
Methicillin-Resistant Staphylococcus aureus Clonal Complex 398 as a Major MRSA Lineage in Dogs and Cats in Thailand

 ,
,
Abstract
:1. Introduction
2. Results
2.1. Numbers and Origins of MRSA
2.2. Genetic Characteristics of Canine and Feline MRSA CC398
2.3. Antimicrobial Resistance Gene Carriage
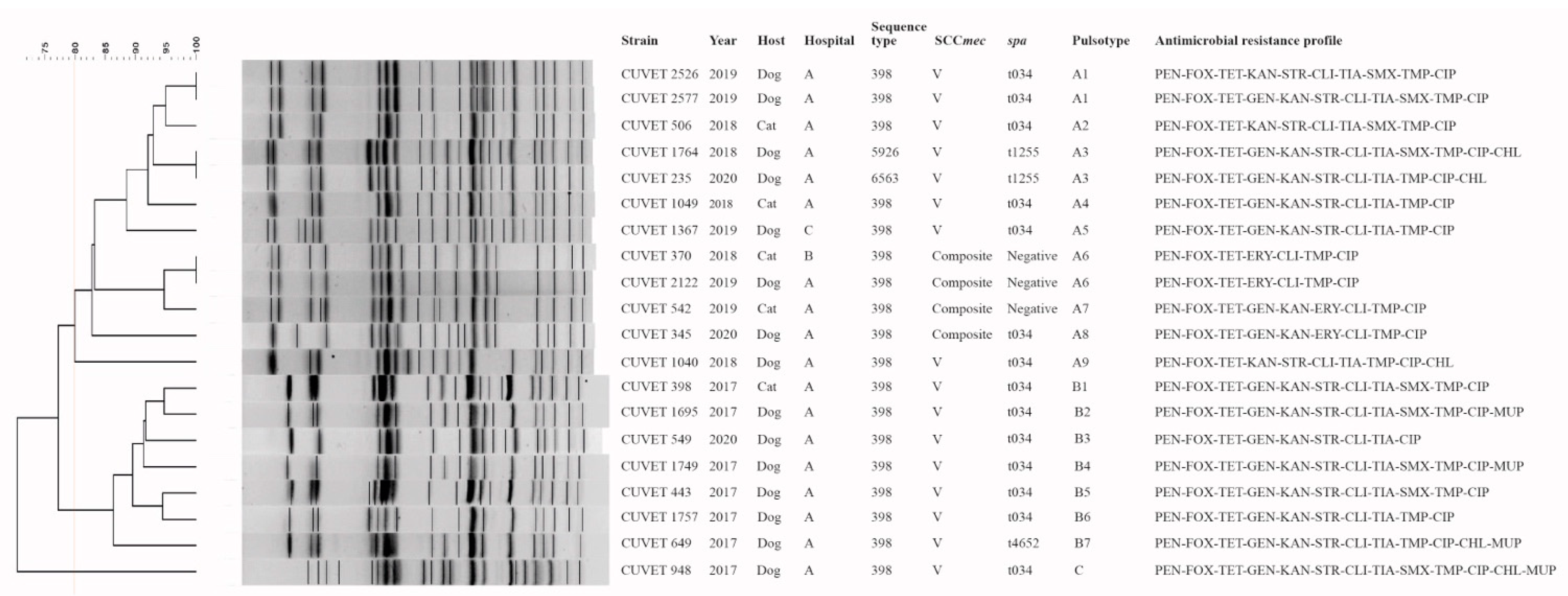
2.4. Intraclonal Variation and Common Features of Canine and Feline MRSA CC398
3. Discussion
4. Materials and Methods
4.1. Bacterial Isolates
4.2. Antimicrobial Susceptibility Testing and Resistance Gene Detection
4.3. DNA Fingerprint Analysis
4.4. spa Typing
4.5. SCCmec Identification and MLST
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Weese, J.S.; Van Duijkeren, E. Methicillin-resistant Staphylococcus aureus and Staphylococcus pseudintermedius in veterinary medicine. Vet. Microbiol. 2010, 140, 418–429. [Google Scholar] [CrossRef] [PubMed]
- Stefani, S.; Chung, D.R.; Lindsay, J.A.; Friedrich, A.W.; Kearns, A.M.; Westh, H.; Mackenzie, F.M. Meticillin-resistant Staphylococcus aureus (MRSA): Global epidemiology and harmonisation of typing methods. Int. J. Antimicrob. Agents 2012, 39, 273–282. [Google Scholar] [CrossRef]
- Mediavilla, J.R.; Chen, L.; Mathema, B.; Kreiswirth, B.N. Global epidemiology of community-associated methicillin resistant Staphylococcus aureus (CA-MRSA). Curr. Opin. Microbiol. 2012, 15, 588–595. [Google Scholar] [CrossRef]
- Popovich, K.J.; Weinstein, R.A.; Hota, B. Are community-associated methicillin-resistant Staphylococcus aureus (MRSA) strains replacing traditional nosocomial MRSA strains? Clin. Infect. Dis. 2008, 46, 787–794. [Google Scholar] [CrossRef] [PubMed]
- Davis, S.L.; Rybak, M.J.; Amjad, M.; Kaatz, G.W.; McKinnon, P.S. Characteristics of patients with healthcare-associated infection due to SCCmec type IV methicillin-resistant Staphylococcus aureus. Infect. Control. Hosp. Epidemiol. 2006, 27, 1025–1031. [Google Scholar] [CrossRef]
- Seybold, U.; Kourbatova, E.V.; Johnson, J.G.; Halvosa, S.J.; Wang, Y.F.; King, M.D.; Ray, S.M.; Blumberg, H.M. Emergence of community-associated methicillin-resistant Staphylococcus aureus USA300 genotype as a major cause of health care-associated blood stream infections. Clin. Infect. Dis. 2006, 42, 647–656. [Google Scholar] [CrossRef] [Green Version]
- Haenni, M.; Saras, E.; Châtre, P.; Médaille, C.; Bes, M.; Madec, J.Y.; Laurent, F. A USA300 variant and other human-related methicillin-resistant Staphylococcus aureus strains infecting cats and dogs in France. J. Antimicrob. Chemother. 2012, 67, 326–329. [Google Scholar] [CrossRef] [Green Version]
- Haenni, M.; Châtre, P.; Dupieux-Chabert, C.; Métayer, V.; Bes, M.; Madec, J.Y.; Laurent, F. Molecular epidemiology of methicillin-resistant Staphylococcus aureus in horses, cats and dogs over a 5-year period in France. Front. Microbiol. 2017, 8, 2493. [Google Scholar] [CrossRef]
- Loncaric, I.; Lepuschitz, S.; Ruppitsch, W.; Trstan, A.; Andreadis, T.; Bouchlis, N.; Marbach, H.; Schauer, B.; Szostak, M.P.; Feßler, A.T.; et al. Increased genetic diversity of methicillin-resistant Staphylococcus aureus (MRSA) isolated from companion animals. Vet. Microbiol. 2019, 235, 118–126. [Google Scholar] [CrossRef] [PubMed]
- Perreten, V.; Kadlec, K.; Schwarz, S.; Grönlund Andersson, U.; Finn, M.; Greko, C.; Moodley, A.; Kania, S.A.; Frank, L.A.; Bemis, D.A.; et al. Clonal spread of methicillin-resistant Staphylococcus pseudintermedius in Europe and North America: An international multicentre study. J. Antimicrob. Chemother. 2010, 65, 1145–1154. [Google Scholar] [CrossRef] [Green Version]
- Vincze, S.; Stamm, I.; Kopp, P.A.; Hermes, J.; Adlhoch, C.; Semmler, T.; Wieler, L.H.; Lübke-Becker, A.; Walther, B. Alarming proportions of methicillin-resistant Staphylococcus aureus (MRSA) in wound samples from companion animals, Germany 2010-2012. PLoS ONE 2014, 9, e85656. [Google Scholar] [CrossRef] [Green Version]
- Vincze, S.; Brandenburg, A.G.; Espelage, W.; Stamm, I.; Wieler, L.H.; Kopp, P.A.; Lübke-Becker, A.; Walther, B. Risk factors for MRSA infection in companion animals: Results from a case-control study within Germany. Int J. Med. Microbiol. 2014, 304, 787–793. [Google Scholar] [CrossRef]
- Taniguchi, Y.; Koide, S.; Maeyama, Y.; Tamai, K.; Hayashi, W.; Tanaka, H.; Iimura, M.; Suzuki, M.; Nagano, Y.; Arakawa, Y.; et al. Predominance of methicillin-resistant Staphylococcus aureus SCCmec type II-CC5 and SCCmec type IV-CC1/CC8 among companion animal clinical isolates in Japan: Findings from phylogenetic comparison with human clinical isolates. J. Glob. Antimicrob. Resist. 2020, 20, 253–259. [Google Scholar] [CrossRef] [PubMed]
- Manian, F.A. Asymptomatic nasal carriage of mupirocin-resistant, methicillin-resistant Staphylococcus aureus (MRSA) in a pet dog associated with MRSA infection in household contacts. Clin. Infect. Dis. 2003, 36, e26–e28. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Asanin, J.; Misic, D.; Aksentijevic, K.; Tambur, Z.; Rakonjac, B.; Kovacevic, I.; Spergser, J.; Loncaric, I. Genetic profiling and comparison of human and animal methicillin-resistant Staphylococcus aureus (MRSA) isolates from Serbia. Antibiotics 2019, 8, 26. [Google Scholar] [CrossRef] [Green Version]
- Drougka, E.; Foka, A.; Koutinas, C.K.; Jelastopulu, E.; Giormezis, N.; Farmaki, O.; Sarrou, S.; Anastassiou, E.D.; Petinaki, E.; Spiliopoulou, I. Interspecies spread of Staphylococcus aureus clones among companion animals and human close contacts in a veterinary teaching hospital. A cross-sectional study in Greece. Prev. Vet. Med. 2016, 126, 190–198. [Google Scholar] [CrossRef] [PubMed]
- Walther, B.; Friedrich, A.W.; Brunnberg, L.; Wieler, L.H.; Lübke-Becker, A. Methicillin-resistant Staphylococcus aureus (MRSA) in veterinary medicine: A “new emerging pathogen”? Berl. Munch. Tierarztl. Wochenschr. 2006, 119, 222–232. [Google Scholar] [PubMed]
- Vincze, S.; Stamm, I.; Monecke, S.; Kopp, P.A.; Semmler, T.; Wieler, L.H.; Lübke-Becker, A.; Walther, B. Molecular analysis of human and canine Staphylococcus aureus strains reveals distinct extended-host-spectrum genotypes independent of their methicillin resistance. Appl Environ. Microbiol. 2013, 79, 655–662. [Google Scholar] [CrossRef] [Green Version]
- Worthing, K.A.; Abraham, S.; Pang, S.; Coombs, G.W.; Saputra, S.; Jordan, D.; Wong, H.S.; Abraham, R.J.; Trott, D.J.; Norris, J.M. Molecular characterization of methicillin-resistant Staphylococcus aureus isolated from Australian animals and veterinarians. Microb. Drug Resist. 2018, 24, 203–212. [Google Scholar] [CrossRef]
- Chanchaithong, P.; Perreten, V.; Schwendener, S.; Tribuddharat, C.; Chongthaleong, A.; Niyomtham, W.; Prapasarakul, N. Strain typing and antimicrobial susceptibility of methicillin-resistant coagulase-positive staphylococcal species in dogs and people associated with dogs in Thailand. J. Appl. Microbiol. 2014, 117, 572–586. [Google Scholar] [CrossRef] [Green Version]
- Chanchaithong, P.; Perreten, V.; Am-In, N.; Lugsomya, K.; Tummaruk, P.; Prapasarakul, N. Molecular characterization and antimicrobial resistance of livestock-associated methicillin-resistant Staphylococcus aureus isolates from pigs and swine workers in central Thailand. Microb. Drug Resist. 2019, 25, 1382–1389. [Google Scholar] [CrossRef] [PubMed]
- Zhang, W.; Hao, Z.; Wang, Y.; Cao, X.; Logue, C.M.; Wang, B.; Yang, J.; Shen, J.; Wu, C. Molecular characterization of methicillin-resistant Staphylococcus aureus strains from pet animals and veterinary staff in China. Vet. J. 2011, 190, e125–e129. [Google Scholar] [CrossRef]
- Chuang, Y.Y.; Huang, Y.C. Livestock-associated meticillin-resistant Staphylococcus aureus in Asia: An emerging issue? Int. J. Antimicrob. Agents 2015, 45, 334–340. [Google Scholar] [CrossRef]
- Verhegghe, M.; Pletinckx, L.J.; Crombé, F.; Vandersmissen, T.; Haesebrouck, F.; Butaye, P.; Heyndrickx, M.; Rasschaert, G. Methicillin-resistant Staphylococcus aureus (MRSA) ST398 in pig farms and multispecies farms. Zoonoses Public Hlth. 2013, 60, 366–374. [Google Scholar] [CrossRef]
- Sahibzada, S.; Abraham, S.; Coombs, G.W.; Pang, S.; Hernández-Jover, M.; Jordan, D.; Heller, J. Transmission of highly virulent community-associated MRSA ST93 and livestock-associated MRSA ST398 between humans and pigs in Australia. Sci. Rep. 2017, 7, 5273. [Google Scholar] [CrossRef] [PubMed]
- Takeuchi, F.; Watanabe, S.; Baba, T.; Yuzawa, H.; Ito, T.; Morimoto, Y.; Kuroda, M.; Cui, L.; Takahashi, M.; Ankai, A.; et al. Whole-genome sequencing of Staphylococcus haemolyticus uncovers the extreme plasticity of its genome and the evolution of human-colonizing staphylococcal species. J. Bacteriol 2005, 187, 7292–7308. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kadlec, K.; Schwarz, S. Antimicrobial resistance of Staphylococcus pseudintermedius. Vet. Dermatol. 2012, 23, 276–282, e255. [Google Scholar] [CrossRef]
- Kern, A.; Perreten, V. Clinical and molecular features of methicillin-resistant, coagulase-negative staphylococci of pets and horses. J. Antimicrob. Chemother. 2013, 68, 1256–1266. [Google Scholar] [CrossRef]
- Perreten, V.; Chanchaithong, P.; Prapasarakul, N.; Rossano, A.; Blum, S.E.; Elad, D.; Schwendener, S. Novel pseudo-staphylococcal cassette chromosome mec element (ψSCCmec57395) in methicillin-resistant Staphylococcus pseudintermedius CC45. Antimicrob. Agents Chemother. 2013, 57, 5509–5515. [Google Scholar] [CrossRef] [Green Version]
- Kadlec, K.; Fessler, A.T.; Hauschild, T.; Schwarz, S. Novel and uncommon antimicrobial resistance genes in livestock-associated methicillin-resistant Staphylococcus aureus. Clin. Microbiol. Infect. 2012, 18, 745–755. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Wendlandt, S.; Kadlec, K.; Feßler, A.T.; Schwarz, S. Identification of ABC transporter genes conferring combined pleuromutilin-lincosamide-streptogramin A resistance in bovine methicillin-resistant Staphylococcus aureus and coagulase-negative staphylococci. Vet. Microbiol. 2015, 177, 353–358. [Google Scholar] [CrossRef]
- Li, B.; Wendlandt, S.; Yao, J.; Liu, Y.; Zhang, Q.; Shi, Z.; Wei, J.; Shao, D.; Schwarz, S.; Wang, S.; et al. Detection and new genetic environment of the pleuromutilin-lincosamide-streptogramin A resistance gene lsa(E) in methicillin-resistant Staphylococcus aureus of swine origin. J. Antimicrob. Chemother. 2013, 68, 1251–1255. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bloom, P. Canine superficial bacterial folliculitis: Current understanding of its etiology, diagnosis and treatment. Vet. J. 2014, 199, 217–222. [Google Scholar] [CrossRef] [PubMed]
- Hillier, A.; Lloyd, D.H.; Weese, J.S.; Blondeau, J.M.; Boothe, D.; Breitschwerdt, E.; Guardabassi, L.; Papich, M.G.; Rankin, S.; Turnidge, J.D.; et al. Guidelines for the diagnosis and antimicrobial therapy of canine superficial bacterial folliculitis (Antimicrobial Guidelines Working Group of the International Society for Companion Animal Infectious Diseases). Vet. Dermatol. 2014, 25, e143–e163. [Google Scholar] [CrossRef]
- Mueller, R.S.; Bergvall, K.; Bensignor, E.; Bond, R. A review of topical therapy for skin infections with bacteria and yeast. Vet. Dermatol. 2012, 23, 330–341, e362. [Google Scholar] [CrossRef]
- Couto, N.; Belas, A.; Rodrigues, C.; Schwarz, S.; Pomba, C. Acquisition of the fexA and cfr genes in Staphylococcus pseudintermedius during florfenicol treatment of canine pyoderma. J. Glob. Antimicrob Resist. 2016, 7, 126–127. [Google Scholar] [CrossRef]
- Kadlec, K.; Van Duijkeren, E.; Wagenaar, J.A.; Schwarz, S. Molecular basis of rifampicin resistance in methicillin-resistant Staphylococcus pseudintermedius isolates from dogs. J. Antimicrob. Chemother. 2011, 66, 1236–1242. [Google Scholar] [CrossRef] [Green Version]
- Feßler, A.T.; Schuenemann, R.; Kadlec, K.; Hensel, V.; Brombach, J.; Murugaiyan, J.; Oechtering, G.; Burgener, I.A.; Schwarz, S. Methicillin-resistant Staphylococcus aureus (MRSA) and methicillin-resistant Staphylococcus pseudintermedius (MRSP) among employees and in the environment of a small animal hospital. Vet. Microbiol. 2018, 221, 153–158. [Google Scholar] [CrossRef] [PubMed]
- Neradova, K.; Jakubu, V.; Pomorska, K.; Zemlickova, H. Methicillin-resistant Staphylococcus aureus in veterinary professionals in 2017 in the Czech Republic. BMC Vet. Res. 2020, 16, 4. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Floras, A.; Lawn, K.; Slavic, D.; Golding, G.R.; Mulvey, M.R.; Weese, J.S. Sequence type 398 meticillin-resistant Staphylococcus aureus infection and colonisation in dogs. Vet. Rec. 2010, 166, 826–827. [Google Scholar] [CrossRef] [PubMed]
- Price, L.B.; Stegger, M.; Hasman, H.; Aziz, M.; Larsen, J.; Andersen, P.S.; Pearson, T.; Waters, A.E.; Foster, J.T.; Schupp, J.; et al. Staphylococcus aureus CC398: Host adaptation and emergence of methicillin resistance in livestock. mBio 2012, 3. [Google Scholar] [CrossRef] [Green Version]
- Viana, D.; Blanco, J.; Tormo-Más, M.A.; Selva, L.; Guinane, C.M.; Baselga, R.; Corpa, J.; Lasa, I.; Novick, R.P.; Fitzgerald, J.R.; et al. Adaptation of Staphylococcus aureus to ruminant and equine hosts involves SaPI-carried variants of von Willebrand factor-binding protein. Mol. Microbiol. 2010, 77, 1583–1594. [Google Scholar] [CrossRef]
- Murray, S.; Pascoe, B.; Méric, G.; Mageiros, L.; Yahara, K.; Hitchings, M.D.; Friedmann, Y.; Wilkinson, T.S.; Gormley, F.J.; Mack, D.; et al. Recombination-mediated host adaptation by avian Staphylococcus aureus. Genome Biol. Evol. 2017, 9, 830–842. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Harrison, E.M.; Weinert, L.A.; Holden, M.T.; Welch, J.J.; Wilson, K.; Morgan, F.J.; Harris, S.R.; Loeffler, A.; Boag, A.K.; Peacock, S.J.; et al. A shared population of epidemic methicillin-resistant Staphylococcus aureus 15 circulates in humans and companion animals. mBio 2014, 5. [Google Scholar] [CrossRef] [Green Version]
- Sasaki, T.; Tsubakishita, S.; Tanaka, Y.; Sakusabe, A.; Ohtsuka, M.; Hirotaki, S.; Kawakami, T.; Fukata, T.; Hiramatsu, K. Multiplex-PCR method for species identification of coagulase-positive staphylococci. J. Clin. Microbiol. 2010, 48, 765–769. [Google Scholar] [CrossRef] [Green Version]
- Strommenger, B.; Kettlitz, C.; Werner, G.; Witte, W. Multiplex PCR assay for simultaneous detection of nine clinically relevant antibiotic resistance genes in Staphylococcus aureus. J. Clin. Microbiol 2003, 41, 4089–4094. [Google Scholar] [CrossRef] [Green Version]
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobial Susceptibility Testing, 30th ed.; 2020; Volume M100, Available online: https://clsi.org/media/3481/m100ed30_sample.pdf (accessed on 17 December 2020).
- Clinical and Laboratory Standards Institute. Performance Standards for Antimicrobsial Disk and Dilution Susceptibility Tests for Bacteria Isolated From Animals, 5th ed.; 2020; Volume VET01S, Available online: https://clsi.org/media/m1gppkx2/vet01sed5_sample.pdf (accessed on 17 December 2020).
- Moreira de Oliveira, N.E.; Cardozo, A.P.C.M.; Marques, E.A.; Netto Dos Santos, K.R.; De Marval, M.G. Interpretive criteria to differentiate low- and high-level mupirocin resistance in Staphylococcus aureus. J. Med. Microbiol. 2007, 56, 937–939. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Skov, R.; Frimodt-Møller, N.; Espersen, F. Correlation of MIC methods and tentative interpretive criteria for disk diffusion susceptibility testing using NCCLS methodology for fusidic acid. Diagn. Microbiol. Infect. Dis. 2001, 40, 111–116. [Google Scholar] [CrossRef]
- Alba, P.; Feltrin, F.; Cordaro, G.; Porrero, M.C.; Kraushaar, B.; Argudín, M.A.; Nykäsenoja, S.; Monaco, M.; Stegger, M.; Aarestrup, F.M.; et al. Livestock-associated methicillin resistant and methicillin susceptible Staphylococcus aureus sequence type (CC)1 in European farmed animals: High genetic relatedness of isolates from Italian cattle herds and humans. PLoS ONE 2015, 10, e0137143. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Argudín, M.A.; Rodicio, M.R.; Guerra, B. The emerging methicillin-resistant Staphylococcus aureus ST398 clone can easily be typed using the Cfr9I SmaI-neoschizomer. Lett. Appl. Microbiol. 2010, 50, 127–130. [Google Scholar] [CrossRef]
- Harmsen, D.; Claus, H.; Witte, W.; Rothgänger, J.; Turnwald, D.; Vogel, U. Typing of methicillin-resistant Staphylococcus aureus in a university hospital setting by using novel software for spa repeat determination and database management. J. Clin. Microbiol 2003, 41, 5442–5448. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strommenger, B.; Braulke, C.; Heuck, D.; Schmidt, C.; Pasemann, B.; Nübel, U.; Witte, W. spa Typing of Staphylococcus aureus as a frontline tool in epidemiological typing. J. Clin. Microbiol. 2008, 46, 574–581. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kondo, Y.; Ito, T.; Ma, X.X.; Watanabe, S.; Kreiswirth, B.N.; Etienne, J.; Hiramatsu, K. Combination of multiplex PCRs for staphylococcal cassette chromosome mec type assignment: Rapid identification system for mec, ccr, and major differences in junkyard regions. Antimicrob. Agents Chemother. 2007, 51, 264–274. [Google Scholar] [CrossRef] [Green Version]
- (IWG-SCC), I.W.G.o.t.C.o.S.C.C.E. Classification of staphylococcal cassette chromosome mec (SCCmec): Guidelines for reporting novel SCCmec elements. Antimicrob. Agents Chemother. 2009, 53, 4961–4967. [Google Scholar] [CrossRef] [Green Version]
- Enright, M.C.; Day, N.P.; Davies, C.E.; Peacock, S.J.; Spratt, B.G. Multilocus sequence typing for characterization of methicillin-resistant and methicillin-susceptible clones of Staphylococcus aureus. J. Clin. Microbiol. 2000, 38, 1008–1015. [Google Scholar] [CrossRef] [PubMed] [Green Version]


{kind=link}
| Antimicrobials * | Resistance Break Point (μg/mL) | Range Tested (μg/mL) | Number of Isolates in Each MIC (μg/mL) ** | Number of Resistant Isolates | Resistance Gene(s) *** | Number of Isolates with the Presence of Resistance Genes *** | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| ≤0.015 | 0.003 | 0.006 | 0.012 | 0.25 | 0.5 | 1 | 2 | 4 | 8 | 16 | 32 | 64 | 128 | 256 | 512 | ||||||
| PEN | ≥0.25 | 0.0012–2 | 2 (=) | 18 (>) | 20/20 | blaZ; mecA | 18; 20 | ||||||||||||||
| FOX | ≥1 | 0.5–16 | 3 (=) | 6 (=) | 11 (>) | 20/20 | mecA | 20 | |||||||||||||
| TET | ≥1 | 0.5–16 | 20 (>) | 20/20 | tet(M); tet(L); tet(K) | 20; 16; 0 | |||||||||||||||
| GEN | ≥16 | 1–16 | 2 (≤) | 3 (=) | 8 (=) | 7 (>) | 15/20 | aac(6′)-Ie-aph(2′)-Ia | 18 | ||||||||||||
| KAN | ≥16 | 4–64 | 2 (≤) | 9 (=); 9 (>) | 18/20 | aac(6′)-Ie-aph(2′)-Ia; ant(4′)-Ia | 18; 17 | ||||||||||||||
| STR | ≥16 | 4–32 | 1 (≤) | 3 (=) | 16 (>) | 16/20 | ant(4′)-Ia; ant(6)-Ia | 17; 16 | |||||||||||||
| ERY | ≥8 | 0.25–8 | 4 (≤) | 10 (=) | 1 (=) | 1 (=) | 4 (>) | 4/20 | erm(B); erm(A); erm(C) | 4; 3; 0 | |||||||||||
| CLI | ≥4 | 0.12–4 | 20 (>) | 20/20 | lnu(B); erm(B); erm(A); erm(C) | 16; 4; 3; 0 | |||||||||||||||
| CHL | ≥32 | 4–64 | 6 (≤) | 5 (=) | 4 (=) | 5 (=) | 5/20 | fexA; catpC221 | 5; 0 | ||||||||||||
| SMX | ≥512 | 64–512 | 6 (≤) | 5 (=) | 7 (>); 2 (=) | 9/20 | ND | ND | |||||||||||||
| TMP | ≥16 | 2–32 | 1 (≤) | 19 (>) | 19/20 | dfrA; dfrG | 17; 4 | ||||||||||||||
| MUP | ≥512 | 0.5–2 & 256 | 16 (≤) | 4 (>) | 4/20 | ileS2 | 4 | ||||||||||||||
| VAN | ≥16 | 1–16 | 19 (≤) | 2 (=) | 0/20 | ND | 0 | ||||||||||||||
| CIP | ≥4 | 0.25–8 | 1 (=); 19 (>) | 20/20 | ND | ND | |||||||||||||||
| RIF | ≥4 | 0.015–0.5 | 19 (≤) | 1 (=) | 0/20 | ND | ND | ||||||||||||||
| LZD | ≥8 | 1–8 | 1 (≤) | 5 (=) | 13 (=) | 1 (=) | 0/20 | ND | ND | ||||||||||||
| FUS | ≥2 | 0.5–4 | 20 (≤) | 0/20 | ND | ND | |||||||||||||||
| TIA | >4 | 0.5–4 | 2 (≤) | 2 (=) | 16 (>) | 16/20 | lsa(E) | 16 | |||||||||||||
| Sequence Type | SCCmec | spa Type | Pulsotype | No. | Year (No.) | Host (No.) | Antimicrobial Resistance Genes * | |||||||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|---|
| mecA | blaZ | tet(L) | tet(M) | ant(4′)-Ia | ant(6)-Ia | aac(6′)-Ie-aph(2)-Ia | dfrA | dfrG | erm(A) | erm(B) | lnu(B) | lsa(E) | spw | fexA | ileS2 | |||||||
| 398 | V | t034 | B | 4 | 2017 (3) 2020 (1) | Dog (3) Cat (1) | ||||||||||||||||
| 398 | V | t034 | B | 2 | 2017 (2) | Dog (2) | ||||||||||||||||
| 398 | V | t4652 | B | 1 | 2017 (1) | Dog (1) | ||||||||||||||||
| 398 | V | t034 | C | 1 | 2017 (1) | Dog (1) | ||||||||||||||||
| 398 | V | t034 | A | 2 | 2018 (2) | Cat (2) | ||||||||||||||||
| 398 | V | t034 | A | 1 | 2018 (1) | Dog (1) | ||||||||||||||||
| 398 | V | t034 | A | 1 | 2019 (1) | Dog (1) | ||||||||||||||||
| 398 | V | t034 | A | 2 | 2019 (2) | Dog (2) | ||||||||||||||||
| 5926 | V | t1255 | A | 1 | 2018 (1) | Dog (1) | ||||||||||||||||
| 6563 | V | t1255 | A | 1 | 2020 (1) | Dog (1) | ||||||||||||||||
| 398 | Composite | t034 | A | 1 | 2020 (1) | Dog (1) | ||||||||||||||||
| 398 | Composite | - | A | 1 | 2019 (1) | Cat (1) | ||||||||||||||||
| 398 | Composite | - | A | 1 | 2018 (1) | Cat (1) | ||||||||||||||||
| 398 | Composite | - | A | 1 | 2019 (1) | Dog (1) | ||||||||||||||||
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chueahiran, S.; Yindee, J.; Boonkham, P.; Suanpairintr, N.; Chanchaithong, P. Methicillin-Resistant Staphylococcus aureus Clonal Complex 398 as a Major MRSA Lineage in Dogs and Cats in Thailand. Antibiotics 2021, 10, 243. https://doi.org/10.3390/antibiotics10030243
Chueahiran S, Yindee J, Boonkham P, Suanpairintr N, Chanchaithong P. Methicillin-Resistant Staphylococcus aureus Clonal Complex 398 as a Major MRSA Lineage in Dogs and Cats in Thailand. Antibiotics. 2021; 10(3):243. https://doi.org/10.3390/antibiotics10030243
Chicago/Turabian StyleChueahiran, Surawit, Jitrapa Yindee, Pongthai Boonkham, Nipattra Suanpairintr, and Pattrarat Chanchaithong. 2021. "Methicillin-Resistant Staphylococcus aureus Clonal Complex 398 as a Major MRSA Lineage in Dogs and Cats in Thailand" Antibiotics 10, no. 3: 243. https://doi.org/10.3390/antibiotics10030243