
Effects of Dietary Chenodeoxycholic Acid Supplementation in a Low Fishmeal Diet Containing Clostridium autoethanogenum Protein on Growth, Lipid and Cholesterol Metabolism, and Hepatopancreas Health of Litopenaeus vannamei
 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Experimental Diets
2.2. Shrimp and Experimental Conditions
2.3. Sampling and Chemical Analysis
2.4. Enzyme Activity Assays
2.5. Quantitative Real-Time PCR Analysis
2.6. Morphological Analysis of Hepatopancreas
2.7. Western Blot Analyses
2.8. Calculations and Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Proximate Composition of Whole Shrimp
3.3. Hemolymph and Hepatopancreas Biochemical Indicators
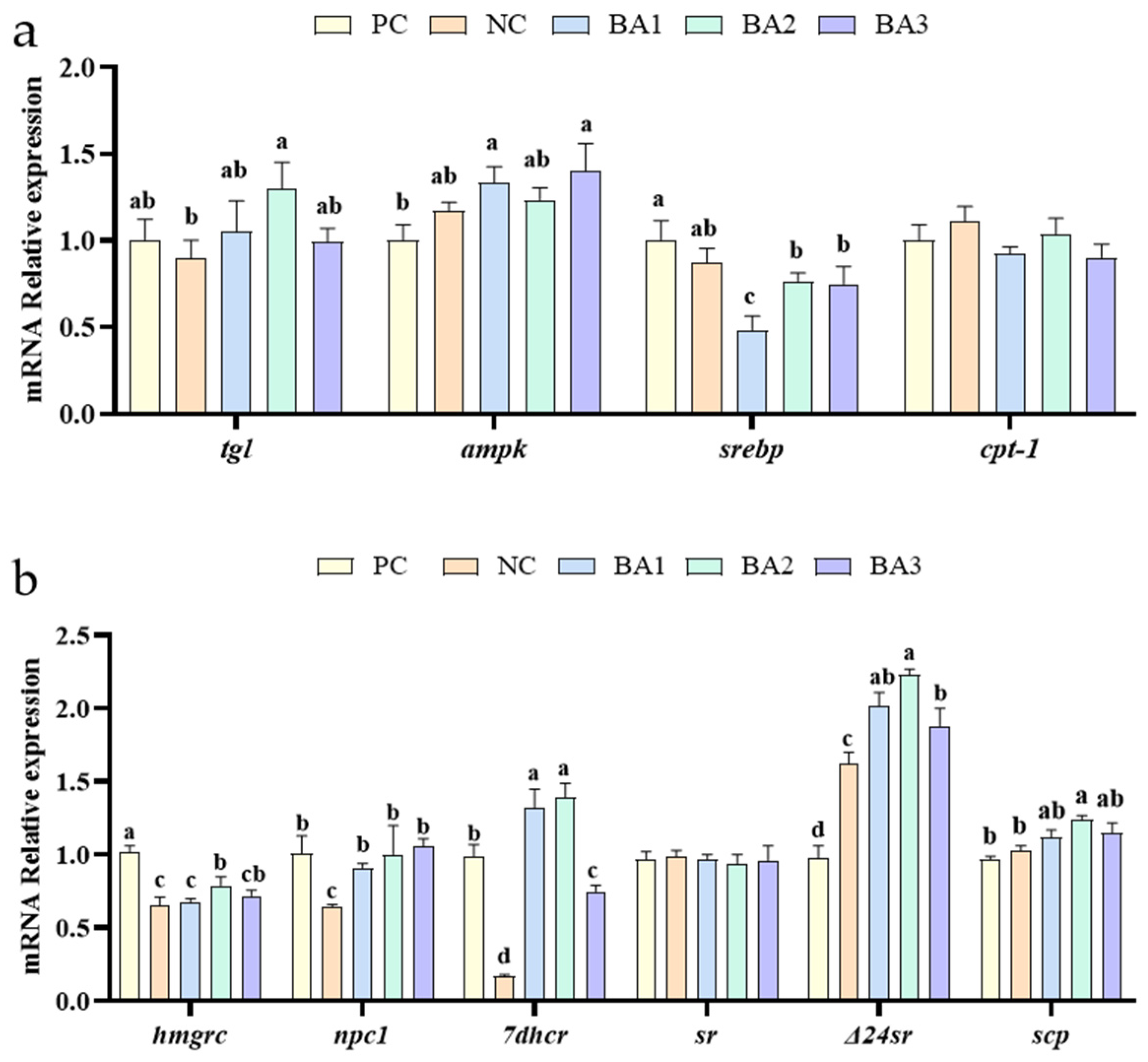
3.4. Lipid and Sterol Metabolism-Related Genes Expression
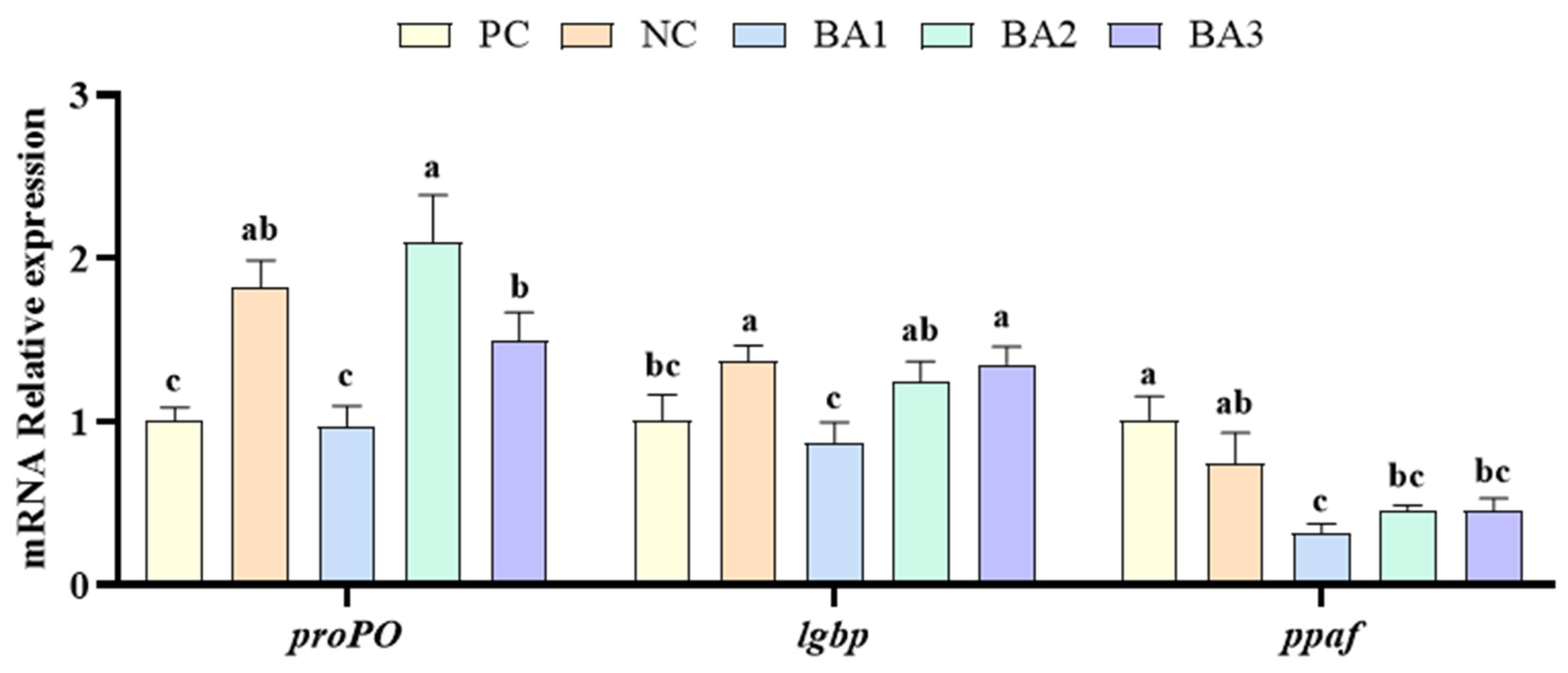
3.5. Phenol Oxidase-Related Genes Expression
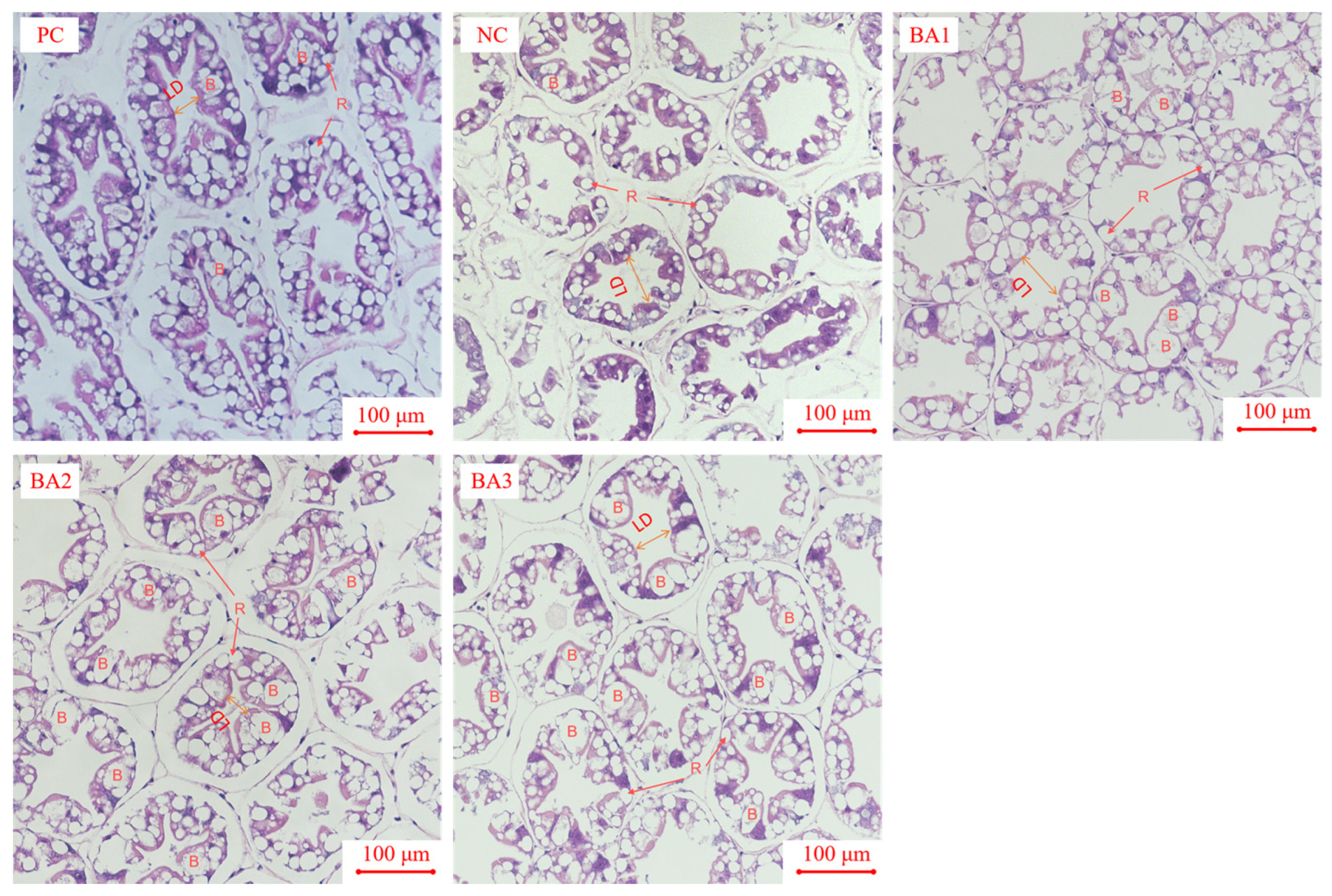
3.6. Morphological Analysis of Hepatopancreas
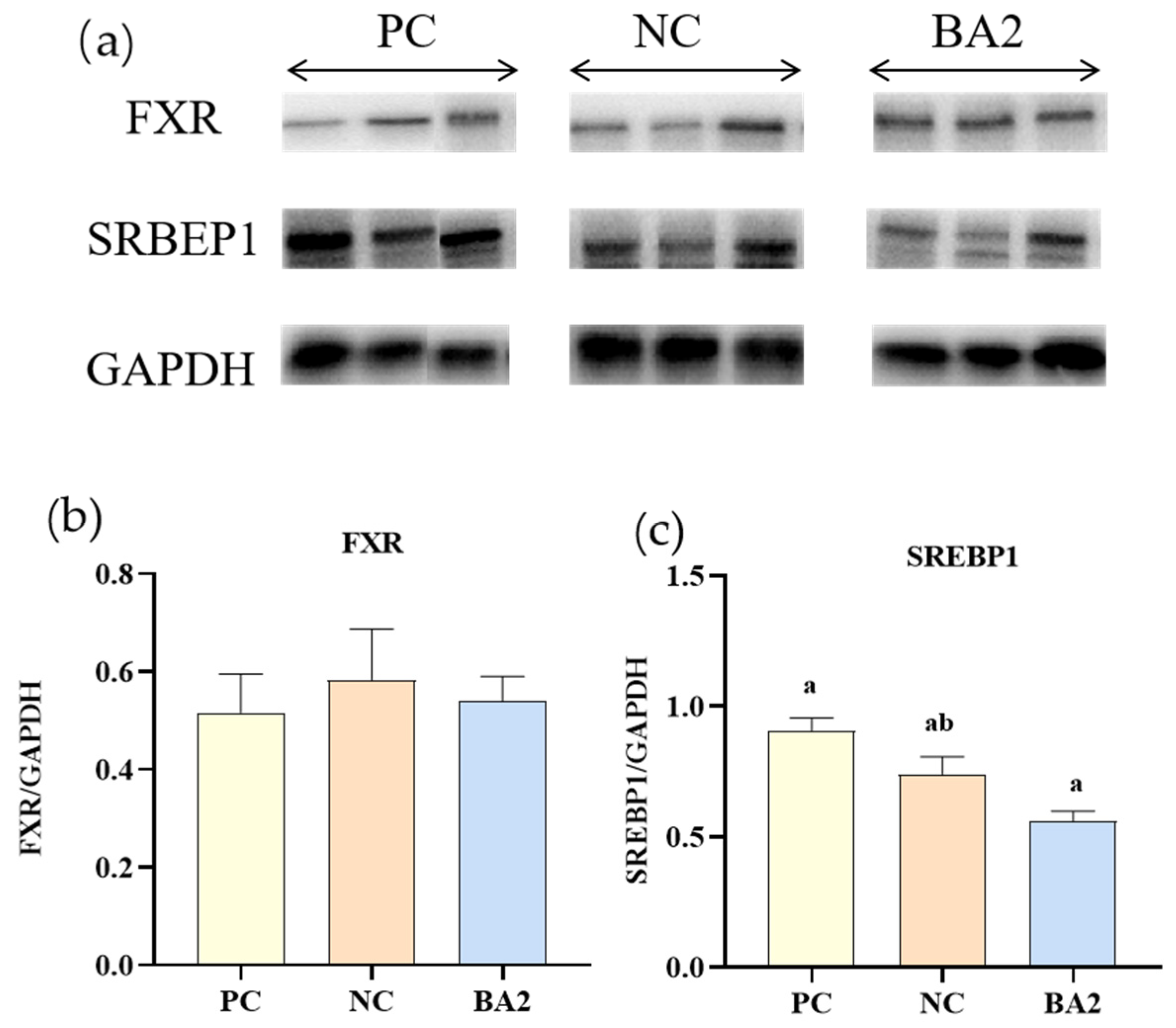
3.7. Western Blot Analysis of Bile Acid Metabolism Related Proteins Expression
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Conflicts of Interest
References
- Li, X.; Yao, X.; Zhang, X.; Dong, X.; Chi, S.; Tan, B.; Zhang, S.; Xie, S. Effects of dietary chenodeoxycholic acid supplementation in a low fishmeal diet on growth performance, lipid metabolism, autophagy and intestinal health of Pacific white shrimp. Litopenaeus vannamei. Fish Shellfish Immunol. 2022, 127, 1088–1099. [Google Scholar] [CrossRef]
- Wu, Y.; Li, R.; Shen, G.; Huang, F.; Yang, Q.; Tan, B.; Chi, S. Effects of dietary small peptides on growth, antioxidant capacity, nonspecific immunity and ingut microflorastructure of Litopenaeus vannamei. J. Guangdong Ocean Univ. 2021, 41, 1–9. [Google Scholar] [CrossRef]
- Huang, B.; Zhang, S.; Dong, X.; Chi, S.; Yang, Q.; Liu, H.; Tan, B.; Xie, S. Effects of fishmeal replacement by black soldier fly on growth performance, digestive enzyme activity, intestine morphology, intestinal flora and immune response of pearl gentian grouper (Epinephelus fuscoguttatus ♀ × Epinephelus lanceolatus ♂). Fish Shellfish. Immunol. 2022, 120, 497–506. [Google Scholar] [CrossRef]
- Xu, J.; Sheng, Z.; Chen, N.; Xie, R.; Zhang, H.; Li, S. Effect of dietary fish meal replacement with spray dried chicken plasma on growth, feed utilization and antioxidant capacity of largemouth bass (Micropterus salmoides). Aquac. Rep. 2022, 24, 101112. [Google Scholar] [CrossRef]
- Food and Agriculture Organization (F.A.O.). The State of World Fisheries and Aquaculture 2020; FAO Yearbook: Rome, Italy, 2020. [Google Scholar]
- Jiang, S.; Chen, Z.; Zhou, F.; Yang, Q.; Huang, J.; Yang, L.; Jiang, S. Study on partial replacement of fish meal with concentrated dephenolized cottonseed protein in feed of Penaeus monodon. Aquac. Res. 2020, 52, 3871–3881. [Google Scholar] [CrossRef]
- Ray, G.W.; Liang, D.; Yang, Q.; Tan, B.; Dong, X.; Chi, S.; Liu, H.; Zhang, S.; Rimei, L. Effects of replacing fishmeal with dietary soybean protein concentrate (SPC) on growth, serum biochemical indices, and antioxidative functions for juvenile shrimp Litopenaeus vannamei. Aquaculture 2020, 516, 734630. [Google Scholar] [CrossRef]
- Zhou, H.; Chen, G.; Ji, D.; Wang, S.; Zhang, J. The effects of partial rReplacement of fish meal by three protein sources on growth performance approximate compositions and energy budget of Juvenile Brown-marbled Grouper (Epinephelus fuscoguttatus). J. Guangdong Ocean Univ. 2012, 32, 10–16. [Google Scholar]
- Zhang, H.; Pu, X.; Yang, Q.; Tan, B.; Dong, X.H.; Chi, S.; Liu, H.Y.; Zhang, S. Effects of replacing fish meal with high-protein cottonseed meal on growth performance, non-specific immune index and disease resistance for Litopenaeus vannamei. J. Guangdong Ocean Univ. 2018, 38, 20–26. [Google Scholar] [CrossRef]
- Tu, G.; Chen, G.; Zhou, H.; Wang, S.; Ji, D.; Zhng, J.; Ye, F. Effects of partial replacement dietary fish meal by three kinds of protein sources on haematological indices of Juvenile Epinephelus fuscoguttatus. J. Guangdong Ocean Univ. 2012, 32, 12–19. [Google Scholar]
- Chen, Y.; Chi, S.; Zhang, S.; Dong, X.; Yang, Q.; Liu, H.; Tan, B.; Xie, S. Evaluation of Methanotroph (Methylococcus capsulatus, Bath) bacteria meal on body composition, lipid metabolism, protein synthesis and muscle metabolites of Pacific white shrimp (Litopenaeus vannamei). Aquaculture 2022, 547, 737517. [Google Scholar] [CrossRef]
- Xie, Y.; Peng, K.; Hu, J.; Wang, G. Review on application of black soldier fly (Hermetia illucens Linnaeus) in aquatic feed. J. Guangdong Ocean Univ. 2022, 42, 144–150. [Google Scholar] [CrossRef]
- Ayiku, S.; Shen, J.; Tan, B.P.; Dong, X.H.; Liu, H.Y. Effects of reducing dietary fishmeal with yeast supplementations on Litopenaeus vannamei growth, immune response and disease resistance against Vibrio harveyi. Microbiol. Res. 2020, 239, 126554. [Google Scholar] [CrossRef]
- Xie, S.; Wei, D.; Tian, L.; Liu, Y. Dietary supplementation of chenodeoxycholic acid improved the growth performance, immune response and intestinal health of juvenile Penaeus monodon fed a low fish-meal diet. Aquac. Rep. 2021, 20, 100773. [Google Scholar] [CrossRef]
- Zhang, W.; Tan, B.; Pang, A.; Deng, J.; Yang, Q.; Zhang, H. Screening of potential biomarkers for soybean meal induced enteritis in pearl gentian Grouper (Epinephelus fuscoguttatus♀ × Epinephelus lanceolatus♂). J. Guangdong Ocean Univ. 2022, 42, 1–12. [Google Scholar] [CrossRef]
- Chen, G.; Liu, B.; Chen, J.; Liu, H.; Tan, B.; Dong, X.; Yang, Q.; Chi, S.; Zhang, S.; Yao, M. Supplementing sulfate-based alginate polysaccharide improves Pacific white shrimp (Litopenaeus vannamei) fed fishmeal replacement with cottonseed protein concrntrate: Effects on growth, intestinal health, and disease resistance. Aquac. Nutr. 2022, 21, 7132362. [Google Scholar] [CrossRef]
- Jiang, X.; Yao, W.; Yang, H.; Tan, S.; Leng, X.; Li, X. Dietary effects of Clostridium autoethanogenum protein substituting fish meal on growth, intestinal histology and immunity of Pacific white shrimp (Litopenaeus vannamei) based on transcriptome analysis. Fish Shellfish. Immunol. 2021, 119, 635–644. [Google Scholar] [CrossRef]
- Yao, W.; Yang, P.; Zhang, X.; Xu, X.; Zhang, C.; Li, X.; Leng, X. Effects of replacing dietary fish meal with Clostridium autoethanogenum protein on growth and flesh quality of Pacific white shrimp (Litopenaeus vannamei). Aquaculture 2022, 549, 737770. [Google Scholar] [CrossRef]
- Zheng, C.; Cao, J.; Chi, S.; Dong, X.; Yang, Q.; Liu, H.; Zhang, S.; Xie, S.; Tan, B. Dietary phosphorus supplementation in the diet of Pacific white shrimp (Litopenaeus vannamei) alleviated the adverse impacts caused by high Clostridium autoethanogenum protein. Fish Shellfish. Immunol. 2022, 131, 137–149. [Google Scholar] [CrossRef]
- Zheng, C.; Xie, S.; Tan, B.; Cao, J.; Chi, S.; Dong, X.; Yang, Q.; Liu, Y.H.; Zhang, S. Effects of dietary lipid sources on alleviating the negative impacts induced by the fishmeal replacement with Clostridium autoethanogenum protein in the diet of pacific white shrimp (Litopenaeus vannamei). Front. Mar. Sci. 2022, 9, 879364. [Google Scholar] [CrossRef]
- Wu, Y.; Tian, S.; Yuan, J.; Zhang, Z.; Zhou, H.; Gao, W.; Zhang, W.; Mai, K. Effects of Clostridium autoethanogenum protein as substitute for dietary fishmeal on the growth, feed utilization, intestinal health and muscle quality of large yellow croaker Larimichthys crocea. Aquaculture 2022, 561, 738591. [Google Scholar] [CrossRef]
- Ma, S.; Liang, X.; Chen, P.; Wang, J.; Gu, X.; Qin, Y.; Blecker, C.; Xue, M. A new single-cell protein from Clostridium autoethanogenum as afunctional protein for largemouth bass (Micropterus salmoides). Animal Nutr. 2022, 10, 99–110. [Google Scholar] [CrossRef]
- Yao, W. Dietary Effects of Replacing Fish Meal with Clostridium autoethanogenum Protein on Growth and Flesh Quality of Pacific White Shrimp (Litopenaeus vannamei) and Nutritional Improvement Strategy; Shanghai Ocean University: Shanghai, China, 2022. [Google Scholar]
- Lu, Q.; Xi, L.; Liu, Y.; Gong, Y.; Su, J.; Han, D.; Yang, Y.; Jin, J.; Liu, H.; Zhu, X. Effects of dietary inclusion of Clostridium autoethanogenum protein on the growth performance and liver health of largemouth bass (Micropterus salmoides). Front. Mar. Sci. 2021, 8, 764964. [Google Scholar] [CrossRef]
- Chen, Y.; Zheng, C.; Zhang, X.; Li, X.; Yao, X.; He, G.; Cao, J.; Chi, S.; Zhang, S.; Tan, B. Evaluation of ratios of fish-to-soybean oils on growth, lipid and cholesterol metabolism and muscle metabolites of Pacific white shrimp (Litopenaeus vannamei) fed low fishmeal diets containing Clostridium autoethanogenum protein. Aquac. Rep. 2022, 27, 101417. [Google Scholar] [CrossRef]
- Hylemon, P.B.; Takabe, K.; Dozmorov, M.; Nagahashi, M.; Zhou, H. Bile acids as global regulators of hepatic nutrient metabolism. Liver Res. 2017, 1, 10–16. [Google Scholar] [CrossRef]
- Su, C.; Liu, X.; Li, J.; Zhang, M.; Pan, L.; Lu, Y.; Wang, Y.; Ding, Y. Effects of bile acids on the growth performance, lipid metabolism, non-specific immunity and intestinal microbiota of Pacific white shrimp (Litopenaeus vannamei). Aquac. Nutr. 2021, 27, 2029–2041. [Google Scholar] [CrossRef]
- Liao, M.; Chen, S.; Wang, W.; Zhong, S.; Yv, S.; Liao, S. Cholesterol-lowering activity of Sargassum zhangii polysaccharide in vitro. J. Guangdong Ocean Univ. 2017, 37, 80–85. [Google Scholar] [CrossRef]
- Li, X.; Shi, M.; Chen, L.; Zhang, S.; Chi, S.; Dong, X.; Deng, J.; Tan, B.; Xie, S. Effects of bile acids supplemented into low fishmeal diet on growth, molting, and intestinal health of Pacific white shrimp, Litopenaeus vannamei. Aquac. Rep. 2023, 29, 101491. [Google Scholar] [CrossRef]
- Du, J.; Xu, H.; Li, S.; Cai, Z.; Mai, K.; Ai, Q. Effects of dietary chenodeoxycholic acid on growth performance, body composition and related gene expression in large yellow croaker (Larimichthys crocea) fed diets with high replacement of fish oil with soybean oil. Aquaculture 2017, 479, 584–590. [Google Scholar] [CrossRef]
- Xu, J.; He, G.; Chen, L.; Xie, S.; Chi, S.; Zhang, S.; Cao, J.; Tan, B. Farnesoid X receptor (FXR) and G protein-coupled bile acid receptor 1 (TGR5) signaling pathways improved the hepatic lipid metabolism in hybrid grouper. Aquac. Rep. 2022, 22, 100997. [Google Scholar] [CrossRef]
- Li, X.; Li, H.; Qu, K.; Liu, Y.; Chi, S.; Yang, Q.; Tan, B.; Zhang, S.; Xie, S. Dietary bile acids promote sterol metabolism, bile acids enterohepatic circulation, and apoptosis in juvenile Pacific white shrimp (Litopenaeus vannamei). Anim. Feed. Sci. Technol. 2023, 115710. [Google Scholar] [CrossRef]
- Iwashita, Y.; Suzuki, N.; Yamamoto, T.; Shibata, J.-i.; Isokawa, K.; Soon, A.H.; Ikehata, Y.; Furuita, H.; Sugita, T.; Goto, T. Supplemental effect of cholyltaurine and soybean lecithin to a soybean meal-based fish meal-free diet on hepatic and intestinal morphology of rainbow trout Oncorhynchus mykiss. Fish. Sci. 2008, 74, 1083–1095. [Google Scholar] [CrossRef]
- Reschly, E.J.; Ai, N.; Ekins, S.; Welsh, W.J.; Hagey, L.R.; Hofmann, A.F.; Krasowski, M.D. Evolution of the bile salt nuclear receptor FXR in vertebrates *s. J. Lipid Res. 2008, 49, 1577–1587. [Google Scholar] [CrossRef] [Green Version]
- Jiang, M.; Wen, H.; Gou, G.; Liu, T.; Lu, X.; Deng, D. Preliminary study to evaluate the effects of dietary bile acids on growth performance and lipid metabolism of juvenile genetically improved farmed tilapia (Oreochromis niloticus) fed plant ingredient-based diets. Aquac. Nutr. 2018, 24, 1175–1183. [Google Scholar] [CrossRef]
- Zhou, J.; Chen, H.; Ji, H.; Shi, X.; Li, X.; Chen, L.; Du, Z.; Yu, H. Effect of dietary bile acids on growth, body composition, lipid metabolism and microbiota in grass carp (Ctenopharyngodon idella). Aquac. Nutr. 2018, 24, 802–813. [Google Scholar] [CrossRef]
- Cao, B.; Liao, Z.; Xiang, X.; He, W.; Cen, Y.; He, S. Effect of bile acids on growth performance, muscle nutrient composition and digestive enzyme activity in grass carp (Ctenopharyngodon idellus). Prog. Fish. Sci. 2017, 38, 99–106. [Google Scholar] [CrossRef]
- Ogata, Y.; Nishi, M.; Nakayama, H.; Kuwahara, T.; Ohnishi, Y.; Tashiro, S. Role of bile in intestinal barrier function and its inhibitory effect on bacterial translocation in obstructive jaundice in rats. J. Surg. Res. 2003, 115, 18–23. [Google Scholar] [CrossRef]
- Chen, Q.; Liu, H.; Tan, B.; Dong, X.; Chi, S.; Yang, Q.; Zhang, S. Effects of dietary cholesterol level on growth performance, blood biochemical parameters and lipid metabolism of juvenile Cobia (Rachycentron canadum). J. Guangdong Ocean Univ. 2016, 36, 35–43. [Google Scholar] [CrossRef]
- Shao, Y.; Chen, C.; Zhang, T.; Li, Y.; Zhang, M.; Li, W. Influence of high temperature stress on survival rate and serum biochemical indexes of 2 Epinephelus Hybrids. J. Guangdong Ocean Univ. 2017, 37, 89–95. [Google Scholar] [CrossRef]
- Rivera-Pérez, C.; García-Carreño, F. Purification and characterization of an intracellular lipase from pleopods of whiteleg shrimp (Litopenaeus vannamei). Comp. Biochem. Physiol. Part B Biochem. Mol. Biol. 2011, 158, 99–105. [Google Scholar] [CrossRef]
- Polakof, S.; Panserat, S.; Craig, P.; Martyres, D.; Plagnes-Juan, E. The Metabolic Consequences of Hepatic AMP-Kinase Phosphorylation in Rainbow Trout. PLoS ONE 2011, 6, e20228. [Google Scholar] [CrossRef] [Green Version]
- Jump, D.B. Dietary polyunsaturated fatty acids and regulation of gene transcription. Curr. Opin. Lipidol. 2002, 13, 155–164. [Google Scholar] [CrossRef]
- Yamauchi, Y.; Rogers, M.A. Sterol metabolism and transport in atherosclerosis and cancer. Front. Endocrinol. 2018, 9, 509. [Google Scholar] [CrossRef] [Green Version]
- Endo, A. The discovery and development of HMG-CoA reductase inhibitors. J. Lipid Res. 1992, 33, 1569–1582. [Google Scholar] [CrossRef]
- Rozner, S.; Garti, N. The activity and absorption relationship of cholesterol and phytosterols. Colloids Surf. A Physicochem. Eng. Asp. 2006, 282, 435–456. [Google Scholar] [CrossRef]
- Liland, N.S.; Espe, M.; Rosenlund, G.; Waagbø, R.; Hjelle, J.I.; Lie, Ø.; Fontanillas, R.; Torstensen, B.E. High levels of dietary phytosterols affect lipid metabolism and increase liver and plasma TAG in Atlantic salmon (Salmo salar L.). Br. J. Nutr. 2013, 110, 1958–1967. [Google Scholar] [CrossRef] [Green Version]
- Fernández, C.; Suárez, Y.; Ferruelo, A.J.; Gómez-Coronado, D.; Lasunción, M.A. Inhibition of cholesterol biosynthesis by Δ22-unsaturated phytosterols via competitive inhibition of sterol Δ24-reductase in mammalian cells. Biochem. J. 2002, 366, 109–119. [Google Scholar] [CrossRef] [Green Version]
- Pfeffer, S.R. NPC intracellular cholesterol transporter 1 (NPC1)-mediated cholesterol export from lysosomes. J. Biol. Chem. 2019, 294, 1706–1709. [Google Scholar] [CrossRef] [Green Version]
- Cao, J.; Yan, J.; Wang, G.; Huang, Y.; Zhang, R.; Zhou, T.; Liu, Q.; Sun, Z. Effects of replacement of fish meal with housefly maggot meal on digestive enzymes, transaminases activities and hepatopancreas histological structure of Litopenaeus vannamei. South China Fish. Sci. 2012, 8, 72–79. [Google Scholar] [CrossRef]
- Xv, D.; Wu, J.; Sun, L.; Qin, X.; Fan, X. Physiological rResponse and metabolicregulation of Litopenaeus vannamei exposed to combination stress of acute cold exposure and chronic waterless duration. J. Guangdong Ocean Univ. 2022, 42, 20–28. [Google Scholar] [CrossRef]
- DJ, J.; CG, A.; Yellowlees, D. Epithelial cytology and function in the digestive gland of Thenus orientalis (Decapoda: Scyllaridae). J. Crustac. Biol. 1998, 18, 271–278. [Google Scholar] [CrossRef]
- Deng, S.; Ye, M.; Zhu, C.; Chen, H.; Li, G. Influence of eyestalk ablation on biochemical composition of ovary, hepatopancreas and serum in pacific white shrimp, Litopenaeus vannamei broodstock. J. Guangdong Ocean Univ. 2016, 36, 29–34. [Google Scholar] [CrossRef]
- Xu, C.; Li, E.; Liu, Y.; Wang, S.; Wang, X.; Chen, K.; Qin, J.; Chen, L. Effect of dietary lipid level on growth, lipid metabolism and health status of the Pacific white shrimp Litopenaeus vannamei at two salinities. Aquac. Nutr. 2018, 24, 204–214. [Google Scholar] [CrossRef]
- Romano, N.; Koh, C.-B.; Ng, W.-K. Dietary microencapsulated organic acids blend enhances growth, phosphorus utilization, immune response, hepatopancreatic integrity and resistance against Vibrio harveyi in white shrimp, Litopenaeus vannamei. Aquaculture 2015, 435, 228–236. [Google Scholar] [CrossRef]
- Chen, W.W.; Romano, N.; Ebrahimi, M.; Natrah, I. The effects of dietary fructooligosaccharide on growth, intestinal short chain fatty acids level and hepatopancreatic condition of the giant freshwater prawn (Macrobrachium rosenbergii) post-larvae. Aquaculture 2017, 469, 95–101. [Google Scholar] [CrossRef]
- Poupon, R. Ursodeoxycholic acid and bile-acid mimetics as therapeutic agents for cholestatic liver diseases: An overview of their mechanisms of action. Clin. Res. Hepatol. Gastroenterol. 2012, 36, S3–S12. [Google Scholar] [CrossRef]





{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Ingredients | Diets | ||||
|---|---|---|---|---|---|
| PC | NC | BA1 | BA2 | BA3 | |
| Fishmeal 1 | 25.00 | 12.50 | 12.50 | 12.50 | 12.50 |
| Clostridium autoethanogenum protein 2 | 0.00 | 10.00 | 10.00 | 10.00 | 10.00 |
| Fish oil | 1.50 | 2.70 | 2.70 | 2.70 | 2.70 |
| Soybean oil | 2.50 | 2.50 | 2.50 | 2.50 | 2.50 |
| Soybean lecithin | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Soybean meal | 25.00 | 25.00 | 25.00 | 25.00 | 25.00 |
| Peanut meal | 10.00 | 10.00 | 10.00 | 10.00 | 10.00 |
| Shrimp shell meal | 5.00 | 5.00 | 5.00 | 5.00 | 5.00 |
| Beer yeast | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 |
| Wheat flour | 23.55 | 23.55 | 23.55 | 23.55 | 23.55 |
| Vitamin C | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 |
| Choline chloride | 0.30 | 0.30 | 0.30 | 0.30 | 0.30 |
| Vitamin and Mineral Premix 3 | 1.00 | 1.00 | 1.00 | 1.00 | 1.00 |
| Ethoxyquin | 0.05 | 0.05 | 0.05 | 0.05 | 0.05 |
| Cellulose microcrystalline | 1.00 | 0.80 | 0.77 | 0.74 | 0.71 |
| CaH2PO4 | 1.00 | 2.50 | 2.50 | 2.50 | 2.50 |
| Chenodeoxycholic acid | 0.00 | 0.00 | 0.03 | 0.06 | 0.09 |
| Proximate composition | |||||
| Dry matter | 89.72 | 89.84 | 89.9 | 89.94 | 89.86 |
| Crude protein | 44.15 | 43.69 | 44.10 | 44.09 | 43.71 |
| Crude lipid | 8.54 | 8.48 | 8.66 | 8.65 | 8.31 |
| Gross energy (KI/g) | 20.04 | 20.33 | 20.37 | 20.35 | 20.29 |
| Gene Name | Forward Primers (5′−3′) | Reverse Primers (5′−3′) | GenBank No. |
|---|---|---|---|
| β-actin | CCACGAGACCACCTACAAC | AGCGAGGGCAGTGATTTC | AF300705.2 |
| proPO | TCCATTCCGTCCGTCTG | GGCTTCGCTCTGGTTAGG | AY723296 |
| lgbp | CCATGTCCGGCGGTGGAA | GTCATCGCCCTTCCAGTTG | AY723297 |
| ppaf | GAGAAGGAGCTGAACCTGTAC | AGCGCCTGAGTTGTAGTTAG | JX644454.1 |
| tgl | ACAAGGTGGATAAGGAAGAG | TAATCAGTAGTTGGCGAAGA | XM_027361886.1 |
| cpt-1 | CAACTTCTACGGCACTGAT | GTCGGTCCACCAATCTTC | XM_027361886·1 |
| ampk | TCAGAGGAGGAGCAGGAAC | CCCGAGGTCTAATAGGCAC | KP272117·1 |
| srebp | ACCATTGCCACTCCCCTA | GTTGCGTTTCTCGCCTTI | MG770374.1 |
| hmgcr | AGGTGCCCACAAAGACACTC | TGATAGTTCCCCAGCCAGGA | XM_027354586.1 |
| scp | TCAGAGGAAATGAACGGGGG | TGGAAGCAGTACACACCCTT | XM_027375905.1 |
| Δ24sr | TGCTGATTGTGCTACCGCTT | TGCTGATTGTGCTACCGCTT | XM_027382756.1 |
| sr | TGCTTGGACCATTCAAGGGG | ACCCGCATAGTCTCTTGTGC | XM_027383297.1 |
| 7dhcr | AGACCTGTTACGGCTGTTGAG | GACTGGTCGGGACTCCAAAA | XM_027377095.1 |
| npc1 | CGAAGGGGAAAAGCCAGAGT | TTGAGGAGGAAGGGAGCGTA | XM_027363410.1 |
| Diets | |||||
|---|---|---|---|---|---|
| PC | NC | BA1 | BA2 | BA3 | |
| IBW (g) | 0.25 ± 0.03 | 0.25 ± 0.03 | 0.25 ± 0.03 | 0.25 ± 0.03 | 0.25 ± 0.03 |
| FBW (g) | 5.15 ± 0.06 a | 4.79 ± 0.05 b | 4.89 ± 0.08 b | 4.96 ± 0.10 ab | 4.77 ± 0.09 b |
| WG (%) | 1927.18 ± 23.78 a | 1787.45 ± 18.08 b | 1823.38 ± 29.92 b | 1853.51 ± 38.18 ab | 1778.39 ± 36.95 b |
| SGR (%d−1) | 5.37 ± 0.02 a | 5.25 ± 0.02 b | 5.28 ± 0.03 b | 5.31 ± 0.04 ab | 5.24 ± 0.04 b |
| SR (%) | 95.56 ± 3.85 | 95.56 ± 7.70 | 100 ± 0.00 | 95.56 ± 3.85 | 94.44 ± 6.94 |
| FCR | 1.17 ± 0.04 | 1.17 ± 0.04 | 1.16 ± 0.01 | 1.22 ± 0.03 | 1.22 ± 0.05 |
| FI (g/shrimp) | 6.35 ± 0.18 | 6.14 ± 0.31 | 5.94 ± 0.12 | 6.35 ± 0.23 | 6.16 ± 0.18 |
| PER | 1.77 ± 0.02 a | 1.67 ± 0.04 c | 1.75 ± 0.02 ab | 1.72 ± 0.01 abc | 1.69 ± 0.02 bc |
| Diets | |||||
|---|---|---|---|---|---|
| PC | NC | BA1 | BA2 | BA3 | |
| Moisture | 77.46 ± 0.27 | 77.43 ± 0.37 | 77.10 ± 0.22 | 77.74 ± 0.23 | 77.51 ± 0.35 |
| Crude protein | 17.28 ± 0.21 ab | 17.01 ± 0.11 b | 17.87 ± 0.11 a | 17.30 ± 0.16 ab | 17.64 ± 0.38 a |
| Crude lipid | 1.85 ± 0.06 a | 1.64 ± 0.06 b | 1.49 ± 0.07 bc | 1.41 ± 0.04 c | 1.33 ± 0.08 c |
| Diets | |||||
|---|---|---|---|---|---|
| PC | NC | BA1 | BA2 | BA3 | |
| Haemolymph | |||||
| TG (mmol/L) | 3.67 ± 0.15 | 3.51 ± 0.36 | 3.63 ± 0.09 | 3.67 ± 0.26 | 3.57 ± 0.22 |
| T-CHOL (mmol/L) | 3.16 ± 0.23 a | 1.62 ± 0.16 b | 1.50 ± 0.17 b | 1.73 ± 0.17 b | 1.87 ± 0.24 b |
| HDL-C(mmol/L) | 1.36 ± 0.29 abc | 0.81 ± 0.32 c | 0.84 ± 0.16 bc | 1.59 ± 0.32 a | 1.56 ± 0.22 ab |
| LDL-C(mmol/L) | 0.62 ± 0.04 a | 0.23 ± 0.03 c | 0.19 ± 0.02 c | 0.39 ± 0.04 b | 0.51 ± 0.09 ab |
| ALT (U/L) | 5.83 ± 1.35 b | 11.33 ± 2.24 a | 5.65 ± 1.04 b | 4.17 ± 1.25 b | 3.75 ± 0.08 b |
| AST (U/L) | 21.29 ± 1.85 ab | 23.94 ± 1.52 a | 18.94 ± 1.95 ab | 18.73 ± 3.03 ab | 16.87 ± 2.50 b |
| Hepatopancreas | |||||
| T-CHOL (mmol/L) | 0.93 ± 0.13 a | 0.60 ± 0.05 cd | 0.51 ± 0.05 d | 0.72 ± 0.05 bc | 0.81 ± 0.04 ab |
| LPS (U/gprot) | 1.07 ± 0.28 b | 1.52 ± 0.50 ab | 2.49 ± 0.54 a | 2.04 ± 0.27 ab | 2.02 ± 0.31 ab |
Disclaimer/Publisher’s Note: The statements, opinions and data contained in all publications are solely those of the individual author(s) and contributor(s) and not of MDPI and/or the editor(s). MDPI and/or the editor(s) disclaim responsibility for any injury to people or property resulting from any ideas, methods, instructions or products referred to in the content. |
© 2023 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Shi, M.; Zheng, C.; Sun, Y.; Li, X.; He, G.; Cao, J.; Tan, B.; Xie, S. Effects of Dietary Chenodeoxycholic Acid Supplementation in a Low Fishmeal Diet Containing Clostridium autoethanogenum Protein on Growth, Lipid and Cholesterol Metabolism, and Hepatopancreas Health of Litopenaeus vannamei. Animals 2023, 13, 2109. https://doi.org/10.3390/ani13132109
Shi M, Zheng C, Sun Y, Li X, He G, Cao J, Tan B, Xie S. Effects of Dietary Chenodeoxycholic Acid Supplementation in a Low Fishmeal Diet Containing Clostridium autoethanogenum Protein on Growth, Lipid and Cholesterol Metabolism, and Hepatopancreas Health of Litopenaeus vannamei. Animals. 2023; 13(13):2109. https://doi.org/10.3390/ani13132109
Chicago/Turabian StyleShi, Menglin, Chaozhong Zheng, Yidan Sun, Xiaoyue Li, Guilun He, Junming Cao, Beiping Tan, and Shiwei Xie. 2023. "Effects of Dietary Chenodeoxycholic Acid Supplementation in a Low Fishmeal Diet Containing Clostridium autoethanogenum Protein on Growth, Lipid and Cholesterol Metabolism, and Hepatopancreas Health of Litopenaeus vannamei" Animals 13, no. 13: 2109. https://doi.org/10.3390/ani13132109