
Characterization and Comparison of Enterococcus spp. Isolates from Feces of Healthy Dogs and Urine of Dogs with UTIs

 , , , and
, , , and
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Bacteria Strains Provenience and Identification
2.2. Phenotypic Resistance
2.3. Genotypic Resistance
2.4. Biofilm Production
2.5. Virulence Genes Detection
2.6. Clustering Analysis
2.7. Statistical Analysis
3. Results
3.1. Bacteria Strains, Provenience and Identification
3.2. Phenotypic Resistance
3.3. Genotypic Resistance
3.4. Resistance Profiles: Comparison of Phenotypic and Genotypic Results
3.5. Biofilm Production
3.6. Virulence Genes and Virulence Profiles
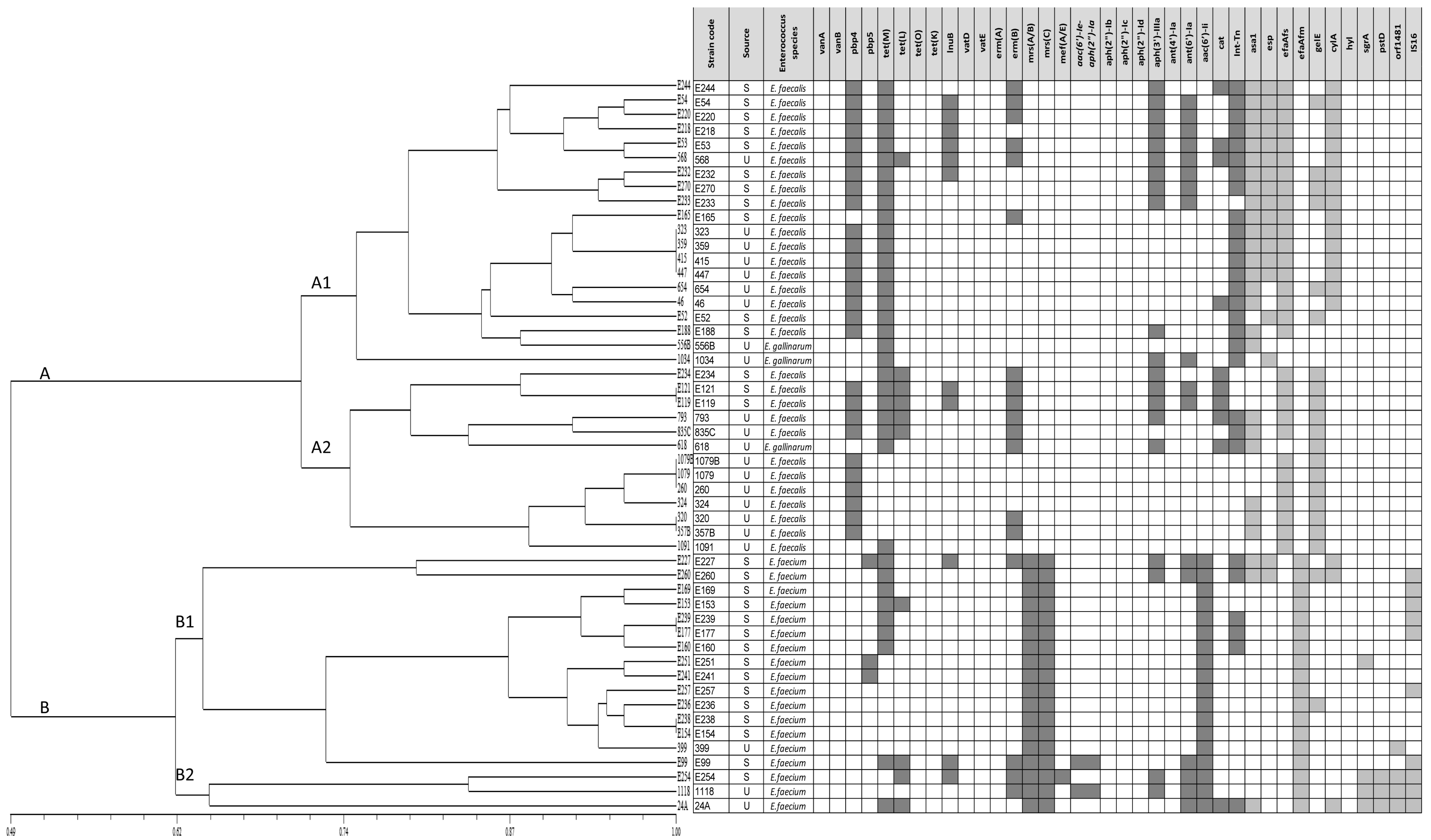
3.7. Clustering Analysis of Relevant Safety Traits
4. Discussion
5. Conclusions
Supplementary Materials
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Ramos, S.; Silva, V.; de Lurdes Enes Dapkevicius, M.; Igrejas, G.; Poeta, P. Enterococci, from harmless bacteria to a pathogen. Microorganisms 2020, 8, 1118. [Google Scholar] [CrossRef]
- Selleck, E.M.; Van Tyne, D.; Gilmore, M.S. Pathogenicity of Enterococci. Microbiol. Spectr. 2019, 7. [Google Scholar] [CrossRef]
- Kubašová, I.; Strompfová, V.; Lauková, A. Safety assessment of commensal enterococci from dogs. Folia Microbiol. 2017, 62, 491–498. [Google Scholar] [CrossRef]
- Torres, C.; Alonso, C.A.; Ruiz-Ripa, L.; León-Sampedro, R.; del Campo, R.; Coque, T.M. Antimicrobial Resistance in Enterococcus spp. of animal origin. Antimicrob. Resist. Bact. Livest. Companion Anim. 2018, 6, 185–227. [Google Scholar] [CrossRef]
- Miller, W.R.; Murray, B.E.; Rice, L.B.; Arias, C.A. Resistance in Vancomycin-Resistant Enterococci. Infect. Dis. Clin. N. Am. 2020, 34, 751–771. [Google Scholar] [CrossRef] [PubMed]
- Kristich, C.J.; Rice, L.B.; Arias, C.A. Enterococcal Infection—Treatment and Antibiotic Resistance. In Enterococci: From Commensals to Leading Causes of Drug Resistant Infection; Gilmore, M., Clewell, D., Ike, Y., Shankar, N., Eds.; Massachusetts Eye and Ear Infirmary: Boston, MA, USA, 2014. [Google Scholar]
- Pillay, S.; Zishiri, O.T.; Adeleke, M.A. Prevalence of virulence genes in enterococcus species isolated from companion animals and livestock. Onderstepoort J. Vet. Res. 2018, 85, 1–8. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bertelloni, F.; Salvadori, C.; Lotti, G.; Cerri, D.; Ebani, V.V. Antimicrobial resistance in enterococcus strains isolated from healthy domestic dogs. Acta Microbiol. Immunol. Hung. 2017, 64, 203–321. [Google Scholar] [CrossRef] [Green Version]
- Bondi, M.; Iseppi, R.; Messi, P.; Anacarso, I.; Sabia, C.; Condò, C.; De Niederhausern, S. Antimicrobial resistance and virulence traits in Enterococcus strains isolated from dogs and cats. New Microbiol. 2015, 38, 369–378. [Google Scholar]
- Van den Bunt, G.; Top, J.; Hordijk, J.; de Greeff, S.C.; Mughini-Gras, L.; Corander, J.; van Pelt, W.; Bonten, M.J.M.; Fluit, A.C.; Willems, R.J.L. Intestinal carriage of ampicillin- and vancomycin-resistant Enterococcus faecium in humans, dogs and cats in the Netherlands. J. Antimicrob. Chemother. 2018, 73, 607–614. [Google Scholar] [CrossRef]
- Oguttu, J.W.; Qekwana, D.N.; Odoi, A. Prevalence and Predictors of Antimicrobial Resistance among Enterococcus spp. from Dogs Presented at a Veterinary Teaching Hospital, South Africa. Front. Vet. Sci. 2021, 7, 589439. [Google Scholar] [CrossRef]
- Trościańczyk, A.; Nowakiewicz, A.; Gnat, S.; Łagowski, D.; Osińska, M. Are dogs and cats a reservoir of resistant and virulent Enterococcus faecalis strains and a potential threat to public health? J. Appl. Microbiol. 2021, 131, 2061–2071. [Google Scholar] [CrossRef]
- Sykes, J.E. Streptococcal and enterococcal infections. In Canine and Feline Infectious Diseases; Sykes, J.E., Ed.; Elsevier Inc.: St. Louis, MO, USA, 2014; pp. 334–346. ISBN 9781437707953. [Google Scholar]
- Byron, J.K. Urinary Tract Infection. Vet. Clin. N. Am.—Small Anim. Pract. 2019, 49, 211–221. [Google Scholar] [CrossRef]
- Thompson, M.F.; Litster, A.L.; Platell, J.L.; Trott, D.J. Canine bacterial urinary tract infections: New developments in old pathogens. Vet. J. 2011, 190, 22–27. [Google Scholar] [CrossRef]
- Olin, S.J.; Bartges, J.W. Urinary Tract Infections. Treatment/Comparative Therapeutics. Vet. Clin. N. Am.—Small Anim. Pract. 2015, 45, 721–746. [Google Scholar] [CrossRef] [PubMed]
- Scarborough, R.; Bailey, K.; Galgut, B.; Williamson, A.; Hardefeldt, L.; Gilkerson, J.; Browning, G. Use of local antibiogram data and antimicrobial importance ratings to select optimal empirical therapies for urinary tract infections in dogs and cats. Antibiotics 2020, 9, 924. [Google Scholar] [CrossRef] [PubMed]
- Yamanaka, A.R.; Hayakawa, A.T.; Rocha, Í.S.M.; Dutra, V.; Souza, V.R.F.; Cruz, J.N.; Camargo, L.M.; Nakazato, L. The occurrence of multidrug resistant bacteria in the urine of healthy dogs and dogs with cystitis. Animals 2019, 9, 1087. [Google Scholar] [CrossRef] [Green Version]
- Yu, Z.; Wang, Y.; Chen, Y.; Huang, M.; Wang, Y.; Shen, Z.; Xia, Z.; Li, G. Antimicrobial resistance of bacterial pathogens isolated from canine urinary tract infections. Vet. Microbiol. 2020, 241, 108540. [Google Scholar] [CrossRef]
- Hernando, E.; Vila, A.; D’Ippolito, P.; Rico, A.J.; Rodon, J.; Roura, X. Prevalence and Characterization of Urinary Tract Infection in Owned Dogs and Cats From Spain. Top. Companion Anim. Med. 2021, 43, 100512. [Google Scholar] [CrossRef] [PubMed]
- Stępień-Pyśniak, D.; Hauschild, T.; Różański, P.; Marek, A. MALDI-TOF mass spectrometry as a useful tool for identification of Enterococcus spp. From wild birds and differentiation of closely related species. J. Microbiol. Biotechnol. 2017, 27, 1128–1137. [Google Scholar] [CrossRef]
- Kariyama, R.; Mitsuhata, R.; Chow, J.W.; Clewell, D.B.; Kumon, H. Simple and reliable multiplex PCR assay for surveillance isolates of vancomycin-resistant enterococci. J. Clin. Microbiol. 2000, 38, 3092–3095. [Google Scholar] [CrossRef] [Green Version]
- Jackson, C.R.; Fedorka-Cray, P.J.; Barrett, J.B. Use of a genus- and species-specific multiplex PCR for identification of enterococci. J. Clin. Microbiol. 2004, 42, 3558–3565. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Clinical and Laboratory Standards Institute. M02-A12—Performance Standards for Antimicrobial Disk Susceptibility Tests. In Approved Standard, 12th ed.; Clinical & Laboratory Standards Institute: Wayne, PA, USA, 2015; pp. 1–96. [Google Scholar]
- EUCAST (The European Committee on Antimicrobial Susceptibility Testing). Breakpoint Tables for Interpretation of MICs and Zone Diameters. Version 10.0, 2020; The European Committee on Antimicrobial Susceptibility Testing: Basel, Switzerland, 2020. [Google Scholar]
- CLSI (Clinical and Laboratory Standards Institute). M100 Performance Standards for Antimicrobial Susceptibility Testing A CLSI Supplement for Global Application, 28th ed.; CLSI Supplement M100: Clinical and Laboratory Standards Institute, Wayne, PA, USA, 2018. [Google Scholar]
- CLSI (Clinical and Laboratory; Standards Institute). M07-A10 Methods for Dilution Antimicrobial Susceptibility Tests for Bacteria That Grow Aerobically. In Approved Standard, 10th ed.; CLSI Document M07-A10: Clinical and Laboratory Standards Institute, Wayne, PA, USA, 2015. [Google Scholar]
- Magiorakos, A.P.; Srinivasan, A.; Carey, R.B.; Carmeli, Y.; Falagas, M.E.; Giske, C.G.; Harbarth, S.; Hindler, J.F.; Kahlmeter, G.; Olsson-Liljequist, B.; et al. Multidrug-resistant, extensively drug-resistant and pandrug-resistant bacteria: An international expert proposal for interim standard definitions for acquired resistance. Clin. Microbiol. Infect. 2012, 18, 268–281. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Hashem, Y.A.; Abdelrahman, K.A.; Aziz, R.K. Phenotype–Genotype Correlations and Distribution of Key Virulence Factors in Enterococcus faecalis Isolated from Patients with Urinary Tract Infections. Infect. Drug Resist. 2021, 14, 1713–1723. [Google Scholar] [CrossRef] [PubMed]
- Bertelloni, F.; Cagnoli, G.; Ebani, V.V. Virulence and antimicrobial resistance in canine staphylococcus spp. Isolates. Microorganisms 2021, 9, 515. [Google Scholar] [CrossRef] [PubMed]
- Wood, M.W.; Lepold, A.; Tesfamichael, D.; Lasarev, M.R. Risk factors for enterococcal bacteriuria in dogs: A retrospective study. J. Vet. Intern. Med. 2020, 34, 2447–2453. [Google Scholar] [CrossRef]
- KuKanich, K.S.; Lubbers, B.V. Review of enterococci isolated from canine and feline urine specimens from 2006 to 2011. J. Am. Anim. Hosp. Assoc. 2015, 51, 148–154. [Google Scholar] [CrossRef]
- Darwich, L.; Seminati, C.; Burballa, A.; Nieto, A.; Durán, I.; Tarradas, N.; Molina-López, R.A. Antimicrobial susceptibility of bacterial isolates from urinary tract infections in companion animals in Spain. Vet. Rec. 2020, 8, 1118. [Google Scholar] [CrossRef]
- Simjee, S.; White, D.G.; McDermott, P.F.; Wagner, D.D.; Zervos, M.J.; Donabedian, S.M.; English, L.L.; Hayes, J.R.; Walker, R.D. Characterization of Tn1546 in vancomycin-resistant Enterococcus faecium isolated from canine urinary tract infections: Evidence of gene exchange between human and animal enterococci. J. Clin. Microbiol. 2002, 40, 4659–4665. [Google Scholar] [CrossRef] [Green Version]
- Iseppi, R.; Di Cerbo, A.; Messi, P.; Sabia, C. Antibiotic resistance and virulence traits in vancomycin-resistant enterococci (Vre) and extended-spectrum β-lactamase/ampc-producing (ESBL/ampc) enterobacteriaceae from humans and pets. Antibiotics 2020, 9, 152. [Google Scholar] [CrossRef] [Green Version]
- Gómez-Beltrán, D.A.; Villar, D.; López-Osorio, S.; Ferguson, D.; Monsalve, L.K.; Chaparro-Gutiérrez, J.J. Prevalence of antimicrobial resistance in bacterial isolates from dogs and cats in a veterinary diagnostic laboratory in Colombia from 2016–2019. Vet. Sci. 2020, 7, 173. [Google Scholar] [CrossRef]
- Bang, K.; An, J.U.; Kim, W.; Dong, H.J.; Kim, J.; Cho, S. Antibiotic resistance patterns and genetic relatedness of enterococcus faecalis and enterococcus faecium isolated from military working dogs in korea. J. Vet. Sci. 2017, 18, 229–236. [Google Scholar] [CrossRef]
- Poeta, P.; Costa, D.; Rodrigues, J.; Torres, C. Antimicrobial resistance and the mechanisms implicated in faecal enterococci from healthy humans, poultry and pets in Portugal. Int. J. Antimicrob. Agents 2006, 27, 131–137. [Google Scholar] [CrossRef] [PubMed]
- Cinquepalmi, V.; Monno, R.; Fumarola, L.; Ventrella, G.; Calia, C.; Greco, M.F.; De Vito, D.; Soleo, L. Environmental contamination by dog’s faeces: A public health problem? Int. J. Environ. Res. Public Health 2013, 10, 72. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Aun, E.; Kisand, V.; Laht, M.; Telling, K.; Kalmus, P.; Väli, Ü.; Brauer, A.; Remm, M.; Tenson, T. Molecular Characterization of Enterococcus Isolates From Different Sources in Estonia Reveals Potential Transmission of Resistance Genes Among Different Reservoirs. Front. Microbiol. 2021, 12, 585. [Google Scholar] [CrossRef] [PubMed]
- Portillo, A.; Ruiz-Larrea, F.; Zarazaga, M.; Alonso, A.; Martinez, J.L.; Torres, C. Macrolide Resistance Genes in Enterococcus spp. Antimicrob. Agents Chemother. 2000, 44, 967. [Google Scholar] [CrossRef] [Green Version]
- Rumi, M.V.; Nuske, E.; Mas, J.; Argüello, A.; Gutkind, G.; Di Conza, J. Antimicrobial resistance in bacterial isolates from companion animals in Buenos Aires, Argentina: 2011–2017 retrospective study. Zoonoses Public Health 2021, 00, 1–11. [Google Scholar] [CrossRef]
- Miller, W.R.; Munita, J.M.; Arias, C.A. Mechanisms of antibiotic resistance in enterococci. Expert Rev. Anti. Infect. Ther. 2014, 12, 1221–1236. [Google Scholar] [CrossRef]
- Ono, S.; Muratani, T.; Matsumoto, T. Mechanisms of resistance to imipenem and ampicillin in Enterococcus faecalis. Antimicrob. Agents Chemother. 2005, 49, 2954–2958. [Google Scholar] [CrossRef] [Green Version]
- Conceição, N.; da Silva, L.E.P.; da Costa Darini, A.L.; Pitondo-Silva, A.; de Oliveira, A.G. Penicillin-resistant, ampicillin-susceptible Enterococcus faecalis of hospital origin: Pbp4 gene polymorphism and genetic diversity. Infect. Genet. Evol. 2014, 28, 289–295. [Google Scholar] [CrossRef]
- Djorić, D.; Little, J.L.; Kristich, C.J. Multiple low-reactivity class B penicillin-binding proteins are required for cephalosporin resistance in enterococci. Antimicrob. Agents Chemother. 2020, 64, e02273-19. [Google Scholar] [CrossRef]
- Choi, J.M.; Woo, G.J. Transfer of Tetracycline Resistance Genes with Aggregation Substance in Food-Borne Enterococcus faecalis. Curr. Microbiol. 2015, 70, 476–484. [Google Scholar] [CrossRef] [Green Version]
- Kubašová, I.; Lauková, A.; Hamarová, Ľ.; Pristaš, P.; Strompfová, V. Evaluation of enterococci for potential probiotic utilization in dogs. Folia Microbiol. 2019, 64, 177–187. [Google Scholar] [CrossRef]
- Balaei Gajan, E.; Shirmohammadi, A.; Aghazadeh, M.; Alizadeh, M.; Sighari Deljavan, A.; Ahmadpour, F. Antibiotic Resistance in Enterococcus faecalis Isolated from Hospitalized Patients. J. Dent. Res. Dent. Clin. Dent. Prospects 2013, 7, 102–104. [Google Scholar] [CrossRef] [PubMed]
- Bertelloni, F.; Salvadori, C.; Moni, A.; Cerri, D.; Mani, P.; Ebani, V.V. Antimicrobial resistance in Enterococcus spp. Isolated from laying hens of backyard poultry flocks. Ann. Agric. Environ. Med. 2015, 22, 665–669. [Google Scholar] [CrossRef] [Green Version]
- Si, H.; Zhang, W.J.; Chu, S.; Wang, X.M.; Dai, L.; Hua, X.; Dong, Z.; Schwarz, S.; Liu, S. Novel plasmid-borne multidrug resistance gene cluster including lsa(E) from a Linezolid-Resistant Enterococcus faecium Isolate of Swine Origin. Antimicrob. Agents Chemother. 2015, 59, 7113–7116. [Google Scholar] [CrossRef] [Green Version]
- Cho, S.; Barrett, J.B.; Frye, J.G.; Jackson, C.R. Antimicrobial resistance gene detection and plasmid typing among multidrug resistant enterococci isolated from freshwater environment. Microorganisms 2020, 8, 1338. [Google Scholar] [CrossRef] [PubMed]
- Mast, Y.; Wohlleben, W. Streptogramins—Two are better than one! Int. J. Med. Microbiol. 2014, 304, 44–50. [Google Scholar] [CrossRef] [PubMed]
- Bortolaia, V.; Guardabassi, L. Zoonotic transmission of antimicrobial resistant enterococci: A threat to public health or an overemphasised risk? In Zoonoses-Infections Affecting Humans and Animals: Focus on Public Health Aspects; Springer Netherlands: Dordrecht, The Netherlands, 2015; pp. 407–431. ISBN 9789401794572. [Google Scholar]
- Ben Said, L.; Dziri, R.; Sassi, N.; Lozano, C.; Ben Slama, K.; Ouzari, I.; Torres, C.; Klibi, N. Species distribution, antibiotic resistance and virulence traits in canine and feline enterococci in Tunisia. Acta Vet. Hung. 2017, 65, 173–184. [Google Scholar] [CrossRef] [PubMed]
- Stȩpień-Pyśniak, D.; Hauschild, T.; Dec, M.; Marek, A.; Urban-Chmiel, R. Clonal structure and antibiotic resistance of enterococcus spp. from wild birds in Poland. Microb. Drug Resist. 2019, 25, 1227–1237. [Google Scholar] [CrossRef] [PubMed]
- Grassotti, T.T.; De Angelis Zvoboda, D.; Da Fontoura Xavier Costa, L.; De Araújo, A.J.G.; Pereira, R.I.; Soares, R.O.; Wagner, P.G.C.; Frazzon, J.; Frazzon, A.P.G. Antimicrobial resistance profiles in enterococcus spp. Isolates from fecal samples of wild and captive black capuchin Monkeys (Sapajus nigritus) in South Brazil. Front. Microbiol. 2018, 9, 2366. [Google Scholar] [CrossRef] [PubMed]
- Bhat, A.H. Bacterial zoonoses transmitted by household pets and as reservoirs of antimicrobial resistant bacteria. Microb. Pathog. 2021, 155, 104891. [Google Scholar] [CrossRef]
- Hammerum, A.M. Enterococci of animal origin and their significance for public health. Clin. Microbiol. Infect. 2012, 18, 619–625. [Google Scholar] [CrossRef]
- Zheng, J.X.; Bai, B.; Lin, Z.W.; Pu, Z.Y.; Yao, W.M.; Chen, Z.; Li, D.Y.; Deng, X.B.; Deng, Q.W.; Yu, Z.J. Characterization of biofilm formation by Enterococcus faecalis isolates derived from urinary tract infections in China. J. Med. Microbiol. 2018, 67, 60–67. [Google Scholar] [CrossRef]
- Ch’ng, J.H.; Chong, K.K.L.; Lam, L.N.; Wong, J.J.; Kline, K.A. Biofilm-associated infection by enterococci. Nat. Rev. Microbiol. 2019, 17, 82–94. [Google Scholar] [CrossRef]
- Fallah, F.; Yousefi, M.; Pourmand, M.R.; Hashemi, A.; Nazari Alam, A.; Afshar, D. Phenotypic and genotypic study of biofilm formation in Enterococci isolated from urinary tract infections. Microb. Pathog. 2017, 108, 85–90. [Google Scholar] [CrossRef]
- Mohamed, J.A.; Huang, D.B. Biofilm formation by enterococci. J. Med. Microbiol. 2007, 56, 1581–1588. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Eaton, T.J.; Gasson, M.J. Molecular Screening of Enterococcus Virulence Determinants and Potential for Genetic Exchange between Food and Medical Isolates. Appl. Environ. Microbiol. 2001, 67, 1628–1635. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Kayaoglu, G.; Ørstavik, D. Virulence factors of Enterococcus faecalis: Relationship to endodontic disease. Crit. Rev. Oral Biol. Med. 2004, 15, 308–320. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Dec, M.; Stȩpień-Pyśniak, D.; Gnat, S.; Fratini, F.; Urban-Chmiel, R.; Cerri, D.; Winiarczyk, S.; Turchi, B. Antibiotic Susceptibility and Virulence Genes in Enterococcus Isolates from Wild Mammals Living in Tuscany, Italy. Microb. Drug Resist. 2020, 26, 505–519. [Google Scholar] [CrossRef] [PubMed]
- Ferguson, D.M.; Talavera, G.N.; Hernández, L.A.R.; Weisberg, S.B.; Ambrose, R.F.; Jay, J.A. Virulence Genes among Enterococcus faecalis and Enterococcus faecium Isolated from Coastal Beaches and Human and Nonhuman Sources in Southern California and Puerto Rico. J. Pathog. 2016, 2016, 1–7. [Google Scholar] [CrossRef] [Green Version]
- Gulhan, T.; Boynukara, B.; Ciftci, A.; Sogut, M.U.; Findik, A. Characterization of Enterococcus faecalis isolates originating from different sources for their virulence factors and genes, antibiotic resistance patterns, genotypes and biofilm production. Iran. J. Vet. Res. 2015, 16, 261–266. [Google Scholar] [CrossRef] [PubMed]
- Kiruthiga, A.; Padmavathy, K.; Shabana, P.; Naveenkumar, V.; Gnanadesikan, S.; Malaiyan, J. Improved detection of esp, hyl, asa1, gelE, cylA virulence genes among clinical isolates of Enterococci. BMC Res. Notes 2020, 13, 1–7. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Antimicrobial | Resistant N (%) | Intermediate N (%) | Susceptible N (%) | |||||||
|---|---|---|---|---|---|---|---|---|---|---|
| Class | Agent | Total | Stool | Urine | Total | Stool | Urine | Total | Stool | Urine |
| Ansamycins | RD | 22 (43.1) | 15 (51.7) | 7 (31.8) | 11 (21.6) | 5 (17.2) | 6 (27.3) | 18 (35.3) | 9 (31.0) | 9 (40.9) |
| Phenicols | C | 21 (41.2) | 16 (55.2) | 5 (22.7) | 8 (15.7) | 5 (17.2) | 3 (13.6) | 22 (43.1) | 8 (27.6) | 14 (63.6) |
| Oxazolidinones | LZD | 21 (41.2) | 21 (72.4) | 0 (0.0) | 5 (9.8) | 3 (10.3) | 2 (9.1) | 25 (49.0) | 5 (17.2) | 20 (90.9) |
| Nitrofurantoins | F | 13 (25.5) | 13 (44.8) | 0 (0.0) | 5 (9.8) | 3 (10.3) | 2 (9.1) | 33 (64.7) | 13 (44.8) | 20 (90.9) |
| Folate pathway antagonists | W | 31 (60.8) | 23 (79.3) | 8 (36.4) | 19 (37.3) | 6 (20.7) | 13 (59.1) | 1 (2.0) | 0 (0.0) | 1 (4.5) |
| Aminoglycoside | N | 46 (90.2) | 28 (96.6) | 18 (81.8) | 4 (7.8) | 1 (3.4) | 3 (13.6) | 1 (2.0) | 0 (0.0) | 1 (4.5) |
| CN | 35 (68.6) | 20 (69.0) | 15 (68.2) | 4 (7.8) | 2 (6.9) | 2 (9.1) | 12 (23.5) | 7 (24.1) | 5 (22.7) | |
| S | 48 (94.1) | 28 (96.6) | 20 (90.9) | 2 (3.9) | 1 (3.4) | 1 (4.5) | 1 (2.0) | 0 (0.0) | 1 (4.5) | |
| Cephems | KF | 32 (62.7) | 17 (58.6) | 15 (68.2) | 14 (27.5) | 10 (34.5) | 4 (18.2) | 5 (9.8) | 2 (6.9) | 3 (13.6) |
| Fluoroquinolones | CIP | 34 (66.7) | 18 (62.1) | 16 (72.7) | 7 (13.7) | 7 (24.1) | 0 (0.0) | 10 (19.6) | 4 (13.8) | 6 (27.3) |
| ENR | 38 (74.5) | 22 (75.9) | 16 (72.7) | 9 (17.6) | 5 (17.2) | 4 (18.2) | 4 (7.8) | 2 (6.9) | 2 (9.1) | |
| Glycopeptides | TEC | 2 (3.9) | 2 (6.9) | 0 (0.0) | 6 (11.8) | 3 (10.3) | 3 (13.6) | 43 (84.3) | 24 (82.8) | 19 (86.4) |
| VA | 7 (13.7) | 2 (6.9) | 5 (22.7) | 15 (29.4) | 7 (24.1) | 8 (36.4) | 29 (56.9) | 20 (69.0) | 9 (40.9) | |
| Macrolides, Streptogramins, Lincosamides | E | 29 (56.9) | 16 (55.2) | 13 (59.1) | 17 (33.3) | 11 (37.9) | 6 (27.3) | 5 (9.8) | 2 (6.9) | 3 (13.6) |
| QD | 40 (78.4) | 19 (65.5) | 21 (95.5) | 7 (13.7) | 6 (20.7) | 1 (4.5) | 4 (7.8) | 4 (13.8) | 0 (0.0) | |
| DA | 43 (84.3) | 21 (72.4) | 22 (100.0) | 6 (11.8) | 6 (20.7) | 0 (0.0) | 2 (3.9) | 2 (6.9) | 0 (0.0) | |
| Penicillins | OX | 50 (98.0) | 28 (96.6) | 22 (100.0) | 1 (2.0) | 1 (3.4) | 0 (0.0) | 0 (0.0) | 0 (0.0) | 0 (0.0) |
| AMC | 6 (11.8) | 1 (3.4) | 5 (22.7) | 6 (11.8) | 6 (20.7) | 0 (0.0) | 39 (76.5) | 22 (75.9) | 17 (77.3) | |
| AMP | 26 (51.0) | 17 (58.6) | 9 (40.9) | 10 (19.6) | 0 (0.0) | 10 (45.5) | 15 (29.4) | 12 (41.4) | 3 (13.6) | |
| Tetracyclines | TE | 40 (78.4) | 24 (82.8) | 16 (72.7) | 5 (9.8) | 2 (6.9) | 3 (13.6) | 6 (11.8) | 3 (10.3) | 3 (13.6) |
| TIG | 15 (29.4) | 15 (51.7) | 0 (0.0) | 8 (15.7) | 6 (20.7) | 2 (9.1) | 28 (54.9) | 8 (27.6) | 20 (90.9) | |
| Origin of Samples | Bacteria Species | Multidrug-Resistant Category | Ampicillin MIC ≥ 64 µg/mL | HLAR | HLAR in Isolates with Ampicillin MIC ≥ 64 µg/mL | |||||
|---|---|---|---|---|---|---|---|---|---|---|
| MDR | XDR | HLSR | HLGR | Both | HLSR | HLGR | Both | |||
| Feces of healthy dogs | E. faecalis | 11 | 3 | 1 | 12 | 10 | 8 | 1 | 0 | 0 |
| E. faecium | 11 | 3 | 13 | 2 | 0 | 0 | 2 | 0 | 0 | |
| Urine of dogs with UTIs | E. faecalis | 16 | 0 | 6 | 4 | 0 | 0 | 2 | 0 | 0 |
| E. faecium | 3 | 0 | 2 | 1 | 2 | 1 | 1 | 2 | 1 | |
| E. gallinarum | 3 | 0 | 2 | 2 | 1 | 1 | 2 | 1 | 1 | |
| Total | 44 | 6 | 24 | 21 | 13 | 10 | 8 | 3 | 2 | |
| Antibiotic | Resistance Gene | Healthy Dog Feces | Sick Dog Urine | Total n = 51 | |||
|---|---|---|---|---|---|---|---|
| E. faecalis n = 14 | E. faecium n = 15 | E. faecalis n = 16 | E. faecium n = 3 | E. gallinarum n = 3 | |||
| Gentamicin (HL) | aac(6’)-Ie-aph(2”)-Ia | 0 | 1 | 0 | 1 | 0 | 2 |
| Kanamycin (HL) | aph(3′)-IIIa | 12 | 3 | 2 | 1 | 2 | 20 |
| Gentamicin and other aminoglycosides (ML) | aac(6′)-Ii | 0 | 15 | 0 | 3 | 0 | 18 |
| Streptomycin (HL) | ant(6’)-Ia | 9 | 4 | 1 | 2 | 1 | 17 |
| Chloramphenicol | cat | 5 | 0 | 3 | 1 | 1 | 10 |
| β lactams | pbp4 | 12 | 0 | 15 | 0 | 0 | 27 |
| pbp5 | 0 | 3 | 0 | 0 | 0 | 3 | |
| Tetracyclines | tet(M) | 14 | 8 | 10 | 1 | 3 | 36 |
| tet(L) | 3 | 3 | 3 | 1 | 0 | 10 | |
| Lincosamides | lnuB | 7 | 3 | 1 | 0 | 0 | 11 |
| Macrolides | erm(B) | 8 | 3 | 5 | 1 | 1 | 18 |
| mrs(A/B) | 0 | 15 | 0 | 3 | 0 | 18 | |
| mrs(C) | 1 | 13 | 0 | 2 | 0 | 16 | |
| mef(A/B) | 0 | 1 | 0 | 0 | 0 | 1 | |
| - | Int-Tn | 10 | 5 | 9 | 1 | 3 | 28 |
| Sample | Species | WBP | MBP | SBP |
|---|---|---|---|---|
| Feces of healthy dogs | E. faecalis | 1 | 3 | 10 |
| E. faecium | 15 | 0 | 0 | |
| Urine of dogs with UTIs | E. faecalis | 3 | 5 | 8 |
| E. faecium | 2 | 1 | 0 | |
| E. gallinarum | 0 | 2 | 1 | |
| Total | 21 | 11 | 19 | |
| Virulence Genes | Feces of Healthy Dogs | Urine of Dogs with UTIs | Total n= 51 (%) | |||
|---|---|---|---|---|---|---|
| E. faecalis n = 14 | E. faecium n = 15 | E. faecalis n = 16 | E. faecium n = 3 | E. gallinarum n = 3 | ||
| cylA | 9 (8 a) | 2 (2 a) | 7 (2 a) | 1 (1 a) | 0 | 19 (37.3) |
| gelE | 8 (8 b) | 2 (1 a) | 10 (5 b) | 0 | 1 | 21 (41.2) |
| asa1 | 10 | 2 | 9 | 4 | 2 | 27 (52.9) |
| esp | 10 | 2 | 5 | 0 | 1 | 18 (35.3) |
| efaAfs | 14 | 0 | 16 | 0 | 0 | 30 (58.8) |
| efaAfm | 0 | 15 | 0 | 3 | 0 | 18 (35.3) |
| sgrA | 0 | 2 | 0 | 2 | 0 | 4 (7.8) |
| pstD | 0 | 1 | 0 | 2 | 0 | 3 (5.9) |
| orf1481 | 0 | 1 | 0 | 3 | 0 | 4 (7.8) |
| IS16 | 0 | 8 | 0 | 2 | 0 | 10 (19.6) |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Stępień-Pyśniak, D.; Bertelloni, F.; Dec, M.; Cagnoli, G.; Pietras-Ożga, D.; Urban-Chmiel, R.; Ebani, V.V. Characterization and Comparison of Enterococcus spp. Isolates from Feces of Healthy Dogs and Urine of Dogs with UTIs. Animals 2021, 11, 2845. https://doi.org/10.3390/ani11102845
Stępień-Pyśniak D, Bertelloni F, Dec M, Cagnoli G, Pietras-Ożga D, Urban-Chmiel R, Ebani VV. Characterization and Comparison of Enterococcus spp. Isolates from Feces of Healthy Dogs and Urine of Dogs with UTIs. Animals. 2021; 11(10):2845. https://doi.org/10.3390/ani11102845
Chicago/Turabian StyleStępień-Pyśniak, Dagmara, Fabrizio Bertelloni, Marta Dec, Giulia Cagnoli, Dorota Pietras-Ożga, Renata Urban-Chmiel, and Valentina Virginia Ebani. 2021. "Characterization and Comparison of Enterococcus spp. Isolates from Feces of Healthy Dogs and Urine of Dogs with UTIs" Animals 11, no. 10: 2845. https://doi.org/10.3390/ani11102845