
Toward Precise Nutrient Value of Feed in Growing Pigs: Effect of Meal Size, Frequency and Dietary Fibre on Nutrient Utilisation

 , ,
, ,
Abstract
:Simple Summary
Abstract
1. Background
2. Natural Feed Intake of the Pig
3. Mechanisms Regulating Meal Size and Frequency
3.1. Distension
3.2. Osmotic Receptors
3.3. Hormonal Receptors
4. Impact of Meal Size and Frequency on Transit Time and Digestibility of Nutrients
5. Impact of Meal Frequency on Blood Profile, Metabolism and Body Composition
6. Impact of Fibres on Digestive Function and Metabolism
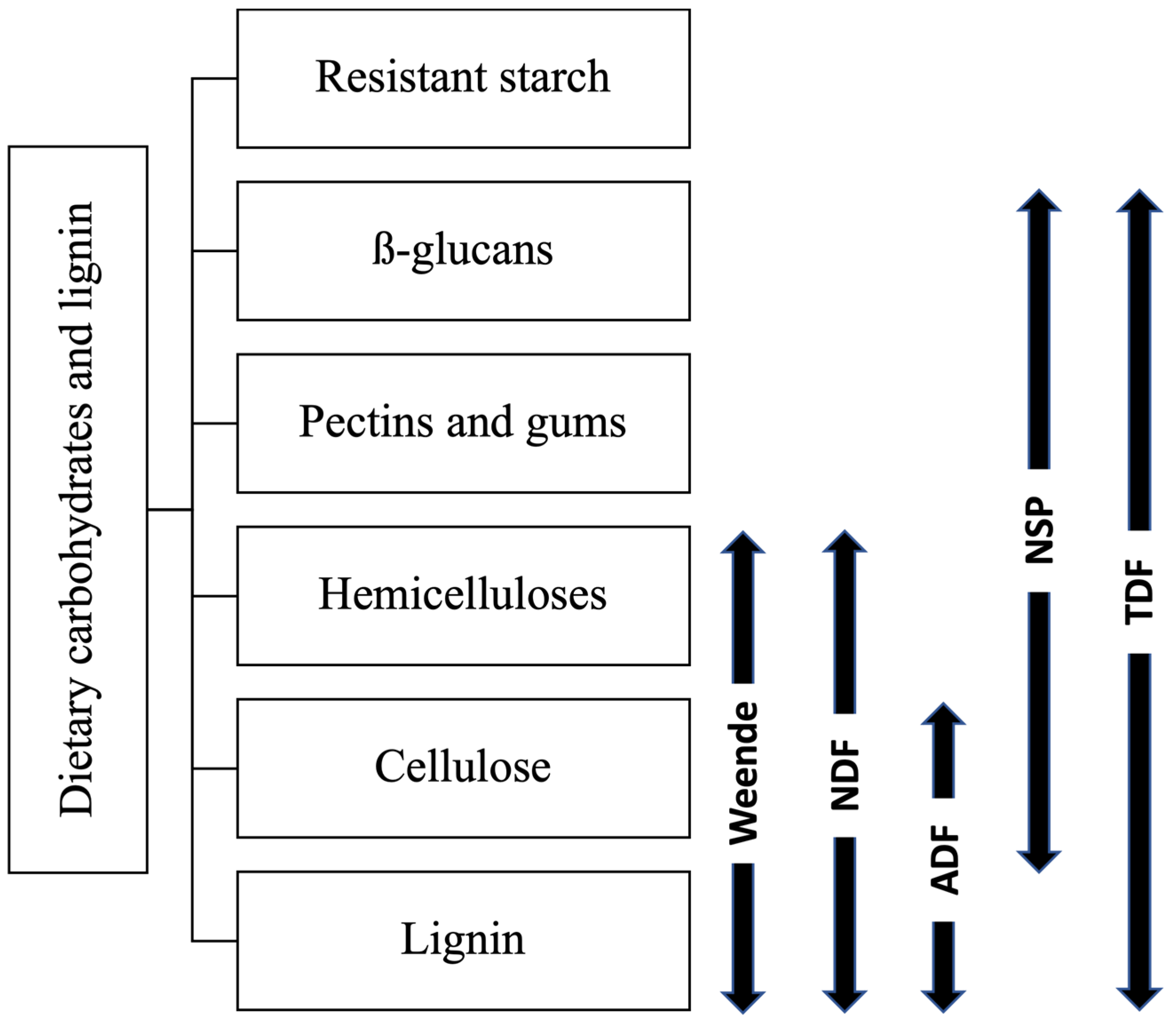
7. Fibre Content in Pig Diets
8. Impact of Fibres on Feed Digestion
8.1. Dilution of Energy Content
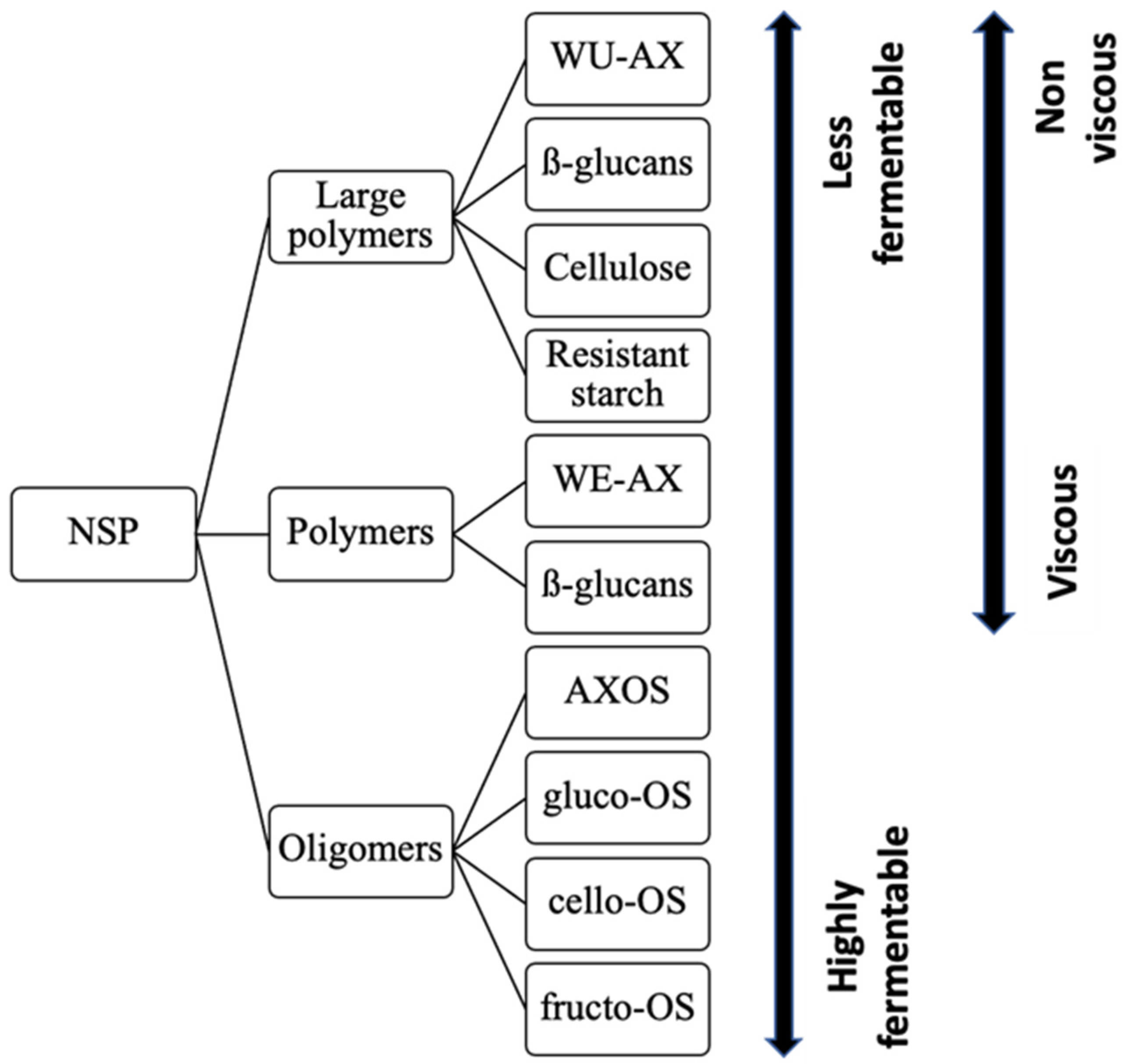
8.2. Viscosity
9. Impact of Exogenous Enzymes on Nutrient Utilisation
9.1. pH
9.2. Retention Time
10. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Conflicts of Interest
References
- Rocadembosch, J.; Amador, J.; Bernaus, J.; Font, J.; Fraile, L.J. Production parameters and pig production cost: Temporal evolution 2010–2014. Porc. Health Manag. 2016, 2, 11. [Google Scholar] [CrossRef] [Green Version]
- CoBank. Surging Feed Prices Will Challenge the Protein Sector’s Recovery. 2020. Available online: https://www.wisfarmer.com/ (accessed on 1 August 2021).
- Woyengo, T.; Beltranena, E.; Zijlstra, R. Nonruminant nutrition symposium: Controlling feed cost by including alternative ingredients into pig diets: A review. J. Anim. Sci. 2014, 92, 1293–1305. [Google Scholar] [CrossRef] [Green Version]
- Zijlstra, R.T.; Owusu-Asiedu, A.; Simmins, P.H. Future of NSP-degrading enzymes to improve nutrient utilization of co-products and gut health in pigs. Livest. Sci. 2010, 134, 255–257. [Google Scholar] [CrossRef]
- Stein, H.H. Distillers dried grains with solubles (DDGS) in diets fed to swine. Swine Focus 2007, 1, 1–8. [Google Scholar]
- Messad, F.; Létourneau-Montminy, M.; Charbonneau, E.; Sauvant, D.; Guay, F. Meta-analysis of the amino acid digestibility of oilseed meal in growing pigs. Animal 2016, 10, 1635–1644. [Google Scholar] [CrossRef]
- Noblet, J.; Shi, X. Comparative digestibility of energy and nutrients in growing pigs fed ad libitum and adults sows fed at maintenance. Livest. Prod. Sci. 1993, 34, 137–152. [Google Scholar] [CrossRef]
- Friend, D.W.; Cunningham, H.M. Growth, Carcass, Blood and Fat Studies with Pigs Fed Once or Five Times Daily. J. Anim. Sci. 1967, 26, 316–322. [Google Scholar] [CrossRef]
- Laplace, J.-P.; Germain, C. Enregistrement du pH intragastrique chez le porc: Variations liees a la nature, a l’importance et a l’intervalle des repas ches l’animal en finition. Ann. Zootech. 1974, 23, 89–104. [Google Scholar] [CrossRef] [Green Version]
- Hee, J.; Sauer, W.C.; Mosenthin, R. The Effect of Frequency of Feeding on the Pancreatic Secretions in the Pig. J. Anim. Physiol. Anim. Nutr.-Z. Tierphysiol. Tierernahr. Futterm. 1988, 60, 249–256. [Google Scholar] [CrossRef]
- Adeola, O.; Lewis, A.; Southern, L. Digestion and balance techniques in pigs. Swine Nutr. 2001, 2, 903–916. [Google Scholar]
- Liu, J.; Liu, Z.; Chen, L.; Zhang, H. Effects of feed intake and dietary nutrient density on apparent ileal and total tract digestibility of nutrients and gross energy for growing pigs. J. Anim. Sci. 2016, 94, 4251–4258. [Google Scholar] [CrossRef] [Green Version]
- Low, A.G. Secretory Response of the Pig Gut to Non-Starch Polysaccharides. Anim. Feed Sci. Technol. 1989, 23, 55–65. [Google Scholar] [CrossRef]
- Bedford, M.R. The evolution and application of enzymes in the animal feed industry: The role of data interpretation. Br. Poult. Sci. 2018, 59, 486–493. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Van Leeuwen, P.; Jansman, A. Effects of dietary water holding capacity and level of fermentable organic matter on digesta passage in various parts of the digestive tract in growing pigs. Livest. Sci. 2007, 109, 77–80. [Google Scholar] [CrossRef]
- Wilfart, A.; Montagne, L.; Simmins, H.; Noblet, J.; Milgen, J. Digesta transit in different segments of the gastrointestinal tract of pigs as affected by insoluble fibre supplied by wheat bran. Br. J. Nutr. 2007, 98, 54–62. [Google Scholar] [CrossRef] [Green Version]
- Zijlstra, R.T.; Jha, R.; Woodward, A.D.; Fouhse, J.; van Kempen, T.A.T.G. Starch and fiber properties affect their kinetics of digestion and thereby digestive physiology in pigs. J. Anim. Sci. 2012, 90, 49–58. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Selle, P.; Ravindran, V.; Partridge, G. Beneficial effects of xylanase and/or phytase inclusions on ileal amino acid digestibility, energy utilisation, mineral retention and growth performance in wheat-based broiler diets. Anim. Feed Sci. Technol. 2009, 153, 303–313. [Google Scholar] [CrossRef]
- Kerr, B.J.; Shurson, G.C. Strategies to improve fiber utilization in swine. J. Anim. Sci. Biotechnol. 2013, 4, 11. [Google Scholar] [CrossRef] [Green Version]
- Dersjant-Li, Y.; Awati, A.; Schulze, H.; Partridge, G. Phytase in non-ruminant animal nutrition: A critical review on phytase activities in the gastrointestinal tract and influencing factors. J. Sci. Food Agric. 2015, 95, 878–896. [Google Scholar] [CrossRef] [Green Version]
- Bastianelli, D.; Sauvant, D.; Rerat, A. Mathematical modeling of digestion and nutrient absorption in pigs. J. Anim. Sci. 1996, 74, 1873–1887. [Google Scholar] [CrossRef] [Green Version]
- Rivest, J.; Bernier, J.F.; Pomar, C. A dynamic model of protein digestion in the small intestine of pigs. J. Anim. Sci. 2000, 78, 328–340. [Google Scholar] [CrossRef] [Green Version]
- Letourneau-Montminy, M.P.; Narcy, A.; Lescoat, P.; Magnin, M.; Bernier, J.F.; Sauvant, D.; Jondreville, C.; Pomar, C. Modeling the fate of dietary phosphorus in the digestive tract of growing pigs. J. Anim. Sci. 2011, 89, 3596–3611. [Google Scholar] [CrossRef] [Green Version]
- Collier, G.; Johnson, D.F.; Mitchell, C. The relation between meal size and the time between meals: Effects of cage complexity and food cost. Physiol. Behav. 1999, 67, 339–346. [Google Scholar] [CrossRef]
- Montgomery, G.W.; Flux, D.S.; Carr, J.R. Feeding patterns in pigs: The effects of amino acid deficiency. Physiol. Behav. 1978, 20, 693–698. [Google Scholar] [CrossRef]
- Hyun, Y.; Ellis, M.; McKeith, F.K.; Wilson, E.R. Feed intake pattern of group-housed growing-finishing pigs monitored using a computerized feed intake recording system. J. Anim. Sci. 1997, 75, 1443–1451. [Google Scholar] [CrossRef] [Green Version]
- Andretta, I.; Pomar, C.; Kipper, M.; Hauschild, L.; Rivest, J. Feeding behavior of growing-finishing pigs reared under precision feeding strategies. J. Anim. Sci. 2016, 94, 3042–3050. [Google Scholar] [CrossRef] [Green Version]
- Dourmad, J.Y. Standing and feeding behaviour of the lactating sow: Effect of feeding level during pregnancy. Appl. Anim. Behav. Sci. 1993, 37, 311–319. [Google Scholar] [CrossRef]
- Vargovic, L.; Hermesch, S.; Athorn, R.Z.; Bunter, K.L. Feed intake and feeding behavior traits for gestating sows recorded using electronic sow feeders. J. Anim. Sci. 2020, 99, skaa395. [Google Scholar] [CrossRef] [PubMed]
- Auffray, P.; Marcilloux, J.C. Étude de la séquence alimentaire du porc adulte. Reprod. Nutr. Dev. 1983, 23, 517–524. [Google Scholar] [CrossRef] [PubMed]
- Auffray, P.; Marcilloux, J.C. Analyse de la séquence alimentaire du porc, du sevrage à l’état adulte. Reprod. Nutr. Dev. 1980, 20, 1625–1632. [Google Scholar] [CrossRef] [PubMed]
- Le Magnen, J.; Devos, M. Parameters of the meal pattern in rats: Their assessment and physiological significance. Neurosci. Biobehav. Rev. 1980, 4, 1–11. [Google Scholar] [CrossRef]
- Bigelow, J.A.; Houpt, T.R. Feeding and drinking patterns in young pigs. Physiol. Behav. 1988, 43, 99–109. [Google Scholar] [CrossRef]
- Chen, D.; Xin, W.S.; Yao, T.X.; Rao, L.; Xu, S.Q.; Xiao, S.J.; Zhang, Z.Y. Multi-breed investigation of pig social rank and biological rhythm based on feeding behaviors at electronic feeding stations. Livest. Sci. 2021, 245, 104419. [Google Scholar] [CrossRef]
- Beaulieu, A.D.; Williams, N.H.; Patience, J.F. Response to dietary digestible energy concentration in growing pigs fed cereal grain-based diets. J. Anim. Sci. 2009, 87, 965–976. [Google Scholar] [CrossRef] [Green Version]
- Quemeneur, K.; Montagne, L.; Le Gall, M.; Lechevestrier, Y.; Labussiere, E. Relation between feeding behaviour and energy metabolism in pigs fed diets enriched in dietary fibre and wheat aleurone. Animal 2020, 14, 508–519. [Google Scholar] [CrossRef]
- Li, Q.Y.; Patience, J.F. Factors involved in the regulation of feed and energy intake of pigs. Anim. Feed Sci. Technol. 2017, 233, 22–33. [Google Scholar] [CrossRef]
- Roura, E. Chapter 8 Dietary fibre, gut sensing and modulation of feed intake in pigs and chickens. In The Value of Fibre: Engaging the Second Brain for Animal Nutrition; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019; pp. 2485–2490. [Google Scholar]
- Tian, M.; Heng, J.H.; Song, H.Q.; Shi, K.; Lin, X.F.; Chen, F.; Guan, W.T.; Zhang, S. Dietary Branched-Chain Amino Acids Regulate Food Intake Partly through Intestinal and Hypothalamic Amino Acid Receptors in Piglets. J. Agric. Food Chem. 2019, 67, 6809–6818. [Google Scholar] [CrossRef] [PubMed]
- Henry, Y.; Seve, B.; Colleaux, Y.; Ganier, P.; Saligaut, C.; Jego, P. Interactive effects of dietary levels of tryptophan and protein on voluntary feed intake and growth performance in pigs, in relation to plasma free amino acids and hypothalamic serotonin. J. Anim. Sci. 1992, 70, 1873–1887. [Google Scholar] [CrossRef]
- Roura, E.; Fu, M. Taste, nutrient sensing and feed intake in pigs (130 years of research: Then, now and future). Anim. Feed Sci. Technol. 2017, 233, 3–12. [Google Scholar] [CrossRef]
- Nyachoti, C.M.; Zijlstra, R.T.; de Lange, C.F.M.; Patience, J.F. Voluntary feed intake in growing-finishing pigs: A review of the main determining factors and potential approaches for accurate predictions. Can. J. Anim. Sci. 2004, 84, 549–566. [Google Scholar] [CrossRef]
- Dänicke, S.; Brüssow, K.-P.; Valenta, H.; Ueberschär, K.-H.; Tiemann, U.; Schollenberger, M. On the effects of graded levels of Fusarium toxin contaminated wheat in diets for gilts on feed intake, growth performance and metabolism of deoxynivalenol and zearalenone. Mol. Nutr. Food Res. 2005, 49, 932–943. [Google Scholar] [CrossRef]
- Schinckel, A.P.; Einstein, M.E.; Jungst, S.; Matthews, J.O.; Booher, C.; Dreadin, T.; Fralick, C.; Wilson, E.; Boyd, R.D. Daily feed intake, energy intake, growth rate and measures of dietary energy efficiency of pigs from four sire lines fed diets with high or low metabolizable and net energy concentrations. Asian-Australas. J. Anim. Sci. 2012, 25, 410–420. [Google Scholar] [CrossRef] [Green Version]
- Da Fonseca de Oliveira, A.C.; Vanelli, K.; Sotomaior, C.S.; Weber, S.H.; Costa, L.B. Impacts on performance of growing-finishing pigs under heat stress conditions: A meta-analysis. Vet. Res. Commun. 2019, 43, 37–43. [Google Scholar] [CrossRef]
- Patience, J.F.; Umboh, J.F.; Chaplin, R.K.; Nyachoti, C.M. Nutritional and physiological responses of growing pigs exposed to a diurnal pattern of heat stress. Livest. Prod. Sci. 2005, 96, 205–214. [Google Scholar] [CrossRef]
- Turner, S.P.; Dahlgren, M.; Arey, D.S.; Edwards, S.A. Effect of social group size and initial live weight on feeder space requirement of growing pigs given food ad libitum. Anim. Sci. 2002, 75, 75–83. [Google Scholar] [CrossRef]
- Stephens, D.B. The effects of alimentary infusions of glucose, amino acids, or neutral fat on meal size in hungry pigs. J. Physiol. 1980, 299, 453–463. [Google Scholar] [CrossRef] [PubMed]
- Houpt, T.R.; Baldwin, B.A.; Houpt, K.A. Effects of duodenal osmotic loads on spontaneous meals in pigs. Physiol. Behav. 1983, 30, 787–795. [Google Scholar] [CrossRef]
- Smeets, A.J.; Westerterp-Plantenga, M.S. Acute effects on metabolism and appetite profile of one meal difference in the lower range of meal frequency. Br. J. Nutr. 2008, 99, 1316–1321. [Google Scholar] [CrossRef]
- Strader, A.D.; Woods, S.C. Gastrointestinal hormones and food intake. Gastroenterology 2005, 128, 175–191. [Google Scholar] [CrossRef]
- Tolkamp, B.J.; Howie, J.A.; Bley, T.A.G.; Kyriazakis, I. Prandial correlations and the structure of feeding behaviour. Appl. Anim. Behav. Sci. 2012, 137, 53–65. [Google Scholar] [CrossRef]
- Young, R.J.; Lawrence, A.B. Feeding behaviour of pigs in groups monitored by a computerized feeding system. Anim. Sci. 2010, 58, 145–152. [Google Scholar] [CrossRef]
- Savory, C.J. Correlations between Meals and Inter-Meal Intervals in Japanese Quail and Their Significance in the Control of Feeding. Behav. Process. 1981, 6, 23–36. [Google Scholar] [CrossRef]
- Houpt, K.A. Gastrointestinal factors in hunger and satiety. Neurosci. Biobehav. Rev. 1982, 6, 145–164. [Google Scholar] [CrossRef]
- Flint, A.; Raben, A.; Astrup, A.; Holst, J.J. Glucagon-like peptide 1 promotes satiety and suppresses energy intake in humans. J. Clin. Investig. 1998, 101, 515–520. [Google Scholar] [CrossRef]
- Salgado, H.H.; Methot, S.; Remus, A.; Letourneau-Montminy, M.P.; Pomar, C. A novel feeding behavior index integrating several components of the feeding behavior of finishing pigs. Animal 2021, 15, 100251. [Google Scholar] [CrossRef] [PubMed]
- D’Eath, R.B.; Jarvis, S.; Baxter, E.M.; Houdijk, J. Chapter 7 Mitigating hunger in pregnant sows. In Advances in Pig Welfare; Špinka, M., Camerlink, I., Eds.; Elsevier Woodhead Publishing: Duxford, UK, 2018; pp. 199–234. [Google Scholar] [CrossRef]
- Kojima, M.; Kangawa, K. Ghrelin: Structure and function. Physiol. Rev. 2005, 85, 495–522. [Google Scholar] [CrossRef]
- Svihus, B.; Hervik, A. Chapter 7 The influence of fibre on gut physiology and feed intake regulation. In The Value of Fibre: Engaging the Second Brain for Animal Nutrition; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019; pp. 683–689. [Google Scholar]
- Auffray, P.; Martinet, J.; Rérat, A. Quelques aspects du transit gastro-intestinal chez le porc. Ann. Biol. Anim. Bioch. Biophys. 1967, 7, 261–279. [Google Scholar] [CrossRef] [Green Version]
- Gregory, P.C.; Mcfadyen, M.; Rayner, D.V. Relation between Gastric-Emptying and Short-Term Regulation of Food-Intake in the Pig. Physiol. Behav. 1989, 45, 677–683. [Google Scholar] [CrossRef]
- Treacy, P.J.; Jamieson, G.G.; Dent, J. Pyloric motility and liquid gastric emptying during barostatic control of gastric pressure in pigs. J. Physiol. 1994, 474, 361–366. [Google Scholar] [CrossRef] [Green Version]
- Lepionka, L.; Malbert, C.H.; Laplace, J.P. Proximal gastric distension modifies ingestion rate in pigs. Reprod. Nutr. Dev. 1997, 37, 449–457. [Google Scholar] [CrossRef] [Green Version]
- Nakazato, M.; Murakami, N.; Date, Y.; Kojima, M.; Matsuo, H.; Kangawa, K.; Matsukura, S. A role for ghrelin in the central regulation of feeding. Nature 2001, 409, 194–198. [Google Scholar] [CrossRef] [PubMed]
- Owusu-Asiedu, A.; Patience, J.F.; Laarveld, B.; Van Kessel, A.G.; Simmins, P.H.; Zijlstra, R.T. Effects of guar gum and cellulose on digesta passage rate, ileal microbial populations, energy and protein digestibility, and performance of grower pigs. J. Anim. Sci. 2006, 84, 843–852. [Google Scholar] [CrossRef] [PubMed]
- Crawley, J.N.; Corwin, R.L. Biological actions of cholecystokinin. Peptides 1994, 15, 731–755. [Google Scholar] [CrossRef]
- Wank, S.A. Cholecystokinin receptors. Am. J. Physiol. 1995, 269, G628–G646. [Google Scholar] [CrossRef] [PubMed]
- McIntosh, C.H.; Widenmaier, S.; Kim, S.J. Glucose-dependent insulinotropic polypeptide (Gastric Inhibitory Polypeptide; GIP). Vitam. Horm. 2009, 80, 409–471. [Google Scholar] [CrossRef]
- Regmi, P.R.; van Kempen, T.A.; Matte, J.J.; Zijlstra, R.T. Starch with high amylose and low in vitro digestibility increases short-chain fatty acid absorption, reduces peak insulin secretion, and modulates incretin secretion in pigs. J. Nutr. 2011, 141, 398–405. [Google Scholar] [CrossRef] [Green Version]
- Holst, J.J. The physiology of glucagon-like peptide 1. Physiol. Rev. 2007, 87, 1409–1439. [Google Scholar] [CrossRef]
- Cummings, J.H.; Edmond, L.M.; Magee, E.A. Dietary carbohydrates and health: Do we still need the fibre concept? Clin. Nutr. Suppl. 2004, 1, 5–17. [Google Scholar] [CrossRef]
- Delzenne, N.M.; Neyrinck, A.M.; Cani, P.D. Modulation of the gut microbiota by nutrients with prebiotic properties: Consequences for host health in the context of obesity and metabolic syndrome. Microb. Cell Fact. 2011, 10 (Suppl. 1), S10. [Google Scholar] [CrossRef] [Green Version]
- Kaczmarczyk, M.M.; Miller, M.J.; Freund, G.G. The health benefits of dietary fiber: Beyond the usual suspects of type 2 diabetes mellitus, cardiovascular disease and colon cancer. Metabolism 2012, 61, 1058–1066. [Google Scholar] [CrossRef] [Green Version]
- Hooda, S.; Metzler-Zebeli, B.U.; Vasanthan, T.; Zijlstra, R.T. Effects of viscosity and fermentability of purified non-starch polysaccharides on ileal and total tract nutrient digestibility in ileal-cannulated grower pigs. Livest. Sci. 2010, 134, 79–81. [Google Scholar] [CrossRef]
- Sleeth, M.L.; Thompson, E.L.; Ford, H.E.; Zac-Varghese, S.E.; Frost, G. Free fatty acid receptor 2 and nutrient sensing: A proposed role for fibre, fermentable carbohydrates and short-chain fatty acids in appetite regulation. Nutr. Res. Rev. 2010, 23, 135–145. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Bach Knudsen, K.; Lærke, H. Carbohydrates and lignin in the feed-from sugars to complex composed fibres. In Feed Evaluation Science; Moughan, P., Hendriks, W.H., Eds.; Wageningen Academic Publishers: Wageningen, The Netherlands, 2018. [Google Scholar]
- Tso, P.; Liu, M. Apolipoprotein A-IV, food intake, and obesity. Physiol. Behav. 2004, 83, 631–643. [Google Scholar] [CrossRef] [PubMed]
- González-Ortiz, G.; Gomes, G.A.; dos Santos, T.T.; Bedford, M.R. Chapter 14 New Strategies influencing gut functionality and animal performance. In The Value of Fibre: Engaging the Second Brain for Animal Nutrition; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019. [Google Scholar]
- Ingerslev, A.K.; Mutt, S.J.; Laerke, H.N.; Hedemann, M.S.; Theil, P.K.; Nielsen, K.L.; Jorgensen, H.; Herzig, K.H.; Bach Knudsen, K.E. Postprandial PYY increase by resistant starch supplementation is independent of net portal appearance of short-chain fatty acids in pigs. PLoS ONE 2017, 12, e0185927. [Google Scholar] [CrossRef] [Green Version]
- Auwerx, J.; Staels, B. Leptin. Lancet (London, England) 1998, 351, 737–742. [Google Scholar] [CrossRef]
- Ahima, R.S.; Flier, J.S. Leptin. Annu. Rev. Physiol. 2000, 62, 413–437. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Barb, C.R.; Yan, X.; Azain, M.J.; Kraeling, R.R.; Rampacek, G.B.; Ramsay, T.G. Recombinant porcine leptin reduces feed intake and stimulates growth hormone secretion in swine. Domest. Anim. Endocrinol. 1998, 15, 77–86. [Google Scholar] [CrossRef]
- Raucci, R.; Rusolo, F.; Sharma, A.; Colonna, G.; Castello, G.; Costantini, S. Functional and structural features of adipokine family. Cytokine 2013, 61, 1–14. [Google Scholar] [CrossRef]
- Suriano, F.; Van Hul, M.; Cani, P.D. Gut microbiota and regulation of myokine-adipokine function. Curr. Opin. Pharmacol. 2020, 52, 9–17. [Google Scholar] [CrossRef]
- Frandson, R.D.; Wilke, W.L.; Fails, A.D. Anatomy and Physiology of Farm Animals, 7th ed.; Wiley-Blackwell: Hoboken, NJ, USA, 2009. [Google Scholar]
- Roth, F.v.; Kirchgessner, M. Verdaulichkeit und intestinale Passagerate beim Schwein in Abhängigkeit vom Fütterungsniveau und Rohfasergehalt des Futters. Z. Tierphysiol. Tierernähr. Futterm. 1985, 53, 254–264. [Google Scholar] [CrossRef]
- De Haer, L.C.M.; de Vries, A.G. Feed intake patterns of and feed digestibility in growing pigs housed individually or in groups. Livest. Prod. Sci. 1993, 33, 277–292. [Google Scholar] [CrossRef]
- Botermans, J.A.M.; Hedemann, M.S.; Sorhede-Winzell, M.; Erlanson-Albertsson, C.; Svendsen, J.; Evilevitch, L.; Pierzynowski, S.G. The effect of feeding time (day versus night) and feeding frequency on pancreatic exocrine secretion in pigs. J. Anim. Physiol. Anim. Nutr.-Z. Tierphysiol. Tierernahr. Futterm. 2000, 83, 24–35. [Google Scholar] [CrossRef]
- Mroz, Z.; Jongbloed, A.M.; Kemme, P.A. Apparent Digestibility and Retention of Nutrients Bound to Phytate Complexes as Influenced by Microbial Phytase and Feeding Regimen in Pigs. J. Anim. Sci. 1994, 72, 126–132. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Jia, M.; Zhang, H.; Xu, J.; Su, Y.; Zhu, W. Feeding frequency affects the growth performance, nutrient digestion and absorption of growing pigs with the same daily feed intake. Livest. Sci. 2021, 250, 104558. [Google Scholar] [CrossRef]
- Ruckenbusch, Y.; Bueno, L. The effect of feeding on the motility of the stomach and small intestine in the pig. Br. J. Nutr. 1976, 35, 397–405. [Google Scholar] [CrossRef] [Green Version]
- Chastanet, F.; Pahm, A.A.; Pedersen, C.; Stein, H.H. Effect of feeding schedule on apparent energy and amino acid digestibility by growing pigs. Anim. Feed Sci. Technol. 2007, 132, 94–102. [Google Scholar] [CrossRef]
- Xu, Y.; Curtasu, M.V.; Bendiks, Z.; Marco, M.L.; Nørskov., P.N.; Knudsen, K.E.B.; Hedemann, M.S.; Laerke, H.N. Effects of dietary fibre and protein content on intestinal fibre degradation, short-chain fatty acid and microbiota composition in a high-fat fructose-rich diet induced obese Gottingen Minipig model. Food Funct. 2020, 11, 10758–10773. [Google Scholar] [CrossRef] [PubMed]
- Glitsø, L.V.; Gruppen, H.; Schols, H.A.; Højsgaard, S.; Sandström, B.; Bach Knudsen, K.E. Degradation of rye arabinoxylans in the large intestine of pigs. J. Sci. Food Agric. 1999, 79, 961–969. [Google Scholar] [CrossRef]
- Jørgensen, H.; Serena, A.; Hedemann, M.S.; Bach Knudsen, K.E. The fermentative capacity of growing pigs and adult sows fed diets with contrasting type and level of dietary fibre. Livest. Sci. 2007, 109, 111–114. [Google Scholar] [CrossRef]
- Tai, M.M.; Castillo, P.; Pi-Sunyer, F.X. Meal size and frequency: Effect on the thermic effect of food. Am. J. Clin. Nutr. 1991, 54, 783–787. [Google Scholar] [CrossRef]
- Le Naou, T.; Le Floc’h, N.; Louveau, I.; van Milgen, J.; Gondret, F. Meal frequency changes the basal and time-course profiles of plasma nutrient concentrations and affects feed efficiency in young growing pigs. J. Anim. Sci. 2014, 92, 2008–2016. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Batterham, E.S.; Bayley, H.S. Effect of frequency of feeding of diets containing free or protein-bound lysine on the oxidation of [14C]lysine or [14C]phenylalanine by growing pigs. Br. J. Nutr. 1989, 62, 647–655. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Palmer, M.A.; Capra, S.; Baines, S.K. Association between eating frequency, weight, and health. Nutr. Rev. 2009, 67, 379–390. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Carlson, O.; Martin, B.; Stote, K.S.; Golden, E.; Maudsley, S.; Najjar, S.S.; Ferrucci, L.; Ingram, D.K.; Longo, D.L.; Rumpler, W.V.; et al. Impact of reduced meal frequency without caloric restriction on glucose regulation in healthy, normal-weight middle-aged men and women. Metab. Clin. Exp. 2007, 56, 1729–1734. [Google Scholar] [CrossRef] [Green Version]
- LeBlanc, J.; Diamond, P. Effect of meal size and frequency on postprandial thermogenesis in dogs. Am. J. Physiol. 1986, 250, E144–E147. [Google Scholar] [CrossRef] [PubMed]
- Fabry, P.; Hejl, Z.; Fodor, J.; Braun, T.; Zvolankova, K. The Frequency of Meals. Its Relation to Overweight, Hypercholesterolaemia, and Decreased Glucose-Tolerance. Lancet 1964, 2, 614–615. [Google Scholar] [CrossRef]
- Cohn, C. Feeding Frequency and Body Composition. Ann. N. Y. Acad. Sci. 1963, 110, 395–409. [Google Scholar] [CrossRef] [PubMed]
- Mohn, S.; Fuller, M.; Ball, R.; De Lange, C. Feeding frequency and type of isotope tracer do not affect direct estimates of lysine oxidation in growing pigs. J. Nutr. 2003, 133, 3504–3508. [Google Scholar] [CrossRef] [Green Version]
- Commission, C.A. Report of the 30th Session of the Codex Committee on Nutrition and Foods for Special Dietary Uses. In Proceedings of the 30th Session of the Codex Committee on Nutrition and Foods for Special Dietary Uses (CCNFSDU), Cape Town, South Africa, 3–7 November 2008. [Google Scholar]
- Bautil, A.; Courtin, C. Chapter 1 Fibres making up wheat cell walls in the context of broiler diets. In The Value of Fibre: Engaging the Second Brain for Animal Nutrition; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019; pp. 148–152. [Google Scholar]
- Cassidy, Y.M.; McSorley, E.M.; Allsopp, P.J. Effect of soluble dietary fibre on postprandial blood glucose response and its potential as a functional food ingredient. J. Funct. Foods 2018, 46, 423–439. [Google Scholar] [CrossRef]
- De Leeuw, J.A.; Jongbloed, A.W.; Spoolder, H.A.M.; Verstegen, M.W.A. Effects of hindgut fermentation of non-starch polysaccharides on the stability of blood glucose and insulin levels and physical activity in empty sows. Livest. Prod. Sci. 2005, 96, 165–174. [Google Scholar] [CrossRef]
- NRC. Nutrient Requirements of Swine, 11th ed.; National Academies Press: Washington, DC, USA, 2012. [Google Scholar] [CrossRef] [Green Version]
- Montoya, C.A.; Henare, S.J.; Rutherfurd, S.M.; Moughan, P.J. Potential misinterpretation of the nutritional value of dietary fiber: Correcting fiber digestibility values for nondietary gut-interfering material. Nutr. Rev. 2016, 74, 517–533. [Google Scholar] [CrossRef]
- Wenk, C. The role of dietary fibre in the digestive physiology of the pig. Anim. Feed Sci. Technol. 2001, 90, 21–33. [Google Scholar] [CrossRef]
- Englyst, H.N.; Quigley, M.E.; Hudson, G.J. Determination of Dietary Fiber as Nonstarch Polysaccharides with Gas-Liquid-Chromatographic, High-Performance Liquid-Chromatographic or Spectrophotometric Measurement of Constituent Sugars. Analyst 1994, 119, 1497–1509. [Google Scholar] [CrossRef] [PubMed]
- Bach Knudsen, K.E. Carbohydrate and lignin contents of plant materials used in animal feeding. Anim. Feed Sci. Technol. 1997, 67, 319. [Google Scholar] [CrossRef]
- Bach Knudsen, K.E.; Hedemann, M.S.; Lærke, H.N. The role of carbohydrates in intestinal health of pigs. Anim. Feed Sci. Technol. 2012, 173, 41–53. [Google Scholar] [CrossRef]
- Noblet, J.; Le Goff, G. Effect of dietary fibre on the energy value of feeds for pigs. Anim. Feed Sci. Technol. 2001, 90, 35–52. [Google Scholar] [CrossRef]
- Högberg, A.; Lindberg, J.E. Influence of cereal non-starch polysaccharides on digestion site and gut environment in growing pigs. Livest. Prod. Sci. 2004, 87, 121–130. [Google Scholar] [CrossRef]
- Le Goff, G.; Dubois, S.; Milgen, J.V.; Noblet, J. Influence of dietary fibre level on digestive and metabolic utilisation of energy in growing and finishing pigs. Anim. Res. 2002, 51, 245–259. [Google Scholar] [CrossRef] [Green Version]
- Yin, Y.L.; McEvoy, J.D.G.; Schulze, H.; Hennig, U.; Souffrant, W.B.; McCracken, K.J. Apparent digestibility (ileal and overall) of nutrients and endogenous nitrogen losses in growing pigs fed wheat (var. Soissons) or its by-products without or with xylanase supplementation. Livest. Prod. Sci. 2000, 62, 119–132. [Google Scholar] [CrossRef]
- Dikeman, C.L.; Fahey, G.C. Viscosity as related to dietary fiber: A review. Crit. Rev. Food Sci. Nutr. 2006, 46, 649–663. [Google Scholar] [CrossRef]
- Li, Y.; Wei, H.; Li, F.; Chen, S.; Duan, Y.; Guo, Q.; Liu, Y.; Yin, Y. Supplementation of branched-chain amino acids in protein-restricted diets modulates the expression levels of amino acid transporters and energy metabolism associated regulators in the adipose tissue of growing pigs. Anim. Nutr. 2016, 2, 24–32. [Google Scholar] [CrossRef]
- Mackie, A.; Rigby, N.; Harvey, P.; Bajka, B. Increasing dietary oat fibre decreases the permeability of intestinal mucus. J. Funct. Foods 2016, 26, 418–427. [Google Scholar] [CrossRef] [Green Version]
- Metzler-Zebeli, B.U.; Hooda, S.; Pieper, R.; Zijlstra, R.T.; van Kessel, A.G.; Mosenthin, R.; Ganzle, M.G. Nonstarch polysaccharides modulate bacterial microbiota, pathways for butyrate production, and abundance of pathogenic Escherichia coli in the pig gastrointestinal tract. Appl. Environ. Microbiol. 2010, 76, 3692–3701. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Raninen, K.; Lappi, J.; Mykkanen, H.; Poutanen, K. Dietary fiber type reflects physiological functionality: Comparison of grain fiber, inulin, and polydextrose. Nutr. Rev. 2011, 69, 9–21. [Google Scholar] [CrossRef] [PubMed]
- Morales, G.A.; Moyano, F.J.; Marquez, L. In vitro assessment of the effects of phytate and phytase on nitrogen and phosphorus bioaccessibility within fish digestive tract. Anim. Feed Sci. Technol. 2011, 170, 209–221. [Google Scholar] [CrossRef]
- Menezes-Blackburn, D.; Gabler, S.; Greiner, R. Performance of Seven Commercial Phytases in an in Vitro Simulation of Poultry Digestive Tract. J. Agric. Food Chem. 2015, 63, 6142–6149. [Google Scholar] [CrossRef] [PubMed]
- Cowieson, A.J.; Hruby, M.; Faurschou Isaksen, M. The effect of conditioning temperature and exogenous xylanase addition on the viscosity of wheat-based diets and the performance of broiler chickens. Br. Poult. Sci. 2005, 46, 717–724. [Google Scholar] [CrossRef]
- Rodehutscord, M.; Siegert, W. Chapter 9 Facts and thoughts on carbohydrase supplementation effects on amino acid digestibility in broiler chickens. In The Value of Fibre: Engaging the Second Brain for Animal Nutrition; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019; pp. 45–64. [Google Scholar]
- Svihus, B. Effect of digestive tract conditions, feed processing and ingredients on response to NSP enzymes. In Enzymes in Farm Animal Nutrition, 2nd ed.; CABI: Wallingford, UK, 2011; pp. 129–159. [Google Scholar]
- Nortey, T.N.; Patience, J.F.; Sands, J.S.; Trottier, N.L.; Zijlstra, R.T. Effects of xylanase supplementation on the apparent digestibility and digestible content of energy, amino acids, phosphorus, and calcium in wheat and wheat by-products from dry milling fed to grower pigs. J. Anim. Sci. 2008, 86, 3450–3464. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Widyaratne, G.P.; Patience, J.F.; Zijlstra, R.T. Effect of xylanase supplementation of diets containing wheat distiller’s dried grains with solubles on energy, amino acid and phosphorus digestibility and growth performance of grower-finisher pigs. Can. J. Anim. Sci. 2009, 89, 91–95. [Google Scholar] [CrossRef]
- Weiland, S.A.; Patience, J.F. Effect of xylanase supplementation on nutrient and energy digestibility at three time periods in growing pigs fed diets based on corn or corn distillers dried grains with solubles. Anim. Feed Sci. Technol. 2021, 276, 114929. [Google Scholar] [CrossRef]
- Laerke, H.N.; Arent, S.; Dalsgaard, S.; Bach Knudsen, K.E. Effect of xylanases on ileal viscosity, intestinal fiber modification, and apparent ileal fiber and nutrient digestibility of rye and wheat in growing pigs. J. Anim. Sci. 2015, 93, 4323–4335. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lagos, L.V.; Lee, S.A.; Bedford, M.R.; Stein, H.H. Reduced concentrations of limestone and monocalcium phosphate in diets without or with microbial phytase did not influence gastric pH, fecal score, or growth performance, but reduced bone ash and serum albumin in weanling pigs. Transl. Anim. Sci. 2021, 5, txab115. [Google Scholar] [CrossRef] [PubMed]
- Ange, K.D.; Eisemann, J.H.; Argenzio, R.A.; Almond, G.W.; Blikslager, A.T. Effects of feed physical form and buffering solutes on water disappearance and proximal stomach pH in swine. J. Anim. Sci. 2000, 78, 2344–2352. [Google Scholar] [CrossRef] [PubMed]
- Morgan, N.K.; Wallace, A.; Bedford, M.R.; Choct, M. Efficiency of xylanases from families 10 and 11 in production of xylo-oligosaccharides from wheat arabinoxylans. Carbohydr. Polym. 2017, 167, 290–296. [Google Scholar] [CrossRef] [PubMed]
- Pace, C.N.; Grimsley, G.R.; Scholtz, J.M. Protein ionizable groups: pK values and their contribution to protein stability and solubility. J. Biol. Chem. 2009, 284, 13285–13289. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Strube, M.L.; Meyer, A.S.; Boye, M. Mini Review: Basic Physiology and Factors Influencing Exogenous Enzymes Activity in the Porcine Gastrointestinal Tract. Anim. Nutr. Feed Technol. 2013, 13, 441–459. [Google Scholar]
- Wilfart, A.; Montagne, L.; Simmins, H.; Noblet, J.; van Milgen, J. Effect of fibre content in the diet on the mean retention time in different segments of the digestive tract in growing pigs. Livest. Sci. 2007, 109, 27–29. [Google Scholar] [CrossRef]
- Metzler-Zebeli, B.U.; Vahjen, W.; Baumgartel, T.; Rodehutscord, M.; Mosenthin, R. Ileal microbiota of growing pigs fed different dietary calcium phosphate levels and phytase content and subjected to ileal pectin infusion. J. Anim. Sci. 2010, 88, 147–158. [Google Scholar] [CrossRef] [Green Version]
- Black, J.; Ratanpaul, V.; Williams, B.; Diffey, S.; Gidley, M. Chapter 2 Variability in cereal grain composition and nutritional value: The importance of fibre. In The Value of Fibre: Engaging the Second Brain for Animal Nutrition; Wageningen Academic Publishers: Wageningen, The Netherlands, 2019; pp. 3195–3202. [Google Scholar]



{kind=link}
{kind=link}
| Hormone | Secretion Site | Location | Effects |
|---|---|---|---|
| Ghrelin | X- and A-like cells | Stomach and duodenum | Increases meal size Energy homeostasis Release of growth hormone Increases gastric movement |
| CCK | I cells | Stomach and duodenum | Decreasing meal size Stimulates gallbladder contractions Reduces gastric emptying and intestinal motility |
| GIP | K cells | Duodenum | Stimulates insulin release and fat storage Optimises nutrient delivery to tissues |
| GLP-1 | L cells | Ileum and colon | Decreases feed intake Stimulates insulin secretion Reduction of intestinal motility Decreases gastric emptying |
| PYY | L cells | Ileum and colon | Regulates energy homeostasis Decreases gastric emptying and intestinal motility |
| Apo A-IV | In the jejunum | Jejunum | Inhibits intestinal motility Reduces feed intake |
| Leptin | Adipocytes | Adipose tissue | Regulates energy balance High levels reduce feed intake |
| Meal Type | Effect on MRT | Effect on Digestibility and Fermentation | Effect on Metabolism | References | |
|---|---|---|---|---|---|
| Meal size | Large meal | Reduced MRT | Decreased digestibility | Roth and Kirchgessner, 1985 [87] | |
| Ad libitum | Reduced fermentation | Chastanet et al., 2007 [93] | |||
| Fibre degraded from distal ileum to mid-colon Depends on AX structure | Xu et al., 2020 [94] Glitsø et al., 1999 [95] | ||||
| Meal frequency | Increased number of meals per day | Continuous flow of digesta | Secretions: Amylase ↑ Trypsin ↑ Chymotrypsin ↑ | Hee et al., 1988 [10] | |
| ATTD of Ca, Trp, Ile, Cys, Arg, Phe and phytic acid ↑ | Mroz et al., 1994 [90] | ||||
| ATTD of CP ↑ | Pepsin secretions ↑ | Jia et al., 2021 [91] | |||
| Reduced fluctuation of gastric emptying | Constant rate of glucose and amino acids digestion | Blood glucose ↓ Insulin fluctuation ↓ | Smeets et al., 2008 [50] Palmer et al., 2009 [100] | ||
| Lipogenesis ↑ | Smeets et al., 2008 [50] Tai et al., 1991 [97] | ||||
| Limited number of meals per day | Reduced motility of intestines and stomach | Ruckenbusch et al., 1976 [92] | |||
| Lipogenesis ↑ | LeBlanc et al., 1986 [102] |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Chassé, É.; Guay, F.; Bach Knudsen, K.E.; Zijlstra, R.T.; Létourneau-Montminy, M.-P. Toward Precise Nutrient Value of Feed in Growing Pigs: Effect of Meal Size, Frequency and Dietary Fibre on Nutrient Utilisation. Animals 2021, 11, 2598. https://doi.org/10.3390/ani11092598
Chassé É, Guay F, Bach Knudsen KE, Zijlstra RT, Létourneau-Montminy M-P. Toward Precise Nutrient Value of Feed in Growing Pigs: Effect of Meal Size, Frequency and Dietary Fibre on Nutrient Utilisation. Animals. 2021; 11(9):2598. https://doi.org/10.3390/ani11092598
Chicago/Turabian StyleChassé, Élisabeth, Frédéric Guay, Knud Erik Bach Knudsen, Ruurd T. Zijlstra, and Marie-Pierre Létourneau-Montminy. 2021. "Toward Precise Nutrient Value of Feed in Growing Pigs: Effect of Meal Size, Frequency and Dietary Fibre on Nutrient Utilisation" Animals 11, no. 9: 2598. https://doi.org/10.3390/ani11092598