
Occurrence of Chlamydiae in Corvids in Northeast Italy
by
 , and
, and
Rachid Aaziz
1,†, 
Karine Laroucau
1,†,
Federica Gobbo
2,
Daniela Salvatore
3,
Christiane Schnee
4,
Calogero Terregino
2,
Caterina Lupini
3 and
Antonietta Di Francesco
3,*
1
Bacterial Zoonoses Unit, Animal Health Laboratory, Anses, University Paris-Est, 94700 Maisons-Alfort, France
2
Istituto Zooprofilattico Sperimentale delle Venezie, 35020 Legnaro, PD, Italy
3
Department of Veterinary Medical Sciences, University of Bologna, 40064 Ozzano dell’Emilia, BO, Italy
4
Institute of Molecular Pathogenesis, Friedrich-Loeffler-Institut (Federal Research Institute for Animal Health), D-07743 Jena, Germany
*
Author to whom correspondence should be addressed.
†
These authors contributed equally to this work.
Animals 2022, 12(10), 1226; https://doi.org/10.3390/ani12101226
Submission received: 8 April 2022
/
Revised: 6 May 2022
/
Accepted: 9 May 2022
/
Published: 10 May 2022
(This article belongs to the Special Issue Animal Chlamydioses)
Abstract
:Simple Summary
The continuous improvement of next-generation sequencing techniques has led to an expansion of the number of Chlamydia species, as well as their host range. Recent studies performed on wild birds have detected Chlamydia strains with characteristics intermediate between Chlamydia psittaci and Chlamydia abortus. In this study, 12/108 corvids tested positive for Chlamydia by real-time PCR. Molecular characterisation at the species level was possible for eight samples, with one positive for C. psittaci and seven for C. abortus. Considering the well-known zoonotic role of C. psittaci and that a potential zoonotic role of avian C. abortus strains cannot be excluded, people who may have professional or other contact with wild birds should take appropriate preventive measures.
Abstract
Chlamydiaceae occurrence has been largely evaluated in wildlife, showing that wild birds are efficient reservoirs for avian chlamydiosis. In this study, DNA extracted from cloacal swabs of 108 corvids from Northeast Italy was screened for Chlamydiaceae by 23S real-time (rt)PCR. The positive samples were characterised by specific rtPCRs for Chlamydia psittaci, Chlamydia abortus, Chlamydia gallinacea, Chlamydia avium, Chlamydia pecorum and Chlamydia suis. Cloacal shedding of Chlamydiaceae was detected in 12 out of 108 (11.1%, 5.9%–18.6% 95% CI) corvids sampled. Molecular characterisation at the species level was possible in 8/12 samples, showing C. psittaci positivity in only one sample from a hooded crow and C. abortus positivity in seven samples, two from Eurasian magpies and five from hooded crows. Genotyping of the C. psittaci-positive sample was undertaken via PCR/high-resolution melting, clustering it in group III_pigeon, corresponding to the B genotype based on former ompA analysis. For C. abortus genotyping, multilocus sequence typing was successfully performed on the two samples with high DNA load from Eurasian magpies, highlighting 100% identity with the recently reported Polish avian C. abortus genotype 1V strain 15-58d44. To confirm the intermediate characteristics between C. psittaci and C. abortus, both samples, as well as two samples from hooded crows, showed the chlamydial plasmid inherent in most C. psittaci and avian C. abortus, but not in ruminant C. abortus strains. The plasmid sequences were highly similar (≥99%) to those of the Polish avian C. abortus genotype 1V strain 15-58d44. To our knowledge, this is the first report of avian C. abortus strains in Italy, specifically genotype 1V, confirming that they are actively circulating in corvids in the Italian region tested.
1. Introduction
Chlamydiae (order Chlamydiales, family Chlamydiaceae, genus Chlamydia) are Gram-negative obligate intracellular bacteria detected worldwide in a broad host range, including humans and livestock, as well as companion, wild and exotic animals [1].
In recent years, the continuous improvement of next-generation sequencing techniques has led to an expansion of the number of Chlamydia species, as well as their host range [2]. Today, the genus Chlamydia includes 14 species, namely C. trachomatis, C. pneumoniae, C. psittaci, C. abortus, C. pecorum, C. suis, C. felis, C. caviae, C. muridarum, C. gallinacea, C. avium, C. serpentis, C. poikilothermis and C. buteonis [3,4,5], plus a further four Candidatus (Ca) species, namely Ca C. ibidis, Ca C. sanzinia, Ca C. corallus and Ca C. testudinis [6,7,8,9]. In addition, the genus Chlamydiifrater (Cf.), with the two species Cf. phoenicopteri and Cf. volucris, was recently introduced [10].
In birds, C. psittaci is the longest known aetiological agent of chlamydiosis, detected in poultry, pet and free-living birds [11]. Depending on the virulence of the strain and the avian host, chlamydiosis can be subclinical or characterised by ocular, respiratory and enteric signs, with intermittent bacterial excretion, especially in stressful situations (migration, breeding, illness). The detection and differentiation of C. psittaci strains were initially performed by monoclonal antibody typing [12,13,14], obtaining six avian serovars (A–F). Later, the serotyping method was replaced by faster genotyping techniques, obtaining A–F genotypes and an additional genotype E/B [15]. The transition to DNA-based typing methods was facilitated by the equivalence detected in most cases between serotypes and genotypes [14]. All genotypes were considered to be readily transmissible to humans. In addition, other avian C. psittaci genotypes, designated 1V, 6N, Mat116, R54, YP84, CPX0308, I, J, G1 and G2, have been proposed [16,17,18]. Recently, C. psittaci genotype M56, originally isolated from muskrat, has also been highlighted in wild raptors [19,20].
For a long time, C. psittaci has been considered the only agent of chlamydiosis in birds. However, recent studies have proposed three new avian species and one Candidatus species: C. gallinacea from poultry, C. avium from pigeons and psittacine birds [21], C. buteonis from raptors [5] and Ca C. ibidis from African Sacred Ibis [6]. Moreover, other chlamydial agents could be involved in avian chlamydiosis, considering that C. abortus, C. pecorum, C. trachomatis, C. suis, C. pneumoniae and C. muridarum were molecularly detected in birds [22,23,24,25,26].
In light of this evidence and the multiple PCR-based detection methods recently developed, the purpose of this study was to investigate the presence of Chlamydiaceae species in corvids in Italy, and to then characterise them by fast and high discriminant molecular techniques such as species-specific real-time (rt) PCR assays, multilocus sequence typing (MLST) and PCR/high-resolution melting (HRM) analysis.
2. Materials and Methods
2.1. Sampling
From April to June 2021, 108 dead birds, including 52 Eurasian magpies (Pica pica), 38 hooded crows (Corvus cornix) and 18 Eurasian jays (Garrulus glandarius), from the Veneto region (northern Italy) were submitted to the Istituto Zooprofilattico Sperimentale delle Venezie for disease surveillance activities (such as Avian Influenza Virus, West Nile Virus and Usutu virus). A cloacal swab was collected from each bird carcass and immediately stored at −20 °C.
2.2. DNA Extraction
Total DNA was individually extracted from each sample using the QIAamp DNA mini kit (Qiagen, 40724 Hilden, Germany), following the manufacturer’s instructions. Positive (C. psittaci Loth strain) and negative (kit reagents only) extraction controls were included in each set of extraction. The DNA extracts were stored at −20 °C before analysis.
2.3. Real-Time PCRs
All DNA extracts were screened using a Chlamydiaceae-specific rtPCR targeting the 23S rRNA gene fragment [27]. An analytical cut-off value was selected at a cycle threshold (Ct) of 39.
All samples that gave a positive signal with the 23S-rtPCR were re-examined with in-house-specific enoA-based C. psittaci and enoA-based C. abortus rtPCRs, developed and already in use in Anses laboratory in order to improve the detection of C. psittaci and avian C. abortus strains compared to traditional methods. For the specific C. psittaci detection, we used enoA_CpsF43 5′-ATTCGCCCTATAGGTGCACAT-3′ and enoA_CpsR162 5′-GCCTTCATCTCCAACTCCTGTAG-3′ primers and the probe enoA_CpsP79 5′-[FAM] GTGCGTATGGGTGCTGATGTTT [BHQ1]-3′. For the specific detection of C. abortus, we used enoA_CabF13 5′-AACAACGGCCTGCAATTTCAAG-3′and enoA_CabR124 5′-TGAGAAGGTTTTTCAATGTATGGAAC-3′ primers, as well as the probe enoA_CabP93 5′-[FAM] GGCACCCATACGTACAGCTTCTTG [BHQ1]-3′. DNA amplification was performed in a final volume of 20 µL containing 10 µL of TaqManTM Fast Advanced Master Mix (Applied Biosystems, Waltham, MA, USA), 0.6 µM of each primer, 0.1 µM of the probe, 2 µL of DNA sample and water (qsp 20 µL). The reaction of rtPCR was carried out in 7500 or ViiA7 apparatus (Applied Biosystems, Waltham, MA, USA) using the following cycling parameters: 50 °C for 2 min, 95 °C for 20 s, 45 cycles of 95 °C for 3 s and 60 °C for 30 s. Specificity and sensitivity of in-house enoA-based C. psittaci and enoA-based C. abortus rtPCRs are shown in Table S1 and Figure S1, respectively.
2.4. Multilocus Sequence Typing (MLST) Analysis
The MLST analysis was performed on the C. abortus-positive samples, according to Pannekoek et al. [31,32]. Fragments of seven housekeeping genes, namely gatA, oppA3, hflX, gidA, enoA, hemN and fumC, were amplified and sequenced using the primers and conditions described on the Chlamydiales MLST website [33]. Sanger sequencing of both DNA strands was performed by Eurofins Genomics (85560 Ebersberg, Germany), and the numbers for alleles and the sequence type (ST) were assigned in accordance with the Chlamydiales MLST database and uploaded on the PubMLST website [33]. For each sample, DNA sequences of the seven alleles were manually assembled to obtain 3098 nucleotides. Multiple alignments of the seven concatenated MLST gene fragments with a large panel of C. psittaci and C. abortus strains were performed using the MEGA7 software [34]. Phylogenetic trees were constructed by the maximum likelihood method based on the general time-reversible model [34].
2.5. Detection of the Chlamydial Plasmid
The presence of the plasmid was investigated on the C. abortus-positive samples by a conventional PCR, using the in-house primer set (pCpsi_Fw 5′-AGCTGTGCATACATGGCTGT-3′ and pCpsi_Rv 5′-CAGTAACTGCGGTAGCTCGT-3′), targeting a 734-nucleotide region within the chlamydial plasmid tyrosine recombinase XerC gene harboured by the plasmid II of C. abortus strain 15-58d44 (GenBank Accession Number OU508368.1). DNA amplification was performed in a final volume of 25 µL containing 2 µL of DNA samples, 1× PCR reaction buffer, 1 U of Hot start Taq DNA polymerase (Qiagen, 40724 Hilden, Germany), 200 µM of each deoxynucleotide triphosphate (Promega, 20126 Milan, Italy) and 0.4 mM of each forward and reverse primers. The following cycling parameters were used: initial denaturation at 94 °C for 10 min, 40 cycles of 94 °C for 30 s, 50 °C for 30 s, 72 °C for 60 s, final extension at 72 °C for 7 min. DNA amplified fragments were sequenced by the Sanger method by Eurofins Genomics (85560 Ebersgerg, Germany) using the same primers. Nucleotide sequences of the plasmid DNA fragments were aligned and analysed in MEGA7 [34]. Phylogenetic trees were constructed by using the maximum likelihood method based on the general time-reversible model. Bootstrap tests were for 1000 repetitions.
2.6. PCR/High-Resolution Melting (HRM) Analysis
Genotyping of the C. psittaci-positive samples was undertaken via PCR/high-resolution melting (HRM), performed according to Vorimore et al. [35].
3. Results
3.1. Results of the 23S rtPCR
The results are shown in Table 1. The PCR targeting the 23S rRNA gene fragment showed cloacal shedding of Chlamydiaceae in 12 out of 108 (11.1%, 5.9%–18.6% 95% CI) birds sampled. The Chlamydiaceae prevalence was higher among hooded crows (9/38, 23.7%, 11.4%–40.2% 95% CI) than Eurasian magpies (2/52, 3.8%, 0.5%–13.2% 95% CI) and Eurasian jays (1/18, 5.6%, 0.1%–27.3% 95% CI). A mean Ct value of 35.1 (Ct range from 26.1 to 38.5) was observed.
3.2. Molecular Characterisation of Chlamydiaceae-Positive Samples
Eight out of twelve Chlamydiaceae-positive samples were characterised by the species-specific rtPCRs. Only one DNA sample from a hooded crow was positive for C. psittaci rtPCR. Interestingly, C. abortus DNA was detected in 5 of 9 (55.5%) and 2/2 (100%) Chlamydiaceae-positive hooded crow and Eurasian magpie samples, respectively.
No positive results were shown for C. avium, C. gallinacea, C. pecorum or C. suis.
With respect to the remaining four Chlamydiaceae-positive samples, the three samples from hooded crows showed a signal above the cut-off value of 39 when tested with rtPCR for C. abortus, while the sample from Eurasian jays showed no signal to specific rtPCRs.
Further attempts at characterisation of these samples by Chlamydiales 16S rRNA PCR or 23S rRNA PCR were unsuccessful, possibly due to the low amount of DNA.
3.3. Genotyping of C. abortus-Positive Samples by MLST and Plasmid Sequencing
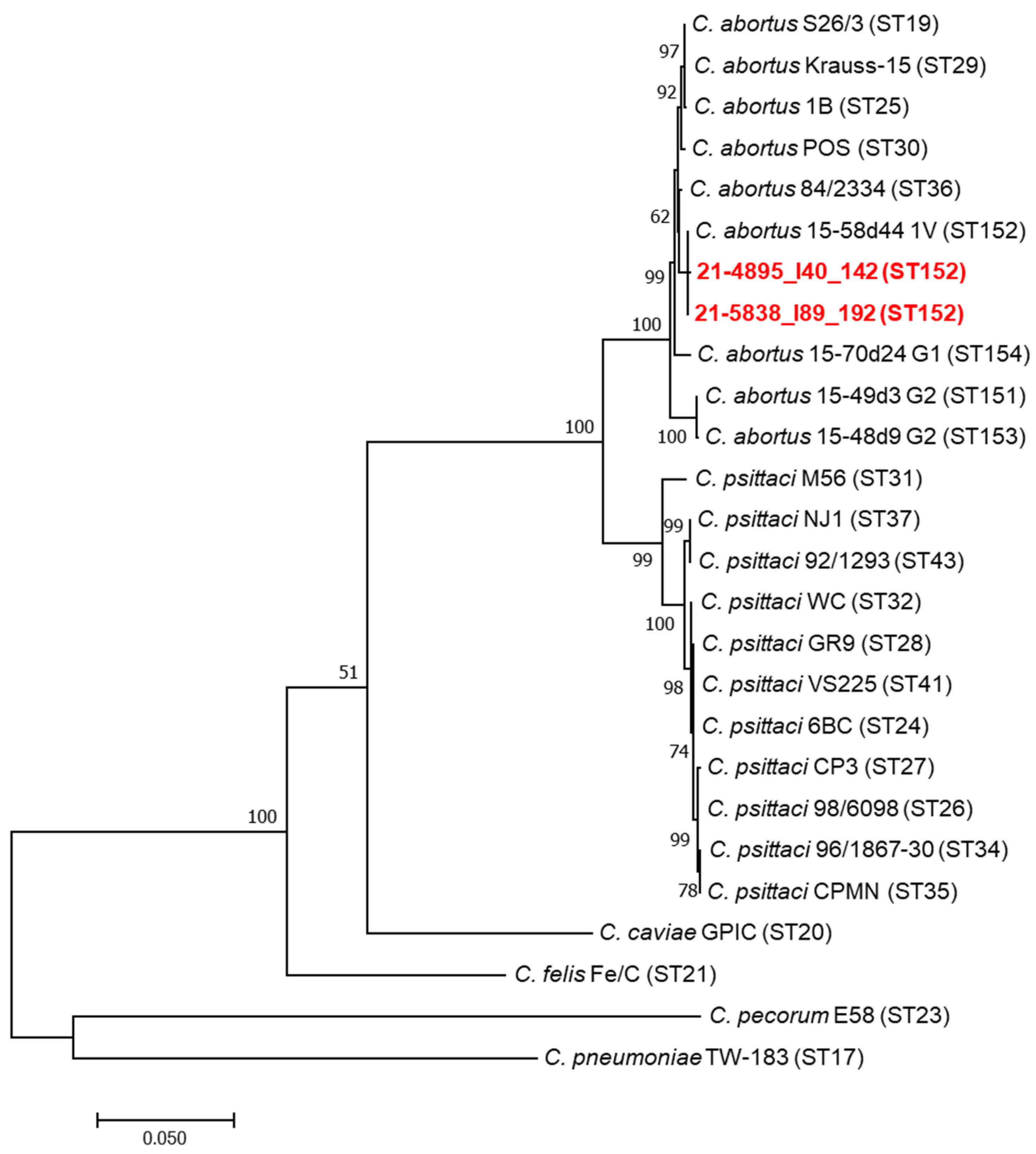
The MLST was successfully performed only on the two C. abortus-positive DNA samples from Eurasian magpies that had shown high levels of chlamydial excretion by rtPCR. The MLST sequences obtained from the two Eurasian magpie samples were identical and identified as ST152. Regarding the five hooded crow samples positive for C. abortus, for three of them, it was possible to amplify only three gene sequences (gidA, enoA, hemN) that were identical to those of the two Eurasian magpie samples. Regarding the remaining two hooded crow samples positive for C. abortus, the low quantity of DNA did not allow an appreciable MLST result. Comparative phylogenetic analysis of the concatenated MLST sequences of the two Eurasian magpie samples and a large panel of C. psittaci and C. abortus strains, including avian and ruminant C. abortus strains, showed a topology Identical to that of the avian C. abortus genotype 1V strain 15-58d44 recently detected in Poland [18] (Figure 1). MLST sequences were uploaded to the PubMLST database, and their ST allelic profile can be consulted [33]. The sequencing of the plasmid DNA fragment of the XerC gene was successfully performed for the two Eurasian magpie samples and two hooded crow samples, showing 100% identity (Eurasian magpie samples) and 99.9% similarity (hooded crow samples) with the same fragment of the 15-58d44 plasmid (Figure 2). The plasmid sequences obtained in this study were submitted to the GenBank database and are available under the following accession numbers: ON165250-ON165253.
3.4. Chlamydia psittaci Genotype Identification
The PCR/high-resolution melting performed on the C. psittaci-positive sample obtained from one hooded crow was consistent to the group III_pigeon, corresponding to the B genotype based on former ompA analysis.
4. Discussion
In Europe, recent decades have seen an adaptation of wild animal populations to specific conditions of the urban environment [36]. Corvids are resident or short-range migratory birds characterised by a generalist behaviour less demanding regarding the environment and the feed [37]. Thus, they have developed a marked synanthropic temperament, taking advantage of the presence of crops, waste and landfills, all of which are derived from human activities. These features are the basis of their successful adaptation to urban ecological niches and their increase, in contrast to the decrease in many bird species due to pollution or habitat modification/destruction. In Italy, for years, there has been an increase in the population levels of corvids, both in urban and rural environments. In the period 2000–2020, an average annual variation of 0.80% (±0.12) for hooded crows and 2.05% (±0.13) for Eurasian magpies has been registered. The conservation status was considered favourable for both avian species [38].
The Chlamydiaceae occurrence in the Corvidae family has been investigated in recent studies [18,20,24,39], showing prevalence values ranging from 5.1% to 29%.
In Italy, a previous investigation [24] performed on 76 corvids showed that 22/76 (29%) birds were PCR Chlamydia positive, with only one C. psittaci-positive sample vs. 21 C. suis-positive animals. The source of C. suis had been related to contact with wild boar, since the corvids sampled were from hilly areas where the presence of wild boar was consistently reported. Some samples showed mixed sequences of C. psittaci-C. abortus but were not included in the study due to the low amount of DNA, impeding further investigations (data not shown).
The results of the present study, performed in another geographical area, confirm Chlamydiaceae circulation in corvids. Chlamydia psittaci positivity was shown in only one DNA sample from a hooded crow, according to the low C. psitttaci prevalence previously detected [24]. Interestingly, the presence of avian C. abortus strains was detected in most of the Chlamydiaceae-positive samples (7/8, 87.5%) characterised at the species level. In addition, three Chlamydiaceae-positive samples from hooded crows reacted to C. abortus rtPCR, but they were not included in the prevalence calculation due to the higher signal cut-off value. The rtPCR results were confirmed by MLST analysis on the two strongest positive samples, allowing to group the two samples with avian C. abortus genotype 1V strain 15-58d44 [18]. To confirm the intermediate characteristics between C. psittaci and C. abortus, in four C. abortus-positive samples, it was possible to partly sequence the chlamydial plasmid inherent in most C. psittaci and avian C. abortus, but not in ruminant C. abortus strains. The fragment plasmid sequences were closely related (≥99%) to those of the Polish avian C. abortus genotype 1V strain 15-58d44 [18]. To our knowledge, this is the first report of avian C. abortus strains in Italy, specifically genotype 1V, which appear to be actively circulating in wild bird populations of the Veneto region, at least in the avian species considered in this study. So far, avian C. abortus strains have been reported in mallard (Anas platyrhynchos), swan (Cignus) and Eurasian teal (Anas crecca), as well as in Eurasian magpie and hooded crow, in Poland [18]; in rook (Corvus frugilegus) and Korean magpie (Pica sericea) in South Korea [39]; and in common buzzard (Buteo buteo) and carrion crow (Corvus corone), as well as in rook, in Switzerland [20]. Furthermore, some C. psittaci isolates from parrots and parakeets were shown to differ from classical avian C. psittaci strains and to be more closely related to C. abortus species [32,40]. Recently, the sequencing of the whole genome of one of these isolates allowed exploring its evolutionary relationship to both C. psittaci and C. abortus, supporting its reclassification as C. abortus species [41]. These acquisitions, if supported by further studies using next-generation sequencing, could suggest some changes in the taxonomy of the Chlamydiaceae family [2]. Considering that data on the prevalence of avian C. abortus strains, as well as their host and geographical distributions, are still limited, our results could supplement the current literature.
At the present state of knowledge, the zoonotic impact of the avian C. abortus strains has not been investigated; however, it cannot be excluded, considering their phylogenetic relationship with C. psittaci and ruminant C. abortus strains. The zoonotic role of C. psittaci has been known for a long time. Zoonotic transmission occurs by the inhalation of respiratory secretions or dried faeces dispersed in the air as fine droplets or dust particles, as well as through handling infected birds [42], particularly in high-risk individuals, such as veterinarians, bird breeders and pet-shop or poultry workers. In humans, symptoms range from mild illness to atypical pneumonia or to serious complications in internal organs [43]. Chlamydia abortus has recently been detected in a wide range of animals, so far mainly associated with enzootic abortions in small ruminants [44]. Most reported cases of human C. abortus infection involve pregnant women, initially showing an influenza-like illness with the consecutive development of placental dysfunction, leading to foetal death, as a result of direct or indirect contact with infected animals [45,46,47,48]. Extra-gestational infections of C. abortus manifested as pelvic inflammatory disease have also been described [49]. More recently, C. abortus was suggested to be the probable causative agent of atypical pneumonia detected in a veterinary researcher working in a laboratory where experimental intranasal infections with C. abortus were developed in sheep [50]. In conclusion, the circulation of a well-known zoonotic agent as C. psittaci and other potential zoonotic chlamydiae in corvid populations should be considered by workers in wildlife centres and people who may have professional or other contact with wild birds, who are urged to take appropriate preventive measures [51].
5. Conclusions
The results of this study on corvids confirm the Chlamydiaceae circulation previously detected in another Italian area [24]. Interestingly, C. psittaci was detected in only one sample, whereas most of the positive samples showed high molecular similarity with avian C. abortus strains, specifically genotype 1V. Further investigations involving more Italian geographical areas and more wild avian species are needed, to confirm and deepen these results.
Moreover, considering the constant increase of the populations of corvids in urban/peri-urban areas and the consequent possibility of human professional and non-professional contacts, the potential zoonotic role of the avian C. abortus strains must be further investigated.
Supplementary Materials
The following supporting information can be downloaded at https://www.mdpi.com/article/10.3390/ani12101226/s1, Table S1: Chlamydia species used to test the specificity of the enoA-based C. psittaci and enoA-based C. abortus rtPCRs. Figure S1: Real-time PCR sensitivity test to detect the genomic DNA from Loth C. psittaci isolate (A) and S26/3 C. abortus strain (B).
Author Contributions
Conceptualisation, R.A., K.L., F.G., C.L. and A.D.F.; methodology, K.L., R.A. and A.D.F.; software, R.A.; validation, R.A. and K.L.; formal analysis, R.A. and A.D.F.; investigation, R.A., F.G., D.S. and C.S.; resources, K.L., C.T., C.L. and A.D.F.; data curation, R.A. and A.D.F.; writing—original draft preparation, R.A. and A.D.F.; writing—review and editing, K.L., R.A., F.G., C.S. and A.D.F.; visualisation, R.A. and A.D.F.; supervision, R.A., K.L. and A.D.F. All authors have read and agreed to the published version of the manuscript.
Funding
This research received no external funding.
Institutional Review Board Statement
Ethical review and approval were waived for this study because the samples examined were taken from dead birds.
Informed Consent Statement
Not applicable.
Data Availability Statement
The plasmid sequences generated in this study are available in GenBank under Accession Numbers ON165250-ON165253; ST allelic profile of MLST sequences can be consulted on the PubMLST database.
Acknowledgments
The authors are grateful to Salvatore Catania from SCT1-Verona, Istituto Zooprofilattico Sperimentale delle Venezie for the aid in collection of samples.
Conflicts of Interest
The authors declare no conflict of interest.
References
- Borel, N.; Polkinghorne, A.; Pospischil, A. A review on chlamydial diseases in animals: Still a challenge for pathologists? Vet. Pathol. 2018, 55, 374–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zaręba-Marchewka, K.; Szymańska-Czerwińska, M.; Niemczuk, K. Chlamydiae—What’s New? J. Vet. Res. 2020, 64, 461–467. [Google Scholar] [CrossRef] [PubMed]
- Sachse, K.; Bavoil, P.M.; Kaltenboeck, B.; Stephens, R.S.; Kuo, C.C.; Rossello-Mora, R.; Horn, M. Emendation of the family Chlamydiaceae: Proposal of a single genus, Chlamydia, to include all currently recognized species. Syst. Appl. Microbiol. 2015, 38, 99–103. [Google Scholar] [CrossRef] [PubMed]
- Staub, E.; Marti, H.; Biondi, R.; Levi, A.; Donati, M.; Leonard, C.A.; Ley, S.D.; Pillonel, T.; Greub, G.; Seth-Smith, H.M.B.; et al. Novel Chlamydia species isolated from snakes are temperature-sensitive and exhibit decreased susceptibility to azithromycin. Sci. Rep. 2018, 8, 5660. [Google Scholar] [CrossRef] [Green Version]
- Laroucau, K.; Vorimore, F.; Aaziz, R.; Solmonson, L.; Hsia, R.C.; Bavoil, P.M.; Fach, P.; Hölzer, M.; Wuenschmann, A.; Sachse, K. Chlamydia buteonis, a new Chlamydia species isolated from a red-shouldered hawk. Syst. Appl. Microbiol. 2019, 42, 125997. [Google Scholar] [CrossRef]
- Vorimore, F.; Hsia, R.C.; Huot-Creasy, H.; Bastian, S.; Deruyter, L.; Passet, A.; Sachse, K.; Bavoil, P.; Myers, G.; Laroucau, K. Isolation of a new Chlamydia species from the Feral Sacred Ibis (Threskiornis aethiopicus): Chlamydia ibidis. PLoS ONE 2013, 8, e74823. [Google Scholar] [CrossRef] [Green Version]
- Taylor-Brown, A.; Bachmann, N.L.; Borel, N.; Polkinghorne, A. Culture-independent genomic characterization of Candidatus Chlamydia sanzinia, a novel uncultivated bacterium infecting snakes. BMC Genom. 2016, 17, 710. [Google Scholar] [CrossRef] [Green Version]
- Taylor-Brown, A.; Spang, L.; Borel, N.; Polkinghorne, A. Culture-independent metagenomics supports discovery of uncultivable bacteria within the genus Chlamydia. Sci. Rep. 2017, 7, 10661. [Google Scholar] [CrossRef] [Green Version]
- Laroucau, K.; Ortega, N.; Vorimore, F.; Aaziz, R.; Mitura, A.; Szymanska-Czerwinska, M.; Cicerol, M.; Salinas, J.; Sachse, K.; Caro, M.R. Detection of a novel Chlamydia species in captive spur-thighed tortoises (Testudo graeca) in southeastern Spain and proposal of Candidatus Chlamydia testudinis. Syst. Appl. Microbiol. 2020, 43, 126071. [Google Scholar] [CrossRef]
- Vorimore, F.; Hölzer, M.; Liebler-Tenorio, E.M.; Barf, L.M.; Delannoy, S.; Vittecoq, M.; Wedlarski, R.; Lécu, A.; Scharf, S.; Blanchard, Y.; et al. Evidence for the existence of a new genus Chlamydiifrater gen. nov. inside the family Chlamydiaceae with two new species isolated from flamingo (Phoenicopterus roseus): Chlamydiifrater phoenicopteri sp. nov. and Chlamydiifrater volucris sp. nov. Syst. Appl. Microbiol. 2021, 44, 126200. [Google Scholar] [CrossRef]
- Kaleta, E.F.; Taday, E.M. Avian host range of Chlamydophila spp. based on isolation, antigen detection and serology. Avian Pathol. 2003, 32, 435–461. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Andersen, A.A. Serotyping of Chlamydia psittaci isolates using serovar-specific monoclonal antibodies with the microimmunofluorescence test. J. Clin. Microbial. 1991, 29, 707–711. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanrompay, D.; Andersen, A.A.; Ducatelle, R.; Haesebrouck, F. Serotyping of European isolates of Chlamydia psittaci from poultry and other birds. J. Clin. Microbiol. 1993, 31, 134–137. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vanrompay, D.; Butaye, P.; Sayada, C.; Ducatelle, R.; Haesebrouck, F. Characterization of avian Chlamydia psittaci strains using omp1 restriction mapping and serovar-specific monoclonal antibodies. Res. Microbiol. 1997, 148, 327–333. [Google Scholar] [CrossRef]
- Geens, T.; Desplanques, A.; Van Loock, M.; Bönner, B.M.; Kaleta, E.F.; Magnino, S.; Andersen, A.A.; Everett, K.D.E.; Vanrompay, D. Sequencing of the Chlamydophila psittaci ompA gene reveals a new genotype, E/B, and the need for a rapid discriminatory genotyping method. J. Clin. Microbiol. 2005, 43, 2456–2461. [Google Scholar] [CrossRef] [Green Version]
- Sachse, K.; Laroucau, K.; Hotzel, H.; Schubert, E.; Ehricht, R.; Slickers, P. Genotyping of Chlamydophila psittaci using a new DNA microarray assay based on sequence analysis of ompA genes. BMC Microbiol. 2008, 8, 63. [Google Scholar] [CrossRef] [Green Version]
- Madani, S.A.; Peighambari, S.M. PCR-based diagnosis, molecular characterization and detection of atypical strains of avian Chlamydia psittaci in companion and wild birds. Avian Pathol. 2013, 42, 38–44. [Google Scholar] [CrossRef] [Green Version]
- Szymańska-Czerwińska, M.; Mitura, A.; Niemczuk, K.; Zaręba, K.; Jodełko, A.; Pluta, A.; Scharf, S.; Vitek, B.; Aaziz, R.; Vorimore, F.; et al. Dissemination and genetic diversity of chlamydial agents in Polish wildfowl: Isolation and molecular characterisation of avian Chlamydia abortus strains. PLoS ONE 2017, 12, e0174599. [Google Scholar] [CrossRef] [Green Version]
- Luján-Vega, C.; Hawkins, M.G.; Johnson, C.K.; Briggs, C.; Vennum, C.; Bloom, P.H.; Hull, J.M.; Cray, C.; Pesti, D.; Johnson, L.; et al. Atypical Chlamydiaceae in wild populations of hawks (Buteo spp.) in California. J. Zoo Wildl. Med. 2018, 49, 108–115. [Google Scholar] [CrossRef]
- Stalder, S.; Marti, H.; Borel, N.; Sachse, K.; Albini, S.; Vogler, B.R. Occurrence of Chlamydiaceae in Raptors and Crows in Switzerland. Pathogens 2020, 9, 724. [Google Scholar] [CrossRef]
- Sachse, K.; Laroucau, K.; Riege, K.; Wehner, S.; Dilcher, M.; Creasy, H.H.; Weidmann, M.; Myers, G.; Vorimore, F.; Vicari, N.; et al. Evidence for the existence of two new members of the family Chlamydiaceae and proposal of Chlamydia avium sp. nov. and Chlamydia gallinacea sp. nov. Syst. Appl. Microbiol. 2014, 37, 79–88. [Google Scholar] [CrossRef] [PubMed]
- Pantchev, A.; Sting, R.; Bauerfeind, R.; Tyczka, J.; Sachse, K. New real-time PCR tests for species-specific detection of Chlamydophila psittaci and Chlamydophila abortus from tissue samples. Vet. J. 2009, 181, 145–150. [Google Scholar] [CrossRef] [PubMed]
- Sachse, K.; Kuehlewind, S.; Ruettger, A.; Schubert, E.; Rohde, G. More than classical Chlamydia psittaci in urban pigeons. Vet. Microbiol. 2012, 157, 476–480. [Google Scholar] [CrossRef] [PubMed]
- Di Francesco, A.; Donati, M.; Laroucau, K.; Balboni, A.; Galuppi, R.; Merialdi, G.; Salvatore, D.; Renzi, M. Chlamydiae in corvids. Vet. Rec. 2015, 177, 466. [Google Scholar] [CrossRef] [PubMed]
- Frutos, M.C.; Monetti, M.S.; Gallo Vaulet, L.; Cadario, M.E.; Rodríguez Fermepin, M.; Ré, V.E.; Cuffini, C.G. Genetic diversity of Chlamydia among captive birds from central Argentina. Avian Pathol. 2015, 44, 50–56. [Google Scholar] [CrossRef]
- Guo, W.; Li, J.; Kaltenboeck, B.; Gong, J.; Fan, W.; Wang, C. Chlamydia gallinacea, not C. psittaci, is the endemic chlamydial species in chicken (Gallus gallus). Sci. Rep. 2016, 6, 19638. [Google Scholar] [CrossRef]
- Ehricht, R.; Slickers, P.; Goellner, S.; Hotzel, H.; Sachse, K. Optimized DNA microarray assay allows detection and genotyping of single PCR-amplifiable target copies. Mol. Cell. Probes 2006, 20, 60–63. [Google Scholar] [CrossRef]
- Laroucau, K.; Aaziz, R.; Meurice, L.; Servas, V.; Chossat, I.; Royer, H.; de Barbeyrac, B.; Vaillant, V.; Moyen, J.L.; Meziani, F.; et al. Outbreak of psittacosis in a group of women exposed to Chlamydia psittaci-infected chickens. Eur. Surveill. 2015, 20, 21155. [Google Scholar] [CrossRef] [Green Version]
- Zocevic, A.; Vicari, N.; Gasparini, J.; Jacquin, L.; Sachese, K.; Magnino, S.; Laroucau, K. A real-time PCR assay for the detection of atypical strains of Chlamydiaceae from pigeons. PLoS ONE 2013, 8, e58741. [Google Scholar] [CrossRef] [Green Version]
- Pantchev, A.; Sting, R.; Bauerfeind, R.; Tyczka, J.; Sachse, K. Detection of all Chlamydophila and Chlamydia spp. of veterinary interest using species-specific real-time PCR assays. Comp. Immunol. Microbiol. Infect. Dis. 2010, 33, 473–484. [Google Scholar] [CrossRef]
- Pannekoek, Y.; Morelli, G.; Kusecek, B.; Morre, S.A.; Ossewaarde, J.M.; Langerak, A.A.; van der Ende, A. Multi locus sequence typing of Chlamydiales: Clonal groupings within the obligate intracellular bacteria Chlamydia trachomatis. BMC Microbiol. 2008, 8, 42. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pannekoek, Y.; Dickx, V.; Beeckman, D.S.A.; Jolley, K.A.; Keijzers, W.C.; Vretou, E.; Maiden, M.C.; Vanrompay, D.; van der Ende, A. Multi locus sequence typing of Chlamydia reveals an association between Chlamydia psittaci genotypes and host species. PLoS ONE 2010, 5, e14179. [Google Scholar] [CrossRef] [PubMed]
- Chlamydiales spp. Available online: https://pubmlst.org/organisms/Chlamydiales-spp (accessed on 22 November 2021).
- Kumar, S.; Stecher, G.; Tamura, K. MEGA7: Molecular Evolutionary Genetics Analysis Version 7.0 for Bigger Datasets. Mol. Biol. Evol. 2016, 33, 1870–1874. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Vorimore, F.; Aaziz, R.; de Barbeyrac, B.; Peuchant, O.; Szymańska-Czerwińska, M.; Herrmann, B.; Schnee, C.; Laroucau, K. A new SNP-based genotyping method for C. psittaci: Application to field samples for quick identification. Microorganisms 2021, 9, 625. [Google Scholar] [CrossRef] [PubMed]
- Luniak, M. Synurbization—Adaptation of Animal Wildlife to Urban Development. In Proceedings 4th International Urban Wildlife Symposium; Shaw, W.W., Harris, L.K., VanDruff, L., Eds.; 2004; Available online: https://cals.arizona.edu (accessed on 18 April 2022).
- Miller, R.; Schiestl, M.; Whiten, A.; Schwab, C.; Bugnyar, T. Tolerance and Social Facilitation in the Foraging Behaviour of Free-Ranging Crows (Corvus corone corone; C. c. cornix). Ethology 2014, 120, 1248–1255. [Google Scholar] [CrossRef] [Green Version]
- Rete Rurale Nazione & LIPU. Uccelli Comuni delle Zone Agricole in Italia. Aggiornamento degli Andamenti di Popolazione e del Farmland Bird Index per la Rete Rurale Nazionale. 2020. Available online: https://www.reterurale.it/ (accessed on 11 February 2022).
- Jeong, J.; An, I.; Oem, J.K.; Wang, S.J.; Kim, Y.; Shin, J.-H.; Woo, C.; Kim, Y.; Jo, S.-D.; Son, K.; et al. Molecular prevalence and genotyping of Chlamydia spp. in wild birds from South Korea. J. Vet. Med. Sci. 2017, 79, 1204–1209. [Google Scholar] [CrossRef] [Green Version]
- Van Loock, M.; Vanrompay, D.; Herrmann, B.; Vander Stappen, J.; Volckaert, G.; Goddeeris, B.M.; Everett, K.D.E. Missing links in the divergence of Chlamydophila abortus from Chlamydophila psittaci. Int. J. Syst. Evol. Microbiol. 2003, 53, 761–770. [Google Scholar] [CrossRef]
- Longbottom, D.; Livingstone, M.; Ribeca, P.; Beeckman, D.S.A.; van der Ende, A.; Pannekoek, Y.; Vanrompay, D. Whole genome de novo sequencing and comparative genomic analyses suggests that Chlamydia psittaci strain 84/2334 should be reclassified as Chlamydia abortus species. BMC Genom. 2021, 22, 159. [Google Scholar] [CrossRef]
- Harkinezhad, T.; Geens, T.; Vanrompay, D. Chlamydophila psittaci infections in birds: A review with emphasis on zoonotic consequences. Vet. Microbiol. 2009, 135, 68–77. [Google Scholar] [CrossRef]
- Rohde, G.; Straube, E.; Essig, A.; Reinhold, P.; Sachse, K. Chlamydial zoonoses. Dtsch Arztebl. Int. 2010, 107, 174–180. [Google Scholar] [CrossRef]
- Cheong, H.C.; Lee, C.Y.Q.; Cheok, Y.Y.; Tan, G.M.Y.; Looi, C.Y.; Wong, W.F. Chlamydiaceae: Diseases in primary hosts and zoonosis. Microorganisms 2019, 7, 146. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pospischil, A.; Thoma, R.; Hilbe, M.; Grest, P.; Gebbers, J.O. Abortion in woman caused by caprine Chlamydophila abortus (Chlamydia psttaci serovar 1). Swiss Med. Wkly. 2002, 132, 64–66. [Google Scholar] [PubMed]
- Meijer, A.; Brandenburg, A.; de Vries, J.; Beentjes, J.; Roholl, P.; Dercksen, D. Chlamydophila abortus infection in a pregnant woman associated with indirect contact with infected goats. Eur. J. Clin. Microbiol. Infect. Dis. 2004, 23, 487–490. [Google Scholar] [CrossRef] [PubMed]
- Walder, G.; Hotzel, H.; Brezinka, C.; Gritsch, W.; Tauber, R.; Würzner, R.; Ploner, F. An unusual cause of sepsis during pregnancy: Recognizing infection with Chlamydophila abortus. Obstet. Gynecol. 2005, 106, 1215–1217. [Google Scholar] [CrossRef]
- Pichon, N.; Guindre, L.; Laroucau, K.; Cantaloube, M.; Nallatamby, A.; Parreau, S. Chlamydia abortus in pregnant woman with acute respiratory distress syndrome. Emerg. Infect. Dis. 2020, 26, 628–629. [Google Scholar] [CrossRef] [Green Version]
- Walder, G.; Meusburger, H.; Hotzel, H.; Oehme, A.; Neunteufel, W.; Dierich, M.P.; Wurzner, R. Chlamydophila abortus pelvic inflammatory disease. Emerg. Infect. Dis. 2003, 9, 1642–1644. [Google Scholar] [CrossRef]
- Ortega, N.; Caro, M.R.; Carmen Gallego, M.; Murcia-Belmonte, A.; Álvarez, D.; del Río, L.; Cuello, F.; Buendía, A.J.; Salinas, J. Isolation of Chlamydia abortus from a laboratory worker diagnosed with atypical pneumonia. Irish Vet.J. 2016, 69, 8. [Google Scholar] [CrossRef] [Green Version]
- Tolba, H.M.N.; Abou Elez, R.M.M.; Elsohaby, I. Risk factors associated with Chlamydia psittaci infections in psittacine birds and bird handlers. J. Appl. Microbiol. 2019, 126, 402–410. [Google Scholar] [CrossRef]
Figure 1.
Phylogenetic analyses of multilocus sequence typing (MLST) concatenated sequences of Chlamydia. Concatenated sequences (3098 nucleotides) were aligned and analysed in MEGA7. Phylogenetic trees were constructed by using the maximum likelihood method based on the general time-reversible model. Bootstrap tests were for 1000 repetitions. Numbers on tree nodes indicate bootstrap values of the main branches. Horizontal line scale indicates the number of nucleotide substitutions per site. The MLST sequence type (ST) is indicated. The red colour represents the avian C. abortus strains analysed in this study.
Figure 1.
Phylogenetic analyses of multilocus sequence typing (MLST) concatenated sequences of Chlamydia. Concatenated sequences (3098 nucleotides) were aligned and analysed in MEGA7. Phylogenetic trees were constructed by using the maximum likelihood method based on the general time-reversible model. Bootstrap tests were for 1000 repetitions. Numbers on tree nodes indicate bootstrap values of the main branches. Horizontal line scale indicates the number of nucleotide substitutions per site. The MLST sequence type (ST) is indicated. The red colour represents the avian C. abortus strains analysed in this study.

Figure 2.
Phylogenetic analyses of plasmid XerC gene fragment sequences of Chlamydia. Nucleotide sequences (734 nucleotides) were aligned and analysed in MEGA7. Phylogenetic trees were constructed by using the maximum likelihood method based on the general time-reversible model. Bootstrap tests were for 1000 repetitions. Numbers on tree nodes indicate bootstrap values of the main branches. Horizontal line scale indicates the number of nucleotide substitutions per site. The outgroup is represented by C. caviae plasmid DNA fragment. Accession numbers of different Chlamydia are indicated. The red colour represents the avian C. abortus strains analysed in this study.
Figure 2.
Phylogenetic analyses of plasmid XerC gene fragment sequences of Chlamydia. Nucleotide sequences (734 nucleotides) were aligned and analysed in MEGA7. Phylogenetic trees were constructed by using the maximum likelihood method based on the general time-reversible model. Bootstrap tests were for 1000 repetitions. Numbers on tree nodes indicate bootstrap values of the main branches. Horizontal line scale indicates the number of nucleotide substitutions per site. The outgroup is represented by C. caviae plasmid DNA fragment. Accession numbers of different Chlamydia are indicated. The red colour represents the avian C. abortus strains analysed in this study.


{kind=link}
{kind=link}
Table 1.
Total number and percentage of Chlamydiaceae-positive corvids per species and number and percentage of chlamydial species identified.
Table 1.
Total number and percentage of Chlamydiaceae-positive corvids per species and number and percentage of chlamydial species identified.
| Corvid Species | Samples | 23S rtPCR | C. psittaci rtPCR | C. abortus rtPCR | C. gallinacea rtPCR | C. avium rtPCR | C. pecorum rtPCR | C. suis rtPCR | Non-Classified Chlamydia |
|---|---|---|---|---|---|---|---|---|---|
| Eurasian magpie | 52 | 2 (3.8%) | - | 2 (3.8%) | - | - | - | - | |
| Hooded crow | 38 | 9 (23.7%) | 1 (2.6%) | 5 (13.1%) | - | - | - | - | 3 (7.9%) |
| Eurasian jays | 18 | 1 (5.6%) | - | - | - | - | - | - | 1 (5.6%) |
| Total | 108 | 12 (11.1%) | 1 (0.9%) | 7 (6.5%) | - | - | - | - | 4 (3.7%) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2022 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
MDPI and ACS Style
Aaziz, R.; Laroucau, K.; Gobbo, F.; Salvatore, D.; Schnee, C.; Terregino, C.; Lupini, C.; Di Francesco, A. Occurrence of Chlamydiae in Corvids in Northeast Italy. Animals 2022, 12, 1226. https://doi.org/10.3390/ani12101226
AMA Style
Aaziz R, Laroucau K, Gobbo F, Salvatore D, Schnee C, Terregino C, Lupini C, Di Francesco A. Occurrence of Chlamydiae in Corvids in Northeast Italy. Animals. 2022; 12(10):1226. https://doi.org/10.3390/ani12101226
Chicago/Turabian StyleAaziz, Rachid, Karine Laroucau, Federica Gobbo, Daniela Salvatore, Christiane Schnee, Calogero Terregino, Caterina Lupini, and Antonietta Di Francesco. 2022. "Occurrence of Chlamydiae in Corvids in Northeast Italy" Animals 12, no. 10: 1226. https://doi.org/10.3390/ani12101226
Note that from the first issue of 2016, this journal uses article numbers instead of page numbers. See further details here.