Effects of Regulating Visitor Viewing Proximity and the Intensity of Visitor Behaviour on Little Penguin (Eudyptula minor) Behaviour and Welfare

 , , and
, , and
Abstract
:Simple Summary
Abstract
1. Introduction
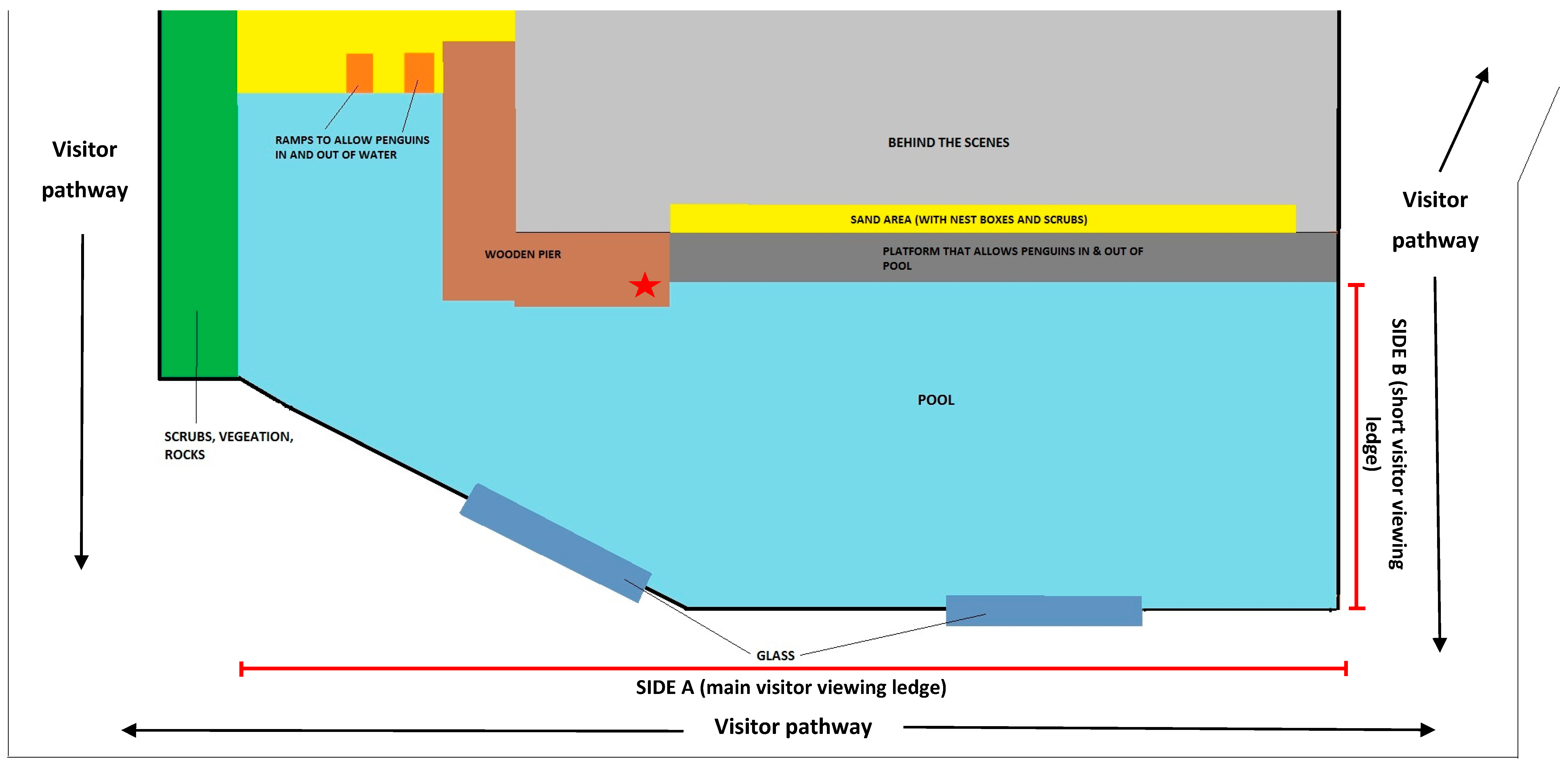
2. Materials and Methods
2.1. Design & Treatments
- Viewing proximity of visitors to enclosure at two levels:
- a.
- ‘Increased viewing distance’—a barrier was set up 2 m from the enclosure to increase the distance between visitors and the enclosure (Figure 2). This allowed unrestricted viewing of the enclosure but was a strong impediment to visitors physically interacting with the glass windows of the pool, pool water and other enclosure features.
- b.
- ‘Normal viewing distance’—no barrier was in place and visitors could approach to the edge of the pool (i.e., visitors could approach within 2 m of the enclosure).
- Intensity of visitor behaviour at two levels:
- a.
- ‘Unregulated visitor behaviour’—visitor behaviours were uncontrolled.
- b.
- ‘Regulated visitor behaviour’—the objective of this treatment was to attempt to reduce the intensity of visitor behaviour using signs requesting visitors to be quiet, move slowly in the exhibit area and avoid physically interacting with the penguins (Figure 3). Also, for this treatment, the researcher was dressed in zoo uniform. This procedure has been successfully used in research on meerkats to reduce the intensity of visitor behaviours [27].
- Normal viewing distance and unregulated visitor behaviour—no barrier in place and visitor behaviour was uncontrolled.
- Increased viewing distance and unregulated visitor behaviour—barrier was in place and visitor behaviour was uncontrolled.
- Normal viewing distance and regulated visitor behaviour—no barrier in place and visitor behaviour was attempted to be controlled.
- Increased viewing distance and regulated visitor behaviour—barrier was in place and visitor behaviour was attempted to be controlled.
- Closed exhibit—penguin exhibit closed to the public.
2.2. Animal Behavioural Observations
2.3. Penguin Faecal Sampling and Analysis: Faecal Glucocorticoid Metabolites (FGM)
2.4. Visitor Behavioural Observations
2.5. Data Analysis
3. Results
3.1. Visitor Variables
3.2. Animal Behaviour
3.2.1. Behavioural States
3.2.2. Behavioural Events
3.2.3. Little Penguin FGM Concentrations
4. Discussion
5. Conclusions
Author Contributions
Funding
Acknowledgments
Conflicts of Interest
References
- Hosey, G. Hediger revisited: How do zoo animals see us? J. Appl. Anim. Welf. Sci. 2013, 16, 338–359. [Google Scholar] [CrossRef] [PubMed]
- Fernandez, E.; Tamborski, M.; Pickens, S.; Timberlake, W. Animal-visitor interactions in the modern zoo: Conflicts and interventions. Appl. Anim. Behav. Sci. 2009, 120, 1–8. [Google Scholar] [CrossRef]
- Giese, M. Effects of human activity on Adelie penguin Phygoscelis adeliae breeding success. Biol. Conserv. 1996, 75, 157–164. [Google Scholar] [CrossRef]
- McClung, M.R.; Seddon, P.J.; Massaro, M.; Setiawan, A.N. Nature-based tourism impacts on Yellow-eyed penguins Megadyptes antipodes: Does unregulated visitor access affect fledging weight and juvenile survival? Biol. Conserv. 2004, 119, 279–285. [Google Scholar] [CrossRef]
- Ellenberg, U.; Setiawan, A.N.; Cree, A.; Houston, D.M.; Seddon, P.J. Elevated hormonal stress response and reduced reproductive output in Yellow-eyed penguins exposed to unregulated tourism. Gen. Comp. Endocrinol. 2007, 152, 54–63. [Google Scholar] [CrossRef]
- Ellenberg, U.; Mattern, T.; Seddon, P.J.; Jorquera, G.L. Physiological and reproductive consequences of human disturbance in Humboldt penguins: The need for species-specific visitor management. Biol. Conserv. 2006, 133, 95–106. [Google Scholar] [CrossRef]
- Holmes, N.D.; Giese, M.; Achurch, H.; Robinson, S.; Kriwoken, L.K. Behaviour and breeding success of Gentoo penguins Pygoscelis papua in areas of low and high human activity. Polar Biol. 2006, 29, 399–412. [Google Scholar] [CrossRef]
- Holmes, N.D. Comparing King, Gentoo, and Royal penguin responses to pedestrian visitation. J. Wildl. Manag. 2007, 71, 2575–2582. [Google Scholar] [CrossRef]
- Giling, D.; Reina, R.D.; Hogg, Z. Anthropogenic influence on an urban colony of the little penguin Eudyptula minor. Mar. Freshw. Res. 2008, 59, 647–651. [Google Scholar] [CrossRef]
- Fowler, G.S. Behavioral and hormonal responses of Magellanic penguins (Spheniscus magellanicus) to tourism and nest site visitation. Biol. Conserv. 1999, 90, 143–149. [Google Scholar] [CrossRef]
- Villanueva, C.; Walker, B.G.; Bertellotti, M. A matter of history: Effects of tourism on physiology, behaviour and breeding parameters in Magellanic penguins (Spheniscus magellanicus) at two colonies in Argentina. J. Ornithol. 2012, 153, 219–228. [Google Scholar] [CrossRef]
- Yorio, P.; Boersma, P.D. The effects of human disturbance on Magellanic penguin Spheniscus magellanicus behaviour and breeding success. Bird Conserv. Int. 1990, 2, 161–173. [Google Scholar] [CrossRef]
- Ozella, L.; Favaro, L.; Carnovale, I.; Pessani, D. Pond use by captive African penguins (Spheniscus demersus) in an immersive exhibit adjacent to human bathers. J. Appl. Anim. Welf. Sci. 2014, 1–7. [Google Scholar] [CrossRef]
- Sherwen, S.L.; Magrath, M.J.L.; Butler, K.L.; Hemsworth, P.H. Little penguins, Eudyptula minor, show increased avoidance, aggression and vigilance in response to zoo visitors. Appl. Anim. Behav. Sci. 2015, 168, 71–76. [Google Scholar] [CrossRef]
- Condon, E.; Wehnelt, S.; Turner, Z. The effect of visitors on the behavior of Humboldt’s penguins at Chester zoo. Fed. Res. Newsl. 2003, 4, 3. [Google Scholar]
- Collins, C.K.; Quirke, T.; Overy, L.; Flannery, K.; O’Riordan, R. The effect of the zoo setting on the behavioural diversity of captive Gentoo penguins and the implications for their educational potential. J. Zoo Aquar. Res. 2016, 4, 85–90. [Google Scholar]
- Learmonth, M.; Sherwen, S.L.; Hemsworth, P.H. The effects of zoo visitors on Quokka (Setonix brachyurus) avoidance behavior in a walk-through exhibit. Zoo Biol. 2018, 37, 223–228. [Google Scholar] [CrossRef]
- Sherwen, S.L.; Magrath, M.J.L.; Butler, K.L.; Fanson, K.V.; Hemsworth, P.H. Effects of visual contact with zoo visitors on Black-capped capuchin welfare. Appl. Anim. Behav. Sci. 2015, 167, 65–73. [Google Scholar] [CrossRef]
- Ainley, D.G. The comfort behaviour of Adélie and other penguins. Behaviour 1974, 50, 16–51. [Google Scholar] [CrossRef]
- Ainley, D.G. Flocking in Adélie penguins. Int. J. Avian Sci. 1972, 114, 388–390. [Google Scholar] [CrossRef]
- Eggleton, P.; Siegfried, W.R. Displays of the Jackass penguin. J. Afr. Ornithol. 1979, 50, 139–167. [Google Scholar] [CrossRef]
- Hughes, K.A.; Waluda, C.M.; Stone, R.E.; Ridout, M.S.; Shears, J.R. Short-term responses of King penguins Aptenodytes patagonicus to helicopter disturbance at South Georgia. Polar Biol. 2008, 31, 1521–1530. [Google Scholar] [CrossRef]
- Black, C.; Collen, B.; Johnston, D.; Hart, T. Why huddle? Ecological drivers of chick aggregations in Gentoo penguins, Pygoscelis papua, across latitudes. PLoS ONE 2016, 11, e0145676. [Google Scholar] [CrossRef] [PubMed]
- Moberg, G.P. Biological response to stress: Implications for animal welfare. In The Biology of Animal Stress; Moberg, G.P., Mench, J.A., Eds.; CABI Publishing: Wallingford, UK, 2000; pp. 123–146. [Google Scholar]
- Sapolsky, R.M. Endocrinology of the stress-response. In Behavioral Endocrinology; Becker, J.B., Breedlove, S.M., Crews, D., McCarthy, M.M., Eds.; MIT Press: Cambridge, MA, USA, 2002; pp. 409–450. [Google Scholar]
- Shepherdson, D.J.; Carlstead, K.C.; Wielebnowski, N. Cross-institutional assessment of stress responses in zoo animals using longitudinal monitoring of faecal corticoids and behaviour. Anim. Welf. 2004, 13, S105–S113. [Google Scholar]
- Sherwen, S.L.; Magrath, M.J.L.; Butler, K.L.; Phillips, C.J.C.; Hemsworth, P.H. A multi-enclosure study investigating the behavioural response of meerkats to zoo visitors. Appl. Anim. Behav. Sci. 2014, 156, 70–77. [Google Scholar] [CrossRef]
- Martin, P.; Bateson, P. Measuring Behaviour: An Introductory Guide; Cambridge University Press: Cambridge, UK, 2007. [Google Scholar]
- Sherwen, S.L.; Fanson, K.V. Validation of a faecal glucocorticoid assay in little penguins (Eudyptula minor). J. Zoo Aquar. Res. 2015, 3, 134–136. [Google Scholar]
- Nakagawa, S.; Möstl, E.; Waas, J.R. Validation of an enzyme immunoassay to measure faecal glucocorticoid metabolites from Adélie penguins (Pygoscelis adeliae): A non-invasive tool for estimating stress? Polar Biol. 2003, 26, 491–493. [Google Scholar]
- Ninnes, C.; Waas, J.; Ling, N.; Nakagawa, S.; Banks, J.; Bell, D.; Bright, A.; Carey, P.; Chandler, J.; Hudson, Q.; et al. Comparing plasma and faecal measures of steroid hormones in Adelie penguins Pygoscelis adeliae. J. Comp. Physiol. Biochem. Syst. Environ. Physiol. 2010, 180, 83–94. [Google Scholar] [CrossRef]
- Park, L.O.; Manning, R.E.; Marion, J.L.; Lawson, S.R.; Jacobi, C. Managing visitor impacts in parks: A multi-method study of the effectiveness of alternative management practices. J. Park Recreat. Adm. 2008, 26, 97–121. [Google Scholar]
- Acevedo-Gutiérrez, A.; Acevedo, L.; Belonovich, O.; Boren, L. How effective are posted signs to regulate tourism? An example with New Zealand Fur seals. Tour. Mar. Environ. 2011, 7, 39–41. [Google Scholar] [CrossRef]
- Kratochvil, H.; Schwammer, H. Reducing acoustic disturbances by aquarium visitors. Zoo Biol. 1997, 16, 349–353. [Google Scholar] [CrossRef]
- Wright, M. Ecotourism on Otago Peninsula: Preliminary Studies of Hoihos (Megadyptes antipodes) and Hookers Sealion (Photcartos hookerii); Department of Conservation: Wellington, New Zealand, 1998; pp. 5–36. [Google Scholar]
- Wilson, R.; Culik, B.; Danfeld, R.; Adelung, D. People in Antarctica—how much do Adélie penguins Pygoscelis adeliae care? Polar Biol. 1991, 11, 363–370. [Google Scholar] [CrossRef]
- Nimon, A.J.; Schroter, R.C.; Stonehouse, B. Heart rate of disturbed penguins. Nature 1995, 374, 415. [Google Scholar] [CrossRef]
- Giese, M. Guidelines for people approaching breeding groups of Adélie penguins (Pygoscelis adeliae). Polar Rec. 1998, 34, 287–292. [Google Scholar] [CrossRef]
- Klomp, N.I.; Meathrel, C.E.; Wienecke, B.C.; Wooller, R.D. Surface nesting by little penguins on Penguin Island, Western Australia. Emu 1991, 91, 190–193. [Google Scholar] [CrossRef]
- Weerheim, M.S.; Klomp, N.I.; Brunsting, A.M.H.; Komdeur, J. Population size, breeding habitat and nest site distribution of little penguins (Eudyptula minor) on Montague Island, New South Wales. Wildl. Res. 2003, 30, 151–157. [Google Scholar] [CrossRef]
- Wallis, R.; King, K.; Wallis, A. The little penguin ‘Eudyptula minor’ on Middle Island, Warrnambool, Victoria: An update on population size and predator management. Vic. Nat. 2017, 134, 48–51. [Google Scholar]
- Dann, P.; Cullen, J.M.; Thoday, R.; Jesso, P.R. Movements and patterns of mortality at sea of little penguins Eudyptula minor from Phillip Island, Victoria. Emu 1991, 91, 278–286. [Google Scholar] [CrossRef]
- Stahel, C.; Gales, R. Little Penguin: Fairy Penguins in Australia; New South Wales University Press: Kensington, Australia, 1987. [Google Scholar]
- Dann, P. Distribution, population trends and factors influencing the population size of little penguins Eudyptula minor on Phillip Island, Victoria. Emu 1992, 91, 263–272. [Google Scholar] [CrossRef]
- Wiebkin, A.S. Conservation Management Priorities for Little Penguin Populations in Gulf St Vincent. Report to Adelaide and Mount Lofty Ranges Natural Resources Management Board. SARDI Research Report Series No. 588; South Australian Research and Development Institute (Aquatic Sciences): Adelaide, Australia, 2011. [Google Scholar]
- Preston, T. Water rats as predators of little penguins. Vic. Nat. 2008, 125, 165–168. [Google Scholar]
- Bool, N.; Page, B.; Goldsworthy, S.D. What Is Causing the Decline of Little Penguins (Eudyptula minor) on Granite Island, South Australia? South Australian Research and Development Institute (Aquatic Sciences): Adelaide, Australia, 2007. [Google Scholar]
- Mckinney, F. The comfort movements of Anatidae. Behaviour 1965, 25, 120–220. [Google Scholar] [CrossRef]
- Delius, J.D. Preening and associated comfort behavior in birds. Ann. N. Y. Acad. Sci. 1988, 525, 40–55. [Google Scholar] [CrossRef]
- Zimmerman, P.H.; Buijs, S.A.F.; Bolhuis, J.E.; Keeling, L.J. Behaviour of domestic fowl in anticipation of positive and negative stimuli. Anim. Behav. 2011, 81, 569–577. [Google Scholar] [CrossRef]
- Dawkins, M.S. Behavioural deprivation: A central problem in animal welfare. Appl. Anim. Behav. Sci. 1988, 20, 209–225. [Google Scholar] [CrossRef] [Green Version]
- Jensen, P.; Toates, F.M. Who needs ‘behavioural needs’? Motivational aspects of the needs of animals. Appl. Anim. Behav. Sci. 1993, 37, 161–181. [Google Scholar] [CrossRef]
- Dawkins, M.S. Using behaviour to assess animal welfare. Anim. Welf. 2004, 13, S3–S7. [Google Scholar]
- Camprasse, E.; Sutton, G.; Berlincourt, M.; Arnould, J. Changing with the times: Little penguins exhibit flexibility in foraging behaviour and low behavioural consistency. Mar. Biol. 2017, 164, 1–10. [Google Scholar] [CrossRef]
- Seligman, M.E.; Meyer, B. Chronic fear and ulcers in rats as a function of the unpredictability of safety. J. Comp. Physiol. Psychol. 1970, 73, 202–207. [Google Scholar] [CrossRef]
- Weiss, J.M. Somatic effects of predictable and unpredictable shock. Psychosom. Med. 1970, 32, 397–408. [Google Scholar] [CrossRef]
- Weiss, J.M. Effects of coping behavior in different warning signal conditions on stress pathology in rats. J. Comp. Physiol. Psychol. 1971, 77, 1–13. [Google Scholar] [CrossRef]
- Boissy, A. Fear and fearfulness in animals. Q. Rev. Biol. 1995, 70, 165–191. [Google Scholar] [CrossRef] [PubMed]
- Mobbs, D.; Hagan, C.C.; Dalgleish, T.; Silston, B.; Prévost, C. The ecology of human fear: Survival optimization and the nervous system. Front. Neurosci. 2015, 9, 1–22. [Google Scholar] [CrossRef] [PubMed]
- Meis, J.; Kashima, Y. Signage as a tool for behavioral change: Direct and indirect routes to understanding the meaning of a sign. PLoS ONE 2017, 12, e0182975. [Google Scholar] [CrossRef] [PubMed]
- Marschall, S.; Granquist, S.M.; Burns, G.L. Interpretation in wildlife tourism: Assessing the effectiveness of signage on visitor behaviour at a seal watching site in Iceland. J. Outdoor Recreat. Tour. 2017, 17, 11–19. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Behaviour | Description |
|---|---|
| States | |
| Distance from visitor viewing area (m) | Side A: long/main visitor viewing ledge of the exhibit, within <1 m or >1 m of visitor viewing area. Side B: short visitor viewing ledge of the exhibit, within <1 m or >1 m of visitor viewing area. |
| Huddling (land) | Stationary, positioned within one flipper distance of at least one other penguin. |
| Resting (land) | Belly on the ground, in a prone position, with eyes open or closed. |
| Idle (land) | Standing on two feet with a relaxed posture, eyes open or closed; not visually scanning the environment. |
| Vigilance (land) | Standing on two feet, visually scanning the environment with head movements from left to right or vice versa. |
| Locomotion (land) | Upright position, moving from one location to another either by walking or running. |
| Surface swimming | Moving or floating on the surface of the water, with head erect or in the water. |
| Diving | Swimming under the water surface. |
| Events | |
| Preening (on land) | Running bill through plumage on land. |
| Preening (in water) | On the surface of the water and running bill through plumage. |
| Allopreening | Running bill through the plumage of another bird(s). |
| Agonistic interactions (one or a combination of these behaviours) | Peck: Directed at another individual in which an individual directly hits or strikes at another bird with its bill. Bill slap: hit or strike with the side of the bill.Bill joust: an individual interlocks bill with another individual’s bill.Lunge: sudden forward thrust of the body towards another individual.Chase: individual runs after another individual. |
| Flee | Diving or moving rapidly away from current position in response to a direct approach or interaction by human. |
| Retreat | Swimming or locomoting (on land) slowly away from human approach or presence. |
| Social interaction | On land: Approaching another individual with flippers back, pushing individual in a circular motion, nibbling and/or preening head or neck of the individual. In water: approaching another individual in the water, in a circular motion, nibbling and/or preening head or neck of the individual. |
| Manipulating inanimate object | Using bill to peck or nibble at an inanimate object such as a plant, stick, rock, grass, etc. |
| Chasing insect | Following a flying insect. |
| Interaction with keeper/staff | Approaching keepers and engaging in play, feeding and/or agonistic behaviours toward keepers. |
| Behaviour | Description |
|---|---|
| Tactile contact with enclosure features | Tapping or banging on the glass windows, barriers, or enclosure features. |
| Loud vocalisations | Shouts, screams, loud whistles to attract the animals’ attention. |
| Looming | Leaning on or over the exhibit barriers to view animals in the water or on land. Penguins > 0.5 m—penguins greater than 0.5 m away from looming position. Penguins < 0.5 m—penguins less than 0.5 m away from looming position. |
| Tactile contact with water | Touching/flicking/slapping the water with the hand(s), which creates ripples in the water. |
| Tactile contact with penguin | Touching penguin with the hand(s). |
| Sudden movement | Running, waving or jumping towards or at the penguin(s) and/or exhibit. |
| Visitor Variables | Covariate | Open Exhibit | p-Value | ||||||
|---|---|---|---|---|---|---|---|---|---|
| Increased Viewing Distance | Normal Viewing Distance | Proximity a | Regulation Effects a | Proximity × Regulation Interaction | |||||
| Regulated | Unregulated | Regulated | Unregulated | s.e.d. c | |||||
| Ambient noise (dB) * | Maximum temperature | 62 | 61 | 61 | 62 | 0.62 | 0.49 | 1.00 | 0.11 |
| Number of visitors * | Visitor gate numbers | 360 | 300 | 500 | 460 | 46 | 0.0047 | 0.080 | 0.77 |
| Behaviour | |||||||||
| Tactile contact with enclosure features | 0.91 (0.83) | 1.7 (2.8) | 5.1 (26) | 6.2 (39) | 1.18 | 0.00076 | 0.30 | 0.84 | |
| Loud vocalisations | Proportion of males | 8.5 (73) | 9.8 (97) | 7.0 (48) | 11 (123) | 1.87 | 0.98 | 0.075 | 0.33 |
| Looming with penguin > 0.5 m (from looming position) | 0.57 (0.32) | 1.6 (2.6) | 22 (492) | 21 (461) | 1.06 | 3.2 × 10−9 | 0.82 | 0.28 | |
| Looming with penguin < 0.5 m (from looming position) | 0 | 0 | 0.86 (0.74) | 1.3 (1.6) | 0.85 d | 0.061 b | 0.64 b | 0.64 b | |
| Tactile contact with water | 0 | 0 | 3.6 (13) | 5.1 (26) | 0.85 d | 0.00054b | 0.09 b | 0.09 b | |
| Tactile contact with penguin | 0 | 0 | 0 | 0.64 (0.41) | 0.36 d | 0.18 b | 0.18 b | 0.18 b | |
| Sudden movement | 2.0 (4) | 1.1 (1) | 5.0 (25) | 5.0 (25) | 1.63 | 0.019 | 0.72 | 0.73 | |
| Behaviour | Covariate | Closed Exhibit | Increased Viewing Distance | Normal Viewing Distance | s.e.d. c | p Value | ||||
|---|---|---|---|---|---|---|---|---|---|---|
| Proximity a | Regulation when Exhibit Open b | Proximity × Regulation Interaction | ||||||||
| Regulated | Unregulated | Regulated | Unregulated | |||||||
| States | ||||||||||
| Penguins visible | Visitor gate numbers | 40 (41) | 37 (36) | 43 (46) | 36 (35) | 39 (40) | 3.3 | 0.77 | 0.070 | 0.48 |
| Huddling | - | 30 (25) | 30 (25) | 40 (42) | 41 (43) | 56 (69) | 4.5 | 0.0011 | 0.0025 | 0.50 |
| <1 m from side A | - | 32 (28) | 22 (14) | 22 (14) | 7 (2) | 7 (1) | 5.4 | 0.00066 | 0.93 | 0.95 |
| <1 m from side B | - | 32 (28) | 27 (21) | 28 (21) | 8 (2) | 11 (4) | 6.4 | 0.0024 | 0.69 | 0.75 |
| Resting | - | 18 (10) | 24 (17) | 19 (11) | 19 (10) | 24 (16) | 4.5 | 0.70 | 0.96 | 0.15 |
| Idle | Maximum temperature | 36 (35) | 34 (31) | 38 (38) | 40 (41) | 45 (51) | 4.2 | 0.081 | 0.14 | 0.91 |
| Locomotion (on land) | Proportion male | 15 (7) | 15 (7) | 15 (7) | 19 (11) | 12 (5) | 1.7 | 0.85 | 0.023 | 0.019 |
| Vigilant | - | 10 (3) | 19 (10) | 19 (11) | 25 (17) | 28 (22) | 4.5 | 0.0060 | 0.53 | 0.60 |
| Surface swimming | - | 38 (39) | 33 (30) | 31 (26) | 16 (7) | 17 (9) | 5.1 | 0.00091 | 0.92 | 0.61 |
| Diving | Proportion male | 13 (5) | 12 (4) | 13 (5) | 8 (2) | 9 (2) | 2.6 | 0.077 | 0.69 | 0.83 |
| Events | ||||||||||
| Preen (land) | - | 19 (10) | 18 (9) | 18 (9) | 24 (16) | 22 (15) | 4.2 | 0.21 | 0.84 | 0.83 |
| Preen (water) | - | 9 (3) | 8 (2) | 11 (4) | 6 (1) | 4 (0.54) | 2.4 | 0.042 | 0.75 | 0.27 |
| Allopreen | - | 6 (1) | 4 (0.54) | 8 (2) | 6 (1) | 8 (2) | 1.7 | 0.57 | 0.039 | 0.45 |
| Peck | - | 3 (0.31) | 2 (0.18) | 3 (0.36) | 3 (0.33) | 5 (0.62) | 0.54 | 0.074 | 0.014 | 0.78 |
| Agonistic interactions | - | 4 (0.48) | 4 (0.39) | 4 (0.58) | 4 (0.51) | 5 (0.74) | 0.63 | 0.47 | 0.10 | 0.92 |
| Flee | Proportion of males | 2 (0.093) | 0.54 (0) | 0.13 (0) | 2 (0.098) | 1 (0.061) | 0.77 | 0.069 | 0.48 | 0.98 |
| Retreat | Proportion of males | 0.36 (0) | 0.17 (0) | 0.53 (0) | 2 (0.16) | 2 (0.14) | 0.38 | 0.00013 | 0.65 | 0.39 |
| Social interaction (land) | Proportion of males | 3 (0.24) | 3 (0.28) | 4 (0.37) | 2 (0.17) | 1 (0.020) | 1.1 | 0.11 | 0.52 | 0.23 |
| Social interaction (water) | - | 0.73 (0.016) | 1 (0.052) | 1 (0.035) | 0 | 0.32 (0) | 0.96 | 0.35 | 0.95 | 0.69 |
| Manipulate object | - | 6 (1) | 6 (1) | 5 (0.76) | 5 (0.74) | 3 (0.35) | 1.8 | 0.52 | 0.41 | 0.74 |
| Chase Insect | - | 1 (0.059) | 0.58 (0.010) | 1 (0.037) | 0.36 (0) | 0.32 (0) | 0.73 | 0.29 | 0.65 | 0.60 |
| Interaction with keeper | Maximum temperature | 0 | 0.59 (0.011) | 0.10 (0) | 0.28 (0) | 0.67 (0.014) | 0.45 | 0.43 | 0.67 | 0.20 |
| Physiology | ||||||||||
| FGM concentration (ng/g) | Maximum temperature | 1600 | 1140 | 1657 | 1272 | 1600 | 284 | 0.71 | 0.060 | 0.67 |
© 2019 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (http://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chiew, S.J.; Butler, K.L.; Sherwen, S.L.; Coleman, G.J.; Fanson, K.V.; Hemsworth, P.H. Effects of Regulating Visitor Viewing Proximity and the Intensity of Visitor Behaviour on Little Penguin (Eudyptula minor) Behaviour and Welfare. Animals 2019, 9, 285. https://doi.org/10.3390/ani9060285
Chiew SJ, Butler KL, Sherwen SL, Coleman GJ, Fanson KV, Hemsworth PH. Effects of Regulating Visitor Viewing Proximity and the Intensity of Visitor Behaviour on Little Penguin (Eudyptula minor) Behaviour and Welfare. Animals. 2019; 9(6):285. https://doi.org/10.3390/ani9060285
Chicago/Turabian StyleChiew, Samantha J., Kym L. Butler, Sally L. Sherwen, Grahame J. Coleman, Kerry V. Fanson, and Paul H. Hemsworth. 2019. "Effects of Regulating Visitor Viewing Proximity and the Intensity of Visitor Behaviour on Little Penguin (Eudyptula minor) Behaviour and Welfare" Animals 9, no. 6: 285. https://doi.org/10.3390/ani9060285