Comparative Transcriptomics Identify Key Hypothalamic Circular RNAs that Participate in Sheep (Ovis aries) Reproduction

 ,
, {kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animal Processing
2.2. Tissue Acquisition, RNA Extraction and Sequencing
2.3. Data Quality Control and Circular RNA Identification
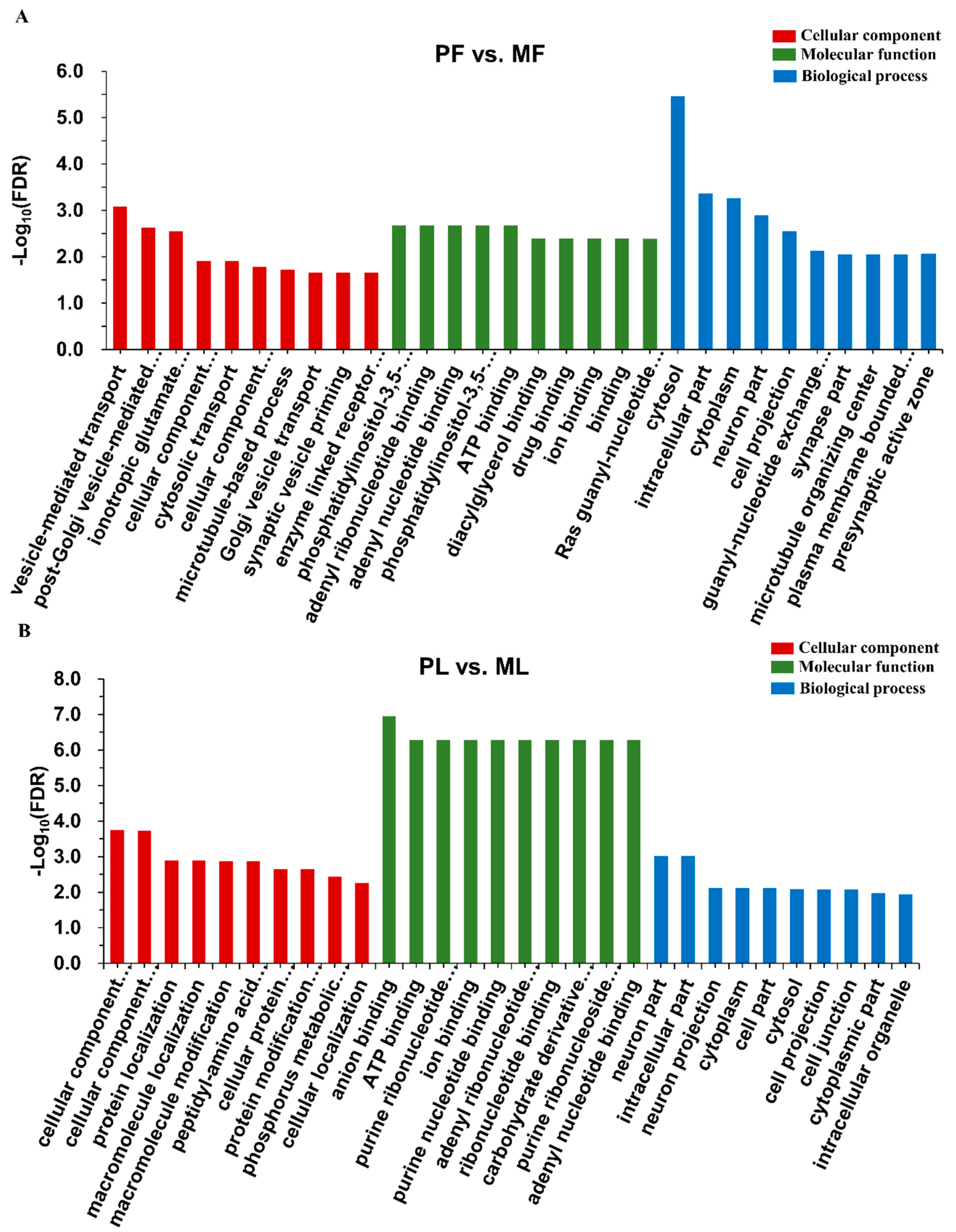
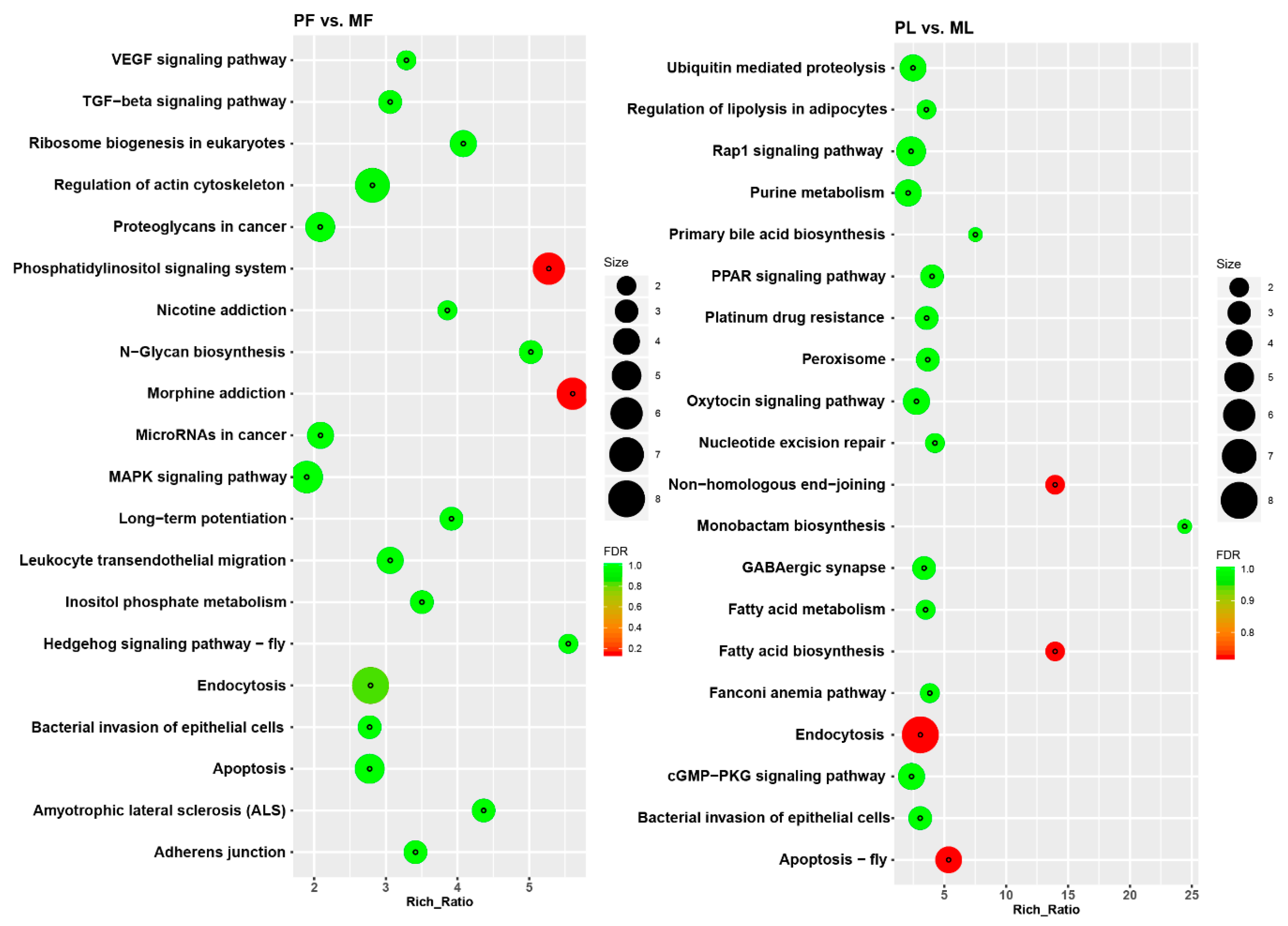
2.4. Analysis of Differentially Expressed circRNAs and Functional Enrichment Analysis
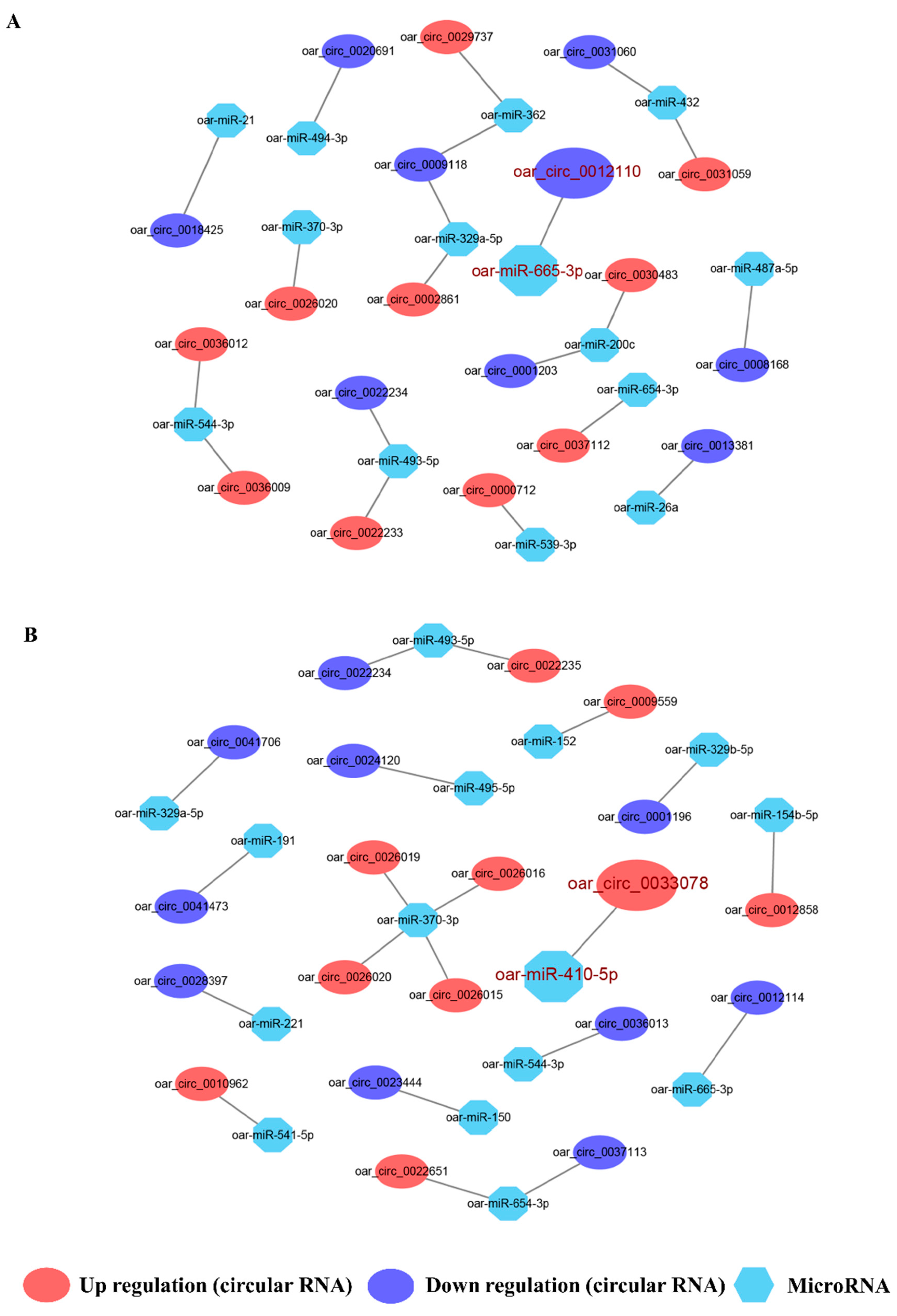
2.5. Integral DE circRNA–miRNA Network Analysis and ceRNA Construction
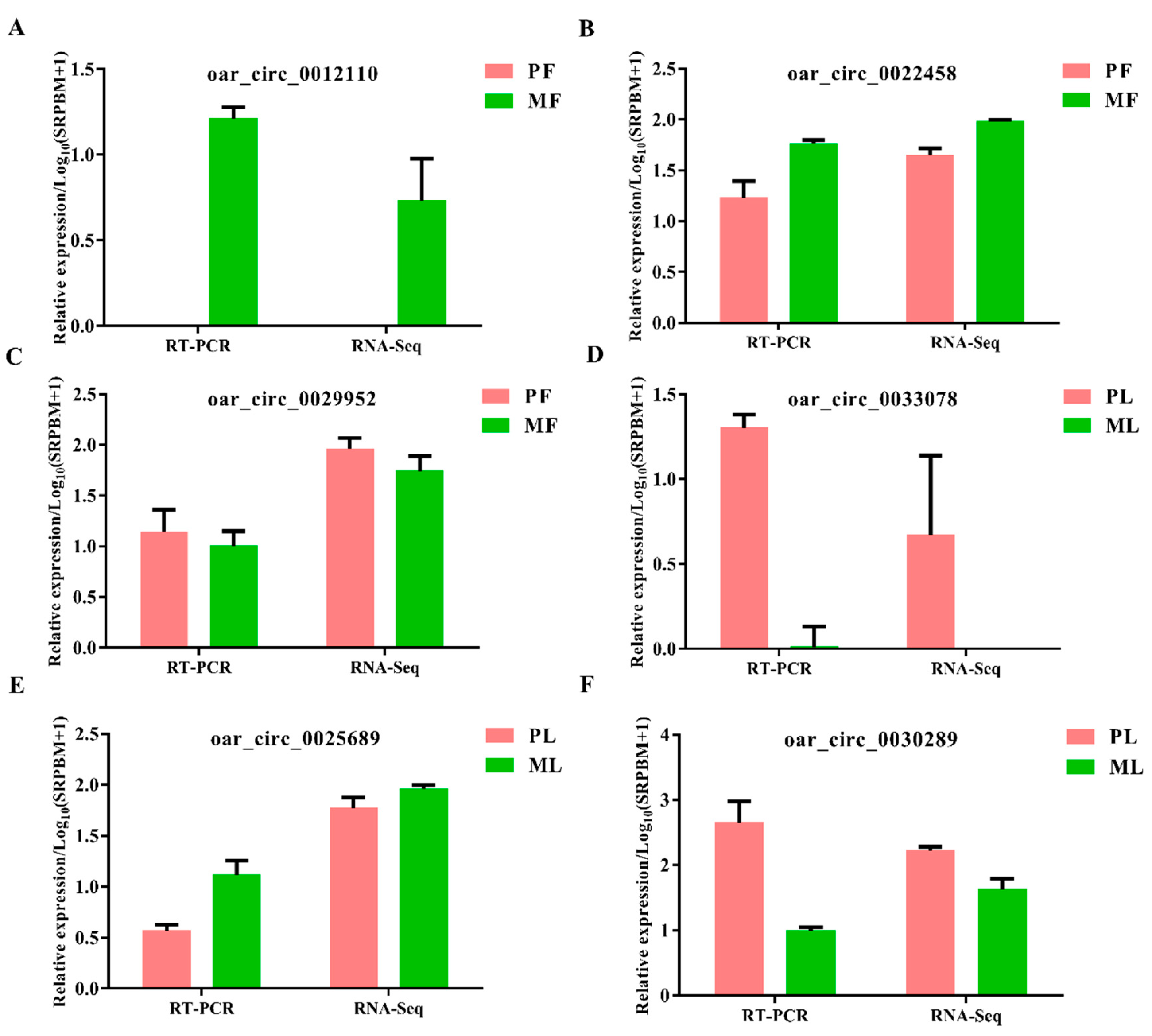
2.6. Data Validation
3. Results
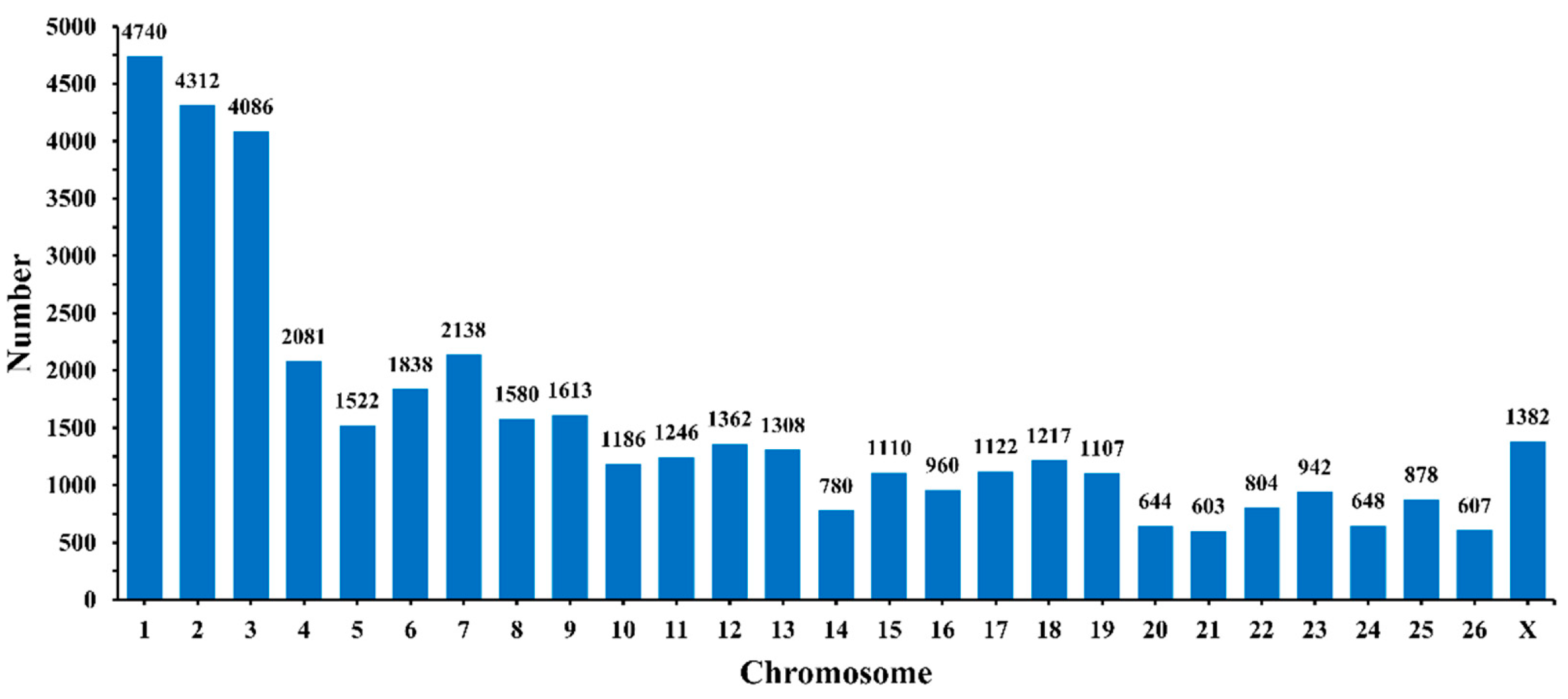
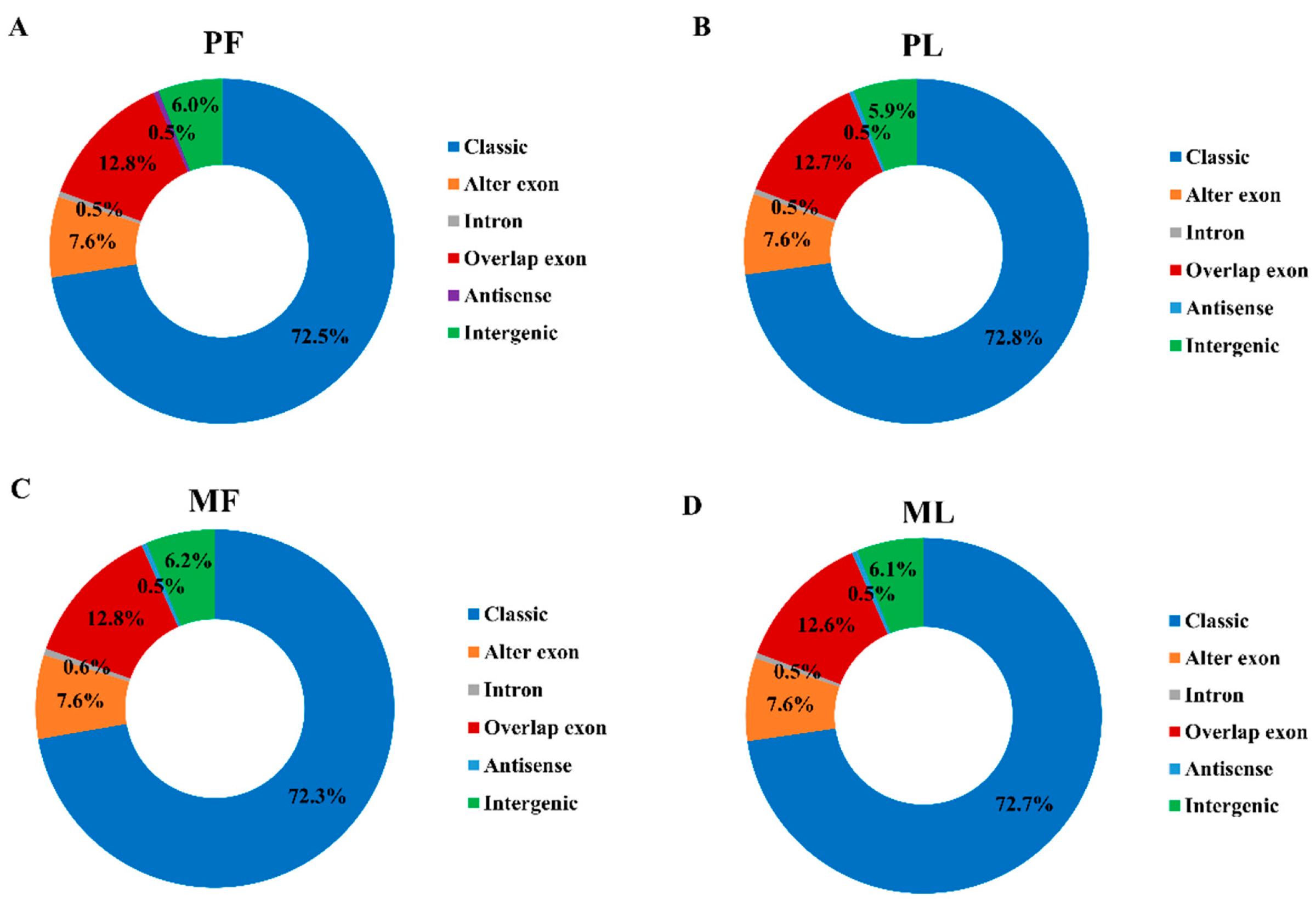
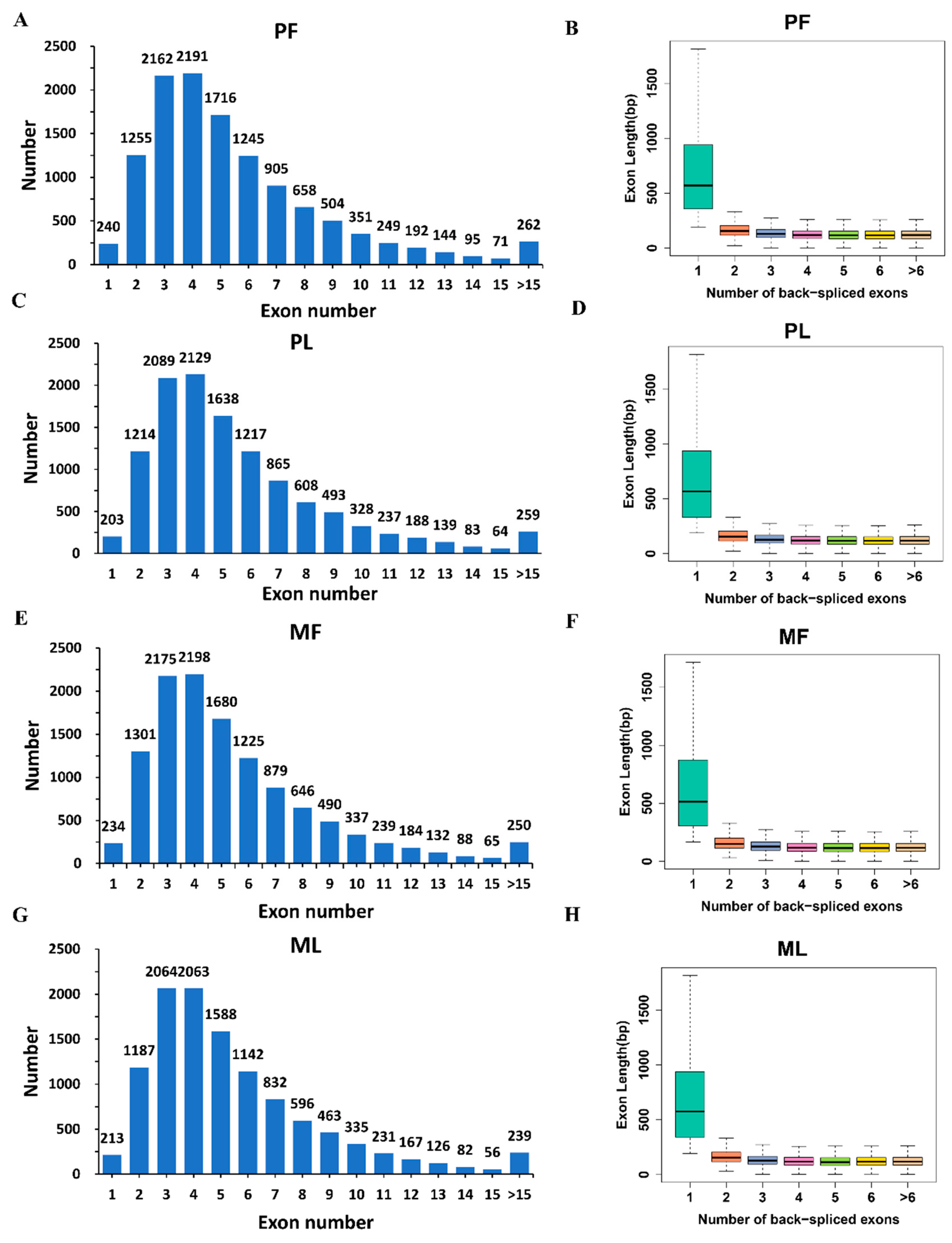
3.1. Circular RNA Expression Profiling
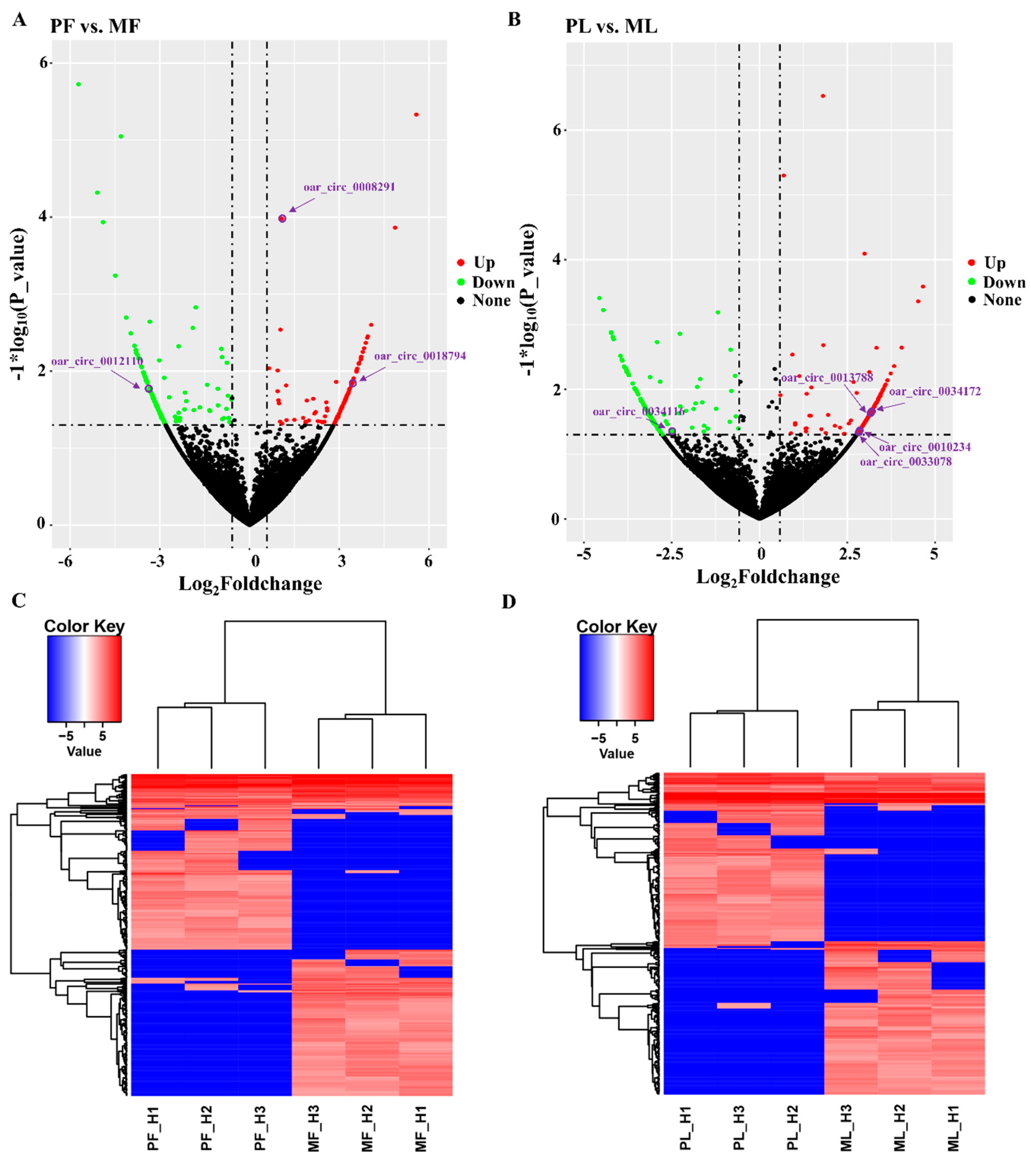
3.2. Identification of Differentially Expressed Circular RNAs (DE circRNAs) and Functional Enrichment Analysis
3.3. Integral circRNA-miRNA Pairs and Competing Endogenous RNAs Analysis
4. Discussion
4.1. Functional Enrichment Analysis of Key circRNAs
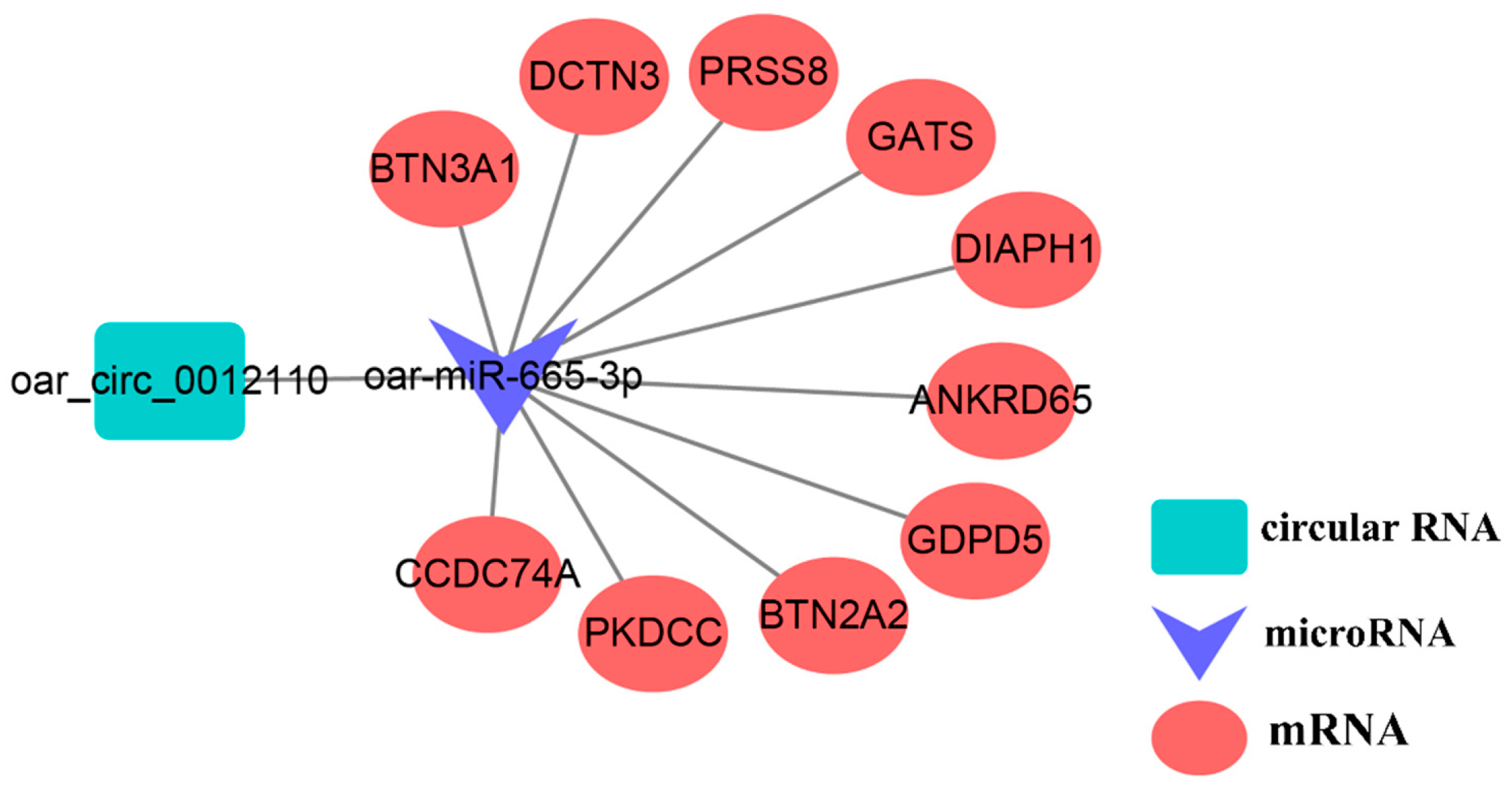
4.2. ceRNA Analysis Involving oar_circ_0012110
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
Data Accessibility
References
- Kaprara, A.; Huhtaniemi, I.T. The hypothalamus-pituitary-gonad axis: Tales of mice and men. Metab. Clin. Exp. 2018, 86, 3–17. [Google Scholar] [CrossRef] [PubMed]
- Marshall, J.C.; Kelch, R.P. Gonadotropin-releasing hormone: Role of pulsatile secretion in the regulation of reproduction. N. Engl. J. Med. 1986, 315, 1459–1468. [Google Scholar] [PubMed]
- Bliss, S.P.; Navratil, A.M.; Xie, J. Roberson MS, GnRH signaling, the gonadotrope and endocrine control of fertility. Front. Neuroendocrinol. 2010, 31, 322–340. [Google Scholar] [CrossRef] [PubMed]
- Haisenleder, D.J.; Dalkin, A.C.; Ortolano, G.A.; Marshall, J.C.; Shupnik, M.A. A pulsatile gonadotropin-releasing hormone stimulus is required to increase transcription of the gonadotropin subunit genes: Evidence for differential regulation of transcription by pulse frequency in vivo. Endocrinology 1991, 128, 509–517. [Google Scholar] [CrossRef] [PubMed]
- Mulsant, P.; Lecerf, F.; Fabre, S.; Schibler, L.; Monget, P.; Lanneluc, I.; Pisselet, C.; Riquet, J.; Monniaux, D.; Callebaut, I.; et al. Mutation in bone morphogenetic protein receptor-IB is associated with increased ovulation rate in Booroola Mérino ewes. Proc. Natl. Acad. Sci. USA 2001, 98, 5104–5109. [Google Scholar] [CrossRef] [PubMed]
- Polley, S.; De, S.; Brahma, B.; Mukherjee, A.; Vinesh, P.V.; Batabyal, S.; Arora, J.S.; Pan, S.; Samanta, A.K.; Datta, T.K.; et al. Polymorphism of BMPR1B, BMP15 and GDF9 fecundity genes in prolific Garole sheep. Trop. Anim. Health Prod. 2010, 42, 985–993. [Google Scholar] [CrossRef] [PubMed]
- Davis, G.H.; Balakrishnan, L.; Ross, I.K.; Wilson, T.; Galloway, S.M.; Lumsden, B.M.; Hanrahan, J.P.; Mullen, M.; Mao, X.Z.; Wang, G.L.; et al. Investigation of the Booroola (FecB) and Inverdale (FecXI) mutations in 21 prolific breeds and strains of sheep sampled in 13 countries. Anim. Reprod. Sci. 2006, 92, 87–96. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Pan, Z.; Wang, X.; Hu, W.; Di, R.; Yao, Y.; Chu, M. Progress on major genes for high fecundity in ewes. Front. Agric. Sci. Eng. 2014, 1, 282–290. [Google Scholar] [CrossRef] [Green Version]
- Li, X.; Li, C.; Wei, J.; Ni, W.; Xu, Y.; Yao, R.; Zhang, M.; Li, H.; Liu, L.; Dang, H.; et al. Comprehensive expression profiling analysis of pituitary indicates that circRNA participates in the regulation of sheep estrus. Genes 2019, 10, 90. [Google Scholar] [CrossRef]
- Li, X.; Li, C.; Ni, W.; Wang, D.; Hou, X.; Liu, Z.; Cao, Y.; Yao, Y.; Zhang, X.; Hu, S. Identification and comparison of microRNAs in pituitary gland during prenatal and postnatal stages of sheep by deep sequencing. J. Genet. 2018, 97, 965–975. [Google Scholar] [CrossRef]
- Cao, C.; Ding, Y.; Kong, X.; Feng, G.; Xiang, W.; Chen, L.; Yang, F.; Zhang, K.; Chu, M.; Wang, P.; et al. Reproductive role of miRNA in the hypothalamic-pituitary axis. Mol. Cell. Neurosci. 2018, 88, 130–137. [Google Scholar] [CrossRef] [PubMed]
- Zheng, J.; Wang, Z.; Yang, H.; Yao, X.; Yang, P.; Ren, C.; Wang, F.; Zhang, Y. Pituitary transcriptomic study reveals the differential regulation of lncRNAs and mRNAs related to prolificacy in different FecB genotyping sheep. Genes 2019, 10, 157. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Yang, L.; Chen, L.L. The biogenesis, functions, and challenges of circular RNAs. Mol. Cell 2018, 71, 428–442. [Google Scholar] [CrossRef] [PubMed]
- Zhang, Z.; Tang, J.; Di, R.; Liu, Q.; Wang, X.; Gan, S.; Zhang, X.; Zhang, J.; Hu, W.; Chu, M. Comparative transcriptomics reveal key sheep (Ovis aries) hypothalamus lncRNAs that affect reproduction. Animals 2019, 9, 152. [Google Scholar] [CrossRef] [PubMed]
- Messina, A.; Langlet, F.; Chachlaki, K.; Roa, J.; Rasika, S.; Jouy, N.; Gallet, S.; Gaytan, F.; Parkash, J.; Tena-Sempere, M.; et al. A microRNA switch regulates the rise in hypothalamic GnRH production before puberty. Nat. Neurosci. 2016, 19, 835–844. [Google Scholar] [CrossRef] [PubMed]
- Liu, Q.; Hu, W.; He, X.; Pan, Z.; Guo, X.; Feng, T.; Cao, G.; Huang, D.; He, J.; Di, R.; et al. Establishment of high-throughput molecular detection methods for ovine high fecundity major gene and their application. Acta Veteinaria Zootech. Sin. 2017, 48, 39–51. [Google Scholar]
- Gao, Y.; Wang, J.; Zhao, F. CIRI: An efficient and unbiased algorithm for de novo circular RNA identification. Genome Biol. 2015, 16, 4. [Google Scholar] [CrossRef]
- Li, Y.; Zheng, Q.; Bao, C.; Li, S.; Guo, W.; Zhao, J.; Chen, D.; Gu, J.; He, X.; Huang, S. Circular RNA is enriched and stable in exosomes: A promising biomarker for cancer diagnosis. Cell Res. 2015, 25, 981–984. [Google Scholar] [CrossRef]
- Wang, L.; Feng, Z.; Wang, X.; Wang, X.; Zhang, X. DEGseq: An R package for identifying differentially expressed genes from RNA-seq data. Bioinformatics 2010, 26, 136–138. [Google Scholar] [CrossRef]
- Hansen, T.B.; Jensen, T.I.; Clausen, B.H.; Bramsen, J.B.; Finsen, B.; Damgaard, C.K.; Kjems, J. Natural RNA circles function as efficient microRNA sponges. Nature 2013, 495, 384–388. [Google Scholar] [CrossRef]
- Memczak, S.; Jens, M.; Elefsinioti, A.; Torti, F.; Krueger, J.; Rybak, A.; Maier, L.; Mackowiak, S.D.; Gregersen, L.H.; Munschauer, M.; et al. Circular RNAs are a large class of animal RNAs with regulatory potency. Nature 2013, 495, 333–338. [Google Scholar] [CrossRef] [PubMed]
- Shannon, P.; Markiel, A.; Ozier, O.; Baliga, N.S.; Wang, J.T.; Ramage, D.; Amin, N.; Schwikowski, B.; Ideker, T. Cytoscape: A software environment for integrated models of biomolecular interaction networks. Genome Res. 2003, 13, 2498–2504. [Google Scholar] [CrossRef] [PubMed]
- Nestor, C.C.; Bedenbaugh, M.N.; Hileman, S.M.; Coolen, L.M.; Lehman, M.N.; Goodman, R.L. Regulation of GnRH pulsatility in sheep. Reproduction 2018, 3, 83–99. [Google Scholar] [CrossRef] [PubMed]
- Navarro, V.M.; Kaiser, U.B. Metabolic influences on neuroendocrine regulation of reproduction. Curr. Opin. Endocrinol. Diabetes Obes. 2013, 20, 335–341. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hill, J.W.; Elias, C.F. Neuroanatomical framework of the metabolic control of reproduction. Physiol. Rev. 2018, 98, 2349–2380. [Google Scholar] [CrossRef] [PubMed]
- Harter, C.J.L.; Kavanagh, G.S.; Smith, J.T. The role of kisspeptin neurons in reproduction and metabolism. J. Endocrinol. 2018, 238, R173–R183. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Plant, T.M. Neuroendocrine control of the onset of puberty. Front. Neuroendocrinol. 2015, 38, 73–88. [Google Scholar] [CrossRef] [Green Version]
- Rybak-Wolf, A.; Stottmeister, C.; Glažar, P.; Jens, M.; Pino, N.; Giusti, S.; Hanan, M.; Behm, M.; Bartok, O.; Ashwal-Fluss, R.; et al. Circular RNAs in the mammalian brain are highly abundant, conserved, and dynamically expressed. Mol. Cell 2015, 58, 870–885. [Google Scholar] [CrossRef] [PubMed]
- Cao, Y.; You, S.; Yao, Y.; Liu, Z.J.; Hazi, W.; Li, C.Y.; Zhang, X.Y.; Hou, X.X.; Wei, J.C.; Li, X.Y.; et al. Expression profiles of circular RNAs in sheep skeletal muscle. Asian Australas. J. Anim. Sci. 2018, 31, 1550–1557. [Google Scholar] [CrossRef]
- Ebbesen, K.K.; Kjems, J.; Hansen, T.B. Circular RNAs: Identification, biogenesis and function. Biochim. Biophys. Acta 2016, 1859, 163–168. [Google Scholar] [CrossRef]
- Luo, S.; Kleemann, G.A.; Ashraf, J.M.; Shaw, W.M.; Murphy, C.T. TGF-β and insulin signaling regulate reproductive aging via oocyte and germline quality maintenance. Cell 2010, 143, 299–312. [Google Scholar] [CrossRef] [PubMed]
- Yang, L.; Du, X.; Liu, L.; Cao, Q.; Pan, Z.; Li, Q. miR-1306 mediates the feedback regulation of the TGF-β/SMAD signaling pathway in granulosa cells. Cells 2019, 8, 298. [Google Scholar] [CrossRef] [PubMed]
- Nakajima, T.; Hata, R.; Kunieda, Y.; Kondo, T. Distribution of Smad mRNA and proteins in the rat brain. J. Chem. Neuroanat. 2018, 90, 11–39. [Google Scholar] [CrossRef] [PubMed]
- Ueberham, U.; Ueberham, E.; Gruschka, H.; Arendt, T. Altered subcellular location of phosphorylated Smads in Alzheimer’s disease. Eur. J. Neurosci. 2010, 24, 2327–2334. [Google Scholar] [CrossRef] [PubMed]
- Ueberham, U.; Lange, P.; Ueberham, E.; Brückner, M.K.; Hartlage-Rübsamen, M.; Pannicke, T.; Rohn, S.; Cross, M.; Arendt, T. Smad2 isoforms are differentially expressed during mouse brain development and aging. Int. J. Dev. Neurosci. 2009, 27, 501–510. [Google Scholar] [CrossRef] [PubMed]
- Bouret, S.; De Seranno, S.; Beauvillain, J.C.; Prevot, V. Transforming growth factor beta1 may directly influence gonadotropin-releasing hormone gene expression in the rat hypothalamus. Endocrinology 2004, 145, 1794–1801. [Google Scholar] [CrossRef] [PubMed]
- Nguyen, T.M.D. Role of AMPK in mammal’s reproduction: Specific controls and whole-body energy sensing. Comptes Rendus Biol. 2019, 342, 1–6. [Google Scholar] [CrossRef]
- Smyth, C.E.; Knee, R.; Wilkinson, M.; Murphy, P.R. Decline in basic fibroblast growth factor (FGF-2) mRNA expression in female rat hypothalamus at puberty. J. Neuroendocrinol. 2010, 9, 151–159. [Google Scholar] [CrossRef]
- Yoshimura, K.; Kaji, H.; Kamidono, S.; Chihara, K. Fasting increases the expression of basic fibroblast growth factor (FGF-2) messenger ribonucleic acid in rat hypothalamus. Horm. Metab. Res. 1995, 27, 363–366. [Google Scholar] [CrossRef]
- Akhter, N.; CarlLee, T.; Syed, M.M.; Odle, A.K.; Cozart, M.A.; Haney, A.C.; Allensworth-James, M.L.; Beneš, H.; Childs, G.V. Selective deletion of leptin receptors in gonadotropes reveals activin and GnRH-binding sites as leptin targets in support of fertility. Endocrinology 2014, 155, 4027–4042. [Google Scholar] [CrossRef]
- Mani, I.; Pandey, K.N. Emerging concepts of receptor endocytosis and concurrent intracellular signaling: Mechanisms of guanylyl cyclase/natriuretic peptide receptor—A activation and trafficking. Cell. Signal. 2019, 60, 17–30. [Google Scholar] [CrossRef] [PubMed]
- Chaudhari, A.; Håversen, L.; Mobini, R.; Andersson, L.; Ståhlman, M.; Lu, E.; Rutberg, M.; Fogelstrand, P.; Ekroos, K.; Mardinoglu, A.; et al. ARAP2 promotes GLUT1-mediated basal glucose uptake through regulation of sphingolipid metabolism. Biochim. Biophys. Acta 2016, 1861, 1643–1651. [Google Scholar] [CrossRef] [PubMed]
- Pal, L.; Chu, H.P.; Shu, J.; Topalli, I.; Santoro, N.; Karkanias, G. In vitro evidence of glucose-induced toxicity in GnRH secreting neurons: High glucose concentrations influence GnRH secretion, impair cell viability, and induce apoptosis in the GT1-1 neuronal cell line. Fertil. Steril. 2007, 88, 1143–1149. [Google Scholar] [CrossRef] [PubMed]
- Sukocheva, O.; Wadham, C.; Xia, P. Role of sphingolipids in the cytoplasmic signaling of estrogens. Steroids 2009, 74, 562–567. [Google Scholar] [CrossRef] [PubMed]
- Das, S.; Maitra, U. Functional significance and mechanism of eIF5-promoted GTP hydrolysis in eukaryotic translation initiation. Prog. Nucleic Acid Res. Mol. Biol. 2001, 70, 207–231. [Google Scholar]
- Asano, K.; Shalev, A.; Phan, L.; Nielsen, K.; Clayton, J.; Valásek, L.; Donahue, T.F.; Hinnebusch, A.G. Multiple roles for the C-terminal domain of eIF5 in translation initiation complex assembly and GTPase activation. EMBO J. 2014, 20, 2326–2337. [Google Scholar] [CrossRef] [PubMed]
- Saini, A.K.; Nanda, J.S.; Martin-Marcos, P.; Dong, J.; Zhang, F.; Bhardwaj, M.; Lorsch, J.R.; Hinnebusch, A.G. Eukaryotic translation initiation factor eIF5 promotes the accuracy of start codon recognition by regulating Pi release and conformational transitions of the preinitiation complex. Nucleic Acids Res. 2014, 42, 9623–9640. [Google Scholar] [CrossRef] [Green Version]
- Jazayeri, A.; Falck, J.; Lukas, C.; Bartek, J.; Smith, G.C.; Lukas, J.; Jackson, S.P. ATM- and cell cycle-dependent regulation of ATR in response to DNA double-strand breaks. Nat. Cell Biol. 2006, 8, 37–45. [Google Scholar] [CrossRef]
- Falck, J.; Coates, J.; Jackson, S.P. Conserved modes of recruitment of ATM, ATR and DNA-PKcs to sites of DNA damage. Nature 2005, 434, 605–611. [Google Scholar] [CrossRef]
- De Klein, A.; Muijtjens, M.; van Os, R.; Verhoeven, Y.; Smit, B.; Carr, A.M.; Lehmann, A.R.; Hoeijmakers, J.H. Targeted disruption of the cell-cycle checkpoint gene ATR leads to early embryonic lethality in mice. Curr. Biol. 2000, 10, 479–482. [Google Scholar] [CrossRef] [Green Version]
- Zhong, Y.; Du, Y.; Yang, X.; Mo, Y.; Fan, C.; Xiong, F.; Ren, D.; Ye, X.; Li, C.; Wang, Y. Circular RNAs function as ceRNAs to regulate and control human cancer progression. Mol. Cancer 2018, 17, 79. [Google Scholar] [CrossRef] [PubMed]
- Li, X.; Ao, J.; Wu, J. Systematic identification and comparison of expressed profiles of lncRNAs and circRNAs with associated co-expression and ceRNA networks in mouse germline stem cells. Oncotarget 2017, 8, 26573–26590. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Li, D.; Sewer, M. RhoA and DIAPH1 mediate adrenocorticotropin-stimulated cortisol biosynthesis by regulating mitochondrial trafficking. Endocrinology 2010, 151, 4313–4323. [Google Scholar] [CrossRef] [PubMed]
- Oakley, A.E.; Breen, K.M.; Clarke, I.J.; Karsch, F.J.; Wagenmaker, E.R.; Tilbrook, A.J. Cortisol reduces gonadotropin-releasing hormone pulse frequency in follicular phase ewes: Influence of ovarian steroids. Endocrinology 2009, 150, 341–349. [Google Scholar] [CrossRef] [PubMed]









© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Zhang, Z.; Tang, J.; He, X.; Zhu, M.; Gan, S.; Guo, X.; Zhang, X.; Zhang, J.; Hu, W.; Chu, M. Comparative Transcriptomics Identify Key Hypothalamic Circular RNAs that Participate in Sheep (Ovis aries) Reproduction. Animals 2019, 9, 557. https://doi.org/10.3390/ani9080557
Zhang Z, Tang J, He X, Zhu M, Gan S, Guo X, Zhang X, Zhang J, Hu W, Chu M. Comparative Transcriptomics Identify Key Hypothalamic Circular RNAs that Participate in Sheep (Ovis aries) Reproduction. Animals. 2019; 9(8):557. https://doi.org/10.3390/ani9080557
Chicago/Turabian StyleZhang, Zhuangbiao, Jishun Tang, Xiaoyun He, Mingxia Zhu, Shangquan Gan, Xiaofei Guo, Xiaosheng Zhang, Jinlong Zhang, Wenping Hu, and Mingxing Chu. 2019. "Comparative Transcriptomics Identify Key Hypothalamic Circular RNAs that Participate in Sheep (Ovis aries) Reproduction" Animals 9, no. 8: 557. https://doi.org/10.3390/ani9080557