
The Phagosome–Lysosome Fusion Is the Target of a Purified Quillaja saponin Extract (PQSE) in Reducing Infection of Fish Macrophages by the Bacterial Pathogen Piscirickettsia salmonis

Abstract
:1. Introduction
2. Materials and Methods
2.1. Bacterial and Cell Culture
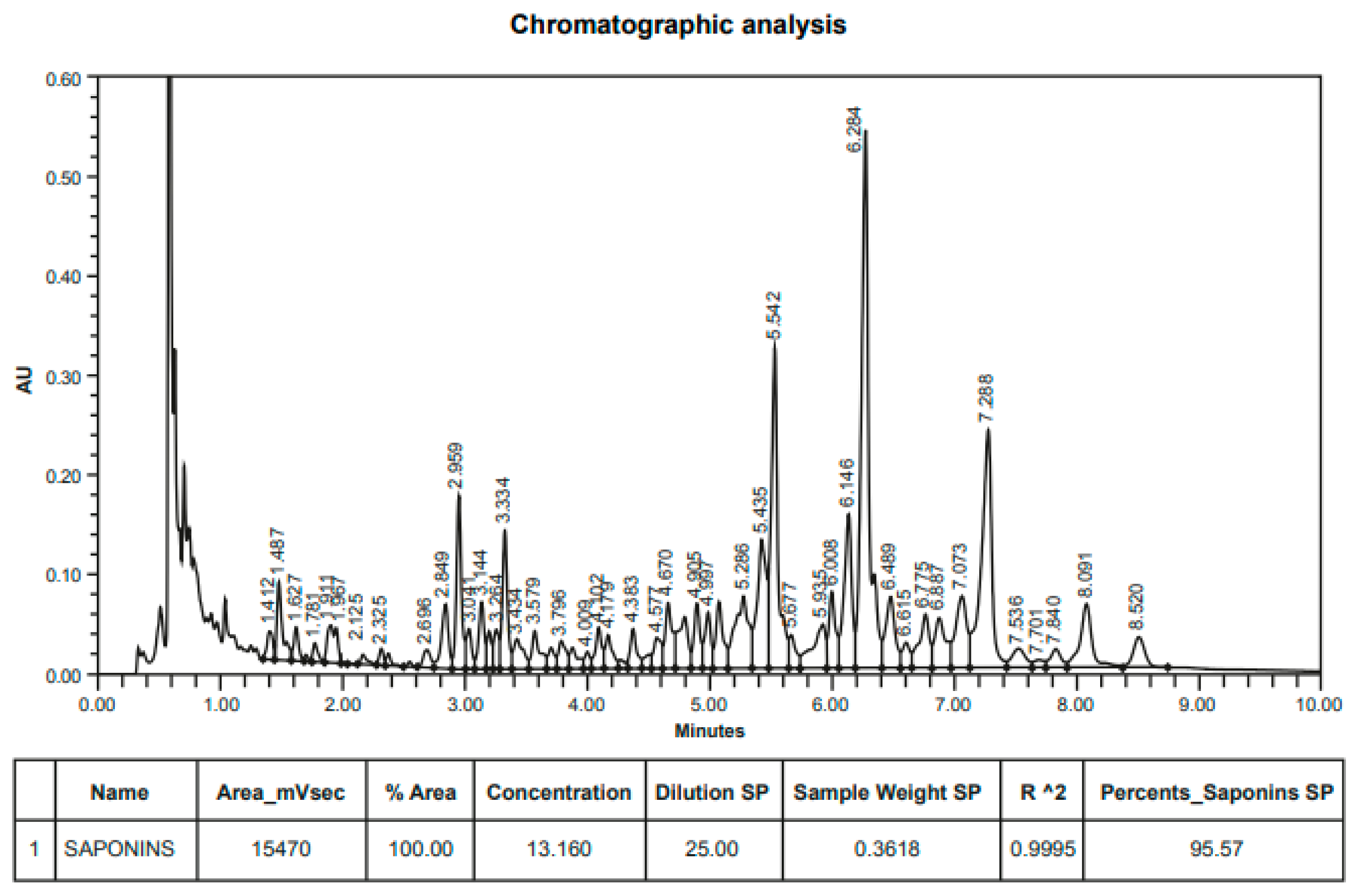
2.2. Direct Quillaja Extract Antimicrobial Analysis
2.3. Cytotoxicity
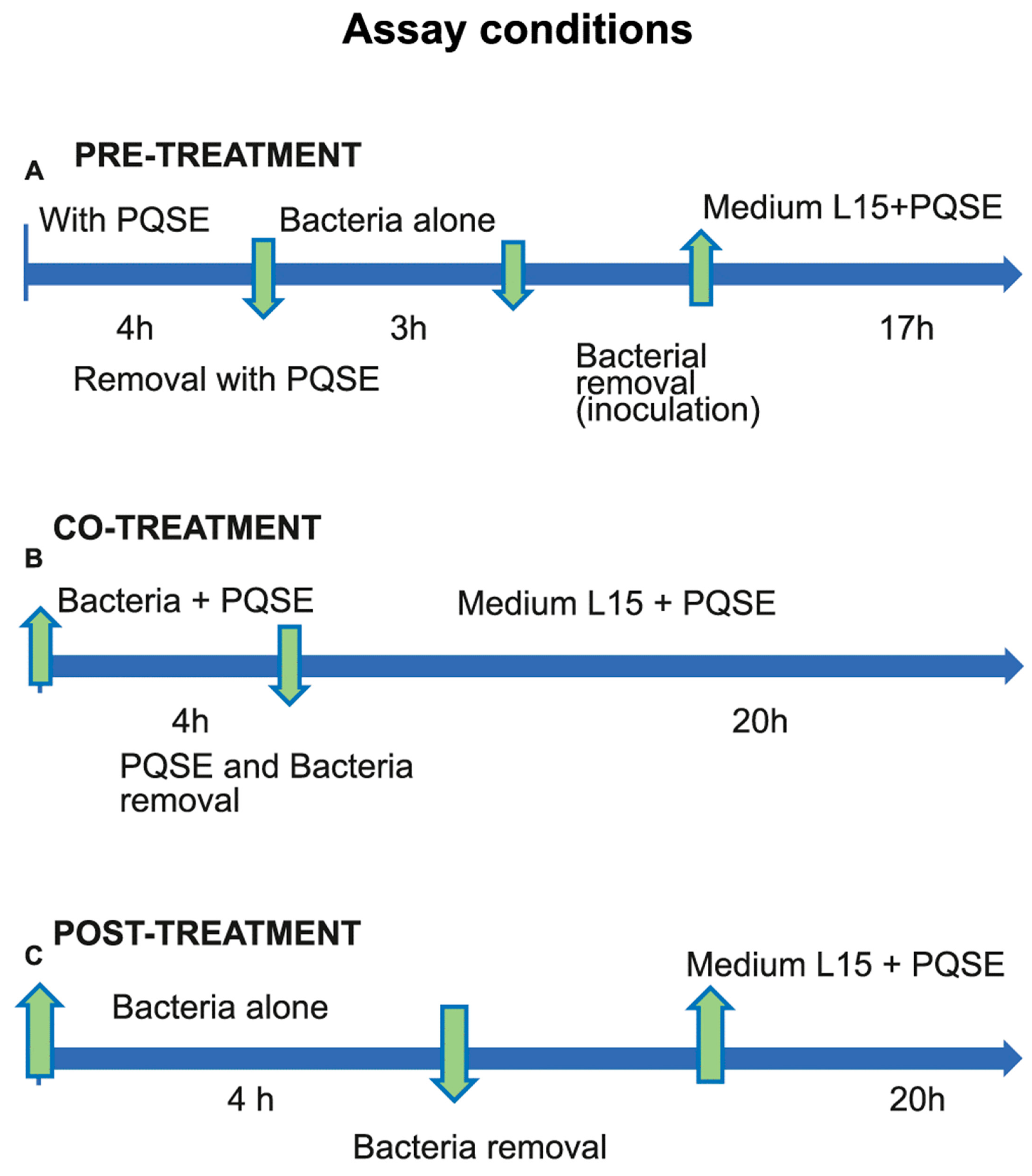
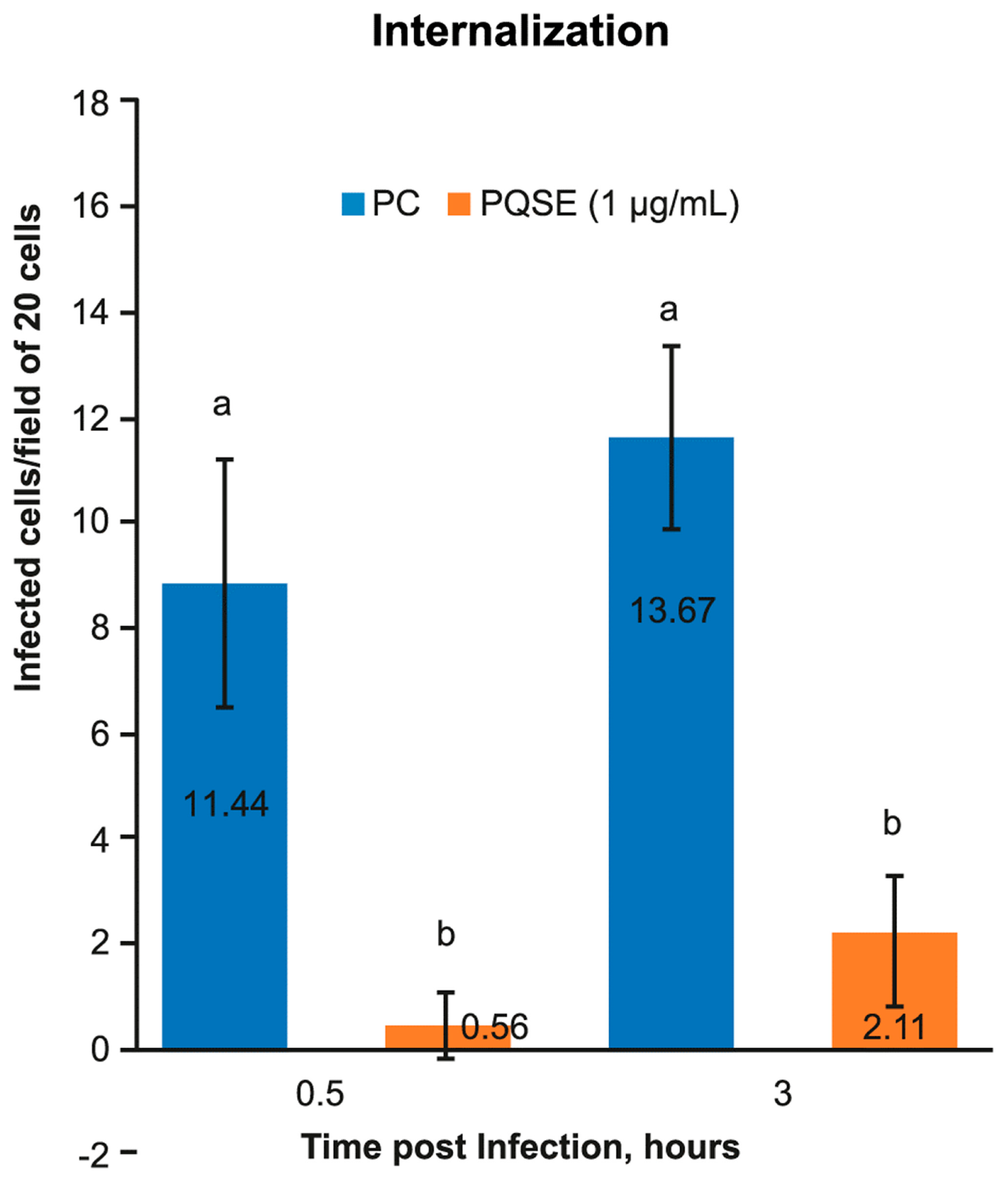
2.4. Internalization of P. salmonis in SHK-1
2.5. Proliferation of P. salmonis in SHK-1 Cells Pre-Incubated with PQSE
2.6. Quantification of P. salmonis DNA: Bacterial Load
2.7. Transcription of the dotB, chaPs, IL-10, and IL-12 Genes
2.8. cDNA Synthesis to Quantify the Transcription of dotB, chaPs, IL-10, and IL-12 Genes
2.9. Expression of IL-12 and IL-10 Genes by qRT-PCR in SHK-1 Cells Pre-Incubated with PQSE
2.10. Evaluation of Phagosome–Lysosome (P-L) Fusion
2.11. Statistical Analysis
3. Results
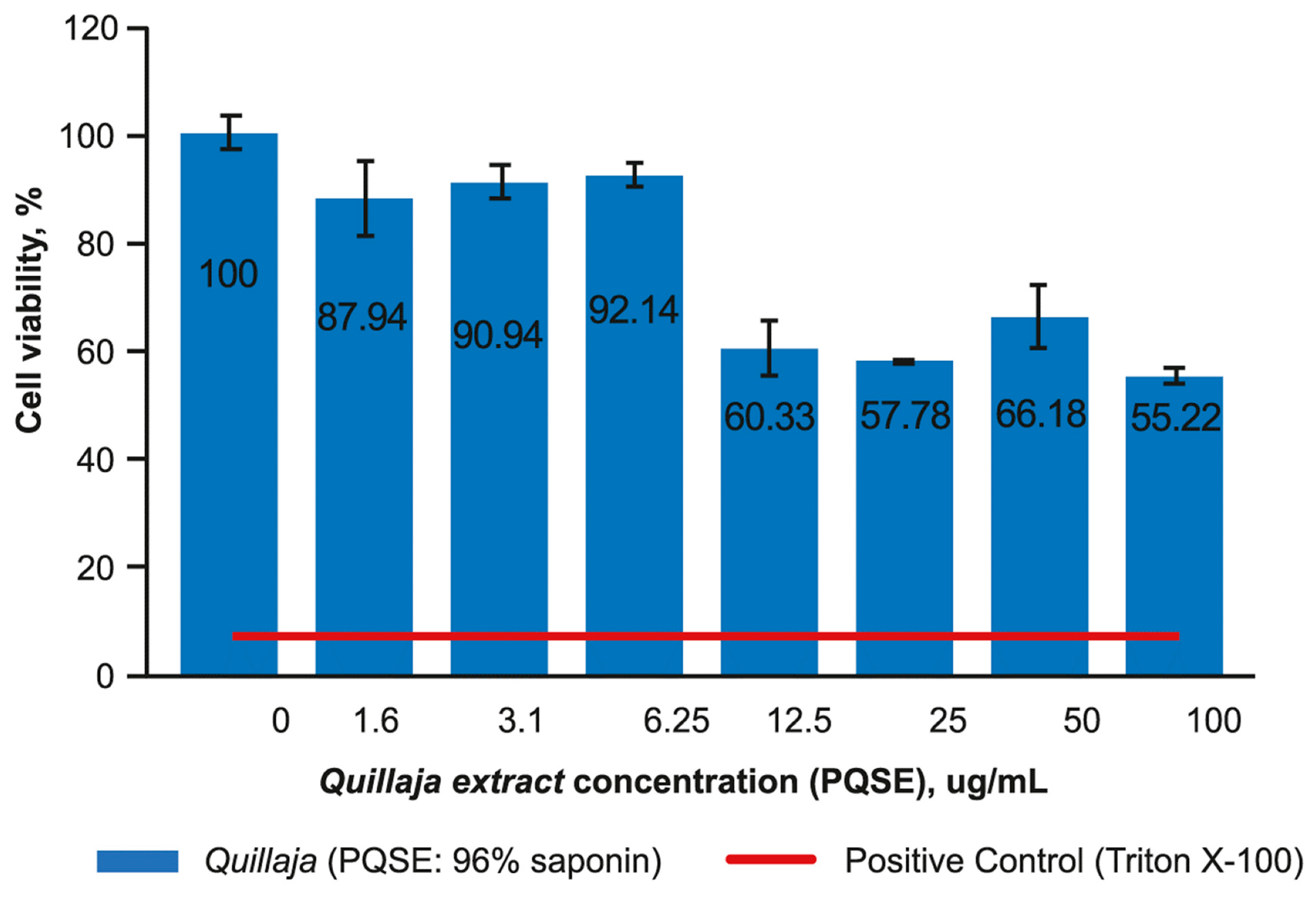
3.1. SHK-1 Cell Lines Are Sensitive to the Quillaja Extract Levels
3.2. The Purified Quillaja Saponin Extracts (PQSEs) Are Not Toxic to P. salmonis at Physiological Doses for the Cell
3.3. Quillaja Saponaria Extracts Reduce the Internalization of P. salmonis in SHK-1
3.4. SHK-1 Cells Pre-Incubated with PQSE Reduce the Intracellular Proliferation (72 hpi) of P. salmonis
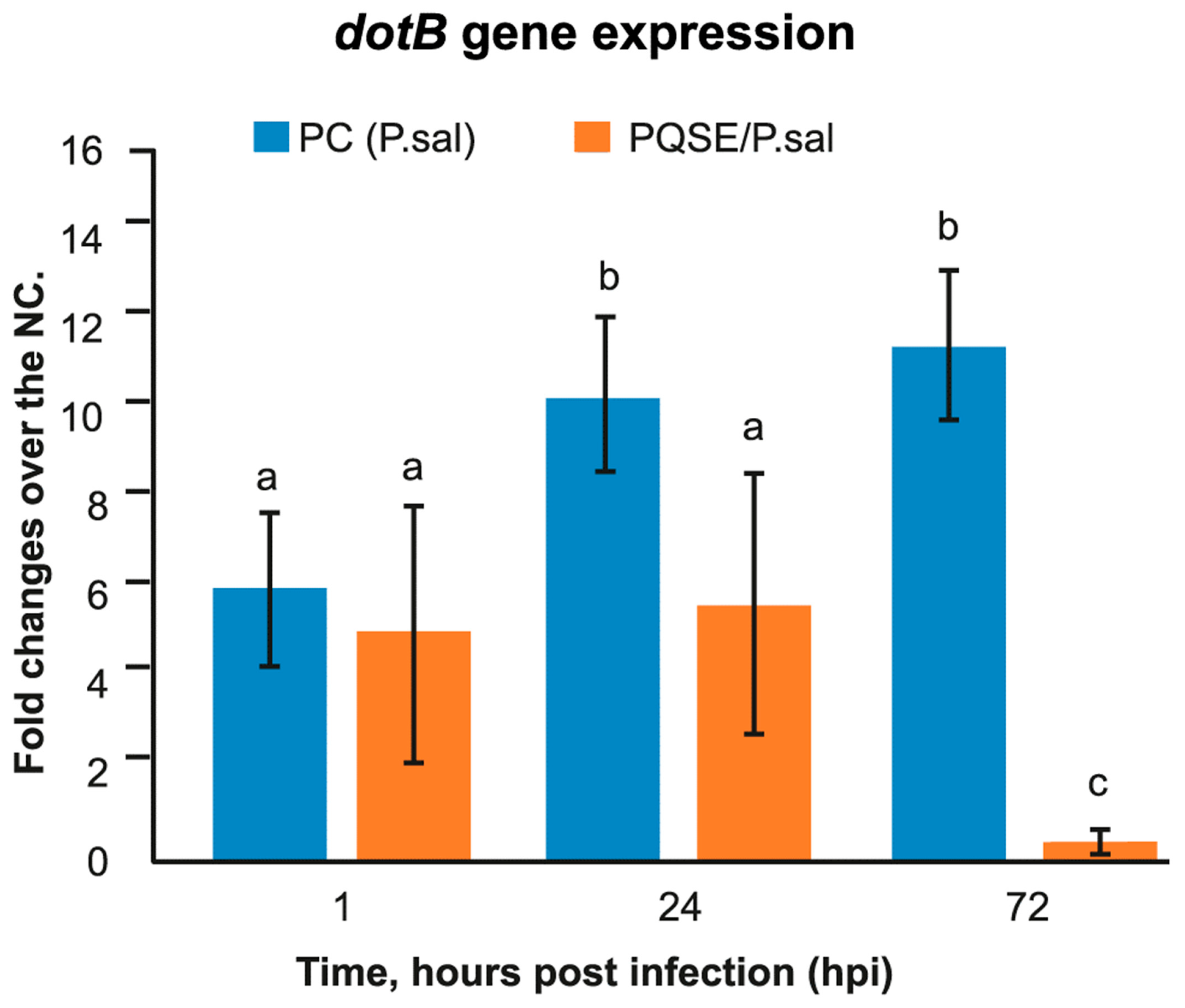
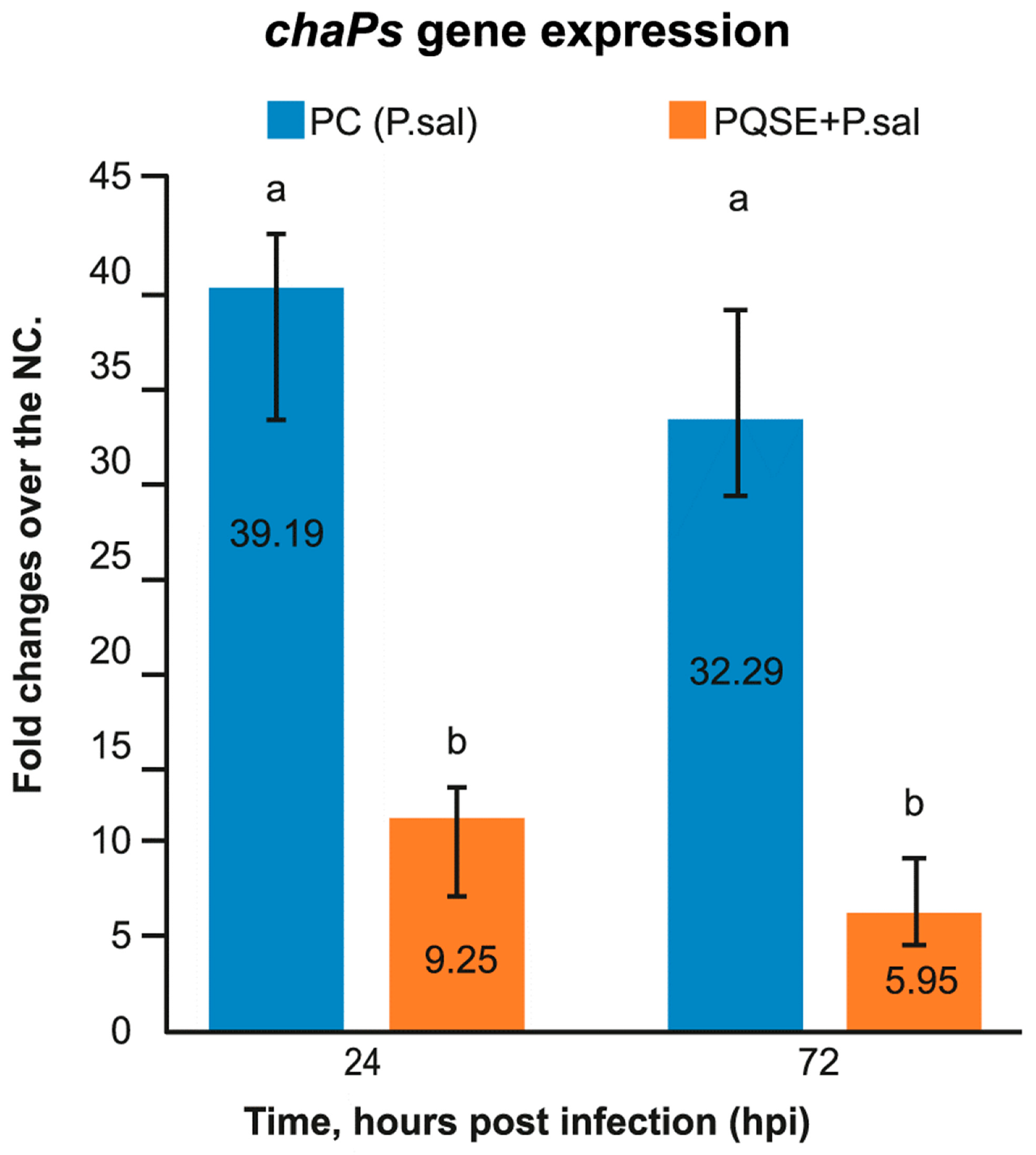
3.5. SHK-1 Cells Pre-Treated with PQSE Reduce the Gene Expression of dotB (T4-BSS) and chaPs of P. salmonis
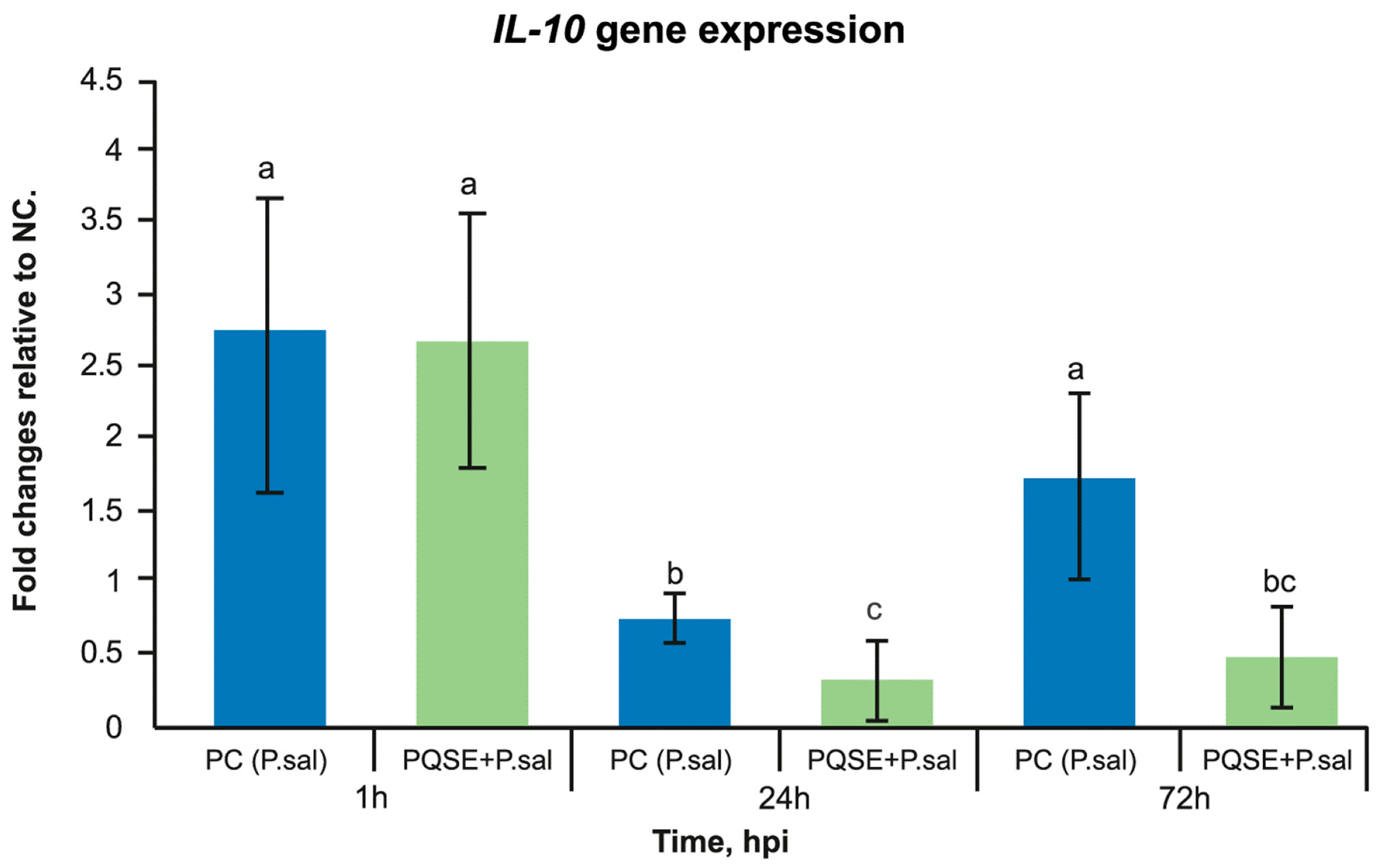
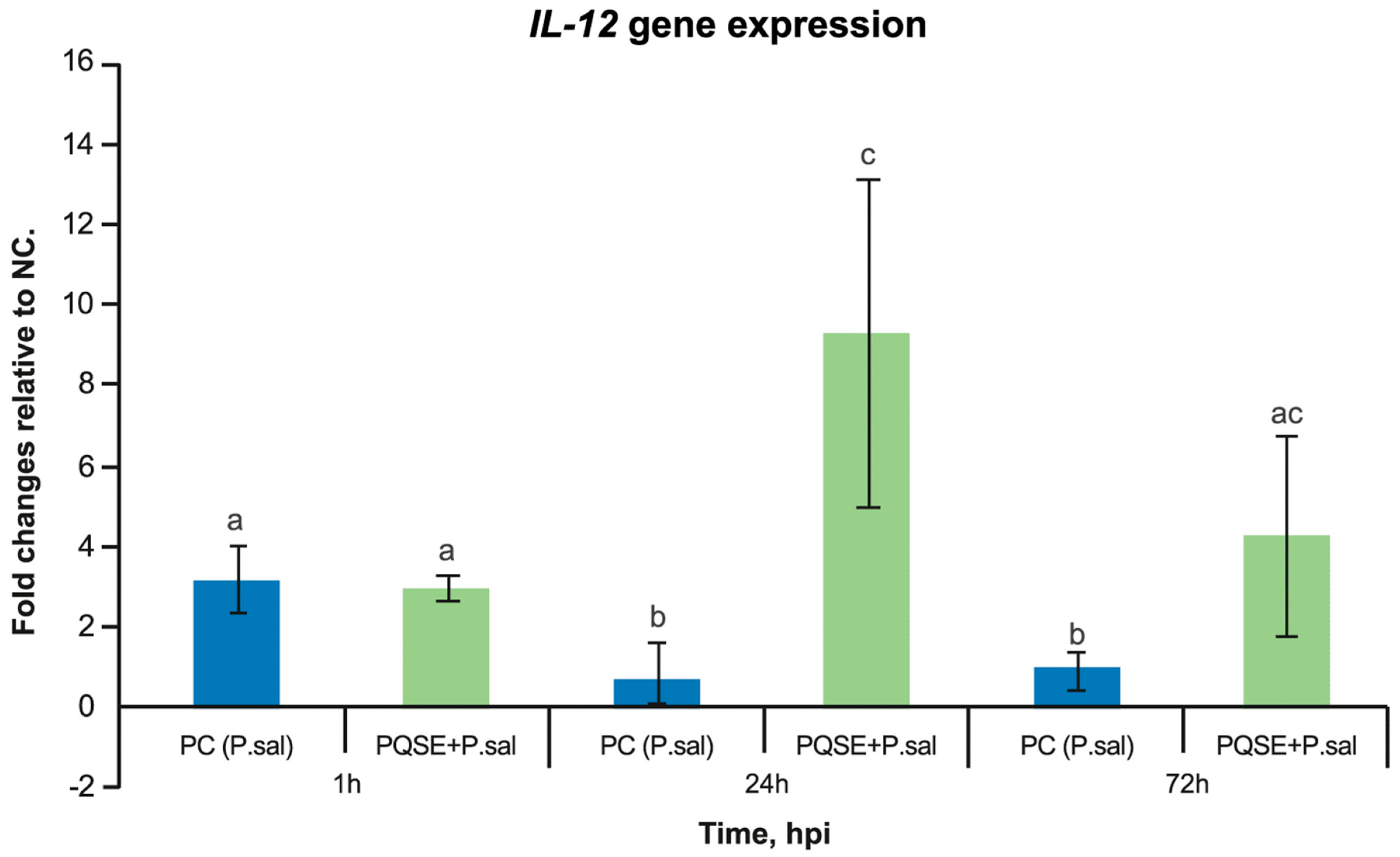
3.6. SHK-1 Cells Pre-Incubated with PQSE Reduce the Gene Expression of Interleukin 10 (IL-10) and Favor the Expression of IL-12 during the Acute Phase of P. salmonis Infection (<72 hpi)
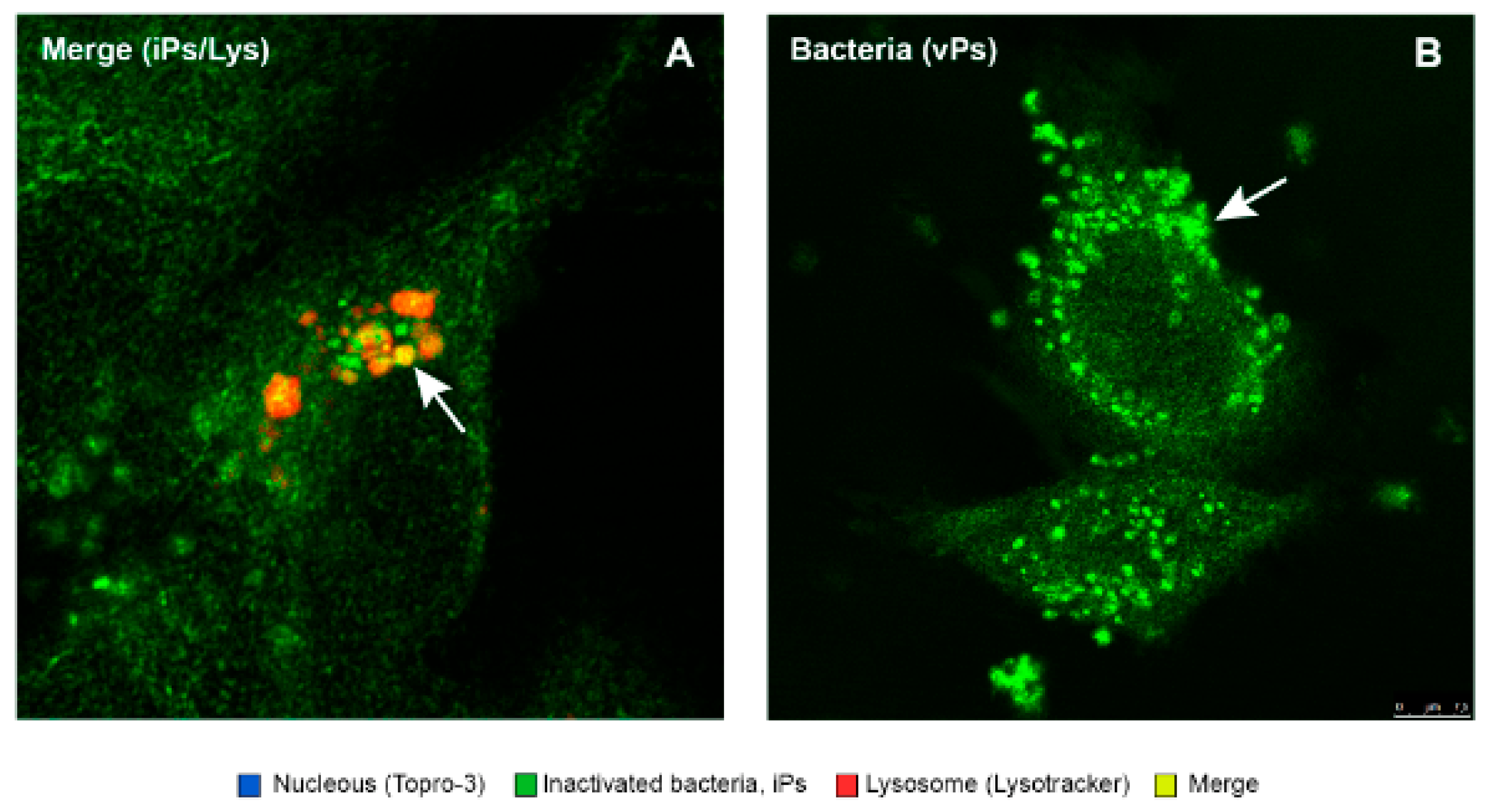
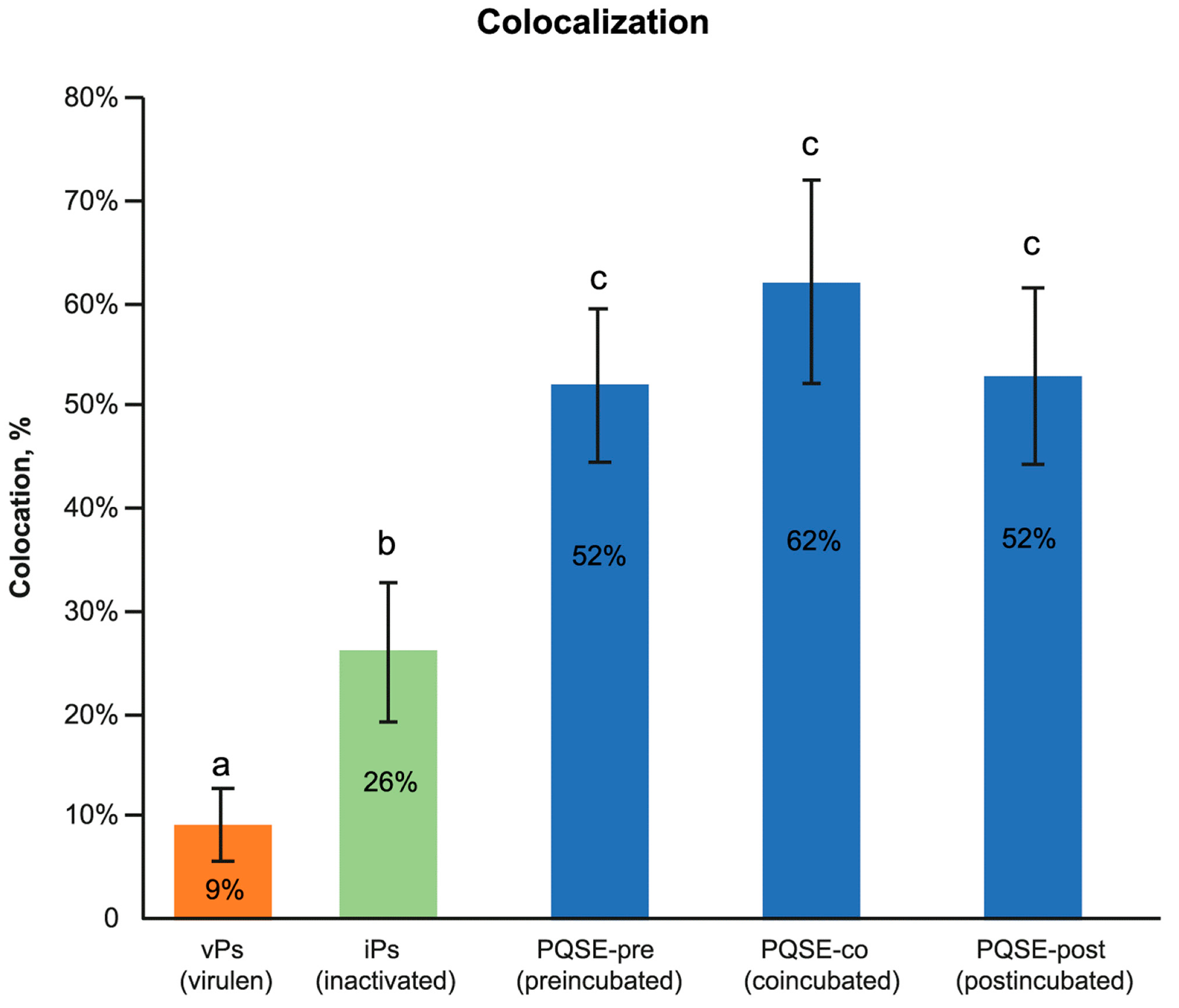
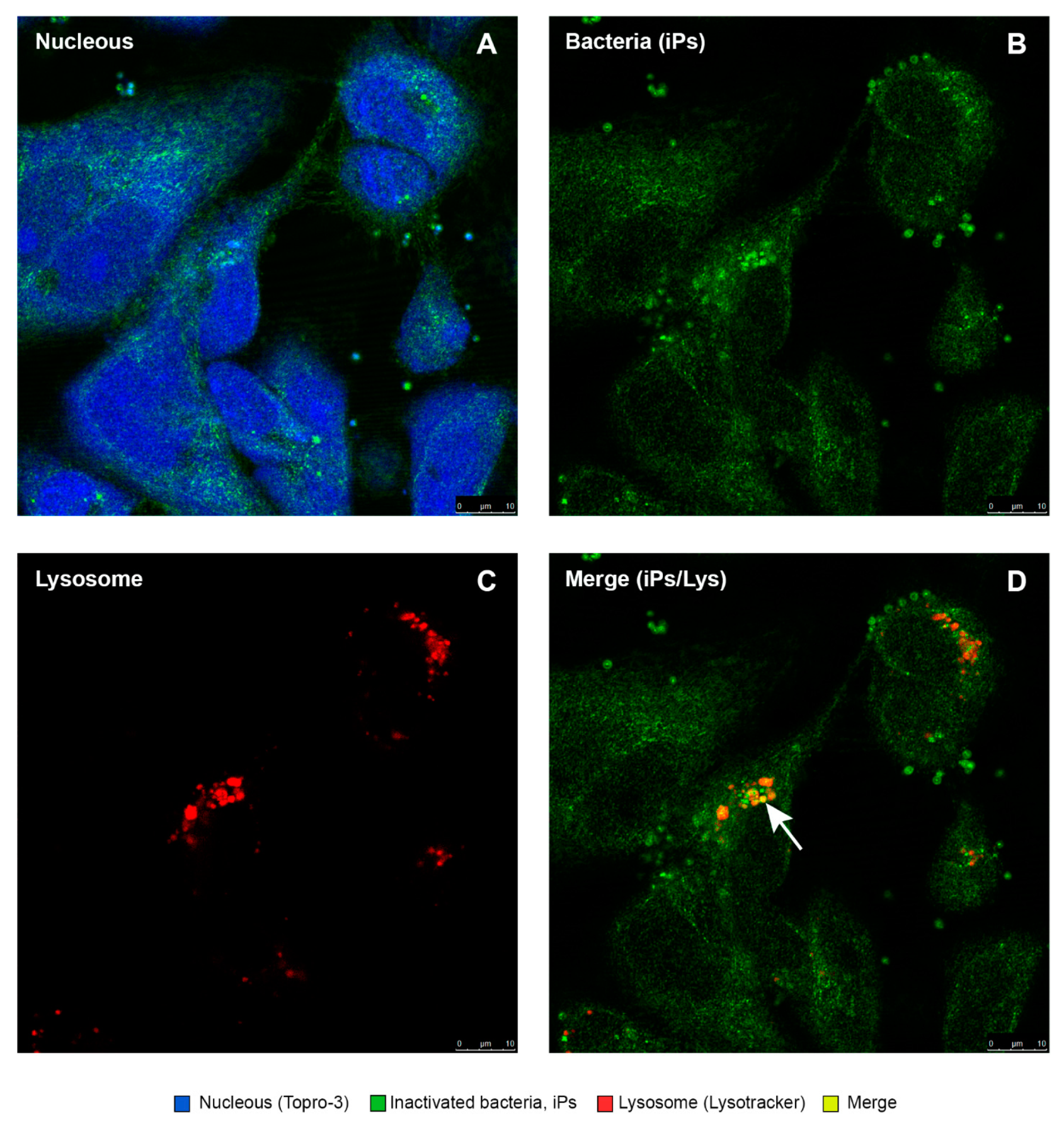
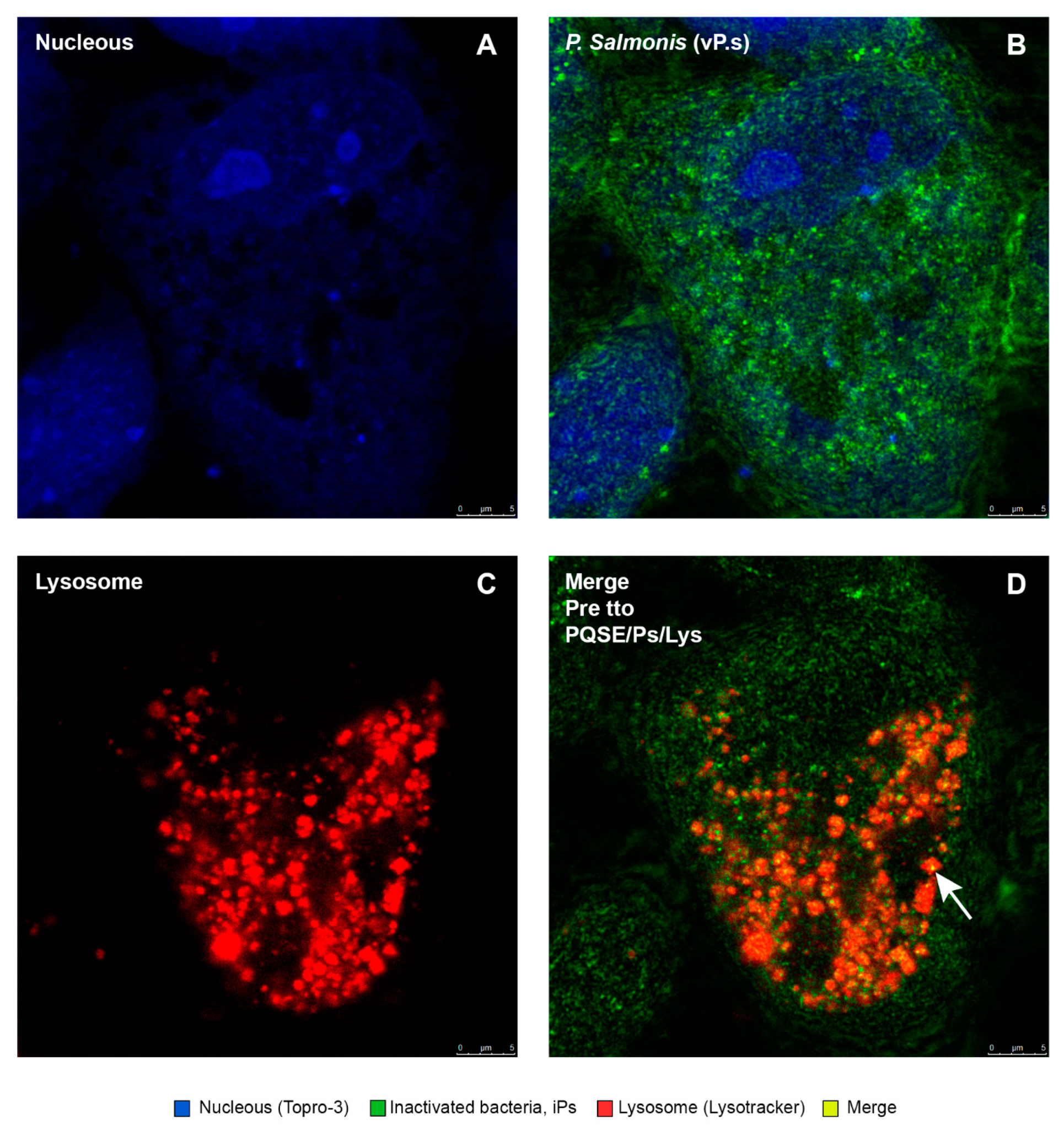
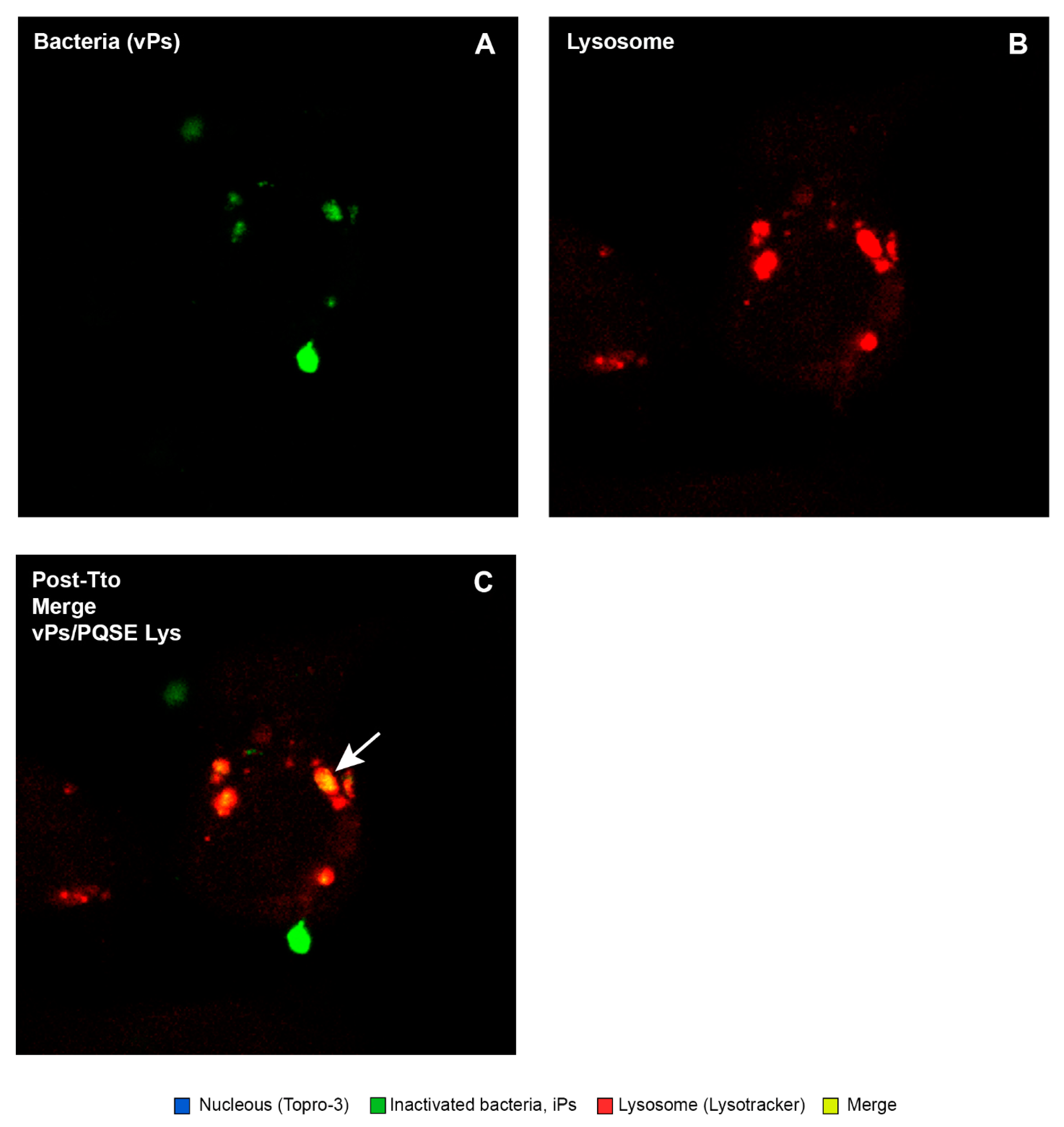
3.7. Purified Quillaja Saponaria Extracts (PQSEs) Favor the Formation of Phago-Lysosomes in SHK-1 Cells
4. Discussion
4.1. SHK-1 Cell Lines Were Sensitive to Quillaja saponaria Extracts
4.2. PQSE Does Not Have a Direct Antimicrobial Effect on P. salmonis at Physiological Doses to the Host Cell
4.3. PQSEs Reduce the Internalization and Intracellular Proliferation of P. salmonis
4.4. PQSE Disrupts Cell Membranes, a Mode of Action That Could Help Reduce the Internalization of P. salmonis in SHK-1 Cells
4.5. SHK-1 Cells Pre-Incubated with PQSE Reduce the Expression of dotB (T4-BSS) and chaPs Protein of P. salmonis
4.6. SHK-1 Cells Pre-Incubated con PQSE Modulate the Expression of Anti-Inflammatory IL-10 and Proinflammatory IL-12 Gene Expression, Favoring a Proinflammatory Environment in the Early Phase of P. salmonis Infection (<72 h)
5. Conclusions
Ending Considerations
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Figueroa, J.; Cárcamo, J.; Yañez, A.; Olavarria, V.; Ruiz, P.; Manríquez, R.; Muñoz, C.; Romero, A.; Avendaño-Herrera, R. Addressing viral and bacterial threats to salmon farming in Chile: Historical contexts and perspectives for management and control. Rev. Aquac. 2019, 11, 299–324. [Google Scholar] [CrossRef]
- Servicio Nacional de Pesca y Acuicultura (Sernapesca). Informe Sanitario de Salmonicultura en Centros Marinos; Primer Semestre: Valparaíso, Chile, 2018.
- Tandberg, J.I.; Lagos, L.X.; Langlete, P.; Berger, E.; Rishovd, A.-L.; Roos, N.; Varkey, D.; Paulsen, I.; Winther-Larsen, H.C. Comparative analysis of membrane vesicles from three Piscirickettsia salmonis isolates reveals differences in vesicle characteristics. PloS ONE 2016, 11, e0165099. [Google Scholar] [CrossRef]
- Miranda, C.D.; Godoy, F.A.; Lee, M.R. Current status of the use of antibiotics and the antimicrobial resistance in the chilean salmon farms. Front. Microbiol. 2018, 9, 1284. [Google Scholar] [CrossRef] [PubMed]
- Cabello, F.C.; Godfrey, H.; Buschmann, A.H.; Dölz, H.J. Aquaculture as yet another environmental gateway to the development and globalisation of antimicrobial resistance. Lancet Infect. Dis. 2016, 16, e127–e133. [Google Scholar] [CrossRef]
- Bohle, H.; Saavedra, J.; Mancilla, M.; Diagnostic, A.D.L. Un plásmido de Psalmonis codifica para resistencia a múltiples antimicrobianos. Report Tech. 2017. [Google Scholar] [CrossRef]
- Henríquez, P.; Bohle, H.; Bustamante, F.; Bustos, P.; Mancilla, M. Polymorphism in gyrA is associated to quinolones resistance in Chilean Piscirickettsia salmonis field isolates. J. Fish Dis. 2015, 38, 415–418. [Google Scholar] [CrossRef]
- Cabello, F.C. Heavy use of prophylactic antibiotics in aquaculture: A growing problem for human and animal health and for the environment. Environ. Microbiol. 2006, 8, 1137–1144. [Google Scholar] [CrossRef] [PubMed]
- Rozas, M.; Enríquez, R. Piscirickettsiosis and Piscirickettsia salmonis in fish: A review. J. Fish Dis. 2014, 37, 163–188. [Google Scholar] [CrossRef]
- Pulgar, R.; Hödar, C.; Travisany, D.; Zuñiga, A.; Domínguez, C.; Maass, A.; Gonzalez, M.J.M.; Cambiazo, V. Transcriptional response of Atlantic salmon families to Piscirickettsia salmonis infection highlights the relevance of the iron-deprivation defence system. BMC Genom. 2015, 16, 495. [Google Scholar] [CrossRef] [Green Version]
- Sandoval, A.; Tapia, E.; Henríquez, P. Estudio de Seguridad y Eficacia Antibacteriana y Antiviral de Extractos Naturales; Informe Final ID-P03-2014; Chile, 2014; Available online: https://docs.bvsalud.org/biblioref/2019/05/997943/estudio-in-vitro-de-la-actividad-antibacteriana-y-antifungica-d_gURYPhH.pdf (accessed on 23 March 2021).
- Maisey, K.; Montero, R.; Corripio-Miyar, Y.; Toro-Ascuy, D.; Valenzuela, B.; Reyes-Cerpa, S.; Sandino, A.M.; Zou, J.; Wang, T.; Secombes, C.J.; et al. Isolation and characterization of salmonid CD4+ T Cells. J. Immunol. 2016, 196, 4150–4163. [Google Scholar] [CrossRef] [Green Version]
- Tobar, I.; Arancibia, S.; Torres, C.; Vera, V.; Soto, P.; Carrasco, C.; Alvarado, M.; Neira, E.; Arcos, S.; Tobar, J.A. Successive Oral Immunizations Against Piscirickettsia Salmonis and Infectious Salmon Anemia Virus are Required to Maintain a Long-Term Protection in Farmed Salmonids. Front. Immunol. 2015, 6, 244. [Google Scholar] [CrossRef] [Green Version]
- Harper, D.R. Criteria for selecting suitable infectious diseases for phage therapy. Viruses 2018, 10, 177. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Martínez, D.; Oyarzún, R.; Pontigo, J.; Romero, A.; Yáñez, A.; Vargas-Chacoff1, L. Nutritional Immunity Triggers the Modulation of Iron Metabolism Genes in the Sub-Antarctic Notothenioid Eleginops maclovinus in Response to Piscirickettsia salmonis. Front. Immunol. 2017, 8, 1153. Available online: https://www.frontiersin.org/article/10.3389/fimmu.2017.01153 (accessed on 23 March 2021). [CrossRef] [PubMed] [Green Version]
- Assefa, A.; Abunna, F. Maintenance of Fish Health in Aquaculture: Review of Epidemiological Approaches for Prevention and Control of Infectious Disease of Fish. Veter. Med. Int. 2018, 2018, 1–10. [Google Scholar] [CrossRef] [PubMed]
- Yã¡ã-Ez, J.M.; Newman, S.; Houston, R.D.; Yáñez, J.M. Genomics in aquaculture to better understand species biology and accelerate genetic progress. Front. Genet. 2015, 6, 1–3. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barría, A.; Christensen, K.A.; Yoshida, G.M.; Correa, K.; Jedlicki, A.; Lhorente, J.P.; Davidson, W.S.; Yáñez, J.M. Genomic Predictions and Genome-Wide Association Study of Resistance Against Piscirickettsia salmonis in Coho Salmon (Oncorhynchus kisutch) Using ddRAD Sequencing. G3 Genes Genomes Genet. 2018, 8, 1183–1194. [Google Scholar] [CrossRef] [Green Version]
- Pérez-Stuardo, D.; Morales-Reyes, J.; Tapia, S.; Ahumada, D.E.; Espinoza, A.; Soto-Herrera, V.; Brianson, B.; Ibaceta, V.; Sandino, A.M.; Spencer, E.; et al. Non-lysosomal activation in macrophages of Atlantic salmon (salmo salar) after infection with Piscirickettsia salmonis. Front. Immunol. 2019, 10, 1–11. [Google Scholar] [CrossRef] [PubMed]
- Rozas-Serri, M.; Peña, A.; Maldonado, L. Transcriptomic profiles of post-smolt Atlantic salmon challenged with Piscirickettsia salmonis reveal a strategy to evade the adaptive immune response and modify cell-autonomous immunity. Dev. Comp. Immunol. 2018, 81, 348–362. [Google Scholar] [CrossRef] [PubMed]
- Ramírez, R.; Gomez, F.; Marshall, S.H. The infection process of Piscirickettsia salmonis in fish macrophages is dependent upon interaction with host-cell clathrin and actin. Fems Microbiol. Lett. 2015, 362, 1–8. [Google Scholar] [CrossRef]
- McCarthy, Ú.M.; Bron, J.; Brown, L.; Pourahmad, F.; Bricknell, I.R.; Thompson, K.D.; Adams, A.; Ellis, A.E. Survival and replication of Piscirickettsia salmonis in rainbow trout head kidney macrophages. Fish. Shellfish. Immunol. 2008, 25, 477–484. [Google Scholar] [CrossRef]
- Rojas, V.; Galanti, N.; Bols, N.C.; Marshall, S. Productive infection of Piscirickettsia salmonisin macrophages and monocyte-like cells from rainbow trout, a possible survival strategy. J. Cell. Biochem. 2009, 108, 631–637. [Google Scholar] [CrossRef] [PubMed]
- Barry, A.O.; Mege, J.-L.; Ghigo, E. Hijacked phagosomes and leukocyte activation: An intimate relationship. J. Leukoc. Biol. 2010, 89, 373–382. [Google Scholar] [CrossRef] [PubMed]
- Flannagan, R.; Cosío, G.; Grinstein, S. Antimicrobial mechanisms of phagocytes and bacterial evasion strategies. Nat. Rev. Genet. 2009, 7, 355–366. [Google Scholar] [CrossRef] [PubMed]
- Rosenberger, C.M.; Finlay, B.B. Phagocyte sabotage: Disruption of macrophage signalling by bacterial pathogens. Nat. Rev. Mol. Cell Biol. 2003, 4, 385–396. [Google Scholar] [CrossRef]
- Thi, E.P.; Lambertz, U.; Reiner, N.E. Sleeping with the Enemy: How Intracellular Pathogens Cope with a Macrophage Lifestyle. PLoS Pathog. 2012, 8, e1002551. [Google Scholar] [CrossRef] [Green Version]
- Alix, E.; Mukherjee, S.; Roy, C.R. Subversion of membrane transport pathways by vacuolar pathogens. J. Cell Biol. 2011, 195, 943–952. [Google Scholar] [CrossRef]
- Mitchell, G.; Chen, C.; Portnoy, D.A. Strategies Used by Bacteria to Grow in Macrophages. Myeloid Cells Health Dis. 2016, 4, 701–725. [Google Scholar] [CrossRef] [Green Version]
- Sgro, G.G.; Oka, G.U.; Souza, D.P.; Cenens, W.; Bayer-Santos, E.; Matsuyama, B.Y.; Bueno, N.F.; Dos Santos, T.R.; Alvarez-Martinez, C.E.; Salinas, R.K.; et al. Bacteria-Killing Type IV Secretion Systems. Front. Microbiol. 2019, 10, 1078. [Google Scholar] [CrossRef]
- Park, D.; Chetrit, D.; Hu, B.; Roy, C.R.; Liu, J. Analysis of Dot/Icm Type IVB Secretion System Subassemblies by Cryoelectron Tomography Reveals Conformational Changes Induced by DotB Binding. mBio 2020, 11, 03328-19. [Google Scholar] [CrossRef] [Green Version]
- Meir, A.; Chetrit, D.; Liu, L.; Roy, C.R.; Waksman, G. Legionella DotM structure reveals a role in effector recruiting to the Type 4B secretion system. Nat. Commun. 2018, 9, 1–12. [Google Scholar] [CrossRef] [Green Version]
- Schroeder, G.N.; Petty, N.K.; Mousnier, A.; Harding, C.R.; Vogrin, A.J.; Wee, B.; Fry, N.; Harrison, T.G.; Newton, H.J.; Thomson, N.; et al. Legionella pneumophila Strain 130b Possesses a Unique Combination of Type IV Secretion Systems and Novel Dot/Icm Secretion System Effector Proteins. J. Bacteriol. 2010, 192, 6001–6016. [Google Scholar] [CrossRef] [Green Version]
- Newton, H.J.; Ang, D.K.Y.; van Driel, I.R.; Hartland, E.L. Molecular Pathogenesis of Infections Caused by Legionella pneumophila. Clin. Microbiol. Rev. 2010, 23, 274–298. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sánchez, P.; Oliver, C.; Hernández, M.; Rauch, M.C.; Valenzuela, K.; Rafael, A.; Avendaño-herrera, R.; Yáñez, A.J. In vitro genomic and proteomic evidence of a type IV pili-like structure in the fish pathogen Piscirickettsia salmonis Huella Profundice en los temas de investigación de’ In vitro genomic and proteomic. FEMS Microbiol. Lett. 2018, 365, fny169. [Google Scholar] [CrossRef]
- Tseng, T.-T.; Tyler, B.M.; Setubal, J.C. Protein secretion systems in bacterial-host associations, and their description in the Gene Ontology. BMC Microbiol. 2009, 9, S2. [Google Scholar] [CrossRef] [Green Version]
- Voth, D.E.; Heinzen, R.A. Lounging in a lysosome: The intracellular lifestyle of Coxiella burnetii. Cell. Microbiol. 2007, 9, 829–840. [Google Scholar] [CrossRef] [PubMed]
- Labra, Á.; Arredondo-Zelada, O.; Flores-Herrera, P.; Marshall, S.H.; Gómez, F.A. In sílico identification and characterization of putative Dot/Icm secreted virulence effectors in the fish pathogen Piscirickettsia salmonis. Microb. Pathog. 2016, 92, 11–18. [Google Scholar] [CrossRef]
- Gomez, F.A.; Tobar, J.A.; Henríquez, V.; Sola, M.; Altamirano, C.; Marshall, S.H. Evidence of the presence of a functional Dot/Icm Type IV-B secretion system in the fish bacterial pathogen Piscirickettsia salmonis. PLoS ONE 2013, 8, e54934. [Google Scholar] [CrossRef]
- Cortés, M.; Sánchez, P.; Ruiz, P.; Haro, R.; Sáez, J.; Sánchez, F.; Hernández, M.; Oliver, C.; Yáñez, A.J. In vitro expression of Sec-dependent pathway and type 4B secretion system in Piscirickettsia salmonis. Microb. Pathog. 2017, 110, 586–593. [Google Scholar] [CrossRef] [PubMed]
- Alvarez, C.; Gomez, F.; Mercado, L.; Ramírez, R.; Marshall, S.H. Piscirickettsia salmonis imbalances the innate immune response to succeed in a productive infection in a salmonid cell line model. PLoS ONE 2016, 11, e0163943. [Google Scholar] [CrossRef] [Green Version]
- Nagamatsu, K.; Kuwae, A.; Konaka, T.; Nagai, S.; Yoshida, S.; Eguchi, M.; Watanabe, M.; Mimuro, H.; Koyasu, S.; Abe, A. Bordetella evades the host immune system by inducing IL-10 through a type III effector, BopN. J. Exp. Med. 2009, 206, 3073–3088. [Google Scholar] [CrossRef] [Green Version]
- Wang, T.; Husain, M. The expanding repertoire of the IL-12 cytokine family in teleost fish: Identification of three paralogues each of the p35 and p40 genes in salmonids, and comparative analysis of their expression and modulation in Atlantic salmon Salmo salar. Dev. Comp. Immunol. 2014, 46, 194–207. [Google Scholar] [CrossRef] [PubMed]
- Chanteux, H.; Guisset, A.C.; Pilette, C.; Sibille, Y. LPS induces IL-10 production by human alveolar macrophages via MAPKinases- and Sp1-dependent mechanisms. Respir. Res. 2007, 8, 71. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Yoshizawa, S.; Tateda, K.; Matsumoto, T.; Gondaira, F.; Miyazaki, S.; Standiford, T.J.; Yamaguchi, K. Legionella pneumophila evades Gamma interferon-mediated growth suppression through Interleukin-10 induction in bone marrow-derived macrophages. Infect. Immun. 2005, 73, 2709–2717. [Google Scholar] [CrossRef] [Green Version]
- Hunt, D.; Wilson, J.E.; Weih, K.A.; Ishido, S.; Harton, J.A.; Roche, P.; Drake, J.R. Francisella tularensis Elicits IL-10 via a PGE2-Inducible Factor, to Drive Macrophage MARCH1 Expression and Class II Down-Regulation. PLoS ONE 2012, 7, e37330. [Google Scholar] [CrossRef]
- O’Leary, S.; O’Sullivan, M.P.; Keane, J. IL-10 Blocks Phagosome Maturation inMycobacterium tuberculosis–Infected Human Macrophages. Am. J. Respir. Cell Mol. Biol. 2011, 45, 172–180. [Google Scholar] [CrossRef] [PubMed]
- Matsunaga, K.; Klein, T.W.; Newton, C.; Friedman, H.; Yamamoto, Y. Legionella pneumophila suppresses Interleukin-12 production by macrophages. Infect. Immun. 2001, 69, 1929–1933. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vargas, D.; Vallejos-Vidal, E.; Reyes-Cerpa, S.; Oyarzún-Arrau, A.; Acuña-Castillo, C.; Imarai, M.; Reyes-López, F.E.; Sandino, A.M. The Analysis of Live-Attenuated Piscirickettsia salmonis Vaccine Reveals the Short-Term Upregulation of Innate and Adaptive Immune Genes in Atlantic Salmon (Salmo salar): An in Situ Open-Sea Cages Study. Microorganisms 2021, 9, 703. [Google Scholar] [CrossRef]
- Marshall, S.H.; Conejeros, P.; Zahr, M.; Olivares, J.; Gomez, F.; Cataldo, P.; Henriquez, V. Immunological characterization of a bacterial protein isolated from salmonid fish naturally infected with Piscirickettsia salmonis. Vaccine 2007, 25, 2095–2102. [Google Scholar] [CrossRef]
- San, M.; Briones, R. Quality control of commercial San Martı Quillaja (Quillaja saponaria Molina) extracts by reverse phase HPLC. J. Sci. Food Agric. 2000, 80, 2063–2068. [Google Scholar]
- Cheeke, P.; Otero, R. Yucca, quillaja may have role in animal nutrition. Feedstuffs 2005, 77, 1–7. [Google Scholar]
- Reichert, C.L.; Salminen, H.; Weiss, J. Quillaja Saponin Characteristics and Functional Properties. Annu. Rev. Food Sci. Technol. 2019, 10, 43–73. [Google Scholar] [CrossRef]
- Marston, H. Chemistry & Pharmacology of Natural Products; Saponins, K., Ed.; Cambridge University Press: New York, NY. USA, 1996. [Google Scholar]
- Marciani, D.J. Elucidating the mechanisms of action of saponin-derived adjuvants. Trends Pharm. Sci. 2018, 39, 573–585. [Google Scholar] [CrossRef] [PubMed]
- Lacaille-Dubois, M.-A.; Wagner, H. New perspectives for natural triterpene glycosides as potential adjuvants. Phytomedicine 2017, 37, 49–57. [Google Scholar] [CrossRef] [PubMed]
- Vinarova, L.; Vinarov, Z.; Atanasov, V.; Pantcheva, I.; Tcholakova, S.; Denkov, N.; Stoyanov, S. Lowering of cholesterol bioaccessibility and serum concentrations by saponins: In vitro and in vivo studies. Food Funct. 2014, 6, 501–512. [Google Scholar] [CrossRef] [PubMed]
- Schlotterbeck, T. Use of extracts of Quillaja saponaria for the prevention and control of bacterial infections in fish. Patent 2016, 1–18. [Google Scholar]
- Johnson, A. Saponins As Agents Preventing Infection Caused by Common Waterborne Pathogens. Ph.D. Thesis, University of Texas at Arlington, Arlington, TX, USA, 2013. [Google Scholar]
- Maier, C.; Conrad, J.; Carle, R.; Weiss, J.; Schweiggert, R.M. Phenolic constituents in commercial Aqueous Quillaja (Quillaja saponaria Molina) wood extracts. J. Agric. Food Chem. 2015, 63, 1756–1762. [Google Scholar] [CrossRef]
- Cheeke, P.R.; Piacente, S.; Oleszek, W. Anti-inflammatory and anti-arthritic effects of yucca schidigera: A review. J. Inflamm. 2006, 3, 6. [Google Scholar] [CrossRef] [Green Version]
- Kamstrup, S.; Martin, R.S.; Doberti, A.; Grande, H.; Dalsgaard, K. Preparation and characterisation of quillaja saponin with less heterogeneity than Quil-A. Vaccine 2000, 18, 2244–2249. [Google Scholar] [CrossRef]
- Mauel, M.J.; Ware, C.; Smith, P.A. Culture of Piscirickettsia salmonis on enriched blood agar. J. Veter. Diagn. Investig. 2008, 20, 213–214. [Google Scholar] [CrossRef] [Green Version]
- Henríquez, M.; Gonzalez, E.; Marshall, S.H.; Henríquez, V.; Gomez, F.A.; Martinez, I.; Altamirano, C. A Novel Liquid Medium for the Efficient Growth of the Salmonid Pathogen Piscirickettsia salmonis and Optimization of Culture Conditions. PLoS ONE 2013, 8, e71830. [Google Scholar] [CrossRef]
- Jensen, I.; Robertsen, B. Effect of double-stranded RNA and interferon on the antiviral activity of Atlantic salmon cells against infectious salmon anemia virus and infectious pancreatic necrosis virus. Fish Shellfish Immunol. 2002, 13, 221–241. [Google Scholar] [CrossRef] [PubMed]
- Gomez, F.A. Descripción, Caracterización y Proyección Biotecnológica de Genes Relacionados con la Virulencia en el Patógeno Piscirickettsia Salmonis, P. Doctorado en Biotecnología; Universidad Católica de Valparaíso—Universidad Técnica Federico Santa María: Valparaíso, Chile, 2012. [Google Scholar]
- Rodriguez-Tudela, J.; Arendrup, M.C.; Barchiesi, F.; Bille, J.; Chryssanthou, E.; Cuenca-Estrella, M.; Dannaoui, E.; Denning, D.; Donnelly, J.; Dromer, F.; et al. EUCAST Definitive Document EDef 7.1: Method for the determination of broth dilution MICs of antifungal agents for fermentative yeasts. Clin. Microbiol. Infect. 2008, 14, 398–405. [Google Scholar] [CrossRef] [PubMed]
- Riss, T.L.; Moravec, R.A.; Niles, A.L.; Duellman, S.; Benink, H.A.; Worzella, T.J.; Minor, L. Cell viability assays. Assay Guid. Man. 2016, 1–31. [Google Scholar]
- Olivares, J.; Marshall, S. Determination of minimal concentration ofPiscirickettsia salmonisin water columns to establish a fallowing period in salmon farms. J. Fish Dis. 2010, 33, 261–266. [Google Scholar] [CrossRef]
- Marshall, S.; Heath, S.; HenríquezV; Orrego, C. Minimally invasive detection of Piscirickettsia salmonis in cultivated Salmonids via the PCR. Appl. Environ. Microbiol. 1998, 64, 3066–3069. [Google Scholar] [CrossRef] [Green Version]
- Livak, K.J.; Schmittgen, T.D. Analysis of relative gene expression data using real-time quantitative PCR and the 2(-Delta Delta C(T)) Method. Methods 2001, 25, 402–408. [Google Scholar] [CrossRef]
- Flores-Herrera, P.; Arredondo-Zelada, O.; Marshall, S.H.; Gómez, F.A. Selection and validation of reliable housekeeping genes to evaluate Piscirickettsia salmonis gene expression. Infect. Genet. Evol. 2018, 63, 151–157. [Google Scholar] [CrossRef]
- Peña, A.; Bols, N.C.; Marshall, S.H. An evaluation of potential reference genes for stability of expression in two salmonid cell lines after infection with either Piscirickettsia salmonis or IPNV. BMC Res. Notes 2010, 3, 101. [Google Scholar] [CrossRef] [Green Version]
- Pfaffl, M.W.; Tichopad, A.; Prgomet, C.; Neuvians, T.P. Determination of stable housekeeping genes, differentially regulated target genes and sample integrity: BestKeeper—Excel-based tool using pair-wise correlations. Biotechnol. Lett. 2004, 26, 509–515. [Google Scholar] [CrossRef]
- Dungan, M. Micellar Properties of Quillaja Saponin 1. Effects of Temperature, Salt, and pH on Solution Properties. J. Agric. Food Chem. 1997, 45, 1587–1595. [Google Scholar]
- Cañon-Jones, H.; Cortes, H.; Castillo-Ruiz, M.; Schlotterbeck, T.; Martín, R.S. Quillaja saponaria (Molina) extracts inhibits in vitro Piscirickettsia salmonis Infections. Animals 2020, 10, 2286. [Google Scholar] [CrossRef]
- Hassan, S.M.; Byrd, J.A.; Cartwright, A.L.; Bailey, C.A. Hemolytic and antimicrobial activities differ among saponin-rich extracts from Guar, Quillaja, Yucca, and Soybean. Appl. Biochem. Biotechnol. 2010, 162, 1008–1017. [Google Scholar] [CrossRef]
- Arabski, M.; Węgierek-Ciuk, A.; Czerwonka, G.; Lankoff, A.; Kaca, W. Effects of saponins against Clinical E. coli Strains and Eukaryotic Cell Line. J. Biomed. Biotechnol. 2012, 2012, 1–6. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Antolak, H.; Mizerska, U.; Berłowska, J.; Otlewska, A.; Kręgiel, D. Quillaja saponaria saponins with potential to enhance the effectiveness of disinfection processes in the beverage industry. Appl. Sci. 2018, 8, 368. [Google Scholar] [CrossRef] [Green Version]
- Sewlikar, S.; D’Souza, D.H. Antimicrobial effects of Quillaja saponaria extract against Escherichia coli O157:H7 and the emerging Non-O157 Shiga toxin-producing E. coli. J. Food Sci. 2017, 82, 1171–1177. [Google Scholar] [CrossRef] [PubMed]
- Sen, S.; Makkar, H.P.; Muetzel, S.; Becker, K. Effect of Quillaja saponaria saponins and Yucca schidigera plant extract on growth of Escherichia coli. Lett. Appl. Microbiol. 1998, 27, 35–38. [Google Scholar] [CrossRef] [PubMed]
- Tam, K.I.; Roner, M.R. Characterization of in vivo anti-rotavirus activities of saponin extracts from Quillaja saponaria Molina. Antivir. Res. 2011, 90, 231–241. [Google Scholar] [CrossRef] [Green Version]
- Roner, M.R.; Sprayberry, J.; Spinks, M.; Dhanji, S. Antiviral activity obtained from aqueous extracts of the Chilean soapbark tree (Quillaja saponaria Molina). J. Gen. Virol. 2007, 88, 275–285. [Google Scholar] [CrossRef]
- Arayan, L.T.; Simborio, H.L.; Reyes, A.W.B.; Hop, H.T.; Min, W.; Lee, H.J.; Rhee, M.H.; Chang, H.H.; Kim, S. The effects of red ginseng saponin fraction-A (RGSF-A) on phagocytosis and intracellular signaling in Brucella abortus infected RAW 264.7 cells. FEMS Microbiol. Lett. 2015, 362, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Tacchi, L.; Bron, J.E.; Taggart, J.B.; Secombes, C.J.; Bickerdike, R.; Adler, M.A.; Takle, H.; Martin, S.A.M. Multiple tissue transcriptomic responses to Piscirickettsia salmonis in Atlantic salmon (Salmo salar). Physiol. Genom. 2011, 43, 1241–1254. [Google Scholar] [CrossRef] [Green Version]
- Salazar, C.; Haussmann, D.; Kausel, G.; Figueroa, J. Molecular cloning of Salmo salar Toll-like receptors (TLR1, TLR22, TLR5M and TLR5S) and expression analysis in SHK-1 cells during Piscirickettsia salmonis infection. J. Fish Dis. 2016, 39, 239–248. [Google Scholar] [CrossRef]
- Katayama, S.; Mine, Y. Quillaja saponin can modulate Ovalbumin-induced IgE allergic responses through regulation of Th1/Th2 balance in a Murine model. J. Agric. Food Chem. 2006, 54, 3271–3276. [Google Scholar] [CrossRef] [PubMed]
- Troian, E.A.; Schallenberger, K.; da Silva, F.P.; Dietrich, G.K.; Ferreira Chiesa, F.; Olivaro, C.; Wallace, F.; Fleck, J.; Verza, S. Screening for antiviral activity of two purified saponin fractions of Quillaja spp. against Yellow Fever Virus and Chikungunya Virus. Int. J. Innov. Educ. Res. 2020, 8, 205–214. [Google Scholar] [CrossRef]
- Roner, M.R.; Tam, K.I.; Kiesling-Barrager, M. Prevention of rotavirus infections in vitro with aqueous extracts of Quillaja Saponaria Molina. Future Med. Chem. 2010, 2, 1083–1097. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sparg, S.G.; Light, M.E.; Van Staden, J. Biological activities and distribution of plant saponins. J. Ethnopharmacol. 2004, 94, 219–243. [Google Scholar] [CrossRef]
- Augustin, J.M.; Kuzina, V.; Andersen, S.B.; Bak, S. Molecular activities, biosynthesis and evolution of triterpenoid saponins. Phytochemistry 2011, 72, 435–457. [Google Scholar] [CrossRef] [PubMed]
- Imarai, M.; Acua, C.; Escobar, A.; Maisey, K.; Reyes-Cerp, S. Mucosal immunity and evasion strategies of Neisseria gonorrhoeae. Sex. Transm. Infect. 2012. [Google Scholar] [CrossRef] [Green Version]
- Rai, A.; Pathak, D.; Thakur, S.; Singh, S.; Dubey, A.K.; Mallik, R. Dynein clusters into lipid microdomains on Phagosomes to drive rapid transport toward Lysosomes. Cell 2016, 164, 722–734. [Google Scholar] [CrossRef] [Green Version]
- Triantafilou, M.; Miyake, K.; Golenbock, U.T.; Triantafilou, K. Mediators of innate immune recognition of bacteria concentrate in lipid rafts and facilitate lipopolysaccharide-induced cell activation. J. Cell Sci. 2002, 115, 2603–2611. [Google Scholar] [CrossRef]
- Böttger, S.; Melzig, M.F. The influence of saponins on cell membrane cholesterol. Bioorg. Med. Chem. 2013, 21, 7118–7124. [Google Scholar] [CrossRef]
- Barman, S.; Nayak, D.P. Lipid Raft Disruption by Cholesterol Depletion Enhances Influenza A Virus Budding from MDCK Cells. J. Virol. 2007, 81, 12169–12178. [Google Scholar] [CrossRef] [Green Version]
- Lee, J.J.; Kim, D.G.; Kim, D.H.; Simborio, H.L.; Min, W.; Lee, H.J.; Her, M.; Jung, S.C.; Watarai, M.; Kim, S. Interplay between Clathrin and Rab5 controls the early phagocytic trafficking and intracellular survival of brucella abortus within HeLa cells. J. Biol. Chem. 2013, 288, 28049–28057. [Google Scholar] [CrossRef] [Green Version]
- Alarcon, P.; Espinosa, G.; Millan, C.; Saravia, J.; Quinteros, V.; Enriquez, R.; Henriquez, C.; Vargas-Chacoff, L.; Burgos, R.A.; Taubert, A.; et al. Piscirickettsia salmonis-Triggered Extracellular Traps Formation as an Innate Immune Response of Atlantic Salmon-Derived Polymorphonuclear Neutrophils. Biology 2021, 10, 206. [Google Scholar] [CrossRef]
- Huynh, K.K.; Gershenzon, E.; Grinstein, S. Cholesterol accumulation by macrophages impairs Phagosome maturation. J. Biol. Chem. 2008, 283, 35745–35755. [Google Scholar] [CrossRef] [Green Version]
- Boyaka, P.N.; Marinaro, M.; Jackson, R.J.; Menon, S.; Kiyono, H.; Jirillo, E.; McGhee, J.R. IL-12 is an effective adjuvant for induction of mucosal immunity. J. Immunol. 1999, 162, 122–128. [Google Scholar] [PubMed]
- Kensil, C.R.; Patel, U.; Lennick, M.; Marciani, D. Separation and characterization of saponins with adjuvant activity from Quillaja saponaria Molina cortex. J. Immunol. 1991, 146, 431–437. [Google Scholar] [PubMed]
- Maharaj, I.; Froh, K.J.; Campbell, J.B. Immune responses of mice to inactivated rabies vaccine administered orally: Potentiation by Quillaja saponin. Can. J. Microbiol. 2010, 32, 414–420. [Google Scholar] [CrossRef] [PubMed]
- Boyaka, P.N.; Marinaro, M.; Jackson, R.J.; Van Ginkel, F.W.; Cormet-Boyaka, E.; Kirk, K.L.; Kensil, C.R.; McGhee, J.R. Oral QS-21 requires early IL-4 Help for induction of mucosal and systemic immunity. J. Immunol. 2001, 166, 2283–2290. [Google Scholar] [CrossRef]
- Kensil, C.R. Immunomodulatory adjuvants from Quillaja saponaria. Immunopotentiators Mod. Vaccines 1996. [Google Scholar] [CrossRef]
- Marty-Roix, R.; Vladimer, G.; Pouliot, K.; Weng, D.; Buglione-Corbett, R.; West, K.; MacMicking, J.D.; Chee, J.D.; Wang, S.; Lu, S.; et al. Identification of QS-21 as an Inflammasome-activating molecular component of saponin adjuvants. J. Biol. Chem. 2016, 291, 1123–1136. [Google Scholar] [CrossRef] [Green Version]
- Wojciechowski, K.; Orczyk, M.; Gutberlet, T.; Trapp, M.; Marcinkowski, K.; Kobiela, T.; Geue, T. Unusual penetration of phospholipid mono- and bilayers by Quillaja bark saponin biosurfactant. Biochim. Biophys. Acta Bba Biomembr. 2014, 1838, 1931–1940. [Google Scholar] [CrossRef] [Green Version]
- Behboudi, S.; Morein, B.; Villacres-Eriksson, M. In vitro activation of antigen-presenting cells (APC) by defined composition of Quillaja saponaria Molina triterpenoids. Clin. Exp. Immunol. 1996, 105, 26–30. [Google Scholar] [CrossRef] [PubMed]
- Takahashi, H.; Takeshfta, T.; Morein, B.; Putney, S.; Germain, R.N.; Berzofsky, J.A. Induction of CD8+ cytotoxic T cells by immunization with purified HIV-1 envelope protein in ISCOMs. Nature 1990, 344, 873–875. [Google Scholar] [CrossRef] [PubMed]
- Didierlaurent, A.M.; Laupèze, B.; Di Pasquale, A.; Hergli, N.; Collignon, C.; Garçon, N. Adjuvant system AS01: Helping to overcome the challenges of modern vaccines. Expert Rev. Vaccines 2017, 16, 55–63. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Welsby, I.; Detienne, S.; N’Kuli, F.; Thomas, S.; Wouters, S.; Bechtold, V.; De Wit, D.; Gineste, R.; Reinheckel, T.; Elouahabi, A.; et al. Lysosome-dependent activation of human Dendritic cells by the vaccine adjuvant QS-21. Front. Immunol. 2017, 7, 526. [Google Scholar] [CrossRef] [Green Version]















{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Gene | Primer Sequence | Reference |
|---|---|---|
| sdhA | For: 5′-ATTTCTTTGGAGCTACGTGAAG-3′ | Flores-Herrera et al., 2018 |
| Rev: 5′-CCACCCATCATATAATGACAAG-3′ | ||
| elf1A | For: 5′-GTC TAC AAA ATC GGC GGT AT-3′ | Peña et al., 2010 |
| Rev: 5′-CTT GAC GGA CAC GTT CTT GA-3′ | ||
| ITS | RTS1; For-223: 5′-TGATTTTATTGTTTAGTGAGAATGA-3′ | Marshall et al., 1998 |
| RTS4; Rev-459: 5′-ATGCACTTATTCACTTGATCATA-3′ | ||
| dotB | Ps-dotB-For: 5′-GCT ACA TCT CCA TTT CTT GAC CAT TTC-3′ | Gómez et al., 2013 |
| Ps-dotB-Rev: 5′- GCA TTA GTG CCG AGC ATT ACA GG-3′ | ||
| chaPs | For: 5′-GATGAAAGAGAAGAAAGACCGC-3′ | Marshall et al., 2007 |
| Rev: 5′-ATGGGCGGCATGGGCGGCATGATG-3′ | ||
| IL-10 | For: GCCCTTCAGTAACTTACACAGATGGAC | Harun et al., 2011 |
| Rev: GTCGTTGTTGTTCTGTGTTCTGTTGT | ||
| IL-12 | For: CCCAACACGGACAGGAACAC | Wang et al., 2014 |
| Rev: GCCCTTCAGTAACTTACACAGATGGAC |
| Panel A—Intracellular Proliferation of P. salmonis at 1 hpi | ||||
| Treatment | Time hpi | Ct (dRn) | No. Copies/Samples | % Inhibition |
| Negative Control, NC | 1 | 38.51 ± 0.11 | 0.0 | N/A |
| Positive Control, PC (P. salmonis) | 1 | 29.68 ± 0.12 | 2377 ± 1642 | 0 |
| PQSE (0.5 µg/mL) + P. salmonis | 1 | 31.66 ± 0.07 | 467 ± 304 | 80 |
| Panel B—Intracellular Proliferation of P. salmonis at 72 hpi | ||||
| Treatment | Time hpi | Ct (dRn) | Number of Copies/Samples | % Inhibition |
| Negative Control, NC | 72 | 38.41 ± 0.22 | 0.0 | |
| Positive Control, PC (P. salmonis) | 72 | 24.44 ± 0.01 | 33,479 ± 7730 | |
| PQSE (0.5 µg/mL) + P. salmonis | 72 | 27.58 ± 1.18 | 8026 ± 4514 | 76% |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Cortés, H.D.; Gómez, F.A.; Marshall, S.H. The Phagosome–Lysosome Fusion Is the Target of a Purified Quillaja saponin Extract (PQSE) in Reducing Infection of Fish Macrophages by the Bacterial Pathogen Piscirickettsia salmonis. Antibiotics 2021, 10, 847. https://doi.org/10.3390/antibiotics10070847
Cortés HD, Gómez FA, Marshall SH. The Phagosome–Lysosome Fusion Is the Target of a Purified Quillaja saponin Extract (PQSE) in Reducing Infection of Fish Macrophages by the Bacterial Pathogen Piscirickettsia salmonis. Antibiotics. 2021; 10(7):847. https://doi.org/10.3390/antibiotics10070847
Chicago/Turabian StyleCortés, Hernán D., Fernando A. Gómez, and Sergio H. Marshall. 2021. "The Phagosome–Lysosome Fusion Is the Target of a Purified Quillaja saponin Extract (PQSE) in Reducing Infection of Fish Macrophages by the Bacterial Pathogen Piscirickettsia salmonis" Antibiotics 10, no. 7: 847. https://doi.org/10.3390/antibiotics10070847