2. Materials and Methods
2.1. Experimental Design
This study was designed in agreement with the Inspection Form for Guizhou University, Experimental Animal Ethics (EAE-GZU-2020-7009, Guiyang, China). The feeding trial was conducted in a commercial goat farm (106.198244 E, 28.26403 N, Xishui, China). The experiment lasted 74 days, from 10 July to 24 September in 2020. The preparation period was 14 days, and the formal experimental period was 60 days. A total of 18 Qianbei Ma wether kids (body weight, 21.38 ± 1.61 kg; mean ± standard deviation) were randomly allotted into three groups using a completely randomized design with six replicates per group. The control group was provided with a basal diet, while treatments 1 (LA) and 2 (HA) were supplemented with 0.5 g/d and 1 g/d PCP, respectively. PCP was purchased from Nanjing Herd Source Bio-technology Co., Ltd., Nanjing, China. The anthocyanin composition of PCP was detected according to Tian et al. [
11], and PCP had 2619.04 µg/g total anthocyanin concentration in this study. The level of purple corn pigment followed Tian et al. [
10]. The PCP was mixed in concentrate, and then mixed with roughage to prepare total mixed rations. All experimental kids were housed in clean individual pens and were provided drinking water
ad libitum. Equal amounts of rations were provided twice daily at 08:30 and 16:30 for
ad libitum intake. The nutrient requirements of experimental animals were according to the National Research Council (NRC) [
12]. The ingredients and nutrient composition of experimental diets was list in
Table 1.
2.2. Sample Collection
Approximately 100 g basal diet was collected once weekly and pooled during the experimental time period. The samples were dried at 65 °C in a vacuum oven for 72 h, then ground and passed through a 1-mm sieve.
One day before the end of the experiment, blood samples (approximately 10 mL) were taken at 0 h and 3 h from the jugular vein using a vacuette tube with heparin sodium (Taizhou Qiujing Medical Instrument Co., Ltd., Taizhou, China). The blood samples were transferred to a 1.5-mL tube after centrifugation (Allegra® X-30R Centrifuge, Beckman Coulter, Brea, CA, USA) at 3000× g for 15 min at 4 °C and kept at −20 °C until plasma lipid metabolism was analyzed.
Six goats per group were slaughtered at the end of the experiment, and the processing of carcasses was as described by Danforth [
13]. Muscle samples were divided into three portions. One aliquot was weighed and put into a freeze dryer (SCIENTZ-18N, Ningbo Scientz Biotechnology Co., Ltd., Ningbo, China) for 72 h to calculate moisture, then ground and passed through a 1-mm sieve until being analyzed for chemical composition. Another aliquot was stored at −20 °C until flavor compounds were detected. The last aliquot was immediately transferred to a 1.5-mL tube with RNA wait (Cat#SR0020, Beijing Solarbio Science and Technology Co., Ltd., Beijing, China) and stored at −80 °C until the gene expression assay was performed.
2.3. Chemical Composition
The contents of moisture/dry matter (DM), crude protein (CP), ether extract (EE), ash, calcium (Ca) and phosphorus (P) were measured in feed and muscle as per the method of Association of Official Analytical Chemists (AOAC) [
14]. Neutral detergent fiber (NDF) and acid detergent fiber (ADF) were determined according to Van Soest et al. [
15]. Gross energy (GE) was analyzed by a calorimeter (WGR-WR3, Changsha BENTE Instrument Co., Ltd., Changsha, China). Each index was run in triplicate.
2.4. Plasma Lipid Mechanism
The concentrations of total cholesterol (T-CHO; code No. A111-2-1), triglyceride (TG; code No. A110-2-1), creatinine (Cr; code No. C011-1-1), low-density lipoprotein cholesterol (LDL-C; code No. A113-1-1), and high-density lipoprotein cholesterol (HDL-C; code No. A112-1-1) were determined using commercial kits purchased from Jiancheng Bioengineering Institute (Nanjing, China). All procedures were strictly completed in accordance with reagent instructions.
2.5. Flavor Compound Profiles
A total of 4 g of chopped and mixed sample was weighed and transferred to a solid phase microextraction instrument and run in a manual injector with a 2 cm–50/30 μm DVB/CAR/PDMS StableFlex fiber tip. The LTL was then subjected to the analysis of volatile compounds using a gas chromatograph (GC; Agilent Technologies, Santa Clara, CA, USA) and tandem mass spectrometer (MS; SCIEX-6500Qtrap; AB Allen-Bradley, Milwaukee, WI, USA) with splitless injecting the sample after headspace extraction-temperature for 60 min at 80 °C. The GC conditions were as follows: the chromatographic column was an Agilent 19091S-436HP-5MSfused silica capillary column (60 m × 250 μm × 0.25 μm). The initial temperature was 40 °C for 2 min, increasing to 180 °C at the rate of 3.5 °C/min, and then to 310 °C at the rate of 10 °C/min. Total run time was 55 min. The temperature of the boil room was 250 °C; the carrier gas was He (99.99%). The pressure before columniation was 15.91 psi; the carrier gas flow rate was 1.0 mL/min, and the solvent delay time was 3 min. The MS conditions were as follows: the ion source was EI ionization; the ionization temperature was 230 °C, the Quadrupole temperature was 150 °C; the energy of ionization was 70 eV, with an emission current of 34.6 μA and multiplier voltage of 1847 v; the interface temperature was 280 °C, and the mass range was 29–500 amu. The peaks in the total ion flow diagram were retrieved by the MS computer data system, and component qualitative analysis was performed via comparison with the standard mass spectra in Nist17 and Wiley275 libraries to determine volatile chemical components. The area normalization method was used to calculate the peak area percentage of each of flavor compound.
2.6. Calculation of Relative Odor Activity Value
The contributions of volatile compounds toward the flavor were analyzed by relative odor activity value (ROAV) using the following equation [
16]:
where C
i and T
i are the relative content and odor threshold concentration, respectively; C
max and T
max are the highest relative content and odor threshold concentration, respectively. The most important flavor component of ROAV was defined as 100. In general, compounds with ROAV >1 contribute to the aroma, whereas compounds with ROAV < 1 may not.
2.7. Gene Expression
Total RNA was extracted from the LTL as follows: a 100 mg sample was transferred into a 1.5-mL tube, and 1-mL of RNA extracting solution (Cat. No. G3013; Wuhan Servicebio Technology Co., Ltd., Wuhan, China) was added and run in a homogenizer. The supernatant was immediately transferred to a 1.5-mL tube after centrifugation at 12,000× g for 10 min at 4 °C (D3024R; DragonLab, Beijing, China). Next, before centrifugation at 12,000× g for 10 min at 4 °C, 250 mL chloroform was added and the tube shaken vigorously, and the mixture was let stand for 3 min. A volume of 400 µL of the supernatant was transferred to a new tube; 320 µL isopropanol (Cat. No. 80109218; Sinopharm Chemical Reagent Co., Ltd., Shanghai, China) was added, and the tube was shaken vigorously and kept at −20 °C for 15 min. The sample was centrifuged at 12,000× g for 10 min at 4 °C; the supernatant was removed, and 1.5 mL of 75% ethanol (Cat. No. 10009218; Sinopharm Chemical Reagent Co., Ltd., Shanghai, China) was added to wash the precipitate. The supernatant was removed after centrifugation at 12,000× g for 10 min at 4 °C, and the centrifuge tube was placed on a super clean table and aerated for 3 min. The total RNA was dissolved with HyPureTM Molecular Biology Grade Water (Cat. No. SH30538.02; HyClone, Logan, UT, USA) and incubated at 55 °C for 5 min. The concentration and purity of RNA were detected by a NanoDrop 2000 (Thermo Fisher Scientific, Waltham, MA, USA). All sample concentrations were adjusted to 200 ng/μL.
The cDNA synthesis was performed using a Servicebio® RT first strand cDNA synthesis kit (Cat. No. G3330; Wuhan Servicebio Technology Co., Ltd., Wuhan, China) with 20 µL reaction volumes containing 4 μL of 5× reaction buffer, 0.5 μL of oligo (dT)18 primer (100 μM), 0.5 μL of random hexamer primer (100 μM), 1 μL of Servicebio® RT enzyme mix, 10 μL of total RNA, and RNase-free water to a final volume of 20 µL.
All primers were designed using the primer 5 software and were synthesized by the Wuhan Servicebio Technology Co., Ltd. (Wuhan, China;
Table 2). The three target genes were peroxisome proliferator-activated receptor gamma (
PPARγ), stearoyl-CoA desaturase (
SCD), and lipoprotein lipase (
LPL). The reference gene was glyceraldehyde-3-phosphate dehydrogenase (GAPDH). The real-time PCR amplifications were performed in a 15-µL reaction volume by a StepOnePlus™ real-time PCR system (Applied Biosystems™, Waltham, MA, USA). The reaction system consisted of 7.5 μL of 2 × SYBR Green qPCR master mix (Cat. No. G3320; Wuhan Servicebio Technology Co., Ltd., Wuhan, China), 1.5 μL of forward and reverse primers (2.5 μM), 2.0 μL of cDNA, and 4.0 μL of ddH
2O. The real-time PCR amplification procedure comprised 10 min at 95 °C for pre-denaturation, 40 cycles of 15 s at 95 °C for denaturation, and 30 s at 60 °C for extension. The fluorescence signal was collected every 0.5 °C from 65 °C to 95 °C. Each sample was performed in triplicate.
2.8. Statistical Analysis
All data analysis was performed through the Statistical Analysis System 9.1.3 (SAS Institute, Cary, NC, USA) software using one-way analysis of variance. Six replications were used in the present study, which could make a significance level of 0.05 and a power greater than 0.80. The replicate was considered the experimental unit in all of the statistical analyses. The relative mRNA abundance was measured according to the 2
−ΔΔCt method. The average abundance of genes in control data was used as the calibrator. The relationships between gene expression and key volatile compounds of
LTL were analyzed by Pearson correlation coefficients (r) [
17].
p-values lower than 0.05 indicated significant differences between samples.
4. Discussion
The FRs may initiate the oxidative degradation of lipids, as lipid radicals can react with oxygen and form new FRs such as O
2−· and
·OH, and these can react with TG or free fatty acids (FA) to reinitiate the process in ruminant meat [
21]. Anthocyanins may affect the initial processes of lipid hydrolysis, micelle formation, and cholesterol transfer to intestinal cells [
22]. Xia et al. [
23] reported that adding anthocyanin-rich extract to the diet of mice reduced the concentrations of serum TG and T-CHO, thus improving the lipid profile. In addition, Hosoda et al. [
24] demonstrated that the feeding of anthocyanin-rich corn silage tended to lower plasma T-CHO concentration in dairy cows. The postulated mechanism of anthocyanins for inhibition of lipid metabolism was as follows: (1) anthocyanins can inhibit cholesterol synthesis by decreasing the gene expression of 3-hydroxy-3-methylglutaryl coenzyme A; (2) anthocyanins may reduce blood apo B–and apo C-III; and (3) anthocyanins may inhibit cholesteryl ester transfer protein [
25]. Yong et al. [
26] revealed that purple sweet potato anthocyanins could decrease sterol regulatory element-binding protein 1 level and the expression levels of the target genes acetyl-coenzyme A carboxylase and FA synthase. Similarly, Lee et al. [
27] demonstrated that anthocyanins could reduce TG in 3T3-L1 cells. Thus, feeding anthocyanin-rich PCP resulted in a reduction of plasma T-CHO, suggesting that anthocyanins may partly contribute to the actions of lipid metabolites. The autoxidation of lipids or the oxidation of lipids with UFAs leads to the formation of meat flavor compounds. Hence, anthocyanins inhibit plasma lipid metabolism that can modulate the formation of flavor compounds in goat muscle.
Animal meat products are composed of water, protein, fats, and minerals, and nutritional status can directly affect the chemical composition of meat. In the present experiment, no significant differences in nutrient composition were noted among the groups, indicating that anthocyanins had no effect on regulating nutritional components of goats. The reason may be related to the low bioavailability of anthocyanins in animals. In addition, factors affecting the stability of anthocyanin include pH, temperature, oxygen, light intensity, and enzymes [
28]. The chemical forms and biological activities of anthocyanins may be different when exposed to different pH and temperatures in the gastrointestinal tract. In addition, the red luteal cation is the most abundant molecular form for anthocyanins when the pH value is less than 2 [
29]. Ruminal fluid pH ranged from 7.19 to 7.22 in this study, and this may also negatively affect the absorption of anthocyanins.
Lipid oxidation in meat is one of the main causes of quality loss; oxidation adversely affects flavor and nutritive value, limiting the shelf-life of meat [
3]. The lipid hydroperoxides are decomposed into hydroxyl and alkoxy radicals, and then the FA chain adjacent to the alkoxy group splits to produce low molecular weight volatile compounds [
9]. Therefore, meat flavor is an important factor affecting the palatability and acceptability of ruminant meat by consumers [
30]. Anthocyanins may be transferred from feed to muscle, subsequently affecting the lipid metabolism of muscle [
31]. Specifically, anthocyanin is a powerful FR scavenger that can not only effectively prevent the oxidation reactions caused by active FRs but also protect the integrity of lipids [
32]. Moreover, the phenolic hydroxyl group of anthocyanins has a strong inhibitory effect on FRs, providing H atoms to FRs and thereby reducing the peroxide value, terminating the reactions of FRs and inhibiting lipid oxidation [
33]. As a result, the anthocyanin groups showed high amounts of volatile compounds, perhaps due to the addition of anthocyanins to the goat diet inhibiting lipid oxidation, delaying the decline of meat flavor quality, and enriching the types of flavor substances.
Although lipid oxidation is one of the main reasons for the deterioration of meat products, it is important for the formation of typical aromas in meat products. Alcohols are mainly derived from the lipid oxidation of meat [
34]. Of interest, the anthocyanin groups had higher relative contents of alcohols, perhaps because the antioxidant system in goats can meet its own needs. Indeed, lipid peroxidation can easily produce various off-flavor volatile compounds, such as alcohols, ketones, and aldehydes [
35]. The polyhydric alcohols in bulk fish oils suppress both oxidation and formation of volatile off-flavors [
36]. Hence, the increased alcohol content in meat after feeding anthocyanin may be a manifestation of the enhancement of antioxidant function. However, the mechanism involved is still unclear. Moreover, while an antioxidant may protect against FRs, it could have no effect at all or in certain circumstances may even act as a “pro-oxidant” that generates toxic oxygen species [
37]. The flavor compounds play a key role in the meat sensory attributes, but they are also considered important indicators for the stability of oxidative of lambs [
38]. It was one of important body protective mechanisms of anthocyanin that regulating the expression of apoptosis-associated protein and antioxidative enzyme by the nuclear factor erythroid 2-related factor 2 (
Nrf2) signing pathway [
39]. Therefore, adding anthocyanin to the diet can improve the antioxidant capacity of muscle and inhibit the excessive oxidation of lipid, reducing the generation of off-flavor and prolonging the shelf life of goat meat. In the present research, the feeding of purple corn anthocyanins showed low total aldehyde and ketone concentrations, suggesting that anthocyanins play a positive role in the aroma of meat.
The volatile compound contents do not reflect their true contributions to the aroma, because these compounds show different odor thresholds, leading to different sensitivity levels for consumers [
40]. As such, ROAV was used to express the contribution of volatile compounds to the aroma of goat meat. In addition, the concentration of flavor is determined by the threshold value, and only volatile components with low odor thresholds can make a direct contribution to flavor. Typically, flavor compounds are considered the key volatile components and are considered to significantly contribute to aroma when the value of ROAV is ≥1; when the value is in the range 0.1 ≤ ROAV < 1, the compounds also contribute to the flavor [
41]. Of interest, the feeding of anthocyanin increased 1-hexanol and 1-octanol concentrations, as well as they were the key flavor compounds in both anthocyanin groups (ROAV > 1), suggesting that anthocyanins could enrichment of vegetal, herbaceous, grease, and fruity flavors for goat meat. In short, adding PCP to the diet enriched the flavor substances of goat meat, making the flavor of meat more harmonious. Indeed, lipid oxidation has a negative effect on meat, whereas it can help to form pleasant aromas in some cases [
42]. Consistent with our results, Prommachart et al. [
43] who showed that purple corn anthocyanin extracted residue in cattle diet could decrease lipid oxidation and increase UFA, might improve flavor compounds in meat.
The UFAs react with molecular oxygen to produce unstable compounds via a FR mechanism, and they produce various flavor compounds that include hydrocarbons, aldehydes, ketones, alcohols, esters, and acids, resulting in off-flavors and off-odors in meat [
44]. Kortenska and Yanishlieva [
45] showed that phenol could inhibit lipid oxidation as a result of the formation of a complex based on the hydrogen bonds between the hydroxy compounds and the phenol antioxidants. The basic chemical structure of anthocyanins is 3,5,7-three hydroxyl-2-phenyl benzopyran, yielding high efficiency in capturing peroxy radicals in the process of lipid oxidation [
46]. Hence, the inclusion of anthocyanins can decrease the levels of total hydrocarbons, aromatics, esters, and miscellaneous compounds in goat muscle. Collectively, adding anthocyanins to the goat diet can improve the types and relative contents of meat flavor and can enrich the vegetal, herbaceous, grease, and fruity flavors.
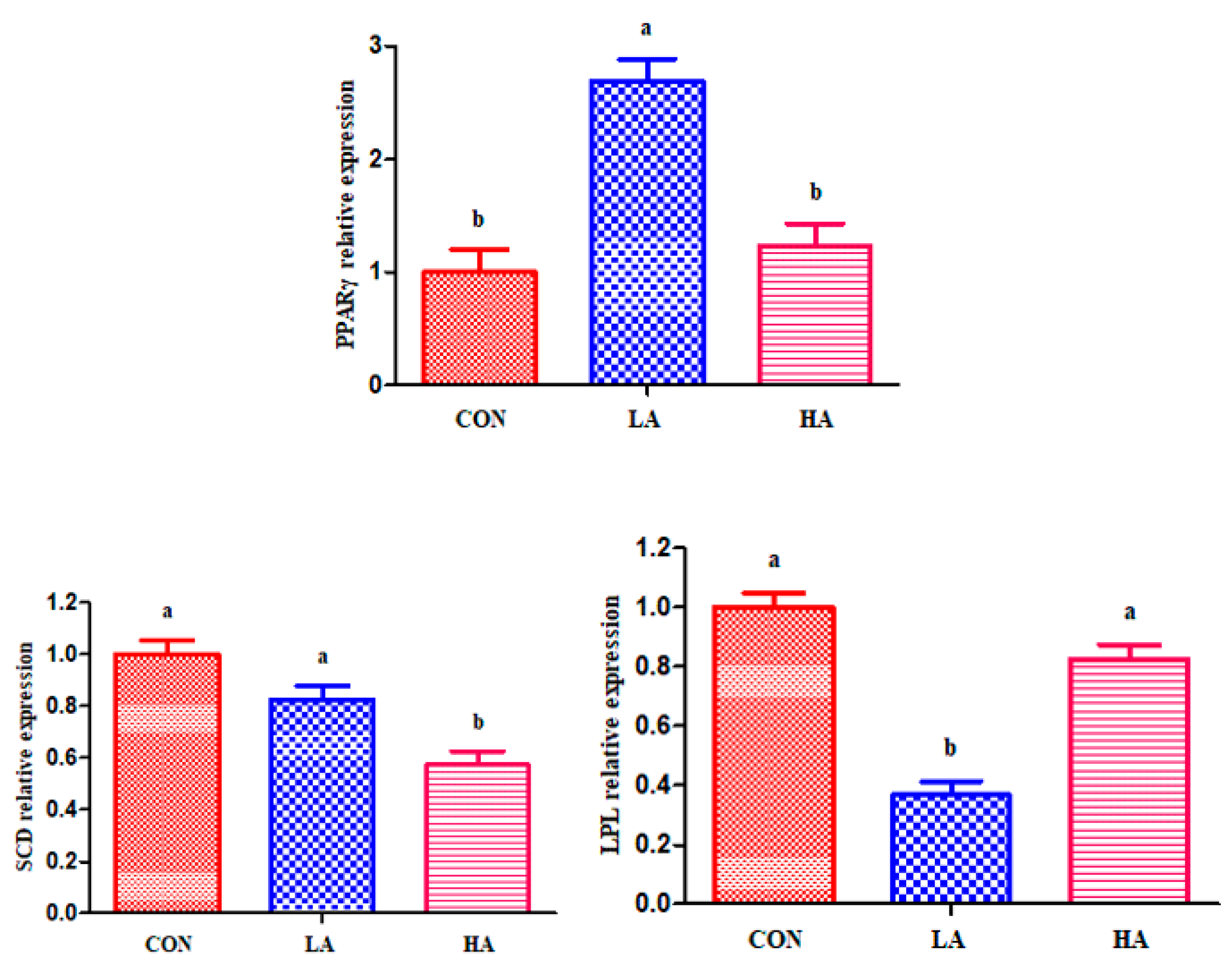
Anthocyanins have been demonstrated to prevent lipid oxidation due to their special properties [
11]. Thus, the formation of flavor compounds was verified by detecting gene expression levels of related signaling pathways. The peroxisome proliferator activated receptors (
PPARs) are nuclear hormone receptors activated by FAs and their derivatives, and they play a central role in lipid homeostasis. Anthocyanins can improve antioxidant capacity by activating the
Nrf2 signaling pathway [
47]. The Nrf2 is involved in the
PPARs pathway because of its mechanisms of action [
48]. In addition, anthocyanins may act as
PPARγ activators, inducing the adipose tissue remodeling and upregulation of peroxisome
PPARγ gene expression [
47]. Therefore, the feeding of 0.5 g/d PCP had significant relative mRNA abundance for
PPARγ. Of interest, feeding high amounts (1 g/d) of anthocyanins did not increase
PPARγ gene expression; this may be because anthocyanins have strong antioxidant activity, and the feeding of low amounts of anthocyanins was unable to alleviate oxidative stress and improve antioxidant capacity.
The
SCD plays an important role in regulating lipogenesis-related gene expression and in modulating mitochondrial UFA oxidation. Nichols et al. [
49] demonstrated that citrus flavonoids can reduce plasma lipid levels, enhance glucose tolerance, and inhibit hyperlipidemia and obesity by reducing the mRNA level of the
SCD1 gene in the liver. In addition, cholesterol was the main factor affecting
SCD activity [
50]. Accordingly, the abundance of muscle
SCD mRNA decreased after feeding goats anthocyanin, perhaps because anthocyanin led to a decrease in plasma T-CHO, inhibiting
SCD activity. Our observations concur with those of Lee et al. [
27], who indicated that anthocyanins suppress lipid accumulation by reducing gene and protein expression levels of lipogenic transcription factors of
PPARγ and
SCD-1.
The
LPL is a key enzyme of lipid metabolism. Its primary function is the hydrolysis of the core TG of circulating chylomicrons and very low-density lipoprotein (VLDL). There are two main sources of FAs involved in the
PPAR signaling pathway: (1) exogenous sources from circulating FA-albumin complexes or from
LPL-mediated hydrolysis of plasma VLDL and chylomicrons; and (2) endogenous de novo synthesis [
51]. Duivenvoorden et al. [
52] showed that mice with increased
LPL activity had higher fat tissue mass and were more insulin-resistant. Therefore, the feeding of 0.5 g/d PCP had increased abundance of
PPARγ, and consequently decreased
LPL gene expression. Consistent with our results, Kowalska et al. [
53] demonstrated that anthocyanin-rich cranberries could inhibit lipid metabolism by down-regulation of the mRNA level of
LPL in the adipose tissue. In short, anthocyanin may change
LTL flavor compound profiles of goats by activating the expression of
PPARγ and inhibiting the expression of
SCD and
LPL.Anthocyanins can decrease hepatic lipid accumulation and counteract oxidative stress and hepatic inflammation [
54]. The flavor substances derived from the oxidative rancidity of FAs and the addition of anthocyanin in the diet of goats consequently decreased the levels of flavor compounds. In addition, anthocyanin could modulate the expression of related lipid metabolism genes. Interestingly,
SCD and
LPL are the key genes that regulate the lipid metabolism in ruminants. Thus, significant positive correlations were detected between
SCD and ethanol, as well as
LPL and ehanol, 1-heptanol, 1-heptanol, (Z)-2-octen-1-ol, hexanal, heptanal, and decanal. Anthocyanin-rich plants could alter UFAs biohydrogenation in the rumen, increasing meat content of beneficial FAs, particularly UFA profiles [
55]. The UFAs are more prone to lipid oxidation that may lead to more flavor substances in food products. In the current study, significant negative correlations were observed between the mRNA expression of
PPARγ and hexanal and heptanal, indicating that anthocyanins may modulate the expression of lipid genes and ultimately increase the flavor of meat. Our data provide evidence that lipid mechanisms in plasma and related gene expression levels in
LTL may be cooperatively involved in anthocyanins regulating the formation of flavor compounds.



 ,
, 

{kind=link}