
Supporting Zoo Asian Elephant (Elephas maximus) Welfare and Herd Dynamics with a More Complex and Expanded Habitat

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals
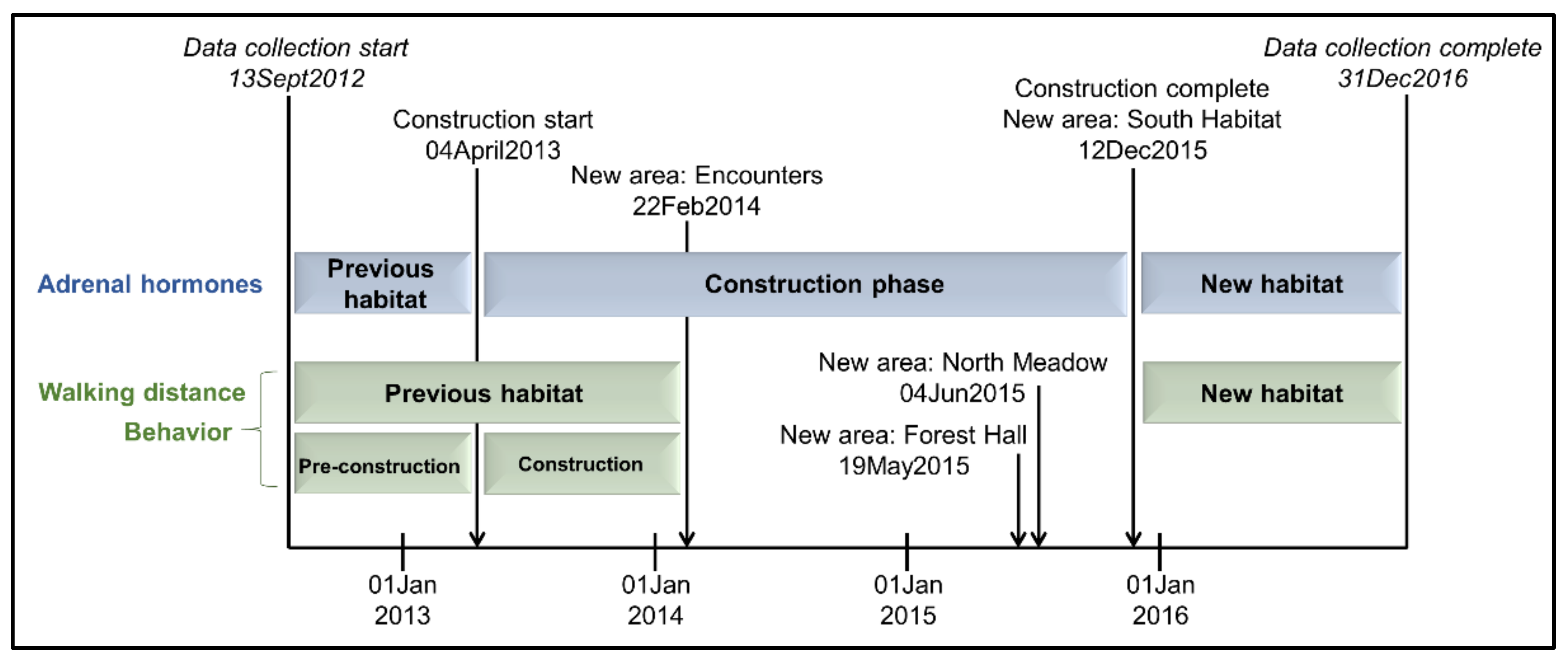
2.2. Study Timeline
2.3. Habitat Descriptions
2.4. Data Analysis
2.5. Walking Distance and Space Use
2.6. Reproductive and Adrenal Hormones
2.7. Behavior and Resource Use
3. Results
3.1. Walking Distance
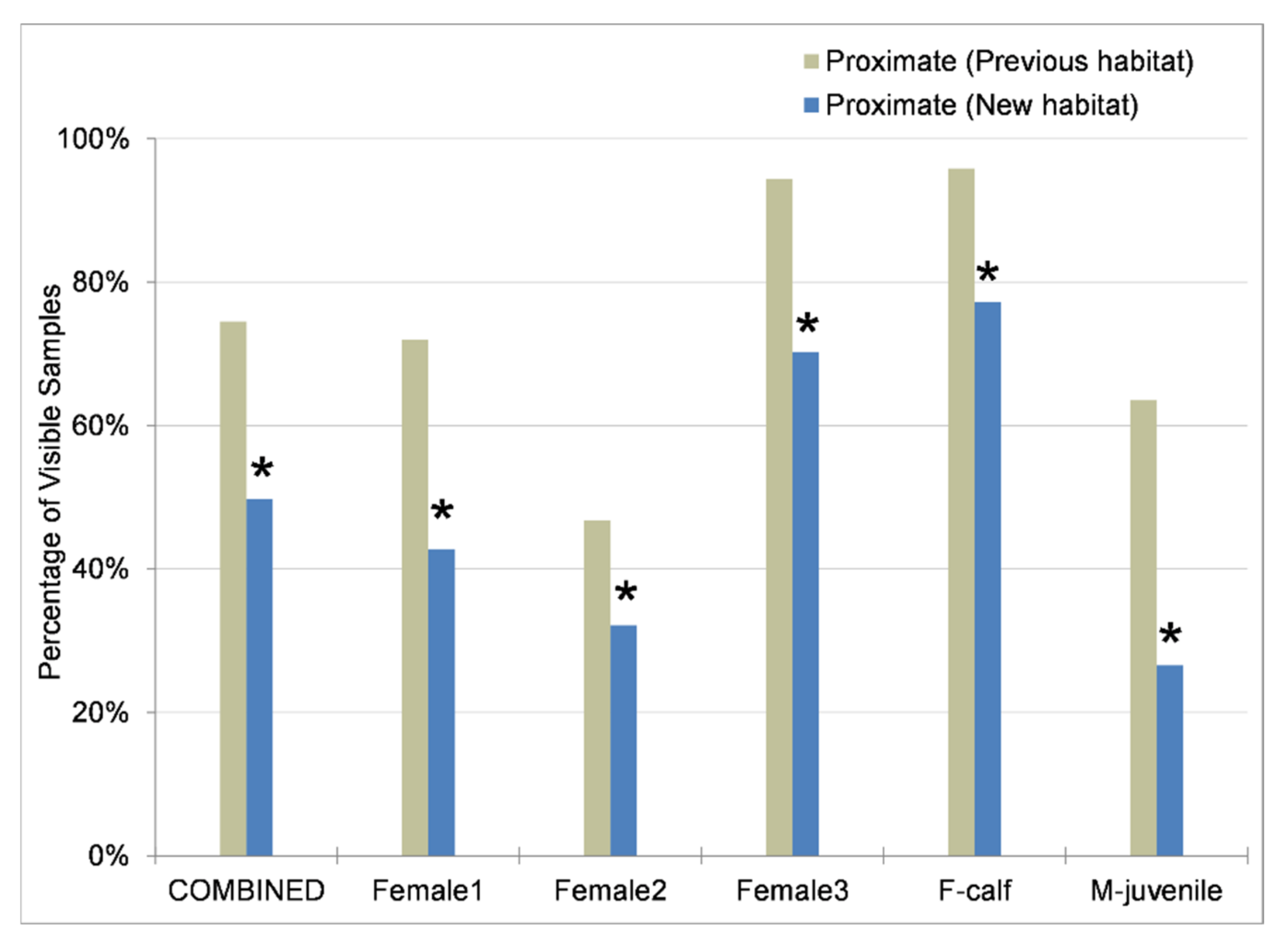
3.2. Space Use
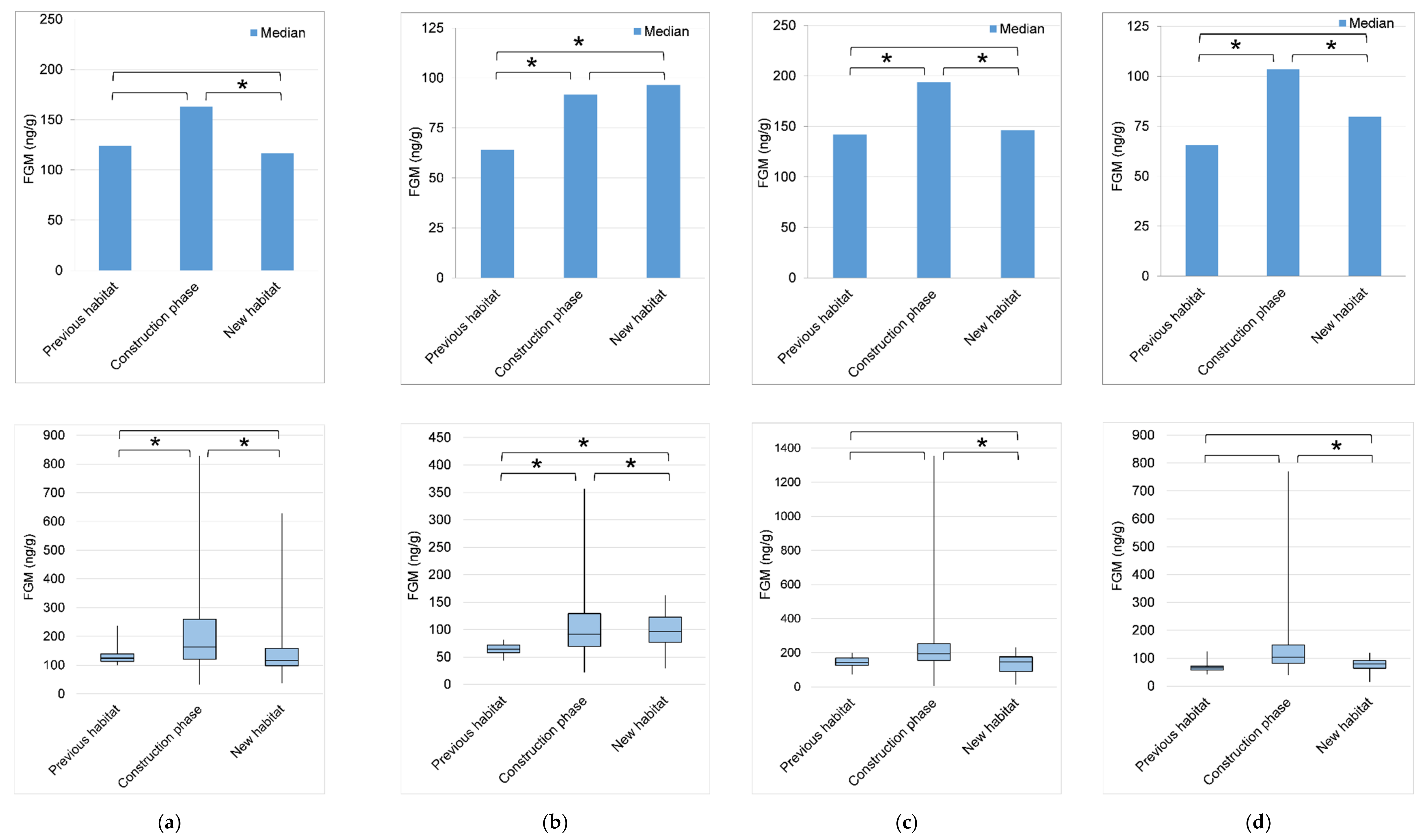
3.3. Reproductive and Adrenal Hormones
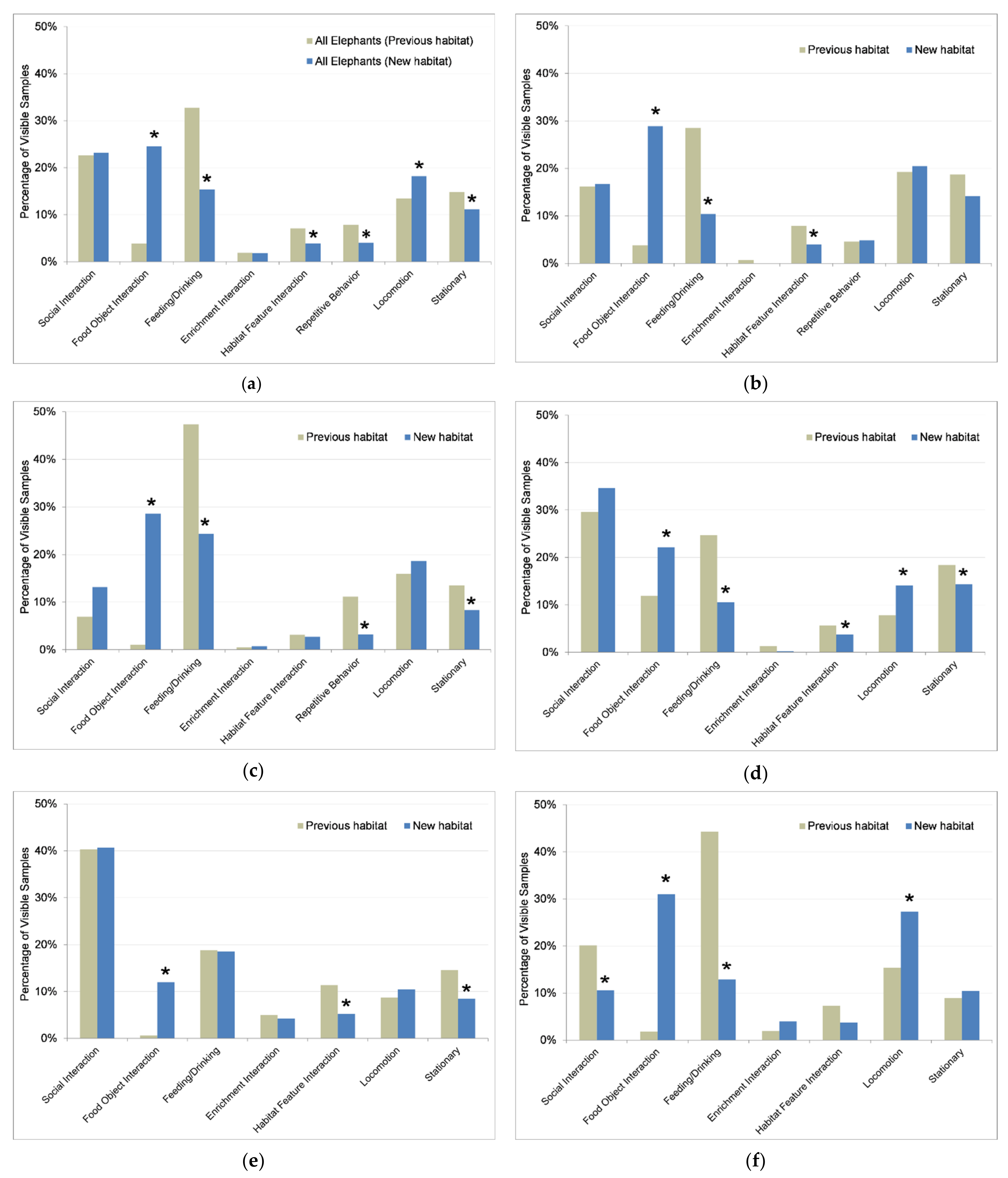
3.4. Behavior
3.4.1. Activity Budgets
3.4.2. Behavior of Individuals
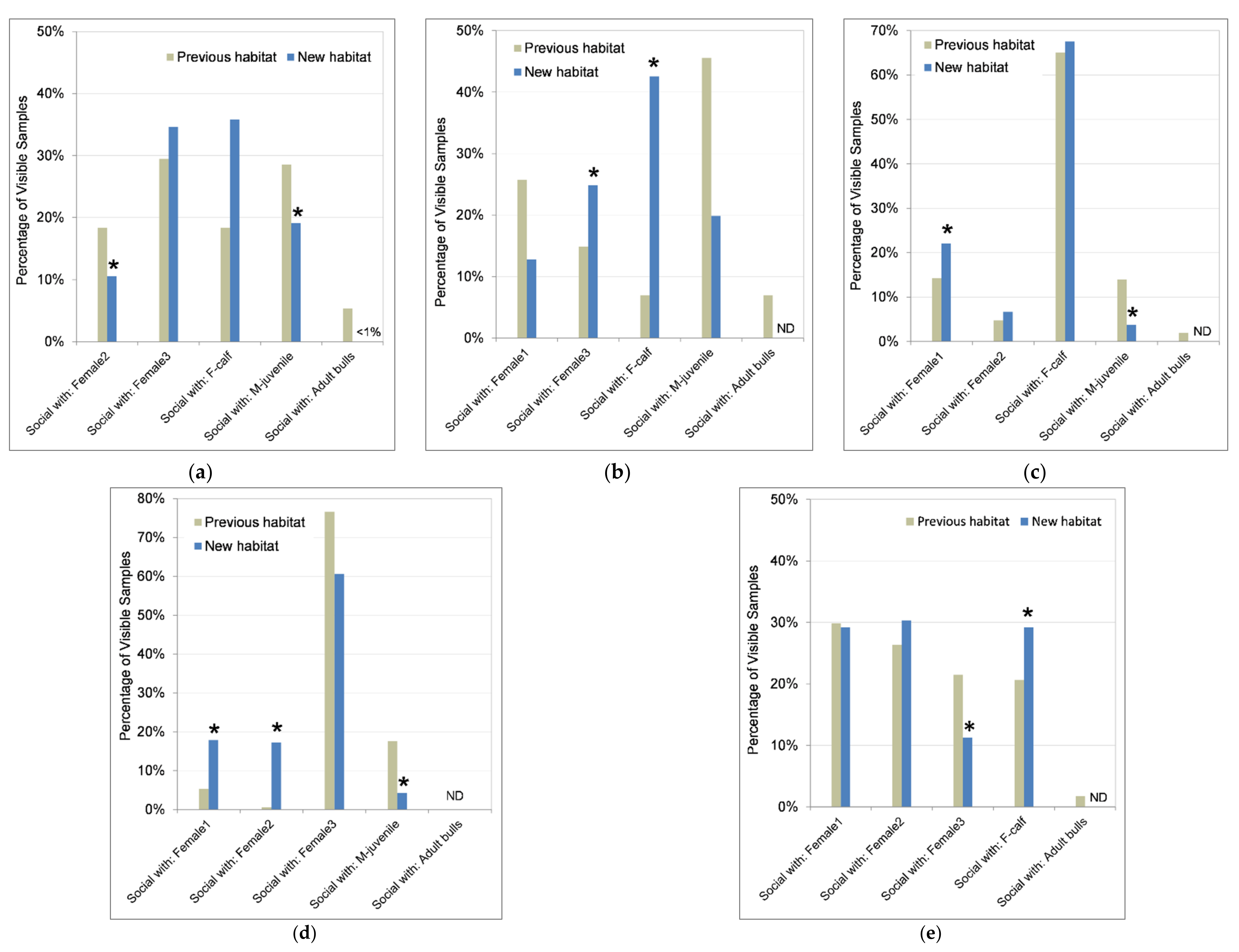
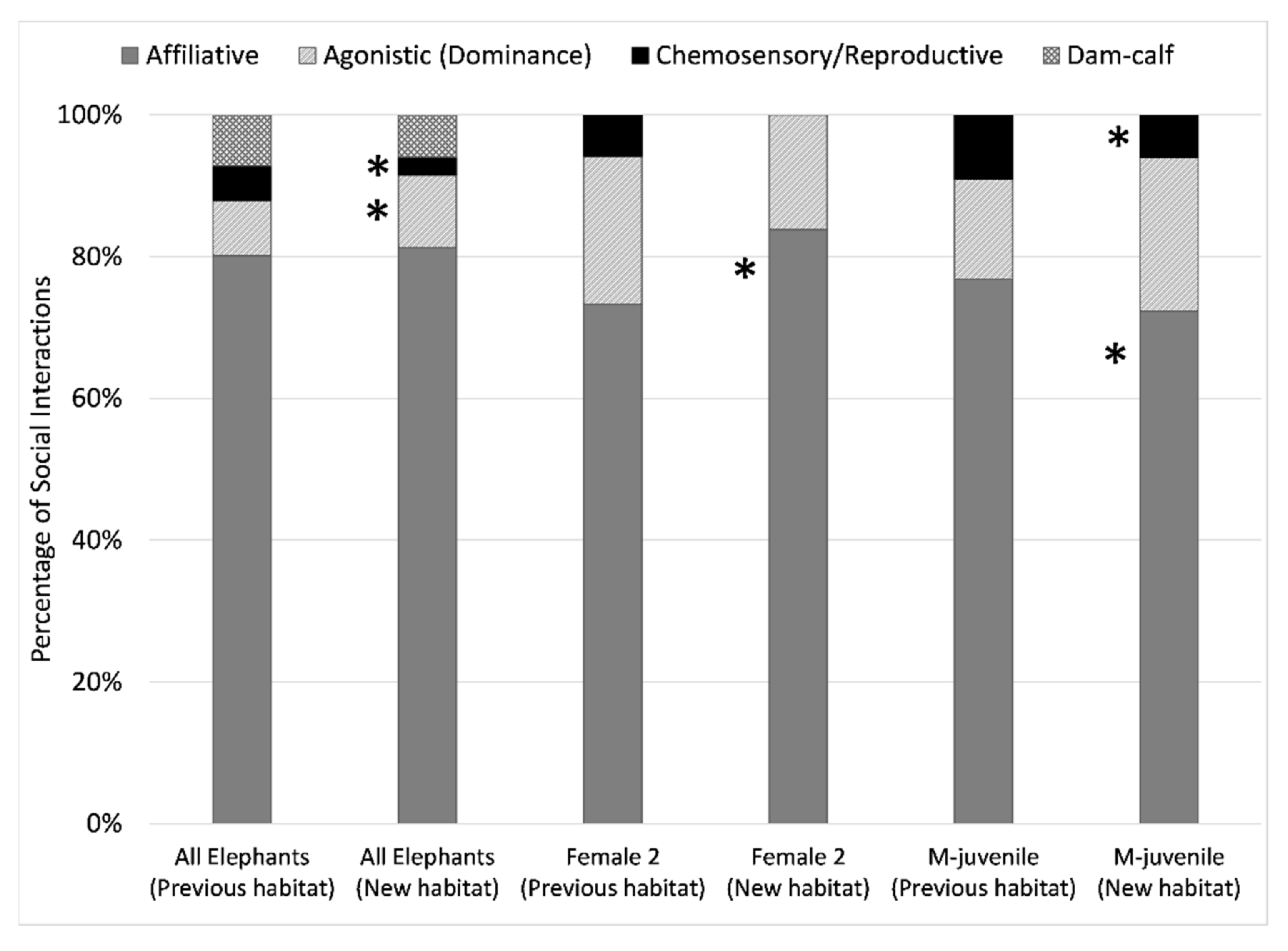
3.4.3. Social Behaviors and Social Partners
3.4.4. Behavior Changes during Construction Prior to Habitat Changes
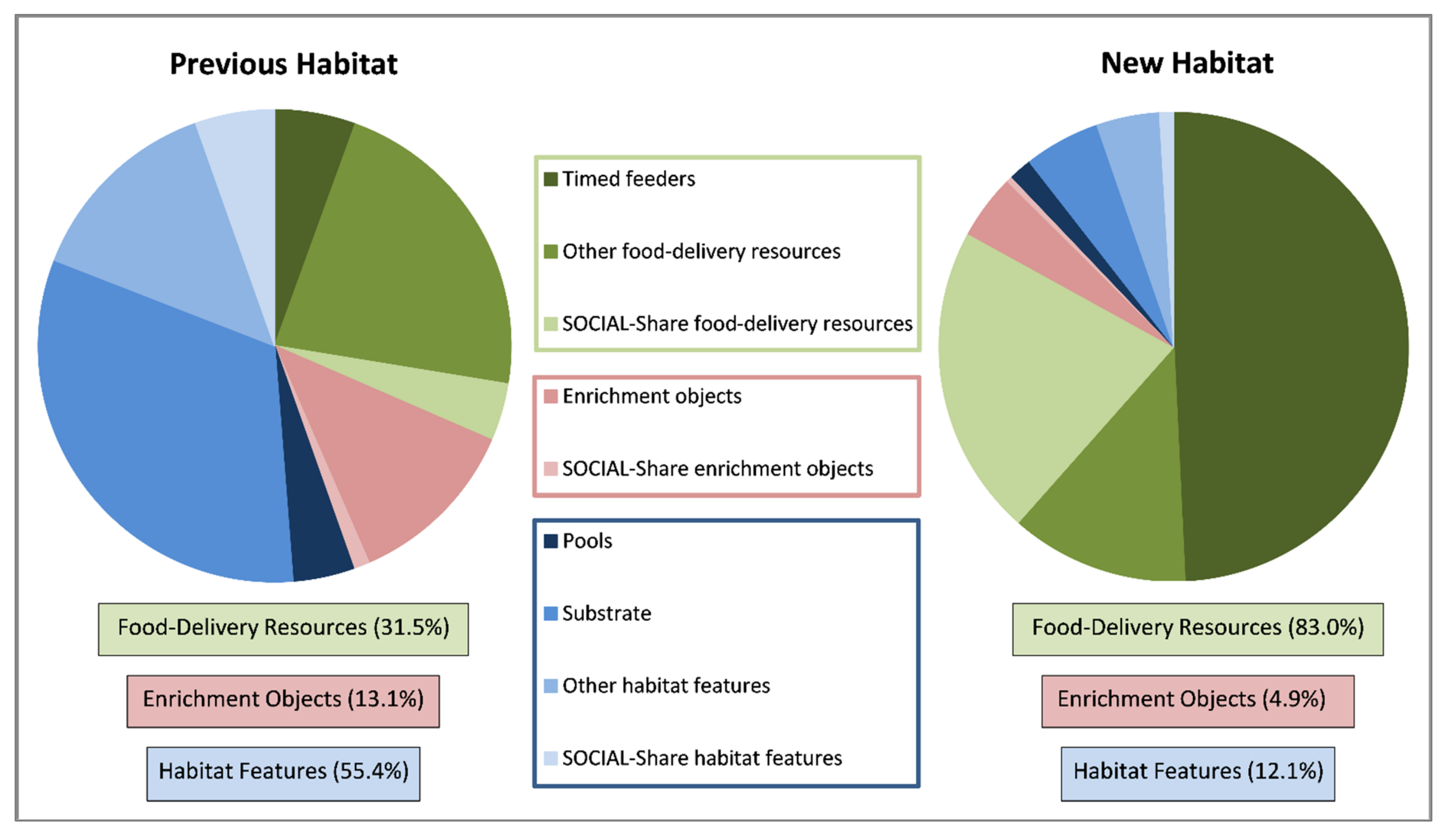
3.5. Resource Use
4. Discussion
4.1. Walking Distance
4.2. Reproductive and Adrenal Hormones
4.3. Behavior
4.3.1. Activity Levels and Locomotion
4.3.2. Foraging, Feeding, and Exploratory Behaviors
4.3.3. Repetitive Behaviors
4.3.4. Social Behaviors and Partners
4.3.5. Behavioral Changes Associated with Construction
4.3.6. Space and Resource Use
4.4. Environmental Complexity and Provisions for Choice and Control
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Informed Consent Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Appendix A. Elephant Behavior Ethogram (Full Version)

{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Proximity | Definition |
| Proximate | Focal animal is within 2 body lengths of or in contact with another individual (defined as 2 adult female body lengths, c.a. 10 m) |
| Proximity Not Visible | Focal animal or other elephants are not visible enough to determine proximity. |
| Behavior Category | Definition |
| Behavior Not Visible | Elephant or activity is not visible enough to determine the behavior. |
| Social Interaction | Interacting with another elephant in a social context, either with physical contact (e.g., trunk twine, play) or without physical contact but within 2 body lengths (e.g., sharing food, displace). LEVEL 2: Social behavior |
| Food-Delivery Object Interaction | Interacting with any object that distributes food, either permanent or provided by caretakers. Behavior includes both seeking food and feeding: Investigating food-delivery object with trunk obvious attempts to reach and manipulate the food-delivery object (e.g., the object is in a position that is challenging to reach), manipulating and putting food item in the mouth before/after interacting with the object. Food Object vs. Enrichment objects: For items that can be either (e.g., barrel), determined by whether the elephant appears to be eating before/after interacting with the object. Food Object vs. Feeding: Determined by whether the elephant must interact with the food-delivery object to get food (e.g., hay net) or whether food is available without interaction with the object. MODIFIER: Food-Delivery Object |
| Feeding/Drinking | Acquiring/gathering and consuming (putting in the mouth) any food item without interacting with a food-delivery object. Drinking water, not bathing. Feeding: Requires the elephant to put something in its mouth; manipulating alone and chewing alone is not sufficient. LEVEL 2: Activity level (stationary, locomoting) MODIFIER: Food/water source |
| Enrichment (non-food) Object Interaction | Interacting objects that do not provision food and are not permanent (e.g., firehose ball). MODIFIER: Enrichment Object |
| Habitat Feature Interaction | Interacting with features that are permanent in the habitat (e.g., dusting, bathing, digging, rubbing body, climbing on logs, investigating features with feet or mouth). MODIFIER: Habitat Feature |
| Repetitive Behaviors (Stereotypy) | Motor: Repeatedly performing a behavior for 3 or more consecutive repetitions without interruption (e.g., route tracing). Non motor: Performing the behavior for 5 s or longer. Stereotypy: Determined by lack of purpose. If a behavior appears to have a purpose (e.g., scooping small food items in a repetitive manner), it may fit in a different category LEVEL 2: Major form of repetitive behavior |
| Locomotion | Walking or running (fast walking) more than 2 body lengths in any direction without stopping for 3 s or longer. |
| Stationary | Any stationary state without engaging in another behavior for 3 s or longer. Standing, walking or shuffling (without moving 2 body lengths), sitting, kneeling, lying (prone or sternal). LEVEL 2: Stationary body position (upright, lay down) MODIFIER: Location |
| Function | Behavior | Definition |
|---|---|---|
| Affiliative | Trunk to: head area, trunk, body, front legs, mammary glands | Extend trunk tip to (within 6 inch) or visibly touches with trunk. MODIFIER: Sender/Receiver |
| Trunk rub | Move trunk back and forth over body of another elephant (not for purpose of taking food atop the body). MODIFIER: Sender/Receiver | |
| Trunk twine | Mutual wrapping of trunks. MODIFIER: Partner(s) | |
| Tail tuck | Grab tail with trunk and tuck under front leg. MODIFIER: Sender/Receiver | |
| Lean/Rub | Lean or rest against or rub against another elephant while standing, sitting, or lying (includes under another elephant). Behavior includes: one elephant lying entirely or partially under another (e.g., calf lying under mom/auntie, elephants lying against each other, one elephant standing partially over another laying down. MODIFIER: Partner(s) | |
| Nudge | Gentle head-to-head, head-to-body, body-to-body contact initiated by one elephant towards another. MODIFIER: Sender/Receiver | |
| Greeting | Coming together, trunk tip to palatal pit (crease of mouth) with reciprocation. May include ear flapping, trunk entwining, arousal. MODIFIER: Partner(s) | |
| Play (Play contact, Play no contact, Play solicit, Self-play) | Play-contact: Sparring (pushing trunks, tusking, shoving, wrestling), back and forth nudging, trunk wrestling, chasing, rolling, climbing on conspecific (at least one foot on the head or body of another elephant). Play-no contact: Clear play with a conspecific (e.g., ball kicking, head wagging, splashing); play soliciting with approach and “run” away or other behaviors that seem to solicit play; other behaviors that seem like play but another elephant is not visible (e.g., sits on rump and throw back head, chasing wildlife, sliding on the mud). MODIFIER: Partner(s) or Unknown (no contact) | |
| Follow or Approach | Follow: Walk along path of conspecific with someone in the lead. Distance between animals almost constant. Approach: Move to within proximity (1 body length) of conspecific Sender = animal that is taking action—animal that is following or approaching. Receiver = animal being followed/approached Approach vs. Displace vs. Claim food/water/object: Depends on the sequence of events. MODIFIER: Sender/Receiver | |
| Share food/water | Simultaneously interacting with the same food-delivery object or feeding from the same food source, pushing food toward another, or drinking from the same water source. Requires that focal is proximate and has line of sight with partners. Behavior includes: Consuming food, sharing space around a food object even if unable to reach the object. Behavior does not include: Interacting with a food object when the elephants cannot see each other (e.g., on opposite sides of the enrichment tree) or bathing from the same water source. MODIFIER: Partner(s) | |
| Share enrichment object | Simultaneously manipulating same enrichment object. MODIFIER: Partner(s) | |
| Share habitat feature | Simultaneously interacting with the same habitat feature. MODIFIER: Partner(s) | |
| Agonistic | Claim-food/object | Approach and take food or object w/in other elephant’s reach, on its body, or in its mouth. Includes displacing from a feeding/drinking source when the other elephant is still eating/drinking. MODIFIER: Sender/Receiver |
| Displace-location | Approach and overtake position of conspecific, receiver moves at least 1 body length. Does not require physical contact. Behavior does not include: Pushing that results in displacement. MODIFIER: Sender/Receiver | |
| Block, resource hold | Place body between a resource and an approaching elephant. Holder may turn rump towards approaching elephant. MODIFIER: Sender/Receiver | |
| Strike or Push | Strike: Forceful body contact initiated by one elephant, including head butt (hit with forehead), trunk strike (hit with trunk), and kick (kick with any foot) Push: Forceful head-to-head, head-to-body, body-to-body contact that typically results in receiver being displaced, or at least it appears there is intent to displace or control. MODIFIER: Sender/Receiver/Partner (if mutual) | |
| Sparring | Mutual striking or pushing with head and trunk, tusking, driving. Sparring vs. Playing: Depends on the partner and appearance of aggression. MODIFIER: Partner | |
| Backup to | Backup rump first towards another individual’s head, side, or rump, sometimes extending tail to touch (moving towards another in a submissive manner). May end in a Rump Present. MODIFIER: Sender/Receiver | |
| Rump present | Back rump under the chin of another (submissive or reproductive, depending on partner and behavior sequence). MODIFIER: Sender/Receiver | |
| Tail pull | Hold tail with trunk and pull or twist. MODIFIER: Sender/Receiver | |
| Drive | Use head, tusks, or trunk to push the rear of another, maintaining contact while both elephants move. Drive vs. Pre-mount: Determined by whether the trunk of the sender is over the back of receiver with an attempt to get in mounting position. MODIFIER: Sender/Receiver | |
| Threat | Ears wide, trunk forward, head raised; may include foot scraping, twitching the tail, head shaking, or weaving. MODIFIER: Sender/Receiver | |
| Charge, mock | Rapidly approach another animal, ears erect, head high, tail extended, trunk sometimes extended. Stops short of contact. Often trumpets. Stops before contact, often in freeze position. MODIFIER: Sender/Receiver | |
| Charge, real | Rapidly approach another animal or object, ears usually close to head, head high, trunk tucked under head. Often silent. Attempts contact (strike, push). MODIFIER: Sender/Receiver | |
| Chase to retreat | Sender rapidly approaches receiver and receiver retreats. MODIFIER: Sender/Receiver | |
| Intervene | Place body between the sender and receiver in an agonistic interaction (typically by a dominant elephant). | |
| Arousal | Spinning U/D–group | Spinning, urinating (U), defecating (D) as a group (usually in close proximity) or by one’s self with no contact. May include Arousal-other. MODIFIER: Partner(s), if any |
| Pause/freeze, alert posture | Pause/freeze: No body movement, absolute stillness, typically with ears erect, sometimes w/ forefoot raised and still. Alert posture: Head raised, ears spread, tail raised, trunk raised. | |
| Defensive circle | Group forms defense circle around an individual MODIFIER: Partners | |
| Arousal-other | Foot scrape, tail erect (parallel to ground), tail up, trunk slap on ground; may include “running” (in the presence of other elephants). Does not include: “running” during play or with no social context or “running” with an enrichment object. | |
| Chemosensory | Trunk to: vulva, penis, anus, back legs, temporal gland | Extend trunk to within 6 inch or visibly touches with trunk. MODIFIER: Sender/Receiver/Partner (if mutual) |
| Trunk to: urine, feces | Extend trunk tip to (within 6 inch) or visibly touches with trunk the urine or feces, either from self or another elephant. | |
| Flehmen | Place dorsal trunk finger onto roof of mouth (VNO openings). | |
| Reproductive | Rump present | Back rump under chin of another (reproductive or submissive depending on partner and behavior sequence). MODIFIER: Sender/Receiver |
| Pre-mount Mount | Pre-mounting (chasing, driving with trunk over back) or mounting behaviors. MODIFIER: Sender/Receiver | |
| Dam-calf | Nurse Attempted nurse | Nursing (dam stands for calf to suckle) or attempt nursing (dam does not move front leg forward or dam moves it leg back to block a teat). |
| Claim calf | Secure space around the calf by physically moving the calf away from another elephant or blocking access to the calf. Appears like an act of protecting or owning. |
| Activity Level | Definition |
|---|---|
| Locomotion | Feeding/drinking while locomoting |
| Stationary | Feeding/drinking while stationary |
| Major Form | Sub-Form | Definition and Examples |
|---|---|---|
| Whole-body stereotypy motor: 3 reps | Limb-swinging | Swinging the trunk or legs in a consistent pattern without body movement. |
| Forward, backward, or vertical movement | Forward, backward, or vertical body movements along the axis parallel to the elephant’s spine. Does not involve travel. Rocking: Body and head move in unison in a forward/backward motion with little vertical movement of the head. Bobbing: Head moves in a vertical motion and may involve turning the chin from shoulder to shoulder. Bobbing typically corresponds with vertical elevation and depression of the spinal column between the shoulder blades and the pelvic girdle. | |
| Side to side movement | Body movement occurs along the axis horizontally perpendicular to the elephant’s spine. Swaying: Body and head move counter to each other, causing the body to undulate horizontally. Weaving: Body undulates side to side while the head is lifted or twisted vertically. | |
| Locomotor stereotypy motor: 3 reps | Locomotor movement | Repeatedly walking the same pattern without leaving the path/route or without performing a competing behavior. Pacing: Following a relatively straight path in a forward or backward direction with patterned turns at each end of the path. Route tracing: Following a specific route. Route tracing patterns are more circuitous and longer than the straight paths associated with pacing. |
| Self-directed stereotypy motor: 3 reps non-motor: 5 s | Self-directed behavior | Self-stimulating behaviors. Limb or head banging: Striking one’s limb or head against objects in a non-species typical manner. Nipple pulling: Using the trunk to pull or manipulate one’s own nipples. |
| Oral stereotypy non-motor: 5 s | Oral behavior | Trunk sucking or “thumb sucking”: Sucking or chewing one’s own trunk Bar biting: Taking a bar into the mouth and biting or sucking on it. Chain chewing: Taking a chain or cable into the mouth and biting or sucking on it |
| Position | Definition |
|---|---|
| Stationary state-Upright | Standing, waling/shuffling, sitting, kneeling. |
| Stationary state-Lay down | Lying (prone or sternal). Sternum or side is in contact with substrate; can vary from curled up to stretched out. Behavior does not include: Lying under another elephant, which is social |
| Location |
|---|
| Gate or door |
| Under shade structure |
| Other location |
| Food-Delivery Object | Enrichment Object | Habitat Feature |
|---|---|---|
| Timed feeder | Firehose ball | Pool—large |
| Enrichment tree | Buckets of snow | Pool—wading |
| Hanging hay net | Log, small | Large logs or rocks |
| Cement herd feeder with food | Tire, small | Tire, large |
| Spinner/pipe puzzle with food | Spinner/pipe puzzle | Cement herd feeders (to climb) |
| Boomer ball with food | Boomer ball | Habitat enclosure (walls, gates, doors, poles) |
| Barrel/keg with food | Barrel/keg | Substrate (dirt/mud, sand, gravel, straw, cedar shavings) |
References
- Binding, S.; Farmer, H.; Krusin, L.; Cronin, K. Status of animal welfare research in zoos and aquariums: Where are we, where to next? J. Zoo Aquar. Res. 2020, 8, 166–174. [Google Scholar] [CrossRef]
- Escobar-Ibarra, I.; Mota-Rojas, D.; Gual-Sill, F.; Sánchez, C.R.; Baschetto, F.; Alonso-Spilsbury, M. Conservation, animal behaviour, and human-animal relationship in zoos. Why is animal welfare so important? J. Anim. Behav. Biometeorol. 2020, 9, 1–17. [Google Scholar]
- AZA. The Accreditation Standards & Related Policies; Association of Zoos and Aquariums (AZA): Silver Spring, MD, USA, 2020. [Google Scholar]
- BIAZA. British and Irish Association of Zoos and Aquariums (BIAZA) Animal Welfare. Available online: https://biaza.org.uk/animal-welfare (accessed on 1 August 2021).
- EAZA. European Association of Zoos and Aquaria (EAZA) Animal Welfare. Available online: https://www.eaza.net/about-us/areas-of-activity/animal-welfare/ (accessed on 1 August 2021).
- SEAZA. Southeast Asian Zoos and Aquariums Association (SEAZA) Standard on Animal Welfare. Available online: http://www.seaza.asia/animal-welfare-and-ethics/ (accessed on 1 August 2021).
- WAZA. World Association of Zoos and Aquariums (WAZA) Approach to Animal Welfare. Available online: https://www.waza.org/priorities/animal-welfare/our-approach-to-animal-welfare/ (accessed on 1 August 2021).
- Veasey, J.S. In pursuit of peak animal welfare; the need to prioritize the meaningful over the measurable. Zoo Biol. 2017, 36, 413–425. [Google Scholar] [CrossRef] [PubMed]
- Meehan, C.L.; Mench, J.A.; Carlstead, K.; Hogan, J.N. Determining Connections between the Daily Lives of Zoo Elephants and Their Welfare: An Epidemiological Approach. PLoS ONE 2016, 11, e0158124. [Google Scholar] [CrossRef] [Green Version]
- Greco, B.J.; Meehan, C.L.; Hogan, J.N.; Leighty, K.A.; Mellen, J.; Mason, G.J.; Mench, J.A. The Days and Nights of Zoo Elephants: Using Epidemiology to Better Understand Stereotypic Behavior of African Elephants (Loxodonta africana) and Asian Elephants (Elephas maximus) in North American Zoos. PLoS ONE 2016, 11, e0144276. [Google Scholar] [CrossRef]
- Brown, J.L.; Paris, S.; Prado-Oviedo, N.A.; Meehan, C.L.; Hogan, J.N.; Morfeld, K.A.; Carlstead, K. Reproductive Health Assessment of Female Elephants in North American Zoos and Association of Husbandry Practices with Reproductive Dysfunction in African Elephants (Loxodonta africana). PLoS ONE 2016, 11, e0145673. [Google Scholar] [CrossRef]
- Miller, M.A.; Hogan, J.N.; Meehan, C.L. Housing and Demographic Risk Factors Impacting Foot and Musculoskeletal Health in African Elephants [Loxodonta africana] and Asian Elephants [Elephas maximus] in North American Zoos. PLoS ONE 2016, 11, e0155223. [Google Scholar] [CrossRef] [PubMed]
- Morfeld, K.A.; Meehan, C.L.; Hogan, J.N.; Brown, J.L. Assessment of Body Condition in African (Loxodonta africana) and Asian (Elephas maximus) Elephants in North American Zoos and Management Practices Associated with High Body Condition Scores. PLoS ONE 2016, 11, e0155146. [Google Scholar] [CrossRef]
- McKay, G.M. Behavior and Ecology of the Asiatic Elephant in Southeastern Ceylon. In Smithsonian Contributions to Zoology; Smithsonian Institution Press: Washington, DC, USA, 1973; Volume 125, pp. 1–113. [Google Scholar]
- Evans, K.E.; Harris, S. Adolescence in male African elephants, Loxodonta africana, and the importance of sociality. Anim. Behav. 2008, 76, 779–787. [Google Scholar] [CrossRef]
- Rees, P.A. Some Preliminary Evidence of the Social Facilitation of Mounting Behavior in a Juvenile Bull Asian Elephant (Elephas maximus). J. Appl. Anim. Welf. Sci. 2004, 7, 49–58. [Google Scholar] [CrossRef]
- Sukumar, R. The Living Elephants: Evolutionary Ecology, Behavior, and Conservation; Oxford University Press: New York, NY, USA, 2003; 478p. [Google Scholar]
- Garai, M.; Kurt, F. The importance of socialisation to the well being of elephants. Z. Des. Kölner Zoo 2006, 2, 85–102. [Google Scholar]
- Eisenberg, J.F.; McKay, G.M.; Jainudeen, M.R. Reproductive behavior of the Asiatic elephant (Elephas maximus maximus L.). Behaviour 1971, 38, 193–225. [Google Scholar] [CrossRef] [PubMed]
- Clark, F. Cognitive Enrichment and Welfare: Current Approaches and Future Directions. Anim. Behav. Cogn. 2017, 4, 52–71. [Google Scholar] [CrossRef] [Green Version]
- Meehan, C.L.; Mench, J.A. The challenge of challenge: Can problem solving opportunities enhance animal welfare? Appl. Anim. Behav. Sci. 2007, 102, 246–261. [Google Scholar] [CrossRef]
- Buchanan-Smith, H.M.; Badihi, I. The psychology of control: Effects of control over supplementary light on welfare of marmosets. Appl. Anim. Behav. Sci. 2012, 137, 166–174. [Google Scholar] [CrossRef]
- Owen, M.A.; Swaisgood, R.R.; Czekala, N.M.; Lindburg, D.G. Enclosure Choice and Well-Being in Giant Pandas: Is It All About Control? Zoo Biol. 2005, 24, 475–481. [Google Scholar] [CrossRef]
- Sambrook, T.D.; Buchanan-Smith, H.M. Control and Complexity in Novel Object Enrichment. Anim. Welf. 1997, 6, 207–216. [Google Scholar]
- Buchanan-Smith, H.M. Environmental enrichment for primates in laboratories. Adv. Sci. Res. 2011, 5, 41–56. [Google Scholar] [CrossRef] [Green Version]
- Swaisgood, R.; Shepherdson, D. Environmental Enrichment as a Strategy for Mitigating Stereotypies in Zoo Animals: A Literature Review and Meta-analysis. In Stereotypic Animal Behaviour: Fundamentals and Applications to Welfare; Mason, G., Rushen, J., Eds.; CABI: Wallingford, UK, 2006; Volume 2, pp. 256–285. [Google Scholar]
- Shepherdson, D.J.; Mellen, J.D.; Hutchins, M. Second Nature: Environmental Enrichment for Captive Animals; Shepherdson, D.J., Mellen, J.D., Hutchins, M., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1998. [Google Scholar]
- Reading, R.P.; Miller, B.; Shepherdson, D. The Value of Enrichment to Reintroduction Success. Zoo Biol. 2013, 32, 332–341. [Google Scholar] [CrossRef]
- Wagman, J.D.; Lukas, K.E.; Dennis, P.M.; Willis, M.A.; Carroscia, J.; Gindlesperger, C.; Schook, M.W. A work-for-food enrichment program increases exploration and decreases stereotypies in four species of bears. Zoo Biol. 2018, 37, 3–15. [Google Scholar] [CrossRef]
- Shepherdson, D. Environmental Enrichment for Elephants: Current Status and Future Directions. J. Elephant Manag. Assoc. 1999, 10, 68–77. [Google Scholar]
- Spinage, C. Elephants; T & AD Poyser Ltd.: London, UK, 1994. [Google Scholar]
- Brando, S.; Buchanan-Smith, H.M. The 24/7 approach to promoting optimal welfare for captive wild animals. Behav. Process. 2018, 156, 83–95. [Google Scholar] [CrossRef]
- Veasey, J.S. Can Zoos Ever Be Big Enough for Large Wild Animals? A Review Using an Expert Panel Assessment of the Psychological Priorities of the Amur Tiger (Panthera tigris altaica) as a Model Species. Animals 2020, 10, 1536. [Google Scholar] [CrossRef]
- Baskaran, N.; Balasubramanian, M.; Swaminathan, S.; Desai, A.A. Feeding ecology of the Asian elephant Elephas maximus Linnaeus in the Nilgiri Biosphere Reserve, southern India. J. Bombay Nat. Hist. Soc. 2010, 107, 3. [Google Scholar]
- Bassett, L.; Buchanan-Smith, H.M. Effects of predictability on the welfare of captive animals. Appl. Anim. Behav. Sci. 2007, 102, 223–245. [Google Scholar] [CrossRef] [Green Version]
- Whitham, J.C.; Wielebnowski, N. New directions for zoo animal welfare science. Appl. Anim. Behav. Sci. 2013, 147, 247–260. [Google Scholar] [CrossRef]
- Holdgate, M.R.; Meehan, C.L.; Hogan, J.N.; Miller, L.J.; Soltis, J.; Andrews, J.; Shepherdson, D.J. Walking Behavior of Zoo Elephants: Associations between GPS-Measured Daily Walking Distances and Environmental Factors, Social Factors, and Welfare Indicators. PLoS ONE 2016, 11, e0150331. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Leighty, K.A.; Soltis, J.; Wesolek, C.M.; Savage, A.; Mellen, J.; Lehnhardt, J. GPS Determination of Walking Rates in Captive African Elephants (Loxodonta africana). Zoo Biol. 2009, 28, 16–28. [Google Scholar] [CrossRef] [PubMed]
- Slotow, R.; Van Dyk, G. Ranging of older male elephants introduced to an existing small population without older males: Pilanesberg National Park. Koedoe 2004, 47, 14. [Google Scholar] [CrossRef] [Green Version]
- Whitehouse, A.M.; Schoeman, D.S. Ranging behaviour of elephants within a small, fenced area in Addo Elephant National Park, South Africa. Afr. Zool. 2003, 38, 95–108. [Google Scholar] [CrossRef]
- Theuerkauf, J.; Ellenberg, H. Movements and defaecation of forest elephants in the moist semi-deciduous Bossematié Forest Reserve, Ivory Coast. Afr. J. Ecol. 2000, 38, 258–261. [Google Scholar] [CrossRef]
- Sach, F.; Dierenfeld, E.S.; Langley-Evans, S.C.; Watts, M.J.; Yon, L. African savanna elephants (Loxodonta africana) as an example of a herbivore making movement choices based on nutritional needs. PeerJ 2019, 7, e6260. [Google Scholar] [CrossRef]
- Wittemyer, G.; Getz, W.M.; Vollrath, F.; Douglas-Hamilton, I. Social dominance, seasonal movements, and spatial segregation in African elephants: A contribution to conservation behavior. Behav. Ecol. Sociobiol. 2007, 61, 1919–1931. [Google Scholar] [CrossRef]
- Boult, V.L.; Sibly, R.M.; Quaife, T.; Fishlock, V.; Moss, C.; Lee, P.C. Modelling large herbivore movement decisions: Beyond food availability as a predictor of ranging patterns. Afr. J. Ecol. 2019, 57, 10–19. [Google Scholar] [CrossRef] [Green Version]
- Mench, J.A. Environmental enrichment and the importance of exploratory behavior. In Second Nature: Environmental Enrichment for Captive Animals; Shepherdson, D.J., Mellen, J.D., Hutchins, M., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1998; pp. 30–46. [Google Scholar]
- Boissy, A.; Manteuffel, G.; Jensen, M.B.; Moe, R.O.; Spruijt, B.; Keeling, L.J.; Winckler, C.; Forkman, B.; Dimitrov, I.; Langbein, J.; et al. Assessment of positive emotions in animals to improve their welfare. Physiol. Behav. 2007, 92, 375–397. [Google Scholar] [CrossRef] [PubMed]
- Shepherdson, D.J.; Carlstead, K.; Mellen, J.D.; Seidensticker, J. The Influence of Food Presentation on the Behavior of Small Cats in Confined Environments. Zoo Biol. 1993, 12, 203–216. [Google Scholar] [CrossRef]
- Lewis, K.D.; Shepherdson, D.J.; Owens, T.M.; Keele, M. A Survey of Elephant Husbandry and Foot Health in North American Zoos. Zoo Biol. 2010, 29, 221–236. [Google Scholar] [CrossRef] [PubMed]
- Wielebnowski, N.C.; Fletchall, N.; Carlstead, K.; Busso, J.M.; Brown, J.L. Noninvasive Assessment of Adrenal Activity Associated with Husbandry and Behavioral Factors in the North American Clouded Leopard Population. Zoo Biol. 2002, 21, 77–98. [Google Scholar] [CrossRef]
- Wielebnowski, N.C.; Ziegler, K.; Wildt, D.E.; Lukas, J.; Brown, J.L. Impact of social management on reproductive, adrenal and behavioural activity in the cheetah (Acinonyx jubatus). Anim. Conserv. 2002, 5, 291–301. [Google Scholar] [CrossRef] [Green Version]
- Wielebnowski, N. Stress and distress: Evaluating their impact for the well-being of zoo animals. J. Am. Vet. Med. Assoc. 2003, 223, 973–977. [Google Scholar] [CrossRef]
- Möstl, E.; Palme, R. Hormones as indicators of stress. Domest. Anim. Endocrinol. 2002, 23, 67–74. [Google Scholar] [CrossRef]
- Buwalda, B.; Scholte, J.; de Boer, S.F.; Coppens, C.M.; Koolhaas, J.M. The acute glucocorticoid stress response does not differentiate between rewarding and aversive social stimuli in rats. Horm. Behav. 2012, 61, 218–226. [Google Scholar] [CrossRef]
- Ralph, C.R.; Tilbrook, A.J. Invited Review: The usefulness of measuring glucocorticoids for assessing animal welfare. J. Anim. Sci. 2016, 94, 457–470. [Google Scholar] [CrossRef] [Green Version]
- Palme, R. Non-invasive measurement of glucocorticoids: Advances and problems. Physiol. Behav. 2019, 199, 229–243. [Google Scholar] [CrossRef]
- St. Juliana, J.R.; Kotler, B.P.; Wielebnowski, N.; Cox, J.G. Stress as an adaptation I: Stress hormones are correlated with optimal foraging behaviour of gerbils under the risk of predation. Evol. Ecol. Res. 2017, 18, 571–585. [Google Scholar]
- McPhee, M.E.; Carlstead, K. The importance of maintaining natural behaviors in captive mammals. In Wild Mammals in Captivity: Principles and Techniques for Zoo Management, 2nd ed.; Kleiman, D.G., Thompson, K.V., Baer, C.K., Eds.; University of Chicago Press: Chicago, IL, USA, 2010; pp. 303–313. [Google Scholar]
- Watters, J.V.; Margulis, S.W.; Atsalis, S. Behavioral Monitoring in Zoos and Aquariums: A Tool for Guiding Husbandry and Directing Research. Zoo Biol. 2009, 28, 35–48. [Google Scholar] [CrossRef] [PubMed]
- Kersey, D.C.; Dehnhard, M. The use of noninvasive and minimally invasive methods in endocrinology for threatened mammalian species conservation. Gen. Comp. Endocrinol. 2014, 203, 296–306. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Meller, C.L.; Croney, C.C.; Shepherdson, D. Effects of rubberized flooring on Asian elephant behavior in captivity. Zoo Biol. 2007, 26, 51–61. [Google Scholar] [CrossRef]
- R Core Team. R: A Language and Environment for Statistical Computing; R Foundation for Statistical Computing: Vienna, Austria, 2021; Available online: https://www.r-project.org/ (accessed on 21 March 2020).
- Kurt, F. Remarks on the social structure and ecology of the Ceylon elephant in the Yala National Park. In The Behavior of Ungulates and Its Relation to Management; Geist, V., Walther, F., Eds.; International Union for Conservation of Nature and Natural Resources: Morges, Switzerland, 1974; Volume 2, pp. 618–634. [Google Scholar]
- Holdgate, M.R. Applying GPS and Accelerometers to the Study of African Savanna (Loxodonta africana) and Asian Elephant (Elephas maximus) Welfare in Zoos; Portland State University: Portland, OR, USA, 2015. [Google Scholar]
- Hutchinson, J.R.; Schwerda, D.; Famini, D.J.; Dale, R.H.; Fischer, M.S.; Kram, R. The locomotor kinematics of Asian and African elephants: Changes with speed and size. J. Exp. Biol. 2006, 209, 3812–3827. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Brown, J.L.; Kersey, D.C.; Freeman, E.W.; Wagener, T. Assessment of Diurnal Urinary Cortisol Excretion in Asian and African Elephants Using Different Endocrine Methods. Zoo Biol. 2010, 29, 274–283. [Google Scholar] [CrossRef]
- Brown, J.L.; Lehnhardt, J. Serum and urinary hormones during pregnancy and the peri- and postpartum period in an Asian elephant (Elephas, maximus). Zoo Biol. 1995, 14, 555–564. [Google Scholar] [CrossRef]
- Brown, J.L.; Walker, S.L.; Moeller, T. Comparative endocrinology of cycling and non-cycling Asian (Elephas maximus) and African (Loxodonta africana) elephants. Gen. Comp. Endocrinol. 2004, 136, 360–370. [Google Scholar] [CrossRef]
- Brown, J.L. Comparative Reproductive Biology of Elephants. In Reproductive Sciences in Animal Conservation: Progress and Prospects; Holt, W.V., Brown, J.L., Comizzoli, P., Eds.; Advances in Experimental Medicine and Biology; Springer Science and Business Media: New York, NY, USA, 2014; pp. 135–169. [Google Scholar]
- Brown, J.L.; Bush, M.; Wildt, D.E.; Raath, J.R.; de Vos, V.; Howard, J.G. Effects of GnRH analogues on pituitary-testicular function in free-ranging African elephants (Loxodonta africana). J. Reprod. Fertil. 1993, 99, 627–634. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.L.; Somerville, M.; Riddle, H.S.; Keele, M.; Duer, C.K.; Freeman, E.W. Comparative endocrinology of testicular, adrenal and thyroid function in captive Asian and African elephant bulls. Gen. Comp. Endocrinol. 2007, 151, 153–162. [Google Scholar] [CrossRef] [PubMed]
- Glaeser, S.S. Assessing Well-Being of Asian Elephants (Elephas maximus) through Major Events and Transitions, with Development and Aging, and Husbandry Changes Dissertation; Portland State University: Portland, OR, USA, 2020. [Google Scholar]
- Scarlata, C.D.; Elias, B.A.; Godwin, J.R.; Powell, R.A.; Shepherdson, D.; Shipley, L.A.; Brown, J.L. Characterizing gonadal and adrenal activity by fecal steroid analyses in pygmy rabbits (Brachylagus idahoensis). Gen. Comp. Endocrinol. 2011, 171, 373–380. [Google Scholar] [CrossRef]
- Brown, J.L.; Carlstead, K.; Bray, J.D.; Dickey, D.; Farin, C.; Ange-van Heugten, K. Individual and environmental risk factors associated with fecal glucocorticoid metabolite concentrations in zoo-housed Asian and African elephants. PLoS ONE 2019, 14, e0217326. [Google Scholar] [CrossRef] [Green Version]
- Watson, R.; Munro, C.; Edwards, K.L.; Norton, V.; Brown, J.L.; Walker, S.L. Development of a versatile enzyme immunoassay for non-invasive assessment of glucocorticoid metabolites in a diversity of taxonomic species. Gen. Comp. Endocrinol. 2013, 186, 16–24. [Google Scholar] [CrossRef] [PubMed]
- Glaeser, S.S.; Hunt, K.E.; Martin, M.S.; Finnegan, M.; Brown, J.L. Investigation of individual and group variability in estrous cycle characteristics in female Asian elephants (Elephas maximus) at the Oregon Zoo. Theriogenology 2012, 78, 285–296. [Google Scholar] [CrossRef] [PubMed]
- Brown, J.L.; Schmitt, D.L.; Bellem, A.; Graham, L.H.; Lehnhardt, J. Hormone Secretion in the Asian Elephant (Elephas maximus): Characterization of Ovulatory and Anovulatory Luteinizing Hormone Surges. Biol. Reprod. 1999, 61, 1294–1299. [Google Scholar] [CrossRef] [Green Version]
- Brown, J.L.; Lehnhardt, J. Secretory Patterns of Serum Prolactin in Asian (Elephas maximus) and African (Loxodonta africana) Elephants During Different Reproductive States: Comparison With Concentrations in a Noncycling African elephant. Zoo Biol. 1997, 16, 149–159. [Google Scholar] [CrossRef] [Green Version]
- Brown, M.B.; Forsythe, A.B. Robust Tests for the Equality of Variances. J. Am. Stat. Assoc. 1974, 69, 364–367. [Google Scholar] [CrossRef]
- Hines, W.G.; Hines, R.J. Increased Power with Modified Forms of the Levene (Med) Test for Heterogeneity of Variance. Biometrics 2000, 56, 451–454. [Google Scholar] [CrossRef]
- Benjamini, Y.; Hochberg, Y. Controlling the False Discovery Rate: A Practical and Powerful Approach to Multiple Testing. J. R. Stat. Soc. Ser. B 1995, 57, 289–300. [Google Scholar] [CrossRef]
- Parra-Frutos, I. Testing homogeneity of variances with unequal sample sizes. Comput. Stat. 2012, 28, 1269–1297. [Google Scholar] [CrossRef]
- Olson, D. (Ed.) Elephant Husbandry Resource Guide; International Elephant Foundation: Azle, TX, USA, 2004. [Google Scholar]
- Eisenberg, J.F.; McKay, C.M.; Seidensticker, J. Asian Elephants; Friends of the National Zoo and National Zoological Park Smithsonian Institution: Washington, DC, USA, 1990; p. 271. [Google Scholar]
- Kahl, M.P.; Santiapillai, C. A glossary of elephant terms. Gajah 2004, 23, 1–36. [Google Scholar]
- Rasmussen, L.E.L. Chemical communication: An integral part of functional Asian elephant (Elephas maximus) society. Ecoscience 1998, 5, 410–426. [Google Scholar] [CrossRef]
- Slade, B.E.; Schulte, B.A.; Rasmussen, L.E.L. Oestrous state dynamics in chemical communication by captive female Asian elephants. Anim. Behav. 2003, 65, 813–819. [Google Scholar] [CrossRef] [Green Version]
- Shepherdson, D. The role of environmental enrichment in the captive breeding and reintroduction of endangered species. In Creative Conservation; Olney, P.J.S., Mace, G.M., Feistner, A.T.C., Eds.; Springer: Dordrecht, The Netherlands, 1994; pp. 167–177. [Google Scholar]
- Martin, P.; Bateson, P. Measuring Behaviour: An Introductory Guide; Cambridge University Press: Cambridge, UK, 1993. [Google Scholar]
- Margulis, S.W.; Westhus, E.J. Evaluation of different observational sampling regimes for use in zoological parks. Appl. Anim. Behav. Sci. 2008, 110, 363–376. [Google Scholar] [CrossRef]
- Fowler, J.; Cohen, L.; Jarvis, P. Practical Statistics for Field Biology, 2nd ed.; John Wiley & Sons: New York, NY, USA, 1999. [Google Scholar]
- Plowman, A.B. BIAZA statistics guidelines: Toward a common application of statistical tests for zoo research. Zoo Biol. 2008, 27, 226–233. [Google Scholar] [CrossRef]
- Signorell, A.; Aho, K.; Alfons, A.; Anderegg, N.; Aragon, T.; Arppe, A.; Baddeley, A.; Barton, K.; Bolker, B.; Borchers, H. DescTools: Tools for Descriptive Statistics; R Package Version 0.99.28; R Foundation for Statistical Computing: Vienna, Austria, 2019; Available online: https://www.r-project.org/ (accessed on 21 March 2020).
- Rowell, Z.E. Locomotion in captive Asian elephants (Elephas maximus). J. Zoo Aquar. Res. 2014, 2, 130–135. [Google Scholar] [CrossRef]
- Brady, A.; McMahon, B.J.; Naulty, F. Estimates of locomotion in Asian elephants Elephas maximus using video monitoring at Dublin Zoo, Ireland. J. Zoo Aquar. Res. 2021, 9, 124–133. [Google Scholar] [CrossRef]
- Merz, G. Movement patterns and group size of the African forest elephant Loxodonta africana cyclotis in the Tai National Park, Ivory Coast. Afr. J. Ecol. 1986, 24, 133–136. [Google Scholar] [CrossRef]
- Wall, J.; Wittemyer, G.; Klinkenberg, B.; LeMay, V.; Douglas-Hamilton, I. Characterizing properties and drivers of long distance movements by elephants (Loxodonta africana) in the Gourma, Mali. Biol. Conserv. 2013, 157, 60–68. [Google Scholar] [CrossRef]
- Viljoen, P.J. Spatial distribution and movements of elephants (Loxodonta africana) in the northern Namib Desert region of the Kaokoveld, South West Africa/Namibia. J. Zool. 1989, 219, 1–19. [Google Scholar] [CrossRef]
- Hertel, A.G.; Niemela, P.T.; Dingemanse, N.J.; Mueller, T. A guide for studying among-individual behavioral variation from movement data in the wild. Mov. Ecol. 2020, 8, 30. [Google Scholar] [CrossRef] [PubMed]
- Chan, A.N.; Wittemyer, G.; McEvoy, J.; Williams, A.C.; Cox, N.; Soe, P.; Grindley, M.; Shwe, N.M.; Chit, A.M.; Oo, Z.M.; et al. Landscape Characteristics Influence Ranging Behavior of Asian Elephants at the Human-wildlands Interface in Myanmar. Mov. Ecol. Prepr. 2021, 27. [Google Scholar] [CrossRef]
- Alfred, R.; Ahmad, A.H.; Payne, J.; Williams, C.; Ambu, L.N.; How, P.M.; Goossens, B. Home range and ranging behaviour of Bornean elephant (Elephas maximus borneensis) females. PLoS ONE 2012, 7, e31400. [Google Scholar] [CrossRef] [Green Version]
- Veasey, J.S. Assessing the Psychological Priorities for Optimising Captive Asian Elephant (Elephas maximus) Welfare. Animals 2019, 10, 39. [Google Scholar] [CrossRef] [Green Version]
- Glaeser, S.S.; Edwards, K.L.; Wielebnowski, N.; Brown, J.L. Effects of physiological changes and social life events on adrenal glucocorticoid activity in female zoo-housed Asian elephants (Elephas maximus). PLoS ONE 2020, 15, e0241910. [Google Scholar] [CrossRef]
- Keerthipriya, P.; Nandini, S.; Gautam, H.; Revathe, T.; Vidya, T.N.C.; Monteith, K. Musth and its effects on male–male and male–female associations in Asian elephants. J. Mammal. 2020, 101, 259–270. [Google Scholar] [CrossRef]
- Chelliah, K.; Sukumar, R. Interplay of male traits, male mating strategies and female mate choice in the Asian elephant, Elephas maximus. Behaviour 2015, 152, 1113–1144. [Google Scholar] [CrossRef]
- Hildebrandt, T.B.; Goritz, F.; Hermes, R.; Reid, C.; Dehnhard, M.; Brown, J.L. Aspects of the reproductive biology and breeding management of Asian and African elephants (Elephas maximus and Loxodonta africana). Int. Zoo Yearb. 2006, 40, 20–40. [Google Scholar] [CrossRef]
- Scott, N.L.; Riddle, H. Assessment of Musth in Captivity: A Survey of Factors Affecting the Frequency and Duration of Musth in Captive Male Elephants Elephas maximus-Loxodonta africana. J. Elephant Manag. Assoc. 2003, 14, 11–15. [Google Scholar]
- Cooper, K.A.; Harder, J.D.; Clawson, D.H.; Fredrick, D.L.; Lodge, G.A.; Peachey, H.C.; Spellmire, T.J.; Winstel, D.P. Serum testosterone and musth in captive male African and Asian elephants. Zoo Biol. 1990, 9, 297–306. [Google Scholar] [CrossRef]
- DeVries, A.C.; Glasper, E.R.; Detillion, C.E. Social modulation of stress responses. Physiol. Behav. 2003, 79, 399–407. [Google Scholar] [CrossRef]
- Plotnik, J.M.; de Waal, F.B.M. Asian elephants (Elephas maximus) reassure others in distress. PeerJ 2014, 2, e278. [Google Scholar] [CrossRef] [Green Version]
- Carlstead, K.; Paris, S.; Brown, J.L. Good keeper-elephant relationships in North American zoos are mutually beneficial to welfare. Appl. Anim. Behav. Sci. 2019, 211, 103–111. [Google Scholar] [CrossRef]
- Cole, J.; Fraser, D. Zoo Animal Welfare: The Human Dimension. J. Appl. Anim. Welf. Sci. 2018, 21, 49–58. [Google Scholar] [CrossRef] [Green Version]
- Mellor, D.J.; Beausoleil, N.J.; Littlewood, K.E.; McLean, A.N.; McGreevy, P.D.; Jones, B.; Wilkins, C. The 2020 Five Domains Model: Including Human-Animal Interactions in Assessments of Animal Welfare. Animals 2020, 10, 1870. [Google Scholar] [CrossRef]
- Rault, J.-L. Friends with benefits: Social support and its relevance for farm animal welfare. Appl. Anim. Behav. Sci. 2012, 136, 1–14. [Google Scholar] [CrossRef]
- Wingfield, J.C. The Concept of Allostasis: Coping With a Capricious Environment. J. Mammal. 2005, 86, 248–254. [Google Scholar] [CrossRef]
- Jakob-Hoff, R.; Kingan, M.; Fenemore, C.; Schmid, G.; Cockrem, J.F.; Crackle, A.; Bemmel, E.V.; Connor, R.; Descovich, K. Potential Impact of Construction Noise on Selected Zoo Animals. Animals 2019, 9, 504. [Google Scholar] [CrossRef] [Green Version]
- Carlstead, K.; Brown, J.L. Relationships Between Patterns of Fecal Corticoid Excretion and Behavior, Reproduction, and Environmental Factors in Captive Black (Diceros bicornis) and White (Ceratotherium simum) Rhinoceros. Zoo Biol. 2005, 24, 215–232. [Google Scholar] [CrossRef]
- Cockrem, J.F. Individual variation in glucocorticoid stress responses in animals. Gen. Comp. Endocrinol. 2013, 181, 45–58. [Google Scholar] [CrossRef]
- Koolhaas, J.M.; Korte, S.M.; De Boer, S.F.; Van Der Vegt, B.J.; Van Reenen, C.G.; Hopster, H.; De Jong, I.C.; Ruis, M.A.W.; Blokhuis, H.J. Coping styles in animals: Current status in behavior and stress-physiology. Neurosci. Biobehav. Rev. 1999, 23, 925–935. [Google Scholar] [CrossRef]
- Pesantez-Pacheco, J.L.; Heras-Molina, A.; Torres-Rovira, L.; Sanz-Fernandez, M.V.; Garcia-Contreras, C.; Vazquez-Gomez, M.; Feyjoo, P.; Caceres, E.; Frias-Mateo, M.; Hernandez, F.; et al. Maternal Metabolic Demands Caused by Pregnancy and Lactation: Association with Productivity and Offspring Phenotype in High-Yielding Dairy Ewes. Animals 2019, 9, 295. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Pokharel, S.S.; Seshagiri, P.B.; Sukumar, R. Influence of the number of calves and lactating adult females in a herd on the adrenocortical activity of free-ranging Asian elephants. Wildl. Res. 2019, 46, 679–689. [Google Scholar] [CrossRef]
- Vijayakrishnan, S.; Kumar, M.A.; Umapathy, G.; Kumar, V.; Sinha, A. Physiological stress responses in wild Asian elephants Elephas maximus in a human-dominated landscape in the Western Ghats, southern India. Gen. Comp. Endocrinol. 2018, 266, 150–156. [Google Scholar] [CrossRef]
- Sapolsky, R.M.; Meaney, M.J. Maturation of the Adrenocortical Stress Response: Neuroendocrine Control Mechanisms and the Stress Hyporesponsive Period. Brain Res. Rev. 1986, 396, 64–76. [Google Scholar] [CrossRef]
- Veasey, J.; Waran, N.; Young, R. On comparing the behaviour of zoo housed animals with wild conspecifics as a welfare indicator. Anim. Welf. 1996, 5, 13–24. [Google Scholar]
- Yeates, J. Naturalness and Animal Welfare. Animals 2018, 8, 53. [Google Scholar] [CrossRef] [Green Version]
- Veasey, J. Concepts in the care and welfare of captive elephants. Int. Zoo Yearb. 2006, 40, 63–79. [Google Scholar] [CrossRef]
- Fazio, J.M.; Barthel, T.; Freeman, E.W.; Garlick-Ott, K.; Scholle, A.; Brown, J.L. Utilizing Camera Traps, Closed Circuit Cameras and Behavior Observation Software to Monitor Activity Budgets, Habitat Use, and Social Interactions of Zoo-Housed Asian Elephants (Elephas maximus). Animals 2020, 10, 2026. [Google Scholar] [CrossRef] [PubMed]
- Rees, P.A. Activity Budgets and the Relationship Between Feeding and Stereotypic Behaviors in Asian elephants (Elephas maximus) in a Zoo. Zoo Biol. 2009, 28, 79–97. [Google Scholar] [CrossRef]
- De Jonge, F.H.; Tilly, S.-L.; Baars, A.M.; Spruijt, B.M. On the rewarding nature of appetitive feeding behaviour in pigs (Sus scrofa): Do domesticated pigs contrafreeload? Appl. Anim. Behav. Sci. 2008, 114, 359–372. [Google Scholar] [CrossRef] [Green Version]
- Montaudouin, S.; Pape, G.L. Comparison between 28 zoological parks: Stereotypic and social behaviours of captive brown bears (Ursus arctos). Appl. Anim. Behav. Sci. 2005, 92, 129–141. [Google Scholar] [CrossRef]
- Cooper, J.J.; McDonald, L.; Mills, D.S. The effect of increasing visual horizons on stereotypic weaving: Implications for the social housing of stabled horses. Appl. Anim. Behav. Sci. 2000, 69, 67–83. [Google Scholar] [CrossRef]
- Carlstead, K. Determining the causes of stereotypic behaviors in zoo carnivores: Toward appropriate enrichment strategies. In Second Nature: Environmental Enrichment for Captive Animals; Shepherdson, D.J., Mellen, J.D., Hutchins, M., Eds.; Smithsonian Institution Press: Washington, DC, USA, 1998; pp. 172–183. [Google Scholar]
- Mason, G.J. Stereotypies: A critical review. Anim. Behav. 1991, 41, 1015–1037. [Google Scholar] [CrossRef] [Green Version]
- Mason, G.; Veasey, J. Chapter 11: How should the psychological welfare of zoo elephants be investigated. In An Elephant in the Room: The Science and Well Being of Elephants in Captivity; Forthman, D., Kane, L., Waldau, P., Eds.; Tufts University Cummings School of Veterinary Medicine’s Center for Animals and Public Policy: North Grafton, MA, USA, 2008. [Google Scholar]
- Mason, G.J.; Latham, N. Can’t stop, won’t stop: Is stereotypy a reliable animal welfare indicator? Anim. Welf. 2004, 13, 57–69. [Google Scholar]
- Bashaw, M.J.; Bloomsmith, M.A.; Marr, M.J.; Maple, T.L. To Hunt or Not to Hunt? A Feeding Enrichment Experiment with Captive Large Felids. Zoo Biol. 2003, 22, 189–198. [Google Scholar] [CrossRef]
- Forthman, D.L.; Elder, S.D.; Bakeman, R.; Kurkowski, T.W.; Noble, C.C.; Winslow, S.W. Effects of Feeding Enrichment on Behavior of Three Species of Captive Bears. Zoo Biol. 1992, 11, 187–195. [Google Scholar] [CrossRef]
- Carlstead, K.; Seidensticker, J.; Baldwin, R. Environmental Enrichment for Zoo Bears. Zoo Biol. 1991, 10, 3–16. [Google Scholar] [CrossRef]
- Swaisgood, R.R.; Shepherdson, D.J. Scientific Approaches to Enrichment and Stereotypies in Zoo Animals: What’s Been done and Where Should We Go Next? Zoo Biol. 2005, 24, 499–518. [Google Scholar] [CrossRef]
- White, B.C.; Houser, L.A.; Fuller, J.A.; Taylor, S.; Elliott, J.L.L. Activity-Based Exhibition of Five Mammalian Species: Evaluation of behavioral Changes. Zoo Biol. 2003, 22, 269–285. [Google Scholar] [CrossRef]
- Powell, D.M.; Vitale, C. Behavioral Changes in Female Asian Elephants When Given Access to an Outdoor Yard Overnight. Zoo Biol. 2016, 35, 298–303. [Google Scholar] [CrossRef]
- Gunawardene, M.; Jayasinghe, L.; Janaka, H.; Weerakoon, D.; Wickramanayake, E.; Fernando, P. Social Organisation of Elephants in Southern Sri Lanka. In Proceedings of the Symposium for Human-Elephant Relationships and Conflicts, Colombo, Sri Lanka, 19–21 September 2003. [Google Scholar]
- Jayantha, D.; Dayawansa, P.; Padmalal, U.; Ratnasooriya, W.D. Social relationships of wild juvenile Asian Elephants Elephas maximus in the Udawalawa National Park, Sri Lanka. J. Threat. Taxa 2009, 1, 211–214. [Google Scholar] [CrossRef] [Green Version]
- Sukumar, R. The Asian Elephant: Ecology and Management; Cambridge University Press: Cambridge, UK, 1989; p. 255. [Google Scholar]
- Lee, P.C.; Moss, C. The social context for learning and behavioural development among wild African elephants. In Mammallian Social Learning: Comparative and Ecological Perspectives, Zoological Society of London, Symposium 73; Box, H.O., Gibson, K.R., Eds.; Cambridge University Press: Cambridge, UK, 1999; pp. 102–125. [Google Scholar]
- Slade-Cain, B.E.; Rasmussen, L.E.; Schulte, B.A. Estrous State Influences on Investigative, Aggressive, and Tail Flicking Behavior in Captive Female Asian Elephants. Zoo Biol. 2008, 27, 167–180. [Google Scholar] [CrossRef] [PubMed]
- Schulte, B.A. Social Structure and Helping Behavior in Captive Elephants. Zoo Biol. 2000, 19, 447–459. [Google Scholar] [CrossRef]
- Freeman, E.W.; Weiss, E.; Brown, J.L. Examination of the Interrelationships of Behavior, Dominance Status, and Ovarian Activity in Captive Asian and African Elephants. Zoo Biol. 2004, 23, 431–448. [Google Scholar] [CrossRef]
- Archie, E.A.; Morrison, T.A.; Foley, C.A.H.; Moss, C.J.; Alberts, S.C. Dominance rank relationships among wild female African elephants, Loxodonta africana. Anim. Behav. 2006, 71, 117–127. [Google Scholar] [CrossRef]
- De Silva, S.; Schmid, V.; Wittemyer, G. Fission–fusion processes weaken dominance networks of female Asian elephants in a productive habitat. Behav. Ecol. 2017, 8, 243–252. [Google Scholar] [CrossRef] [Green Version]
- Pinto, A.; Stelvig, M.; Costa, C.; Colaço, J.; Colaço, B. Influence of male presence on female Asian elephants (Elephas maximus) behaviour in captivity. J. Zoo Aquar. Res. 2020, 8, 45–49. [Google Scholar]
- Bonaparte-Saller, M.; Mench, J.A. Assessing the dyadic social relationships of female african (Loxodonta africana) and asian (Elephas maximus) zoo elephants using proximity, tactile contact, and keeper surveys. Appl. Anim. Behav. Sci. 2018, 199, 45–51. [Google Scholar] [CrossRef]
- Leighty, K.A.; Soltis, J.; Savage, A. GPS Assessment of the Use of Exhibit Space and Resources by African Elephants (Loxodonta africana). Zoo Biol. 2010, 29, 210–220. [Google Scholar] [CrossRef] [PubMed]
- Robitaille, J.-F.; Prescott, J. Use of space and activity budgets in relation to age and social status in a captive herd of American bison, Bison bison. Zoo Biol. 1993, 12, 367–379. [Google Scholar] [CrossRef]
- Badihi, I. The Effects of Complexity, Choice and Control on the Behaviour and the Welfare of Captive Common Marmosets (Callithrix jacchus); University of Stirling: Stirling, UK, 2006. [Google Scholar]
- Kagan, R.; Veasey, J. Challenges of zoo animal welfare. In Wild Mammals in Captivity: Principles and Techniques for Zoo Management, 2nd ed.; Kleiman, D.G., Thompson, K.V., Baer, C.K., Eds.; University of Chicago Press: Chicago, IL, USA, 2010; pp. 11–21. [Google Scholar]
- Taylor, P.E.; Coerse, N.C.; Haskell, M. The effects of operant control over food and light on the behaviour of domestic hens. Appl. Anim. Behav. Sci. 2001, 71, 319–333. [Google Scholar] [CrossRef]
- Joffe, J.M.; Rawson, R.A.; Mulick, J.A. Control of Their Environment Reduces Emotionality in Rats. Science 1973, 180, 1383–1384. [Google Scholar] [CrossRef]
- Carlstead, K.; Shepherdson, D. Effects of Environmental Enrichment on Reproduction. Zoo Biol. 1994, 13, 447–458. [Google Scholar] [CrossRef]
- Ross, S.R.; Calcutt, S.; Schapiro, S.J.; Hau, J. Space Use Selectivity by Chimpanzees and Gorillas in an Indoor-Outdoor Enclosure. Am. J. Primatol. 2011, 73, 197–208. [Google Scholar] [CrossRef]
- Herrelko, E.S.; Buchanan-Smith, H.M.; Vick, S.J. Perception of Available Space During Chimpanzee Introductions: Number of Accessible Areas is More Important Than Enclosure Size. Zoo Biol. 2015, 34, 397–405. [Google Scholar] [CrossRef]
- Markowitz, H. Behavioral Enrichment in the Zoo; Van Nostrand Reinhold: New York, NY, USA, 1982. [Google Scholar]
- Rauvola, R.S. Aging, Control, & Well-Being: A Longitudinal Investigation of Actual, Perceived, and Enacted Control across Contexts. Ph.D. Thesis, Saint Louis University, St. Louis, MO, USA, 2020. [Google Scholar]
- Wallhagen, M.I.; Kagan, S.H. Staying within bounds: Perceived control and the experience of elderly caregivers. J. Aging Stud. 1993, 7, 197–213. [Google Scholar] [CrossRef]
- Robinson, S.A.; Lachman, M.E. Perceived Control and Aging: A Mini-Review and Directions for Future Research. Gerontology 2017, 63, 435–442. [Google Scholar] [CrossRef] [PubMed]
- Leotti, L.A.; Iyengar, S.S.; Ochsner, K.N. Born to choose: The origins and value of the need for control. Trends Cogn. Sci. 2010, 14, 457–463. [Google Scholar] [CrossRef] [Green Version]
- Carlstead, K. Effects of captivity on the behavior of wild mammals. In Wild Mammals in Captivity: Principles and Techniques, 1st ed.; Kleiman, D.G., Allen, M.E., Harris, H., Thompson, K.V., Lumpkin, S., Eds.; University of Chicago Press: Chicago, IL, USA, 1996; pp. 317–333. [Google Scholar]
- Baker, K.; Melfi, V.A. The Ultimate Benefits of Learning. In Zoo Animal Learning and Training; Melfi, V.A., Dorey, N.R., Ward, S.J., Eds.; John Wiley & Sons Ltd.: New York, NY, USA, 2020; pp. 35–51. [Google Scholar]
- Scott, N.L.; LaDue, C.A. The behavioral effects of exhibit size versus complexity in African elephants: A potential solution for smaller spaces. Zoo Biol. 2019, 38, 448–457. [Google Scholar] [CrossRef] [PubMed]
- Mellor, D.J. Updating Animal Welfare Thinking: Moving beyond the “Five Freedoms” towards “A Life Worth Living”. Animals 2016, 6, 21. [Google Scholar] [CrossRef] [PubMed]











| Elephant | Sex ♂/♀ | Origin | Date of Birth or Transfer to OZ | Age at Start of Sample Collection |
|---|---|---|---|---|
| Female 1 | ♀ | Zoo-born Oregon Zoo | 26 December 1982—Birth | 29 years |
| Female 2 1 | ♀ | Wild Borneo, Malaysia | ~1993—Birth 20 November 1999—Transfer to Oregon Zoo | 19 years |
| Female 3 | ♀ | Zoo-born Oregon Zoo | 31 August 1994—Birth | 18 years |
| M-juvenile 2 | ♂ | Zoo-born Oregon Zoo | 23 August 2008—Birth | 4 years |
| F-calf 2 | ♀ | Zoo-born Oregon Zoo | 30 November 2012—Birth | 2 Months |
| Proximity | Definition |
| Proximate | Focal animal is within 2 body lengths of or in contact with another individual (defined as 2 adult female body lengths, c.a. 10 m.) |
| Proximity Not Visible | Focal animal or other elephants are not visible enough to determine proximity. |
| Behavior Category | Definition |
| Behavior Not Visible | Elephant or activity is not visible enough to determine the behavior. |
| Social Interaction | Interacting with another elephant in a social context, either with physical contact (e.g., trunk twine, play) or without physical contact but within 2 body lengths (e.g., sharing food, displace). |
| Food-Delivery Object Interaction | Interacting with any object that distributes food, either permanent or provided by caretakers. |
| Feeding/Drinking | Acquiring/gathering and consuming (putting in the mouth) any food item without interacting with a food-delivery object. Drinking water, not bathing. |
| Enrichment (non-food) Object Interaction | Interacting objects that do not provision food and are not permanent (e.g., firehose ball or braid). |
| Habitat Feature Interaction | Interacting with features that are permanent in the habitat (e.g., dusting, bathing, digging, rubbing body, climbing on logs, investigating features with feet or mouth). |
| Repetitive Behaviors (Stereotypy) | Motor: Repeatedly performing a behavior for 3 or more consecutive repetitions without interruption (e.g., route tracing). Non motor stereotypy: Performing the behavior for 5 s or more. |
| Locomotion | Walking or running (fast walking) more than 2 body lengths in any direction, without stopping for 3 s or more. |
| Stationary | Any stationary state without engaging in another behavior for 3 s or longer. Standing, walking or shuffling (without moving 2 body lengths), sitting, kneeling, lying (prone or sternal). |
| Elephant | Previous Habitat | Construction Phase | New Habitat | |||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|---|
| N | Median (Range) | Q1–Q3 | IQR | N | Median (Range) | Q1–Q3 | IQR | N | Median (Range) | Q1–Q3 | IQR | |
| Female 1 | 19 | 124.21 (99.51–237.21) | 112.71–138.57 | 25.86 | 85 | 162.81 (32.08–829.08) | 120.34–260.41 | 140.07 | 49 | 116.57 (36.80–628.53) | 98.22–158.55 | 60.03 |
| Female 2 | 16 | 64.02 (43.37–81.65) | 57.89–71.49 | 13.61 | 104 | 91.68 (21.60–356.66) | 69.44–129.16 | 59.72 | 52 | 96.41 (29.59–162.84) | 76.78–122.82 | 46.03 |
| Female 3 | 12 | 141.91 (72.89–198.49) | 124.80–168.32 | 43.52 | 97 | 193.59 (4.62–1355.22) | 154.98–252.37 | 97.40 | 49 | 146.10 (14.20–231.54) | 91.48–175.85 | 84.37 |
| M-juvenile | 18 | 65.57 (40.85–124.26) | 57.44–71.20 | 13.76 | 115 | 103.50 (40.02–770.50) | 81.55–147.03 | 65.48 | 53 | 79.79 (14.1–120.30) | 63.92–91.13 | 27.21 |
| F-calf | ND | ND | ND | ND | 117 | 136.88 (7.12–730.38) | 101.03–195.88 | 94.85 | 51 | 100.19 (26.05–262.32) | 74.93–136.11 | 61.18 |
| Elephant | Kruskal-Wallis Test | Dunn’s Post-Hoc Pairwise Comparisons | |||
|---|---|---|---|---|---|
| Previous Habitat | Construction Phase | New Habitat | Dunn’s Multiple Comparisons | ||
| Female 1 | χ2 = 16.04, df = 2, p < 0.001 | X | X | Z = 1.90, p = 0.085 | |
| X-higher | X | Z = 3.90, p < 0.001 | |||
| X | X | Z = −0.80, p = 0.425 | |||
| Female 2 | χ2 = 16.10, df = 2, p < 0.001 | X | X-higher | Z = 3.93, p < 0.001 | |
| X | X | Z = 0.06, p = 0.949 | |||
| X | X-higher | Z = 3.66, p < 0.001 | |||
| Female 3 | χ2 = 30.61, df = 2, p < 0.001 | X | X-higher | Z = 2.81, p = 0.007 | |
| X-higher | X | Z = 5.22, p < 0.001 | |||
| X | X | Z = −0.17, p = 0.87 | |||
| M-juvenile | χ2 = 51.64, df =2, p < 0.001 | X | X-higher | Z = 5.61, p < 0.001 | |
| X-higher | X | Z = 5.55, p < 0.001 | |||
| X | X | Z = 1.83, p = 0.067 | |||
| F-calf | χ2 = 51.81, df = 1, p < 0.001 | ND | X-higher | X | |
| Elephant | Levene’s Test for Equality of Variance | ||||
|---|---|---|---|---|---|
| Across All Phases | Previous Habitat | Construction Phase | New Habitat | Dunn’s Multiple Comparisons | |
| Female 1 | F = 6.19, df = 2, p = 0.003 | X | X-higher | F = 5.33, p = 0.023 | |
| X-higher | X | F = 7.82, p = 0.006 | |||
| X | X | F = 0.72, p = 0.400 | |||
| Female 2 | F = 6.82, df = 2, p = 0.001 | X | X-higher | F = 7.61, p = 0.007 | |
| X-higher | X | F = 5.75, p = 0.018 | |||
| X | X-higher | F = 11.61, p = 0.001 | |||
| Female 3 | F = 2.89, df = 2, p = 0.059 | X | X | F = 1.71, p = 0.194 | |
| X-higher | X | F = 4.06, p = 0.046 | |||
| X | X | F = 2.83, p = 0.136 | |||
| M-juvenile | F = 5.26, df = 2, p = 0.006 | X | X | F = 3.24, p = 0.074 | |
| X-higher | X | F = 7.36, p = 0.007 | |||
| X | X | F = 1.51, p = 0.224 | |||
| F-calf | F = 8.09, df = 1, p = 0.005 | ND | X-higher | X | |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Glaeser, S.S.; Shepherdson, D.; Lewis, K.; Prado, N.; Brown, J.L.; Lee, B.; Wielebnowski, N. Supporting Zoo Asian Elephant (Elephas maximus) Welfare and Herd Dynamics with a More Complex and Expanded Habitat. Animals 2021, 11, 2566. https://doi.org/10.3390/ani11092566
Glaeser SS, Shepherdson D, Lewis K, Prado N, Brown JL, Lee B, Wielebnowski N. Supporting Zoo Asian Elephant (Elephas maximus) Welfare and Herd Dynamics with a More Complex and Expanded Habitat. Animals. 2021; 11(9):2566. https://doi.org/10.3390/ani11092566
Chicago/Turabian StyleGlaeser, Sharon S., David Shepherdson, Karen Lewis, Natalia Prado, Janine L. Brown, Bob Lee, and Nadja Wielebnowski. 2021. "Supporting Zoo Asian Elephant (Elephas maximus) Welfare and Herd Dynamics with a More Complex and Expanded Habitat" Animals 11, no. 9: 2566. https://doi.org/10.3390/ani11092566