene abundances associated with antibiotics and heavy metals in animal manures and agricultural soils adjacent to feedlots in Shanghai; China. J. Hazard. Mater. 2012, 235, 178–185. [Google Scholar] [CrossRef] [PubMed]
- Yang, Q.; Zhang, H.; Guo, Y.; Tian, T. Influence of Chicken Manure Fertilization on Antibiotic-Resistant Bacteria in Soil and the Endophytic Bacteria of Pakchoi. Int. J. Environ. Res. Public Health 2016, 13, 662. [Google Scholar] [CrossRef] [PubMed]
- Gao, L.; Tan, Y.; Zhang, X.; Hu, J.; Miao, Z.; Wei, L.; Chai, T. Emissions of Escherichia coli carrying extended-spectrum β-lactamase resistance from pig farms to the surrounding environment. Int. J. Environ. Res. Public Health 2015, 12, 4203–4213. [Google Scholar] [CrossRef]
- Songe, M.M.; Hang’ombe, B.M.; Knight-Jones, T.J.; Grace, D. Antimicrobial resistant enteropathogenic Escherichia coli and Salmonella spp. in houseflies infesting fish in food markets in Zambia. Int. J. Environ. Res. Public Health 2016, 14, 21. [Google Scholar] [CrossRef]
- Massé, D.I.; Saady, N.M.; Gilbert, Y. Potential of biological processes to eliminate antibiotics in livestock manure: An overview. Animals 2014, 4, 146–163. [Google Scholar] [CrossRef]
- WHO. Global Action Plan on Antimicrobial Resistance; World Health Organization: Geneva, Switzerland, 2015; p. 28. ISBN 978-92-4-150976-3. [Google Scholar]
- WHO. Antimicrobial Resistance: A Manual for Developing National Action Plans; Published by the WHO and the Food and Agriculture Organization of the United Nations and the World Organization for Animal Health; World Health Organization: Geneva, Switzerland, 2016; p. 26. ISBN 978-92-4-154953-0. [Google Scholar]
- King, L.J.; Anderson, L.R.; Blackmore, C.G.; Blackwell, M.J.; Lautner, E.A.; Marcus, L.C.; Meyer, T.E.; Monath, T.P.; Nave, J.E.; Ohle, J.; et al. Executive summary of the AVMA one health initiative task force report. J. Am. Vet. Med. Assoc. 2008, 233, 259–261. [Google Scholar] [CrossRef]
- Rousham, E.K.; Unicomb, L.; Islam, M.A. Human, animal and environmental contributors to antibiotic resistance in low-resource settings: Integrating behavioural, epidemiological and One Health approaches. Proc. R. Soc. Lond. B 2018, 285, 20180332. [Google Scholar] [CrossRef]
- Amador, P.; Fernandes, R.; Prudêncio, C.; Barreto, M.; Duarte, I. Antibiotic resistance in wastewater: Occurrence and fate of Enterobacteriaceae producers of Class A and Class C β-lactamases. J. Environ. Sci. Health A 2015, 50, 26–39. [Google Scholar] [CrossRef]
- CLSI. Performance Standards for Antimicrobial Susceptibility Testing, 26th ed.; CLSI Supplement M100S; Clinical and Laboratory Standards Institute: Wayne, PA, USA, 2016. [Google Scholar]
- Magiorakos, A.P.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; Paterson, D.L.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef]
- Amador, P.; Fernandes, R.; Brito, L.; Prudêncio, C. Antibiotic resistance in Enterobacteriaceae isolated from Portuguese deli meats. J. Food Saf. 2011, 31, 1–20. [Google Scholar] [CrossRef]
- Šeputienė, V.; Povilonis, J.; Ružauskas, M.; Pavilonis, A.; Sužiedėlienė, E. Prevalence of trimethoprim resistance genes in Escherichia coli isolates of human and animal origin in Lithuania. J. Med. Microbiol. 2010, 59, 315–322. [Google Scholar] [CrossRef]
- Szmolka, A.; Nagy, B. Multidrug resistant commensal Escherichia coli in animals and its impact for public health. Front. Microbiol. 2013, 4, 258. [Google Scholar] [CrossRef]
- De Jong, A.; Smet, A.; Ludwig, C.; Stephan, B.; De Graef, E.; Vanrobaeys, M.; Haesebrouck, F. Antimicrobial susceptibility of Salmonella isolates from healthy pigs and chickens (2008–2011). Vet. Microbiol. 2014, 171, 298–306. [Google Scholar] [CrossRef]
- Moawad, A.A.; Hotzel, H.; Awad, O.; Tomaso, H.; Neubauer, H.; Hafez, H.M.; El Adawy, H. Occurrence of Salmonella enterica and Escherichia coli in raw chicken and beef meat in northern Egypt and dissemination of their antibiotic resistance markers. Gut Pathog. 2017, 9, 57. [Google Scholar] [CrossRef]
- EC. Community procedure on measures to monitor certain substances and residues thereof in live animals and animal products and repealing. Directives 85/358/EEC and 86/469/EEC and Decisions 89/187/EEC and 91/664/EEC. Council Directive 96/23/EC of 29 April 1996. Offic. J. Eur. Communities 1996, 125, 10. [Google Scholar]
- Rasmussen, M.M.; Opintan, J.A.; Frimodt-Møller, N.; Styrishave, B. Beta-lactamase producing Escherichia coli isolates in imported and locally produced chicken meat from Ghana. PLoS ONE 2015, 10, e0139706. [Google Scholar] [CrossRef]
- Gay, N.; Leclaire, A.; Laval, M.; Miltgen, G.; Jégo, M.; Stéphane, R.; Jaubert, J.; Belmonte, O.; Cardinale, E. Risk factors of extended-spectrum β-lactamase producing Enterobacteriaceae occurrence in farms in Reunion, Madagascar and Mayotte Islands, 2016–2017. Vet. Sci. 2018, 5, 22. [Google Scholar] [CrossRef]
- Wu, H.; Wang, M.; Liu, Y.; Wang, X.; Wang, Y.; Lu, J.; Xu, H. Characterization of antimicrobial resistance in Klebsiella species isolated from chicken broilers. Int. J. Food Microbiol. 2016, 232, 95–102. [Google Scholar] [CrossRef]
- Mendonça, N.; Figueiredo, R.; Mendes, C.; Card, R.M.; Anjum, M.F.; Silva, G.J. Microarray evaluation of antimicrobial resistance and virulence of Escherichia coli isolates from Portuguese poultry. Antibiotics 2016, 5, 4. [Google Scholar] [CrossRef]
- Agunos, A.; Léger, D.; Avery, B.P.; Parmley, E.J.; Deckert, A.; Carson, C.A.; Dutil, L. Ciprofloxacin-resistant Campylobacter spp. in retail chicken, western Canada. Emerg. Infect. Dis. 2013, 19, 1121–1124. [Google Scholar] [CrossRef]
- Xie, R.; Huo, S.; Li, Y.; Chen, L.; Zhang, F.; Wu, X. Molecular epidemiological survey on quinolone resistance genotype and phenotype of Escherichia coli in septicemic broilers in Hebei, China. Poult. Sci. 2014, 93, 335–339. [Google Scholar] [CrossRef]
- Gouvêa, R.; dos Santos, F.F.; de Aquino, M.H.C. Fluoroquinolones in industrial poultry production, bacterial resistance and food residues: A review. Rev. Bras. Cienc. Avic. 2015, 17, 1–10. [Google Scholar] [CrossRef]
- Bhushan, C.; Khurana, A.; Sinha, R.; Nagaraju, M. Antibiotic Resistance in Poultry Environment: Spread of Resistance from Poultry Farm to Agricultural Field; Centre for Science and Environment: New Delhi, India, 2017; 36p, Available online: www.cseindia.org (accessed on 14 July 2018).
- Franco, A.; Leekitcharoenphon, P.; Feltrin, F.; Alba, P.; Cordaro, G.; Iurescia, M.; Tolli, R.; D’Incau, M.; Staffolani, M.; Di Giannatale, E.; et al. Emergence of a clonal lineage of multidrug-resistant ESBL-producing Salmonella Infantis transmitted from broilers and broiler meat to humans in Italy between 2011 and 2014. PLoS ONE 2015, 10, e0144802. [Google Scholar] [CrossRef]
- Gao, L.; Hu, J.; Zhang, X.; Wei, L.; Li, S.; Miao, Z.; Chai, T. Application of swine manure on agricultural fields contributes to extended-spectrum β-lactamase-producing Escherichia coli spread in Tai’an, China. Front. Microbiol. 2015, 6, 313. [Google Scholar] [CrossRef]
- Li, L.; Ye, L.; Yu, L.; Zhou, C.; Meng, H. Characterization of extended spectrum β-lactamase producing Enterobacteria and methicillin-resistant Staphylococcus aureus isolated from raw pork and cooked pork products in South China. J. Food Sci. 2016, 81, M1773–M1777. [Google Scholar] [CrossRef]
- Jiang, H.X.; Lü, D.H.; Chen, Z.L.; Wang, X.M.; Chen, J.R.; Liu, Y.H.; Liao, X.P.; Liu, J.H.; Zeng, Z.L. High prevalence and widespread distribution of multi-resistant Escherichia coli isolates in pigs and poultry in China. Vet. J. 2011, 187, 99–103. [Google Scholar] [CrossRef]
- Koovapra, S.; Bandyopadhyay, S.; Das, G.; Bhattacharyya, D.; Banerjee, J.; Mahanti, A.; Samanta, I.; Nanda, P.K.; Kumar, A.; Mukherjee, R.; et al. Molecular signature of extended spectrum β-lactamase producing Klebsiella pneumoniae isolated from bovine milk in eastern and north-eastern India. Infect. Genet. Evol. 2016, 44, 395–402. [Google Scholar] [CrossRef]
- Clemente, L.; Manageiro, V.; Jones-Dias, J.; Correia, C.; Patricia Themudo, P.; Albuquerque, T.; Geraldes, M.; Matos, F.; Almendra, C.; Ferreira, E.; et al. Antimicrobial susceptibility and oxymino-β-lactam resistance mechanisms in Salmonella enterica and Escherichia coli isolates from different animal sources. Res. Microbiol. 2015, 166, 574–583. [Google Scholar] [CrossRef]
- Marchant, M.; Vinué, L.; Torres, C.; Moreno, M.A. Change of integrons over time in Escherichia coli isolates recovered from healthy pigs and chickens. Vet. Microbiol. 2013, 163, 124–132. [Google Scholar] [CrossRef]
- Clemente, L.; Manageiro, V.; Ferreira, E.; Jones-Dias, D.; Correia, I.; Themudo, P.; Albuquerque, T.; Caniça, M. Occurrence of extended-spectrum β-lactamases among isolates of Salmonella enterica subsp. enterica from food-producing animals and food products, in Portugal. Int. J. Food Microbiol. 2013, 167, 221–228. [Google Scholar] [CrossRef]
- Jones-Dias, D.; Manageiro, V.; Martins, A.P.; Ferreira, E.; Caniça, M. New class 2 integron in 2-4 among IncI1-positive Escherichia coli isolates carrying ESBL and PMAβ genes from food animals in Portugal. Foodborne Pathog. Dis. 2015, 13, 36–39. [Google Scholar] [CrossRef]
- Karczmarczyk, M.; Walsh, C.; Slowey, R.; Leonard, N.; Fanning, S. Molecular characterization of multidrug-resistant Escherichia coli isolates from Irish cattle farms. Appl. Environ. Microbiol. 2011, 77, 7121–7127. [Google Scholar] [CrossRef]
- Frye, J.G.; Jackson, C.R. Genetic mechanisms of antimicrobial resistance identified in Salmonella enterica, Escherichia coli and Enteroccocus spp. isolated from U.S. food animals. Front. Microbiol. 2013, 4, 135. [Google Scholar] [CrossRef]
- Shaw, W.V. Chloramphenicol acetyltransferase: Enzymology and molecular biology. Crit. Rev. Biochem. 1983, 14, 1–46. [Google Scholar] [CrossRef]
- Yoo, M.H.; Huh, M.D.; Kim, E.H.; Lee, H.H.; Do Jeong, H. Characterization of chloramphenicol acetyltransferase gene by multiplex polymerase chain reaction in multidrug-resistant strains isolated from aquatic environments. Aquaculture 2003, 217, 11–21. [Google Scholar] [CrossRef]
- Niero, G.; Bortolaia, V.; Vanni, M.; Intorre, L.; Guardabassi, L.; Piccirillo, A. High diversity of genes and plasmids encoding resistance to third-generation cephalosporins and quinolones in clinical Escherichia coli from commercial poultry flocks in Italy. Vet. Microbiol. 2018, 216, 93–98. [Google Scholar] [CrossRef]
- Röderova, M.; Halova, D.; Papousek, I.; Dolejska, M.; Masarikova, M.; Hanulik, V.; Pudova, V.; Broz, P.; Htoutou-Sedlakova, M.; Sauer, P.; et al. Characteristics of quinolone resistance in Escherichia coli isolates from humans, animals and the environment in the Czech Republic. Front. Microbiol. 2017, 7, 2147. [Google Scholar] [CrossRef]
- Akinbami, O.R.; Olofinsae, S.; Ayeni, F.A. Prevalence of extended spectrum beta lactamase and plasmid mediated quinolone resistant genes in strains of Klebsiella pneumonia, Morganella morganii, Leclercia adecarboxylata and Citrobacter freundii isolated from poultry in South Western Nigeria. PeerJ 2018, 6, e5053. [Google Scholar] [CrossRef]
- Li, J.; Wang, T.; Shao, B.; Shen, J.; Wang, S.; Wu, Y. Plasmid-mediated quinolone resistance genes and antibiotic residues in wastewater and soil adjacent to swine feedlots: Potential transfer to agricultural lands. Environ. Health Perspect. 2012, 120, 1144–1149. [Google Scholar] [CrossRef]
- Nguyen, F.; Starosta, A.L.; Arenz, S.; Sohmen, D.; Dönhöfer, A.; Wilson, D.N. Tetracycline antibiotics and resistance mechanisms. Biol. Chem. 2014, 395, 559–575. [Google Scholar] [CrossRef]
- Zhang, S.; Gu, J.; Wang, C.; Wang, P.; Jiao, S.; He, Z.; Han, B. Characterization of antibiotics and antibiotic resistance genes on an ecological farm system. J. Chem. 2015, 2015, 526143. [Google Scholar] [CrossRef]
- Ghosh, S.; LaPara, T.M. The effects of subtherapeutic antibiotic use in farm animals on the proliferation and persistence of antibiotic resistance among soil bacteria. ISME J. 2007, 1, 191–203. [Google Scholar] [CrossRef] [Green Version]
- Koo, H.J.; Woo, G.J. Distribution and transferability of tetracycline resistance determinants in Escherichia coli isolated from meat and meat products. Int. J. Food Microbiol. 2011, 145, 407–413. [Google Scholar] [CrossRef]
- McNeece, G.; Naughton, V.; Woodward, M.J.; Dooley, J.S.G.; Naughton, P.J. Array based detection of antibiotic resistance genes in Gram negative bacteria isolated from retail poultry meat in the UK and Ireland. Int. J. Food Microbiol. 2014, 179, 24–32. [Google Scholar] [CrossRef]
- Chee-Sanford, J.; Maxwell, S.; Tsau, K.; Merrick, K.; Aminov, R. Antibiotic resistance in swine-manure-impacted environments. In Antimicrobial Resistance in the Environment; Keen, P.L., Mark, H.M.M., Eds.; John Wiley & Sons, Inc.: Hoboken, NJ, USA, 2012; pp. 203–223. ISBN 9780470905425. [Google Scholar]
- Hammerum, A.M.; Sandvang, D.; Andersen, S.R.; Seyfarth, A.M.; Porsbo, L.J.; Frimodt-Møller, N.; Heuer, O.E. Detection of sul1, sul2 and sul3 in sulphonamide resistant Escherichia coli isolates obtained from healthy humans, pork and pigs in Denmark. Int. J. Food Microbiol. 2006, 106, 235–237. [Google Scholar] [CrossRef]
- Wu, S.; Dalsgaard, A.; Hammerum, A.M.; Porsbo, L.J.; Jensen, L.B. Prevalence and characterization of plasmids carrying sulfonamide resistance genes among Escherichia coli from pigs, pig carcasses and human. Acta Vet. Scand. 2010, 52, 47. [Google Scholar] [CrossRef]
- Vuthy, Y.; Lay, K.S.; Seiha, H.; Kerleguer, A.; Aidara-Kane, A. Antibiotic susceptibility and molecular characterization of resistance genes among Escherichia coli and among Salmonella subsp. in chicken food chains. Asian Pac. J. Trop. Biomed. 2017, 7, 670–674. [Google Scholar] [CrossRef]
- Tacão, M.; Moura, A.; Correia, A.; Henriques, I. Co-resistance to different classes of antibiotics among ESBL-producers from aquatic systems. Water Res. 2014, 48, 100–107. [Google Scholar] [CrossRef]
- EFSA—European Food Safety Authority and European Centre for Disease Prevention and Control (EFSA and ECDC). The European Union Summary Report on antimicrobial resistance in zoonotic and indicator bacteria from humans, animals and food in 2010. EFSA J. 2012, 10, 2598. [Google Scholar] [CrossRef] [Green Version]
- Dillon, B.; Thomas, L.; Mohmand, G.; Zelynski, A.; Iredell, J. Multiplex PCR for screening of integrons in bacterial lysates. J. Microbiol. Methods 2005, 62, 221–232. [Google Scholar] [CrossRef]
- Goldstein, C.; Lee, M.D.; Sanchez, S.; Hudson, C.; Phillips, B.; Register, B.; Grady, M.; Liebert, C.; Summers, A.O.; White, D.G.; et al. Incidence of class 1 and 2 integrases in clinical and commensal bacteria from livestock, companion animals and exotics. Antimicrob. Agents Chemother. 2001, 45, 723–726. [Google Scholar] [CrossRef]
- Akya, A.; Azam Elahi, A.; Chegenelorestani, R.; Rezaee, M. Dissemination of multidrug-resistant, class I and II integrons and molecular typing of CTX-M-producing Klebsiella pneumoniae. Int. J. Appl. Basic Med. Res. 2018, 8, 100–105. [Google Scholar] [CrossRef]
- Koczura, R.; Mokracka, J.; Jabłońska, L.; Gozdecka, E.; Kubek, M.; Kaznowski, A. Antimicrobial resistance of integron-harboring Escherichia coli isolates from clinical samples, wastewater treatment plant and river water. Sci. Total Environ. 2012, 414, 680–685. [Google Scholar] [CrossRef]
- Ravi, A.; Avershina, E.; Ludvigsen, J.; L’Abée-Lund, T.M.; Rudi, K. Integrons in the intestinal microbiota as reservoirs for transmission of antibiotic resistance genes. Pathogens 2014, 3, 238–248. [Google Scholar] [CrossRef]
Figure 1.
Location of sampling sites depicted on (a) a map of Europe (approximate scale 1:100,000,000, (b) a map of Portugal (approximate scale 1:10,000,000) and (c) a map of Coimbra and Leiria administrative regions (dashed lines) (approximate scale 1:1,000,000). Enterprise types are represented as follows: ![Antibiotics 08 00023 i001]() dairy cattle farm;
dairy cattle farm; ![Antibiotics 08 00023 i002]() poultry farm;
poultry farm; ![Antibiotics 08 00023 i003]() slaughterhouse;
slaughterhouse; ![Antibiotics 08 00023 i004]() pig farm.
pig farm.
 dairy cattle farm;
dairy cattle farm;  poultry farm;
poultry farm;  slaughterhouse;
slaughterhouse;  pig farm.
pig farm.
Figure 1.
Location of sampling sites depicted on (a) a map of Europe (approximate scale 1:100,000,000, (b) a map of Portugal (approximate scale 1:10,000,000) and (c) a map of Coimbra and Leiria administrative regions (dashed lines) (approximate scale 1:1,000,000). Enterprise types are represented as follows: ![Antibiotics 08 00023 i001]() dairy cattle farm;
dairy cattle farm; ![Antibiotics 08 00023 i002]() poultry farm;
poultry farm; ![Antibiotics 08 00023 i003]() slaughterhouse;
slaughterhouse; ![Antibiotics 08 00023 i004]() pig farm.
pig farm.
dairy cattle farm; poultry farm; slaughterhouse; pig farm.
Figure 2.
Antibiotic resistance of Enterobacteriaceae isolates obtained from 12 sampling sites. Relative frequency of resistance patterns exhibited by resistant and intermediate resistant isolates to 13 AB. Tested AB: eight β-lactams: amoxicillin/clavulanic acid (AMC), ceftazidime (CAZ), cefotaxime (CTX), cefpirome (CPO), aztreonam (ATM), cefoxitin (FOX), imipenem (IPM), meropenem (MEM) and non-β-lactams: chloramphenicol (CHL), tetracycline (TET), gentamicin (GEN), trimethoprim/sulfamethoxazol (SXT) and ciprofloxacin (CIP). Enterprise type: ![Antibiotics 08 00023 i005]() poultry,
poultry, ![Antibiotics 08 00023 i006]() pig,
pig, ![Antibiotics 08 00023 i007]() dairy,
dairy, ![Antibiotics 08 00023 i008]() slaughterhouse.
slaughterhouse.
 poultry,
poultry,  pig,
pig,  dairy,
dairy,  slaughterhouse.
slaughterhouse.
Figure 2.
Antibiotic resistance of Enterobacteriaceae isolates obtained from 12 sampling sites. Relative frequency of resistance patterns exhibited by resistant and intermediate resistant isolates to 13 AB. Tested AB: eight β-lactams: amoxicillin/clavulanic acid (AMC), ceftazidime (CAZ), cefotaxime (CTX), cefpirome (CPO), aztreonam (ATM), cefoxitin (FOX), imipenem (IPM), meropenem (MEM) and non-β-lactams: chloramphenicol (CHL), tetracycline (TET), gentamicin (GEN), trimethoprim/sulfamethoxazol (SXT) and ciprofloxacin (CIP). Enterprise type: ![Antibiotics 08 00023 i005]() poultry,
poultry, ![Antibiotics 08 00023 i006]() pig,
pig, ![Antibiotics 08 00023 i007]() dairy,
dairy, ![Antibiotics 08 00023 i008]() slaughterhouse.
slaughterhouse.
poultry, pig, dairy, slaughterhouse.
Figure 3.
Relative frequency of the number of antibiotics to which each isolate resists. Enterprise type: ![Antibiotics 08 00023 i005]() poultry,
poultry, ![Antibiotics 08 00023 i006]() pig,
pig, ![Antibiotics 08 00023 i007]() dairy,
dairy, ![Antibiotics 08 00023 i008]() slaughterhouse.
slaughterhouse.
poultry, pig, dairy, slaughterhouse.
Figure 3.
Relative frequency of the number of antibiotics to which each isolate resists. Enterprise type: ![Antibiotics 08 00023 i005]() poultry,
poultry, ![Antibiotics 08 00023 i006]() pig,
pig, ![Antibiotics 08 00023 i007]() dairy,
dairy, ![Antibiotics 08 00023 i008]() slaughterhouse.
slaughterhouse.
poultry, pig, dairy, slaughterhouse.

{kind=link}
{kind=link}
{kind=link}
Table 1.
Animal heads per sampling site, average microbiological and physical-chemical parameters of samples.
Table 1.
Animal heads per sampling site, average microbiological and physical-chemical parameters of samples.
| Characterisation of Sampling Sites | Characterisation of Samples | ||||
|---|---|---|---|---|---|
| Site Code | Enterprise Type | Animal Heads | Enterobacteriaceae (cfu/mL) | Temperature (°C) | pH |
| A1 | Poultry | 10,000 chicks Legorne/pavilion | 1.4 × 107 | 10.2 | 6.80 |
| A2 | Poultry | 72,000 caged laying hens Legorne | 6.9 × 109 | 24.1 | 7.24 |
| A3 | Poultry | 28,000 soil, cage-free laying hens Legorne | 7.6 × 109 | 24.0 | 7.06 |
| D1 | Dairy cattle | 100 Holstein Friesian | 1.7 × 105 | 9.4 | 8.81 |
| D2 | Dairy cattle | 30 Holstein Friesian | 5.7 × 105 | 14.6 | 7.46 |
| D3 | Dairy cattle | 54 Holstein Friesian | 2.4 × 105 | 16.9 | 8.47 |
| P1 | Pig | 48 breeding sow Large Write | 3.4 × 105 | 14.7 | 7.71 |
| P2 | Pig | 700 breeding sow, 5300 piglets Large Write | 5.0 × 105 | 14.0 | 7.16 |
| P3 | Pig | 1640 fattening pigs Large Write | 1.7 × 106 | 13.8 | 7.33 |
| S1 | Slaughterhouse | 505 piglets, 62 cattle (daily abattoir) | 1.0 × 106 | 14.1 | 6.52 |
| S2 | Slaughterhouse | 381 hogs, 740 cattle (daily abattoir) | 1.7 × 106 | 16.0 | 4.92 |
| S3 | Slaughterhouse | 190 hogs, 40 cattle (daily abattoir) | 1.1 × 109 | 23.8 | 7.31 |
Temperature (°C)—determined by thermometry; pH—determined by potentiometry.
Table 2.
List of AB resistance profile, AB resistance and integrons genes harboured per isolate species and sample origin.
Table 2.
List of AB resistance profile, AB resistance and integrons genes harboured per isolate species and sample origin.
| Sample (a) | Species | Resistance Phenotype (b) | AB Resistance Genes | intI | ||||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| AMC | FOX | CTX | CAZ | CPO | ATM | IPM | MEM | CHL | CIP | GEN | SXT | TET | cat | qnr/oqx | tet | sul | dfr | |||
| Poultry | S. enterica | x | x | x | x | x | x | x | x | - | - | A, M, L, K | 1, 2, 3 | Ia, IIb | - | |||||
| Enterobacter cloacae | x | x | x | x | x | x | x | x | x | I | D, S | A, E, B, K, O | 1, 3 | Ia | - | |||||
| E. hermannii | x | x | x | x | x | x | x | x | x | - | - | C | 1, 2 | Ic | - | |||||
| C. freundii | x | x | x | x | x | x | x | x | I | C, S | A | 1, 2 | Ia, IIb, IIIc | - | ||||||
| C. freundii | x | x | x | x | x | x | x | x | I | S | A, L, K | 1, 2 | Ia, Ic, IIIa, IIIc | - | ||||||
| R. ornithinolytica | x | x | x | x | - | nt | - | 3 | Ia | 1 | ||||||||||
| E. coli | x | x | x | x | x | x | x | x | x | - | nt | A, C | 1, 2, 3 | Ia, Ib, IIIa | - | |||||
| E. coli | x | x | x | x | x | - | nt | A, E, C, M, L | 2, 3 | Ia, IIa | - | |||||||||
| E. coli | x | x | x | x | x | x | x | - | nt | A | 1, 3 | Ia, Ib | - | |||||||
| E. coli | x | x | x | x | x | x | x | x | - | nt | A | 1, 3 | Ia, Ib, Ic, IIIa | - | ||||||
| E. coli | x | x | x | x | x | - | nt | A | 2 | Ia, Ic, IIb | - | |||||||||
| E. coli | x | x | x | x | x | x | x | - | nt | A | 3 | Ia, Ib | - | |||||||
| nt | x | x | x | x | x | x | x | x | I | S | A, M | 1, 2, 3 | Ia, IIb | - | ||||||
| nt | x | x | x | x | x | x | x | x | x | I | S | A, A(P) | 1, 3 | Ia | - | |||||
| nt | x | x | x | x | x | x | x | - | C, S | A | 1, 3 | Ia | 1 | |||||||
| nt | x | x | x | x | x | x | x | x | I | C, S | A | 1, 2, 3 | Ia | 1 | ||||||
| nt | x | x | x | x | x | x | I | B, S | A | 1, 3 | - | I | ||||||||
| nt | x | x | x | x | x | x | I | B, C, D, S | - | 1 | Ia | 1 | ||||||||
| nt | x | x | x | x | - | nt | A, A(P), M | - | IIb | - | ||||||||||
| nt | x | x | x | x | x | x | - | nt | A | - | - | - | ||||||||
| nt | x | x | x | x | x | - | B, S | A, L, K | 3 | - | - | |||||||||
| nt | x | x | x | x | x | - | B, S | A, L | 1, 3 | Ia, Ic, IIb | - | |||||||||
| Pig | S. enterica | x | x | x | x | x | - | nt | A | - | Ia, Ic | 2 | ||||||||
| K. oxytoca | x | x | x | x | x | - | oqxB | A | 1, 2, 3 | Ic, IIa | - | |||||||||
| R. ornithinolytica | x | x | x | x | x | x | x | I | B | M | 2, 3 | - | - | |||||||
| Kluyvera spp. | x | x | x | x | x | x | x | x | x | - | nt | A | 2 | Ia | 2 | |||||
| E. coli | x | x | x | x | - | nt | A, B | - | Ic | - | ||||||||||
| E. coli | x | x | x | x | x | x | x | I | - | A | 1, 2, 3 | Ia, Ic | 1 | |||||||
| E. coli | x | x | x | x | x | x | - | B | A | 1, 2, 3 | Ia | 2 | ||||||||
| E. coli | x | x | x | x | x | x | - | - | - | - | Ia, IIIb | - | ||||||||
| E. coli | x | x | x | x | x | x | x | x | I | - | B, M | 3 | Ia, IIa | 2 | ||||||
| E. coli | x | x | x | x | x | x | x | - | B | A | - | Ia | - | |||||||
| E. coli | x | x | x | x | x | x | x | x | x | - | B | A | - | Ia, IIIb | 1 | |||||
| E. coli | x | x | x | x | x | x | x | x | x | - | B | A | 1, 2 | Ia, Ic | - | |||||
| nt | x | x | x | x | - | oqxB | A, M | 1, 2, 3 | - | 1 | ||||||||||
| nt | x | x | x | x | x | x | - | nt | - | - | - | 2 | ||||||||
| nt | x | x | x | x | x | x | - | B, S | A | 2 | IIIb | - | ||||||||
| nt | x | x | x | x | x | x | - | nt | A, M, K | 2, 3 | - | 2 | ||||||||
| nt | x | x | x | x | x | x | - | S | A, M | 3 | - | 1 | ||||||||
| nt | x | x | x | x | x | x | x | - | nt | A | 2 | IIIb | - | |||||||
| nt | x | x | x | x | x | x | x | - | nt | A | 2, 3 | - | 1 | |||||||
| Dairy | M. morganii | x | x | x | x | x | II | nt | A, O | 1, 2 | - | - | ||||||||
| M. morganii | x | x | x | x | x | x | x | II | nt | A, K | 1, 2 | - | - | |||||||
| C. freundii | x | x | x | x | x | x | x | I | S | A | 1, 2 | Ia | - | |||||||
| C. freundii | x | x | x | x | x | x | I | C, S | - | 1, 3 | Ia | 1 | ||||||||
| C. freundii | x | x | x | x | x | x | x | I | C, S | A | 1, 3 | Ia | 1 | |||||||
| C. braakii | x | x | x | x | x | x | x | - | nt | A | - | Ib | - | |||||||
| C. koseri | x | x | x | x | x | x | x | x | x | - | nt | A, E | 1, 3 | Ib, IIIb | - | |||||
| R. ornithinolytica | x | x | x | x | - | nt | A, E | - | Ia, Ib | - | ||||||||||
| C. koseri | x | x | x | x | - | nt | - | - | IIIc | - | ||||||||||
| E. coli | x | x | x | x | - | nt | A | 2 | Ia, IIa | - | ||||||||||
| E. coli | x | x | x | x | x | x | - | nt | A, E | 1 | IIc | - | ||||||||
| E. coli | x | x | x | x | x | - | nt | - | - | Ia | - | |||||||||
| nt | x | x | x | x | x | x | - | nt | M | 3 | IIIb, IIIc | - | ||||||||
| Slaughterhouse | E. coli | x | x | x | - | nt | A(P), L | - | Ia, Ic | 2 | ||||||||||
| E. coli | x | x | x | x | x | - | nt | O, M | 3 | Ia | - | |||||||||
| E. coli | x | x | x | x | x | x | - | nt | A, M | - | Ic | - | ||||||||
| E. coli | x | x | x | x | x | x | - | - | A, B, O, M | 3 | Ic | - | ||||||||
| E. coli | x | x | x | x | x | x | x | x | x | - | nt | K | 1, 3 | Ia | 1 | |||||
| E. coli | x | x | x | x | x | x | x | x | - | nt | A | 2, 3 | IIb | 1 | ||||||
| E. coli | x | x | x | x | x | x | x | x | I | - | A, M | 2, 3 | Ic, IIb | 1 | ||||||
| S. enterica | x | x | x | x | - | nt | - | 3 | Ia, IIIb | 2 | ||||||||||
| E. vulneris | x | x | x | x | x | - | - | B, M | 1, 2, 3 | Ic, IIb | - | |||||||||
| Enterobacter cloacae | x | x | x | x | x | - | nt | - | - | - | - | |||||||||
| C. freundii | x | x | x | x | x | x | x | - | nt | - | 3 | IIIb | - | |||||||
| nt | x | x | x | - | nt | A(P) | - | - | 1 | |||||||||||
| nt | x | x | x | x | - | B, S | A, M | 3 | - | - | ||||||||||
| nt | x | x | x | x | x | - | B, S | B | 3 | - | - | |||||||||
| nt | x | x | x | x | x | - | nt | A, A(P), M | 3 | IIIb | 2 | |||||||||
| nt | x | x | x | x | x | - | nt | A | 1 | - | - | |||||||||
(a) Isolates origin per enterprise type: Poultry, Pig, Dairy and Slaughterhouse; (b) AB (antibiotics): AMC, amoxicillin/clavulanic acid; FOX, cefoxitin; CTX, cefotaxime; CAZ, ceftazidime; CPO, cefpirome; ATM, aztreonam; IPM, imipenem; MEM, meropenem; CIP, ciprofloxacin; GEN, gentamicin; CHL, chloramphenicol; SXT, trimethoprim/sulfamethoxazol; TET, tetracycline; nt: not tested; -: gene not present.
Table 3.
Absolute and relative frequency of isolates with cat, qnr, oqx, aac(6’)-Ib, qep, tet, sul, dfr and intI genes per enterprise type.
Table 3.
Absolute and relative frequency of isolates with cat, qnr, oqx, aac(6’)-Ib, qep, tet, sul, dfr and intI genes per enterprise type.
| Target Gene/Group | Enterprise Type | Total * | ||||
|---|---|---|---|---|---|---|
| Poultry * | Pig * | Dairy * | Slaughterhouse * | |||
| cat | I | 8 (36.4) | 3 (15.7) | 3 (23.1) | 1 (6.3) | 15 (21.4) |
| II | 2 (15.4) | 2 (2.9) | ||||
| III, IV | 0 (0.0) | |||||
| qnr | A | 0 (0.0) | ||||
| B | 4 (30.8) | 6 (50.0) | 2 (40.0) | 12 (36.4) | ||
| C | 4 (30.8) | 2 (66.7) | 6 (18.1) | |||
| D | 2 (15.4) | 2 (6.1) | ||||
| S | 11 (84.6) | 2 (16.7) | 3 (100.0) | 2 (40.0) | 18 (54.6) | |
| oqx | A | 0 (0.0) | ||||
| B | 2 (16.7) | 2 (6.1) | ||||
| aac(6’)-Ib | 0 (0.0) | |||||
| qep | A | 0 (0.0) | ||||
| tet | A | 19 (86.4) | 15 (78.9) | 9 (69.2) | 7 (43.8) | 50 (71.4) |
| B | 1 (4.6) | 2 (10.5) | 3 (18.8) | 6 (8.6) | ||
| C | 3 (13.6) | 3 (4.3) | ||||
| E | 2 (9.1) | 3 (23.1) | 5 (7.1) | |||
| K | 4 (18.2) | 1 (5.3) | 1 (7.7) | 1 (6.3) | 7 (10.0) | |
| L | 5 (22.7) | 1 (6.3) | 6 (8.6) | |||
| M | 4 (18.2) | 5 (26.3) | 1 (7.7) | 7 (43.8) | 17 (24.3) | |
| O | 1 (4.6) | 1 (7.7) | 2 (12.5) | 4 (5.7) | ||
| A(P) | 2 (9.1) | 3 (18.8) | 5 (7.1) | |||
| D, G, S, Q, X | 0 (0.0) | |||||
| sul | 1 | 15 (68.2) | 5 (26.3) | 7 (53.8) | 3 (18.8) | 30 (42.9) |
| 2 | 9 (40.9) | 11 (57.9) | 4 (30.8) | 3 (18.8) | 27 (38.6) | |
| 3 | 15 (68.2) | 9 (47.4) | 4 (30.8) | 11 (68.8) | 40 (57.1) | |
| dfr | Ia | 17 (77.2) | 9 (47.4) | 6 (46.2) | 4 (25.0) | 36 (51.4) |
| Ib | 4 (18.2) | 3 (23.1) | 7 (10.0) | |||
| Ic | 5 (22.7) | 5 (26.3) | 5 (31.3) | 15 (21.4) | ||
| IIa | 1 (4.6) | 2 (10.5) | 1 (7.7) | 4 (5.7) | ||
| IIb | 6 (27.3) | 3 (18.8) | 9 (12.9) | |||
| IIc | 1 (7.7) | 1 (1.4) | ||||
| IIIa | 3 (13.6) | 3 (4.3) | ||||
| IIIb | 4 (21.1) | 2 (15.4) | 3 (18.8) | 9 (12.9) | ||
| IIIc | 2 (9.1) | 2 (15.4) | 4 (5.7) | |||
| IVa, IVb, IVc | 0 (0.0) | |||||
| Va, Vb, Vc, Vd | 0 (0.0) | |||||
| intI | 1 | 5 (22.7) | 5 (26.3) | 2 (15.4) | 4 (25.0) | 16 (22.9) |
| 2 | 6 (31.6) | 3 (18.8) | 9 (12.9) | |||
| 3 | 0 (0.0) | |||||
* n (%), absolute and (relative) frequency of isolates carrying the gene. The blank space means a negative result, i.e., 0 (0%).
Table 4.
Resistance phenotype of isolates from four livestock enterprises to non-β-lactamic AB and corresponding presence of integrons class.
Table 4.
Resistance phenotype of isolates from four livestock enterprises to non-β-lactamic AB and corresponding presence of integrons class.
| AB | Resistant Isolates (N) | Resistant Isolates with Integrons, N (%) | Resistant Isolates without Integrons, N (%) | |
|---|---|---|---|---|
| intI1 | intI2 | |||
| CHL | 58 | 14 (24.1%) | 8 (13.8%) | 36 (62.1%) |
| CIP | 29 | 10 (34.5%) | 1 (3.4%) | 18 (62.1%) |
| GEN | 7 | 1 (14.3%) | 0 (0.0%) | 6 (85.7%) |
| SXT | 56 | 15 (26.8%) | 8 (14.3%) | 33 (58.9%) |
| TET | 63 | 13 (20.6%) | 8 (12.7%) | 42 (66.7%) |
N (%), absolute and (relative) frequency of isolates carrying the gene.
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Amador, P.; Fernandes, R.; Prudêncio, C.; Duarte, I. Prevalence of Antibiotic Resistance Genes in Multidrug-Resistant Enterobacteriaceae on Portuguese Livestock Manure. Antibiotics 2019, 8, 23. https://doi.org/10.3390/antibiotics8010023
AMA Style
Amador P, Fernandes R, Prudêncio C, Duarte I. Prevalence of Antibiotic Resistance Genes in Multidrug-Resistant Enterobacteriaceae on Portuguese Livestock Manure. Antibiotics. 2019; 8(1):23. https://doi.org/10.3390/antibiotics8010023
Chicago/Turabian StyleAmador, Paula, Ruben Fernandes, Cristina Prudêncio, and Isabel Duarte. 2019. "Prevalence of Antibiotic Resistance Genes in Multidrug-Resistant Enterobacteriaceae on Portuguese Livestock Manure" Antibiotics 8, no. 1: 23. https://doi.org/10.3390/antibiotics8010023
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.