
Assessment of Intestinal Immunity and Permeability of Broilers on Partial Replacement Diets of Two-Stage Fermented Soybean Meal by Bacillus velezensis and Lactobacillus brevis ATCC 367

 ,
,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Preparation and Characteristics of FSBM
2.2. Animal Experiment
2.2.1. Animal Feeding and Housing
2.2.2. Growth Performance and Sample Collection
2.2.3. Jejunum Total RNA Isolation and qPCR
2.3. Statistical Analysis
3. Results
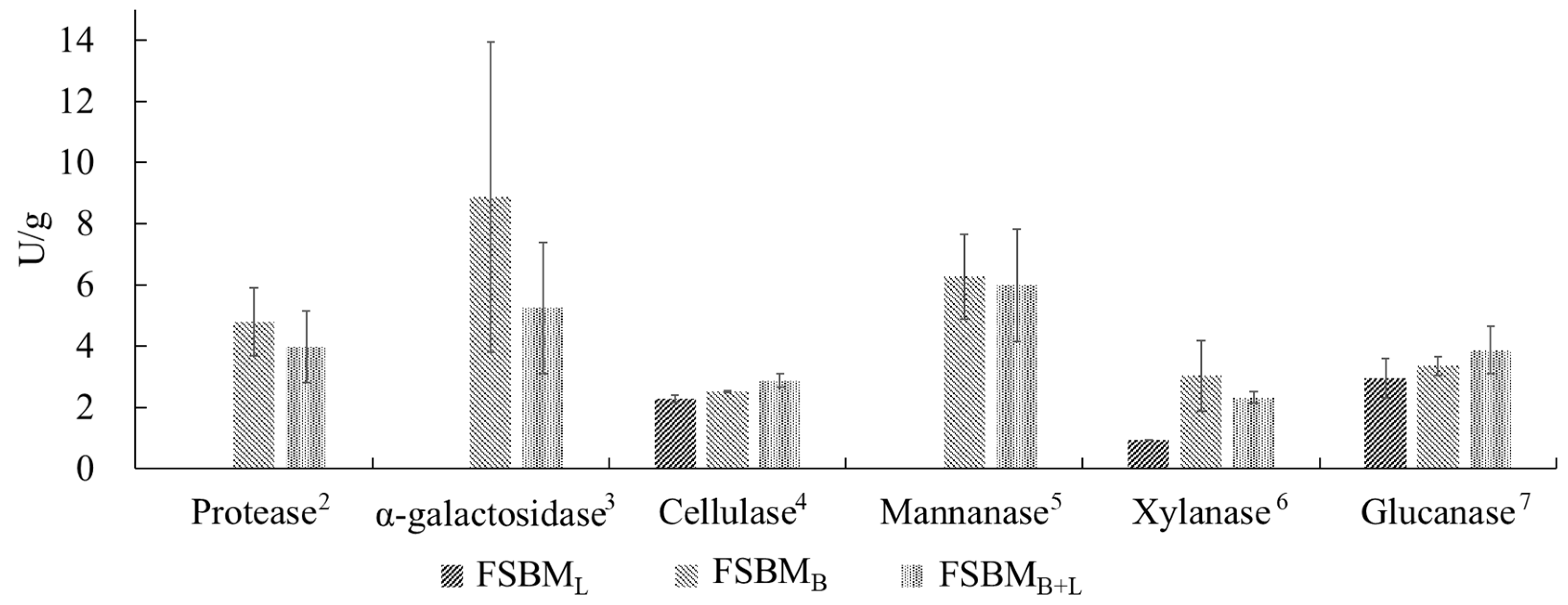
3.1. The Composition and Enzyme Activity of SBM and FSBM by Bv and Lb
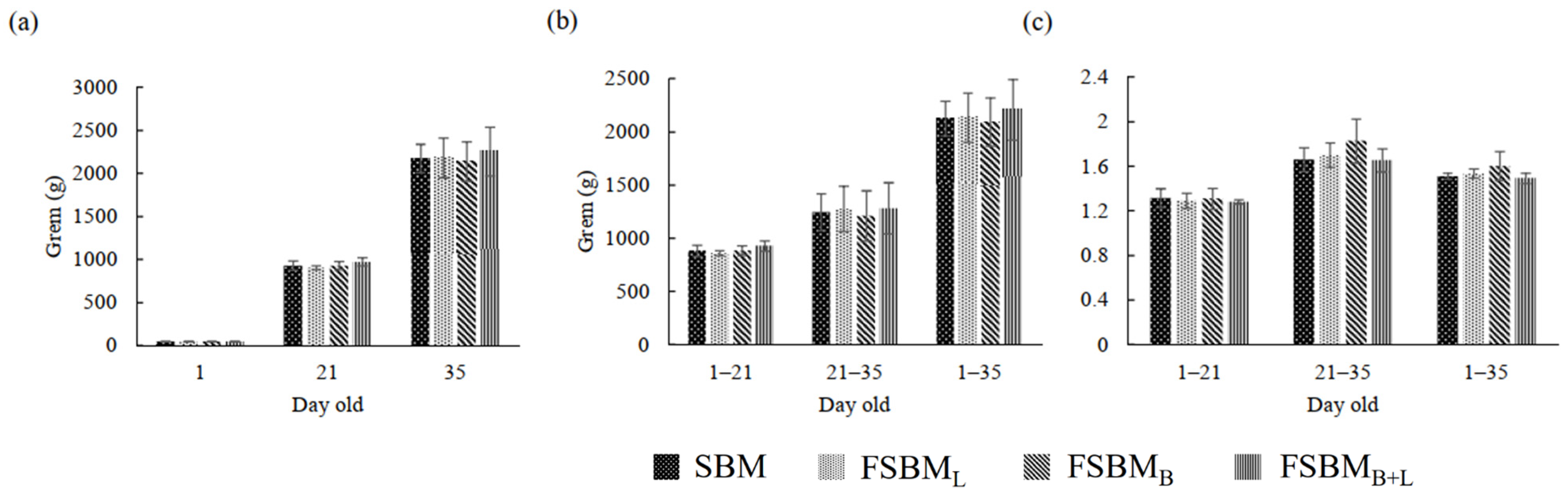
3.2. Effect of Two-Stage FSBM on Growth Performance of 1 to 35-Day-Old Broilers
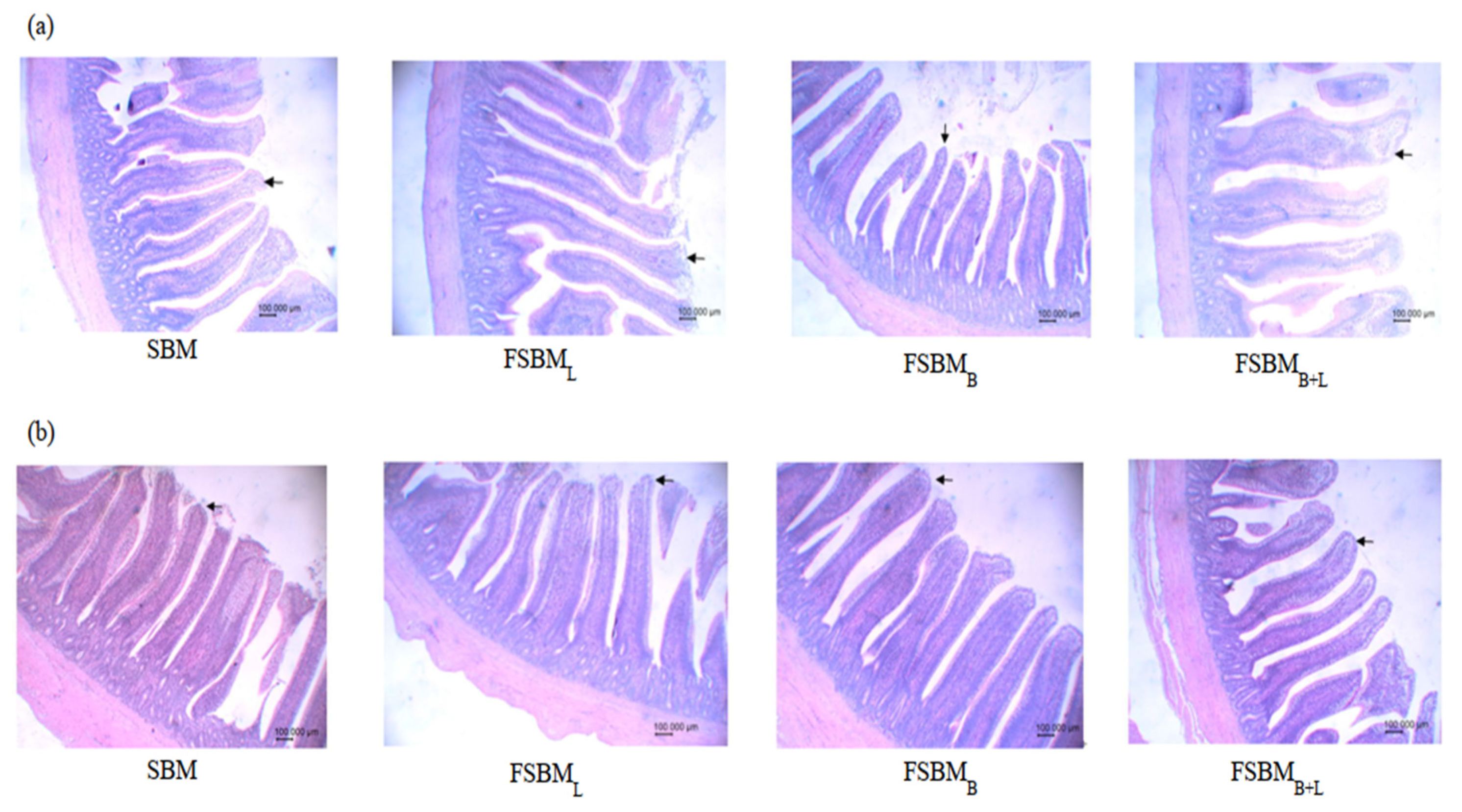
3.3. Effect of Two-Stage FSBM on Intestinal Morphology of 35-Day-Old Broilers
3.4. Effect of Two-Stage FSBM on Serum and Jejunum’ s Immunity of 35 Day-Old Broilers
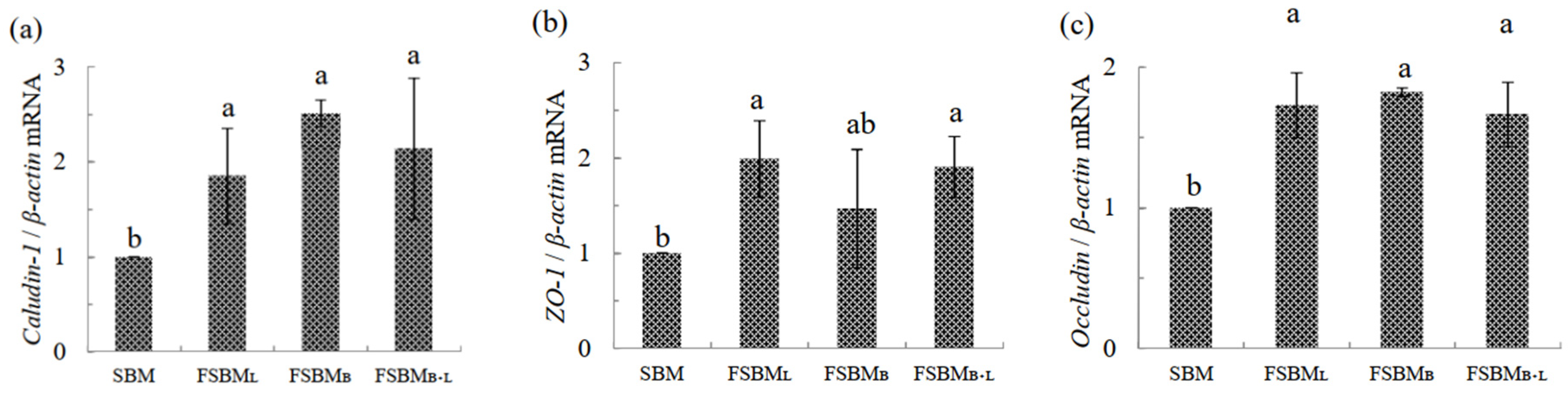
3.5. Effect of Two-Stage FSBM on Relative mRNA Expression in Jejunum of 35 Day-Old Broilers
3.6. Effect of Two-Stage FSBM on Economic Benefits
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
Abbreviations
| FSBM | Fermented soybean meal |
| Bv | Bacillus velezensis |
| Lb | Lactobacillus brevis ATCC 367 |
| FSBMB+L | Two-stage fermented soybean meal by Bacillus velezensis and Lactobacillus brevis ATCC 367 |
| SBM | Soybean meal |
| GOS | Galacto-oligosaccharide |
| FSBMB | One-stage fermented soybean meal by Bacillus velezensis |
| FSBML | Commercial fermented soybean meal products fermented by lactobacillus spp. |
| CLDN-1 | Claudin-1 |
| ZO-1 | Zonula occludens 1 |
| NFκB | Nuclear factor kappa B p 65 |
| IL-6 | Interleukin-6 |
| MUC2 | Mucin2 |
| ACE-inhibitory | Angiotensin-converting-enzyme inhibitory |
| Th1 | T Helper 1 Cells |
| IL-12 | Interleukin- |
| TNF-α | Tumor Necrosis Factor-α |
| IFN-γ | Interferon-γ |
| TGF-β | Transforming Growth Factor Beta |
| IL-17 | Interleukin- |
| IL-10 | Interleukin- |
| PepT-1 | Peptide transporter 1 |
| SEM | Standard error of mean |
| TCA- soluble protein | Trichloroethanoic acid- soluble protein |
| BW | Body weight |
| FI | Feed intake |
| BWG | Body weight gain |
| FCR | Feed conversion rate |
| IL-1ß | Interleukin-1ß |
| ROS | Reactive oxygen species |
References
- Nahashon, S.N.; Kilonzo-Nthenge, A.K. Advances in soybean and soybean by-products in monogastric nutrition and health. Soybean Nutr. 2011, 7, 125–156. [Google Scholar]
- Soumeh, E.A.; Mohebodini, H.; Toghyani, M.; Shabani, A.; Ashayerizadeh, A.; Jazi, V. Synergistic effects of fermented soybean meal and mannan-oligosaccharide on growth performance, digestive functions, and hepatic gene expression in broiler chickens. Poult. Sci. 2019, 98, 6797–6807. [Google Scholar] [CrossRef]
- Guo, S.; Zhang, Y.; Cheng, Q.; Xv, J.; Hou, Y.; Wu, X.; Du, E.; Ding, B. Partial substitution of fermented soybean meal for soybean meal influences the carcass traits and meat. Animals 2020, 10, 225. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Sembratowicz, I.; Chachaj, R.; Krauze, M.; Ognik, K. The effect of diet with fermented soybean meal on blood metabolites and redox status of chickens. Ann. Anim. Sci. 2020, 20, 599–611. [Google Scholar] [CrossRef]
- Chachaj, R.; Sembratowicz, I.; Krauze, M.; Ognik, K. The effect of partial replacement of soybean meal with fermented soybean meal on chicken performance and immune status. J. Anim. Feed Sci. 2019, 28, 263–271. [Google Scholar] [CrossRef]
- Young, D.; Ibuki, M.; Nakamori, T.; Fan, M.; Mine, Y. Soy-derived di- and tripeptides alleviate colon and ileum inflammation in pigs with dextran sodium sulfate-induced colitis. J. Nutr. 2012, 142, 363–368. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, B.; Moon, K.M.; Kim, C.Y. Tight junction in the intestinal epithelium: Its association with diseases and regulation by phytochemicals. J. Immunol. Res. 2018, 2018, 2645465. [Google Scholar] [CrossRef] [Green Version]
- Barve, A.; Khor, T.O.; Nair, S.; Lin, W.; Yu, S.; Jain, M.R.; Chan, J.Y.; Kong, A. Pharmacogenomic profile of soy isoflavone concentrate in the prostate of Nrf2 deficient and wild-type mice. J. Pharm. Sci. 2008, 97, 4528–4545. [Google Scholar] [CrossRef]
- Tonolo, F.; Moretto, L.; Grinzato, A.; Fiorese, F.; Folda, A.; Scalcon, V.; Ferro, S.; Arrigoni, G.; Bellamio, M.; Feller, E.; et al. Fermented soy-derived bioactive peptides selected by a molecular docking approach show antioxidant properties involving the Keap1/Nrf2 pathway. Antioxidants 2020, 9, 1306. [Google Scholar] [CrossRef]
- Sanjukta, S.; Rai, A.K. Production of bioactive peptides during soybean fermentation and their potential health benefits. Trends Food Sci. Technol. 2016, 50, 1–10. [Google Scholar] [CrossRef]
- Hou, Y.; Wu, Z.; Dai, Z.; Wang, G.; Wu, G. Protein hydrolysates in animal nutrition: Industrial production, bioactive peptides, and functional significance. J. Anim. Sci. Biotechnol. 2017, 8, 24. [Google Scholar] [CrossRef] [Green Version]
- Chatterjee, C.; Gleddie, S.; Xiao, C.W. Soybean bioactive peptides and their functional properties. Nutrients 2018, 10, 1211. [Google Scholar] [CrossRef] [Green Version]
- Karaś, M. Influence of physiological and chemical factors on the absorption of bioactive peptides. Int. J. Food Sci. Technol. 2019, 54, 1486–1496. [Google Scholar] [CrossRef]
- Cui, J.; Xia, P.; Zhang, L.; Hu, Y.; Xie, Q.; Xiang, H. A novel fermented soybean, inoculated with selected Bacillus, Lactobacillus and Hansenula strains, showed strong antioxidant and anti-fatigue potential activity. Food Chem. 2020, 333, 127527. [Google Scholar] [CrossRef]
- Zhang, Y.; Chen, S.; Zong, X.; Wang, C.; Shi, C.; Wang, F.; Wang, Y.; Lu, Z. Peptides derived from fermented soybean meal suppresses intestinal inflammation and enhances epithelial barrier function in piglets. Food Agr. Immunol. 2020, 31, 120–135. [Google Scholar] [CrossRef] [Green Version]
- Yang, X.J.; Li, W.L.; Feng, Y.; Yao, J.H. Effects of immune stress on growth performance, immunity, and cecal microflora in chickens. Poult. Sci. 2011, 90, 2740–2746. [Google Scholar] [CrossRef]
- Mishra, B.; Jha, R. Oxidative stress in the poultry gut: Potential challenges and interventions. Front. Vet. Sci. 2019, 6, 60. [Google Scholar] [CrossRef] [PubMed]
- Cheng, Y.; Hsiao, F.S.; Wen, C.; Wu, C.; Dybus, A.; Yu, Y. Mixed fermentation of soybean meal by protease and probiotics and its effects on the growth performance and immune response in broilers. J. Appl. Anim. Res. 2019, 47, 339–348. [Google Scholar] [CrossRef] [Green Version]
- AOAC. Official Methods of Analysis of AOAC International, 19th ed.; AOAC International: Gaithersburg, MD, USA, 2012. [Google Scholar]
- Faridah, D.N.; Andarwulan, N.; Purnomo, E.P.; Saifatah, L.; Kurniawan, S.K. Oligosaccharide content and in vitro protein digestibility of twenty commercial soy-based powder drinks in Indonesia. J. Chem. Soc. Chem. Commun. 2014, 26, 7687–7692. [Google Scholar] [CrossRef]
- Yin, L.; Tai, H.; Lee, H.; Jiang, S. Proteolysis and lactobacillus fermentation effect on the isoflavones biotransformation and removal of anti-nutritional factors of soy bean. J. Mar. Sci. Technol. 2014, 22, 525–530. [Google Scholar]
- Xie, J.; Du, M.; Shen, M.; Wu, T.; Lin, L. Physico-chemical properties, antioxidant activities and angiotensin-I converting enzyme inhibitory of protein hydrolysates from mung bean (Vigna radiate). Food Chem. 2019, 270, 243–250. [Google Scholar] [CrossRef]
- Olukomaiya, O.; Fernando, C.; Mereddy, R.; Li, X.; Sultanbawa, Y. Solid-state fermented plant protein sources in the diets of broiler chickens: A review. Anim. Nutr. 2019, 5, 319–330. [Google Scholar] [CrossRef]
- Wang, L.C.; Wen, C.; Jiang, Z.Y.; Zhou, Y.M. Evaluation of the partial replacement of high-protein feedstuff with fermented soybean meal in broiler diets. J. Appl. Poult. Res. 2012, 21, 849–855. [Google Scholar] [CrossRef]
- Choct, M.; Dersjant-Li, Y.; McLeish, J.; Peisker, M. Soy oligosaccharides and soluble non-starch polysaccharides: A review of digestion, nutritive and anti-nutritive effects in pigs and poultry. Asian-Aust. J. Anim. Sci. 2010, 23, 1386–1398. [Google Scholar] [CrossRef]
- Ibrahim, O.O. Functional oligosaccharides: Chemicals structure, manufacturing, health benefits, applications and regulations. J. Food Chem. Nanotechnol 2018, 4, 65–76. [Google Scholar] [CrossRef] [Green Version]
- Aliakbarpour, H.R.; Chamani, M.; Rahimi, G.; Sadeghi, A.A.; Qujeq, D. The Bacillus subtilis and lactic acid bacteria probiotics influences intestinal mucin gene expression, histomorphology and growth performance in broilers. Asian-Aust. J. Anim. Sci. 2012, 25, 1285–1293. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Lee, M.T.; Lin, W.C.; Lee, T.T. Potential crosstalk of oxidative stress and immune response in poultry through phytochemicals—A review. Asian-Aust. J. Anim. Sci. 2019, 32, 309–319. [Google Scholar] [CrossRef] [PubMed]
- Zihni, C.; Mills, C.; Matter, K.; Balda, M.S. Tight junctions: From simple barriers to multifunctional molecular gates. Nat. Rev. Mol. Cell Biol. 2016, 17, 564–580. [Google Scholar] [CrossRef] [PubMed]
- Peng, C.; Ding, X.; Zhu, L.; He, M.; Shu, Y.; Zhang, Y.; Li, Y.; Wang, X.; Feng, S.; Li, J.; et al. β-conglycinin-induced intestinal porcine epithelial cell damage via the nuclear factor κB/mitogen-activated protein kinase signaling pathway. J. Agric. Food Chem. 2019, 67, 9009–9021. [Google Scholar] [CrossRef] [PubMed]
- Ren, J.; Yang, B.; Lv, Y.; Guo, S. Protective and reparative effects of peptides from soybean β-conglycinin on mice intestinal mucosa injury. Int. J. Food Sci. Nutr. 2014, 65, 345–350. [Google Scholar] [CrossRef] [PubMed]
- Liu, T.; Zhang, L.; Joo, D.; Sun, S. NF-κB signaling in inflammation. Signal Transduct. Target. Ther. 2017, 2, e17023. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Ipseiz, N.; Pickering, R.J.; Rosas, M.; Tyrrell, V.J.; Davies, L.C.; Selinda, J.O.; Czubala, M.A.; Fathalla, D.; Robertson, A.A.; Bryant, C.E.; et al. Tissue-resident macrophages actively suppress IL-1beta release via a reactive prostanoid/IL-10 pathway. EMBO J. 2020, 39, e103454. [Google Scholar] [CrossRef] [PubMed]
- Carter, A.J.; Adams, M.R.; Woodward, M.J.; La Ragione, R.M. Control strategies for Salmonella colonisation of poultry: The probiotic perspective. Food Nutr. Bull. 2009, 5, 103–115. [Google Scholar]
- Bouma, G.; Strober, W. The immunological and genetic basis of inflammatory bowel disease. Nat. Rev. Immunol. 2003, 3, 521–533. [Google Scholar] [CrossRef] [PubMed]
- Song, Y.S.; Frias, J.; Martinez-Villaluenga, C.; Vidal-Valdeverde, C.; de Mejia, E.G. Immunoreactivity reduction of soybean meal by fermentation, effect on amino acid composition and antigenicity of commercial soy products. Food Chem. 2008, 108, 571–581. [Google Scholar] [CrossRef] [PubMed]
- Singh, B.P.; Vij, S.; Hati, S. Functional significance of bioactive peptides derived from soybean. Peptides 2014, 54, 171–179. [Google Scholar] [CrossRef] [PubMed]
- Xia, J.; Zu, Q.; Yang, A.; Wu, Z.; Li, X.; Tong, P.; Yuan, J.; Wu, Y.; Fan, Q.; Chen, H. Allergenicity reduction and rheology property of lactobacillus-fermented soymilk. J. Sci. Food Agric. 2019, 99, 6841–6849. [Google Scholar] [CrossRef]
- Wang, T.; Qin, G.; Sun, Z.; Zhao, Y. Advances of research on glycinin and β-conglycinin: A review of two major soybean allergenic proteins. Crit. Rev. Food Sci. Nutr. 2013, 54, 850–862. [Google Scholar] [CrossRef]
- Netea, M.G.; Domínguez-Andrés, J.; Barreiro, L.B.; Chavakis, T.; Divangahi, M.; Fuchs, E.; Joosten, L.A.B.; van der Meer, J.W.M.; Mhlanga, M.M.; Mulder, W.J.M.; et al. Defining trained immunity and its role in health and disease. Nat. Rev. Immunol. 2020, 20, 375–388. [Google Scholar] [CrossRef] [Green Version]
- Zhu, J.; Gao, M.; Zhang, R.; Sun, Z.; Wang, C.; Yang, F.; Huang, T.; Qu, S.; Zhao, L.; Li, Y.; et al. Effects of soybean meal fermented by L. plantarum, B. subtilis and S. cerevisieae on growth, immune function and intestinal morphology in weaned piglets. Microb. Cell Factories 2017, 16, 191–201. [Google Scholar] [CrossRef] [PubMed]







{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
{kind=link}
| Nutrient | SBM | FSBML | FSBMB | FSBMB+L | SEM | p-Value |
|---|---|---|---|---|---|---|
| Composition | ||||||
| DM (%) | 88.4 c | 93.2 a | 88.2 c | 89.9 b | 0.001 | <0.001 |
| CP (% DM) | 43.0 | 45.1 | 50.8 | 48.1 | 1.72 | 0.104 |
| TCA-soluble protein (% DM) | 4.21 d | 7.42 c | 15.17 a | 10.29 b | 0.49 | <0.001 |
| Lactic acid Bacteria (Log CFU/g DM) | 5.54 c | 8.82 a | 8.00 b | 8.97 a | 0.08 | <0.001 |
| Anti-nutritive factors | ||||||
| Raffinose (% DM) | 1.29 a | 0.11 b | 0.08 b | 0.0 3b | 0.04 | <0.001 |
| Stachyose (% DM) | 4.15 a | 1.19 b | ND | ND | 0.08 | <0.001 |
| Allergen protein (mg/g DM) | 505 a | 226 b | 183 b | 198 b | 35.8 | 0.001 |
| Trpsin inhibitor (mg/g DM) | 17.5 a | 9.03 b | 0.43 d | 1.57 c | 0.16 | <0.001 |
| Ingredients | Starter Diet (Day 1–21) | Finisher Diet (Day 22–35) | ||||||
|---|---|---|---|---|---|---|---|---|
| SBM | FSBML | FSBMB | FSBMB+L | SBM | FSBML | FSBMB | FSBMB+L | |
| Composition, % | ||||||||
| Corn, yellow | 52.99 | 53.27 | 54.18 | 53.49 | 57.15 | 57.39 | 58.45 | 57.61 |
| Soybean meal (CP-44%) | 34.0 | 28.0 | 28.0 | 28.0 | 28.0 | 22.0 | 22.0 | 22.0 |
| Fermented soybean meal | - | 6.0 | 6.0 | 6.0 | - | 6.0 | 6.0 | 6.0 |
| Full fat soybean meal | 3.00 | 2.99 | 2.00 | 2.47 | 4.15 | 4.18 | 3.04 | 3.65 |
| Soybean oil | 3.16 | 2.89 | 2.97 | 3.20 | 4.13 | 3.86 | 3.93 | 4.17 |
| Fish meal (CP-65%) | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 | 3.00 |
| Monocalcium phosphate | 1.32 | 1.32 | 1.32 | 1.32 | 1.25 | 1.25 | 1.25 | 1.25 |
| Calcium carbonate | 1.36 | 1.36 | 1.36 | 1.36 | 1.28 | 1.28 | 1.28 | 1.28 |
| NaCl | 0.34 | 0.34 | 0.34 | 0.34 | 0.34 | 0.34 | 0.34 | 0.34 |
| DL-Methionine | 0.35 | 0.35 | 0.35 | 0.35 | 0.27 | 0.27 | 0.27 | 0.27 |
| L-Lysine HCl | 0.20 | 0.20 | 0.20 | 0.20 | 0.16 | 0.16 | 0.16 | 0.16 |
| Choline-Cl | 0.08 | 0.08 | 0.08 | 0.08 | 0.08 | 0.08 | 0.08 | 0.08 |
| Vitamin premix 2 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 |
| Mineral premix 3 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 | 0.10 |
| Total | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 | 100.0 |
| Calculated nutrient levels | ||||||||
| Crude protein, % | 23.0 | 23.0 | 23.0 | 23.0 | 21.0 | 21.0 | 21.0 | 21.0 |
| Crude fat, % | 6.6 | 6.4 | 6.3 | 6.6 | 7.8 | 7.7 | 7.6 | 7.8 |
| ME, kcal/kg | 3050.0 | 3050.0 | 3050.0 | 3050.0 | 3175.0 | 3175.0 | 3175.0 | 3175.0 |
| Calcium, % | 0.96 | 0.96 | 0.96 | 0.96 | 0.90 | 0.91 | 0.91 | 0.91 |
| Total phosphorus, % | 0.70 | 0.71 | 0.71 | 0.71 | 0.66 | 0.68 | 0.67 | 0.67 |
| Available phosphorus, % | 0.48 | 0.47 | 0.46 | 0.46 | 0.46 | 0.44 | 0.44 | 0.44 |
| Methionine,+Cysteine, % | 1.08 | 1.08 | 1.07 | 1.07 | 0.95 | 0.95 | 0.94 | 0.95 |
| Gene Name 1 | Primer Sequence | Genbank No. |
|---|---|---|
| ß-actin | F: 5′-CTGGCACCTAGCACAATGAA-3′ R: 5′-ACATCTGCTGGAAGGTGGAC-3′ | X00182.1 |
| NFκB | F: 5′-CCAGGTTGCCATCGTGTTCC-3′ R: 5′-GCGTGCGTTTGCGCTTCT-3′ | D13719.1 |
| IFN-γ | F: 5′-CTCCCGATGAACGACTTGAG-3′ R: 5′-CTGAGACTGGCTCCTTTTCC-3′ | Y07922 |
| IL-1ß | F: 5′-GCTCTACATGTCGTGTGTGATGAG-3′ R: 5′-TGTCGATGTCCCGCATGA-3′ | NM_204524 |
| IL-6 | F: 5′-AGGACGAGATGTGCAAGAAGTTC-3′ R: 5′-TTGGGCAGGTTGAGGTTGTT-3′ | NM_204628 |
| IL-10 | F: 5′-TTCAGCTTGGATGTGTGAGC-3′ R: 5′-TGTCAGTTCTGCATGCTTCC-3′ | XM_025143715.1 |
| Claudin-1 | F: 5′-GGAGGATGACCAGGTGAAGA-3′ R: 5′-TCTGGTGTTAACGGGTGTGA-3′ | NM_001013611.2 |
| MUC-2 | F: 5′-GCTACAGGATCTGCCTTTGC-3′ R: 5′-AATGGGCCCTCTGAGTTTTT-3′ | NM_001318434.1 |
| Occludin | F: 5′-GTCTGTGGGTTCCTCATCGT-3′ R: 5′-GTTCTTCACCCACTCCTCCA-3′ | NM_205128.1 |
| ZO-1 | F: 5′-AGGTGAAGTGTTTCGGGTTG-3′ R: 5′-CCTCCTGCTGTCTTTGGAAG-3′ | XM_015278975.1 |
| Items | Experimental Diet | |||||
|---|---|---|---|---|---|---|
| SBM | FSBML | FSBMB | FSBMB+L | SEM | p-Value | |
| Jejunum | ||||||
| Villus height (μm) | 1181 b | 1326 a | 1293 a | 1312 a | 26.2 | <0.001 |
| Crypt depth (μm) | 210 a | 206 a | 175 b | 196 a | 5.38 | <0.001 |
| Villus height/crypt depth | 5.62 c | 6.44 b | 7.39 a | 6.69 b | 0.18 | <0.001 |
| Ileum | ||||||
| Villus height (μm) | 955 | 971 | 965 | 985 | 17.5 | 0.676 |
| Crypt depth (μm) | 202 | 204 | 204 | 193 | 5.79 | 0.461 |
| Villus height/crypt depth | 4.73 | 4.76 | 4.73 | 5.10 | 0.15 | 0.125 |
| Items | Experimental Diet | |||||
|---|---|---|---|---|---|---|
| SBM | FSBML | FSBMB | FSBMB+L | SEM | p-Value | |
| Jejunum | ||||||
| IL-1β (pg/mg protein) | 10.2 b | 16.9 a | 11.7 b | 12.5 b | 1.15 | 0.016 |
| IL-10 (pg/mg protein) | 134 b | 208 a | 130 b | 135 b | 4.44 | <0.001 |
| IL-6 (pg/mg protein) | 655 b | 769 a | 510 c | 500 c | 36.3 | 0.002 |
| Serum | ||||||
| IL-1β (pg/mL) | 90.1 a | 43.1 c | 70.9 ab | 51.5 bc | 7.95 | 0.018 |
| Item | Experimental Diets | |||
|---|---|---|---|---|
| SBM | FSBML | FSBMB | FSBMB+L | |
| Feed cost, TWD/bird | ||||
| 1–35 days | 46.6 | 49.0 | 49. 6 | 49.3 |
| Meat income, TWD/bird | ||||
| 1–35 days | 108.6 | 108.9 | 106.8 | 112.6 |
| Income over feed cost, TWD/bird | ||||
| 1–35 days | 61.9 | 59.9 | 57.2 | 63.3 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Tsai, C.F.; Lin, L.J.; Wang, C.H.; Tsai, C.S.; Chang, S.C.; Lee, T.T. Assessment of Intestinal Immunity and Permeability of Broilers on Partial Replacement Diets of Two-Stage Fermented Soybean Meal by Bacillus velezensis and Lactobacillus brevis ATCC 367. Animals 2021, 11, 2336. https://doi.org/10.3390/ani11082336
Tsai CF, Lin LJ, Wang CH, Tsai CS, Chang SC, Lee TT. Assessment of Intestinal Immunity and Permeability of Broilers on Partial Replacement Diets of Two-Stage Fermented Soybean Meal by Bacillus velezensis and Lactobacillus brevis ATCC 367. Animals. 2021; 11(8):2336. https://doi.org/10.3390/ani11082336
Chicago/Turabian StyleTsai, Chia Fen, Li Jen Lin, Chao Humg Wang, Ching Sung Tsai, Shang Chang Chang, and Tzu Tai Lee. 2021. "Assessment of Intestinal Immunity and Permeability of Broilers on Partial Replacement Diets of Two-Stage Fermented Soybean Meal by Bacillus velezensis and Lactobacillus brevis ATCC 367" Animals 11, no. 8: 2336. https://doi.org/10.3390/ani11082336