
Effects of Cold Exposure on Performance and Skeletal Muscle Fiber in Weaned Piglets

 , , , , ,
, , , , ,
Abstract
:Simple Summary
Abstract
1. Introduction
2. Materials and Methods
2.1. Animals, Diets and Experimental Design
2.2. Growth Performance
2.3. Diarrhea Score
2.4. Sample Collection
2.5. Chemical Analysis
2.6. Determination of Antioxidant Parameters in Serum and Skeletal Muscle
2.7. Total RNA Extraction, Reverse Transcription and Quantitative Real-Time PCR
2.8. Statistical Analysis
3. Results
3.1. Growth Performance
3.2. Diarrhea Score
3.3. Nutrient Digestibility
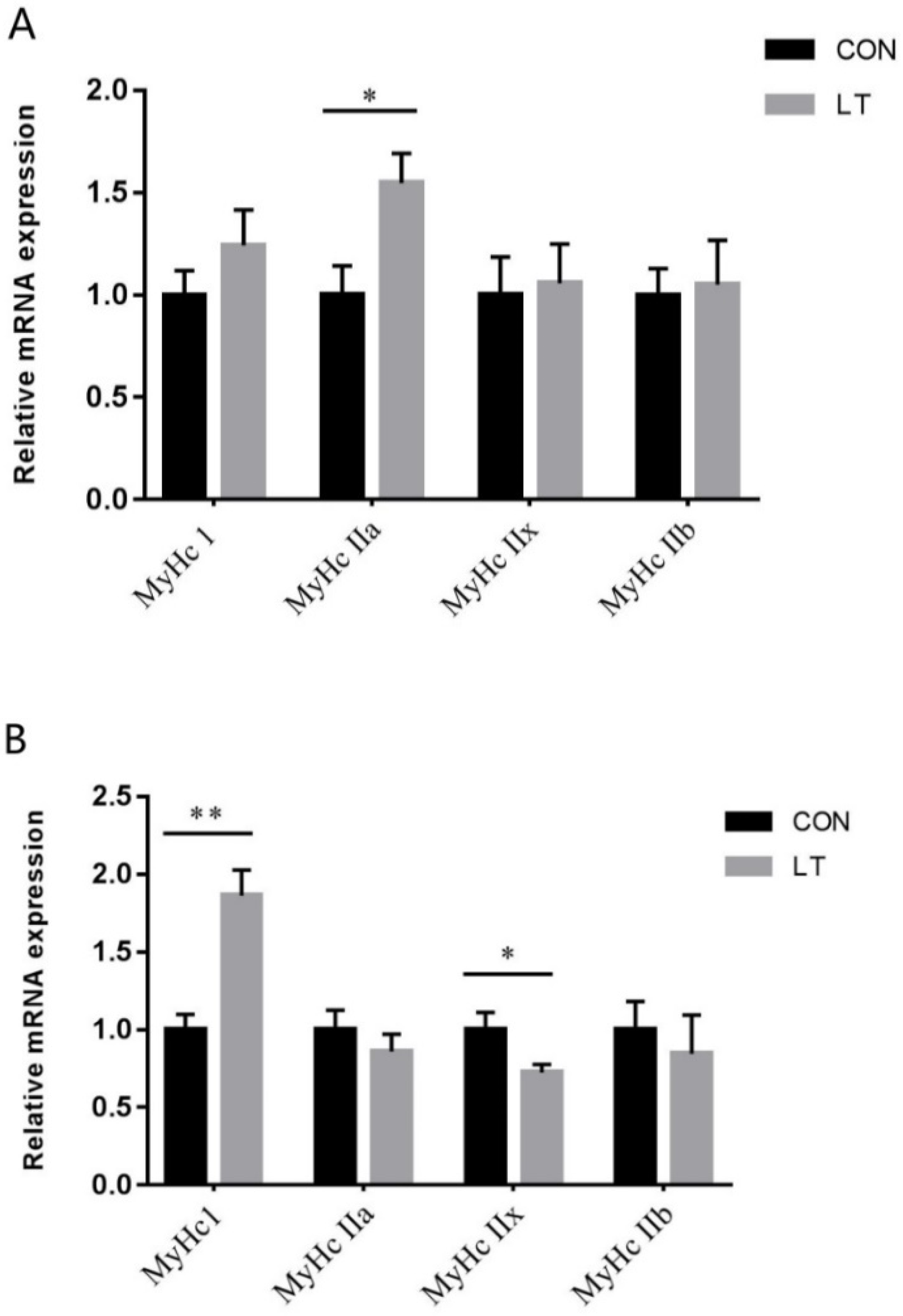
3.4. Skeletal Muscle Fiber Type-Related Gene Expression
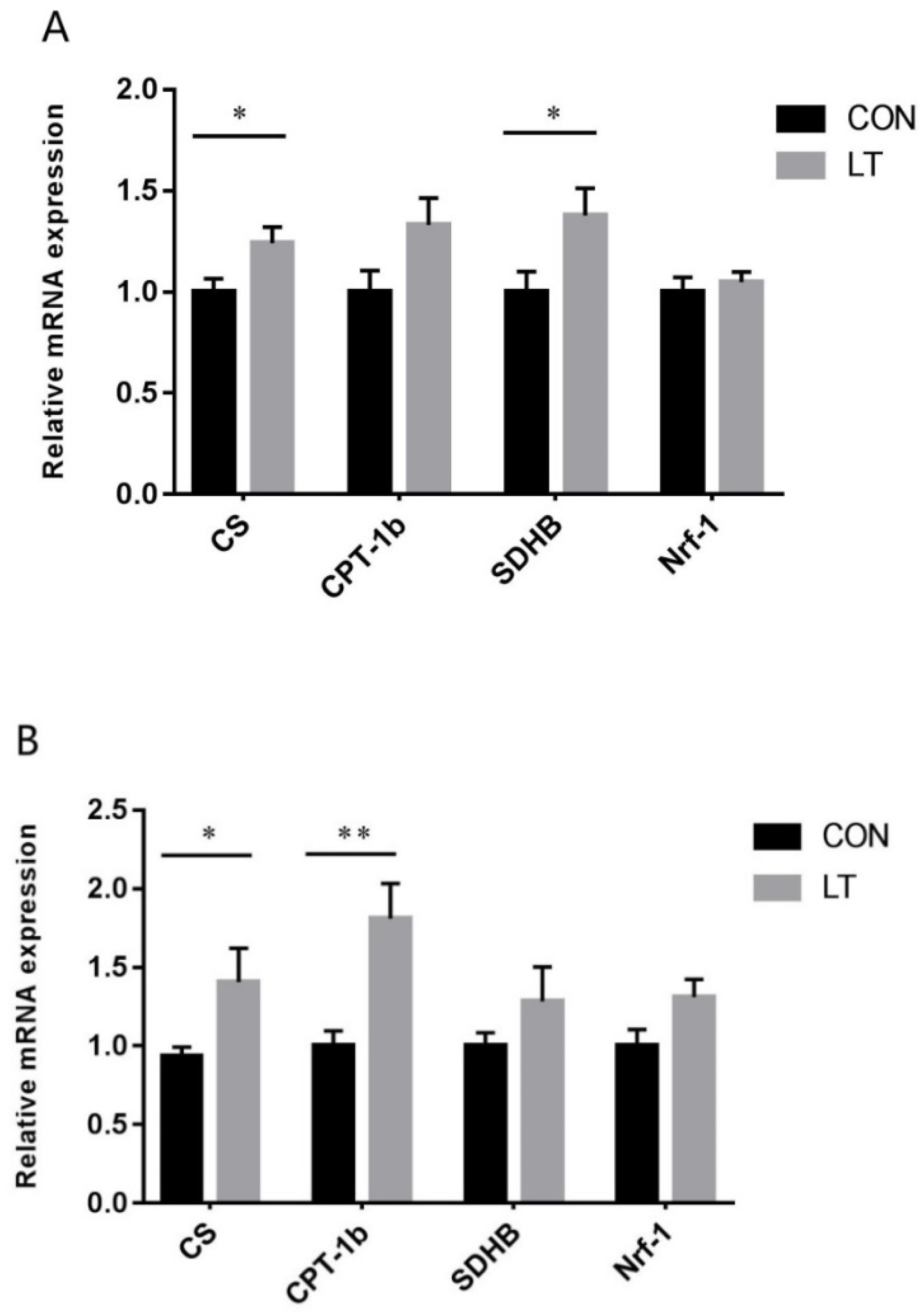
3.5. Mitochondrial Function-Related Gene Expression
3.6. Antioxidant Capacity
4. Discussion
5. Conclusions
Author Contributions
Funding
Institutional Review Board Statement
Data Availability Statement
Acknowledgments
Conflicts of Interest
References
- Mendes, A.A.; Watkins, S.E.; England, J.A.; Saleh, E.A.; Waldroup, A.L.; Waldroup, P.W. Influence of dietary lysine levels and arginine:lysine ratios on performance of broilers exposed to heat or cold stress during the period of three to six weeks of age. Poult. Sci. 1997, 76, 472–481. [Google Scholar] [CrossRef] [PubMed]
- Li, J.; Huang, F.; Li, X.; Su, Y.; Li, H.; Bao, J. Effects of intermittent cold stimulation on antioxidant capacity and mRNA expression in broilers. Livest. Sci. 2017, 204, 110–114. [Google Scholar] [CrossRef]
- Janský, L. Non-shivering thermogenesis and its thermoregulatory significance. Biol. Rev. 2010, 48, 85–132. [Google Scholar] [CrossRef] [PubMed]
- Henneman, E.; Clamann, H.P.; Gillies, J.D.; Skinner, R.D. Rank order of motoneurons within a pool: Law of combination. J. Neurophysiol. 1974, 37, 1338–1349. [Google Scholar] [CrossRef] [PubMed]
- Lefaucheur, L.; Hoffman, R.K.; Gerrard, D.E.; Okamura, C.S.; Rubinstein, N.; Kelly, A. Evidence for three adult fast myosin heavy chain isoforms in type II skeletal muscle fibers in pigs. J. Anim. Sci. 1998, 76, 1584–1593. [Google Scholar] [CrossRef] [PubMed]
- Berchtold, M.W.; Brinkmeier, H.; Muntener, M. Calcium ion in skeletal muscle: Its crucial role for muscle function, plasticity, and disease. Phys. Rev. 2000, 80, 1215–1265. [Google Scholar] [CrossRef] [Green Version]
- Dividich, J.L.; Vermorel, M.; Noblet, J.; Bouvier, J.C.; Aumaitre, A. Effects of environmental temperature on heat production, energy retention, protein and fat gain in early weaned piglets. Br. J. Nutr. 1980, 44, 313–323. [Google Scholar] [CrossRef] [Green Version]
- McCarthy, J.; Aherne, F.; Okai, D. Use of HCl insoluble ash as an index material for determining apparent digestibility with pigs. Can. J. Anim. Sci. 1974, 54, 107–109. [Google Scholar] [CrossRef]
- AOAC International. Official Methods of Analysis of AOAC International, 16th ed.; Trends in Food Science & Technology; AOAC International: Rockville, MD, USA, 1995; Volume 1, p. 382. [Google Scholar]
- Schmittgen, T.D. Real-time quantitative PCR. Methods 2001, 25, 383–385. [Google Scholar] [CrossRef] [Green Version]
- Stahly, T.; Cromwell, G.; Aviotti, M. The effect of environmental temperature and dietary lysine source and level on the performance and carcass characteristics of growing swine. J. Anim. Sci. 1979, 49, 1242–1251. [Google Scholar] [CrossRef]
- Lefaucheur, L.; Le, D.J.; Mourot, J.; Monin, G.; Ecolan, P.; Krauss, D. Influence of environmental temperature on growth, muscle and adipose tissue metabolism, and meat quality in swine. J. Anim. Sci. 1991, 69, 2844. [Google Scholar] [CrossRef] [PubMed]
- Kelley, K.W.; Blecha, F.; Regnier, J.A. Cold exposure and absorption of colostral immunoglobulins by neonatal pigs. J. Anim. Sci. 1982, 552, 363–368. [Google Scholar] [CrossRef] [PubMed]
- Sun, S.; Huang, X.; Hou, X.; Xie, X. Impaired accommodation of the proximal stomach to a colder meal in healthy adults. J. Clin. Intern. Med. 2005, 22, 678–681. [Google Scholar]
- Wigmore, P.M.; Stickland, N.C. Muscle development in large and small pig fetuses. J. Anat. 1983, 137, 235–245. [Google Scholar] [PubMed]
- Stickland, N.C.; Handel, S.E. The numbers and types of muscle fibres in large and small breeds of pigs. J. Anat. 1986, 147, 181–189. [Google Scholar] [PubMed]
- Picard, B.; Lefaucheur, L.C.; Duclos, M.J. Muscle fibre ontogenesis in farm animal species. Reprod. Nutr. Dev. 2002, 42, 415–431. [Google Scholar] [CrossRef] [Green Version]
- Chang, K.C. Key signalling factors and pathways in the molecular determination of skeletal muscle phenotype. Animal 2007, 1, 681–698. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Chang, K.C.; Costa, N.D.; Blackley, R.; Southwood, O.; Evans, G.; Plastow, G.; Wood, J.D.; Richardson, R.I. Relationships of myosin heavy chain fibre types to meat quality traits in traditional and modern pigs. Meat Sci. 2003, 64, 93–103. [Google Scholar] [CrossRef]
- Heath, M.; Ingram, D.L. Thermoregulatory heat production in cold-reared and warm-reared pigs. Am. J. Physiol. 1983, 244, 273–278. [Google Scholar] [CrossRef]
- Dauncey, M.J.; Ingram, D.L. Acclimatization to warm or cold temperatures and the role of food intake. J. Therm. Biol. 1986, 11, 89–93. [Google Scholar] [CrossRef]
- Bottinelli, R.; Canepari, M.; Reggiani, C.; Stienen, G.J. Myofibrillar ATPase activity during isometric contraction and isomyosin composition in rat single skinned muscle fibres. J. Physiol. 1994, 481, 663–675. [Google Scholar] [CrossRef] [PubMed]
- Dauncey, M.J.; Ingham, D.L. Respiratory enzymes in muscle: Interaction between environmental temperature, nutrition and growth. J. Therm. Biol. 1990, 15, 325–328. [Google Scholar] [CrossRef]
- Bee, G.; Guex, G.; Herzog, W. Free-range rearing of pigs during the winter: Adaptations in muscle fiber characteristics and effects on adipose tissue composition and meat quality traits. J. Anim. Sci. 2004, 4, 1206–1218. [Google Scholar] [CrossRef]
- Lefaucheur, L.; Ecolan, P.; Lossec, G.; Gabillard, J.C.; Butler-Browne, G.S.; Herpin, P. Influence of early postnatal cold exposure on myofiber maturation in pig skeletal muscle. J. Muscle Res. Cell. Motil. 2001, 22, 439–452. [Google Scholar] [CrossRef] [PubMed]
- Gentry, J.G.; Mcglone, J.J.; Miller, M.F.; Blanton, J. Environmental effects on pig performance, meat quality, and muscle characteristics. J. Anim. Sci. 2004, 82, 209–217. [Google Scholar] [CrossRef] [PubMed]
- Sherratt, H.S.A. Mitochondria: Structure and function. Rev. Neurol. 1991, 147, 417–430. [Google Scholar]
- Spina, R.J.; Chi, M.M.; Hopkins, M.G.; Nemeth, P.M.; Lowry, O.H.; Holloszy, J.O. Mitochondrial enzymes increase in muscle in response to 7-10 days of cycle exercise. J. Appl. Physiol. 1996, 80, 2250–2254. [Google Scholar] [CrossRef]
- Bruce, C.R.; Hoy, A.J.; Turner, N.; Watt, M.J.; Allen, T.L.; Carpenter, K.; Cooney, G.J.; Febbraio, M.A.; Kraegen, E.W. Overexpression of carnitine palmitoyltransferase-1 in skeletal muscle is sufficient to enhance fatty acid oxidation and improve high-fat diet–induced insulin resistance. Diabetes 2009, 58, 550–558. [Google Scholar] [CrossRef] [Green Version]
- Hannon, J.P. Effect of prolonged cold exposure on components of the electron transport system. Am. J. Physiol. 1960, 198, 740–744. [Google Scholar] [CrossRef]
- Herpin, S.P. Postnatal Changes in Mitochondrial Protein Mass and Respiration in Skeletal Muscle from the Newborn Pig. Biochem. Mol. Biol. 1997, 118, 639–647. [Google Scholar]
- Brand-Williams, W.M.; Cuvelier, M.E.; Berset, C. Use of free radical method to evaluate antioxidant activity. LWT Food Sci. Technol. 1995, 28, 25–30. [Google Scholar] [CrossRef]
- Djordjevi, V.B. Free radicals in cell biology. Int. Rev. Cytol. 2004, 237, 57–89. [Google Scholar]
- Ramnath, V.; Rekha, P. Brahma Rasayana enhances in vivo antioxidant status in cold-stressed chickens (Gallus gallus domesticus). Indian J. Pharmacol. 2009, 41, 115–119. [Google Scholar] [CrossRef] [Green Version]
- Venditti, P.; Pamplona, R.; Ayala, V.; De Rosa, R.; Caldarone, G.; Di Meo, S. Differential effects of experimental and cold-induced hyperthyroidism on factors inducing rat liver oxidative damage. J. Exp. Biol. 2006, 209, 817–825. [Google Scholar] [CrossRef] [PubMed] [Green Version]
- Zhao, F.-Q.; Zhang, Z.-W.; Qu, J.-P.; Yao, H.-D.; Li, M.; Xu, S.-W. Cold stress induces antioxidants and Hsps in chicken immune organs. Cell Stress Chaperones 2014, 19, 635–648. [Google Scholar] [CrossRef] [PubMed] [Green Version]



{kind=link}
{kind=link}
| Ingredients | Content (%) |
|---|---|
| Extruded corn | 30.00 |
| Corn | 25.00 |
| Soybean meal | 10.50 |
| Extruded soybean | 5.50 |
| Rice screenings | 8.15 |
| Wheat bran | 1.50 |
| Soybean protein concentrate | 4.00 |
| Spray-dried animal plasma | 1.50 |
| Fish meal | 3.50 |
| Whey powder (low protein) | 3.80 |
| Soybean oil | 2.00 |
| Sucrose | 2.00 |
| Limestone | 0.88 |
| Dicalcium phosphate | 0.40 |
| NaCl | 0.30 |
| L-Lysine HCl (78%) | 0.42 |
| DL-Methionine | 0.14 |
| L-Threonine (98.5%) | 0.06 |
| Chloride choline | 0.10 |
| Vitamin premix a | 0.05 |
| Mineral premix b | 0.20 |
| Nutrition level | |
| DE (Mcal/kg) | 3.50 |
| Crude protein | 18.03 |
| Calcium | 0.80 |
| Total phosphorus | 0.56 |
| Available phosphorus | 0.36 |
| Lysine | 1.35 |
| Methionine | 0.44 |
| Threonine | 0.79 |
| Tryptophan | 0.24 |
| Gene Name | Primer | Sequences (5′→3′) | Product Length (bp) | Accession No. |
|---|---|---|---|---|
| β-actin | Forward | GCGGCATCCACGAAACTAC | 138 | DQ-845171.1 |
| Reverse | TGATCTCCTTCTGCATCCTGTC | |||
| MyHC I | Forward | AAGGGCTTGAACGAGGAGTAGA | 114 | AB-053226 |
| Reverse | TTATTCTGCTTCCTCCAAAGGG | |||
| MyHC IIa | Forward | GCTGAGCGAGCTGAAATCC | 137 | AB-025260 |
| Reverse | ACTGAGACACCAGAGCTTCT | |||
| MyHC IIx | Forward | AGAAGATCAACTGAGTGAACT | 149 | AB-025262 |
| Reverse | AGAGCTGAGAAACTAACGTG | |||
| MyHC IIb | Forward | ATGAAGAGGAACCACATTA | 166 | AB-025261 |
| Reverse | TTATTGCCTCAGTAGCTTG | |||
| CS | Forward | GGAAGTCGGCAAAGATGTGT | 162 | NM-214276.1 |
| Reverse | TCATGAGGCAGGTGTTTCAG | |||
| CPT-1b | Forward | GCTATCTGTGTCCGCCTTCT | 151 | NM-001007191.1 |
| Reverse | GGCTGTATTCCTCGTCATCC | |||
| SDHB | Forward | TGTGGTCCTATGGTGTTGGA | 168 | NM-001104953.1 |
| Reverse | TTTGTCGAGGTTGGTGTCAA | |||
| Nrf-1 | Forward | TCCATCAATCCGGAAGAGAC | 170 | XM-021078993.1 |
| Reverse | GCACCACATTCTCCAAAGGT |
| Item | Treatments | p-Value | |
|---|---|---|---|
| CON a | LT b | ||
| ADG, g | 269.21 ± 20.93 | 323.60 ± 36.41 | 0.214 |
| ADFI, g | 428.49 ± 24.95 | 557.72 ± 48.66 | 0.036 |
| F/G | 1.61 ± 0.05 | 1.77 ± 0.07 | 0.073 |
| Item | Treatments | p-Value | |
|---|---|---|---|
| CON | LT | ||
| 1–7 d | |||
| Diarrhea incidence (%) | 31.42 ± 3.40 | 52.38 ± 3.99 | 0.002 |
| Diarrhea index | 0.47 ± 0.07 | 0.90 ± 0.09 | 0.003 |
| 8–14 d | |||
| Diarrhea incidence (%) | 22.86 ± 1.84 | 23.81 ± 1.59 | 0.702 |
| Diarrhea index | 0.27 ± 0.04 | 0.33 ± 0.03 | 0.276 |
| 15–21 d | |||
| Diarrhea incidence (%) | 17.14 ± 3.60 | 20.63 ± 3.78 | 0.516 |
| Diarrhea index | 0.23 ± 0.04 | 0.24 ± 0.04 | 0.880 |
| 1–21 d | |||
| Diarrhea incidence (%) | 23.81 ± 2.12 | 32.28 ± 3.67 | 0.054 |
| Diarrhea index | 0.32 ± 0.04 | 0.49 ± 0.07 | 0.051 |
| Item | Treatments | p-Value | |
|---|---|---|---|
| CON | LT | ||
| CP a, % | 86.13 ± 0.67 | 83.35 ± 0.73 | 0.014 |
| DM b, % | 90.28 ± 0.24 | 89.16 ± 0.30 | 0.012 |
| OM c, % | 91.98 ± 0.23 | 90.96 ± 0.27 | 0.013 |
| GE d, % | 90.60 ± 0.28 | 89.68 ± 0.33 | 0.051 |
| Item | Treatments | p-Value | |
|---|---|---|---|
| CON | LT | ||
| Serum | |||
| MDA, nmol/mL | 3.86 ± 0.18 | 3.91 ± 0.11 | 0.805 |
| T-AOC, U/mL | 0.87 ± 0.05 | 1.25 ± 0.09 | 0.002 |
| T-SOD, U/mL | 145.25 ± 4.16 | 146.50 ± 3.56 | 0.825 |
| LM | |||
| MDA, nmol/mg prot | 0.16 ± 0.01 | 0.14 ± 0.02 | 0.402 |
| T-AOC, U/mg prot | 0.16 ± 0.02 | 0.27 ± 0.03 | 0.006 |
| T-SOD, U/mg prot | 39.31 ± 1.06 | 42.93 ± 1.88 | 0.121 |
| PM | |||
| MDA, nmol/mg prot | 0.24 ± 0.02 | 0.26 ± 0.02 | 0.466 |
| T-AOC, U/mg prot | 0.20 ± 0.02 | 0.24 ± 0.03 | 0.340 |
| T-SOD, U/mg prot | 35.11 ± 1.53 | 40.19 ± 1.62 | 0.039 |
Publisher’s Note: MDPI stays neutral with regard to jurisdictional claims in published maps and institutional affiliations. |
© 2021 by the authors. Licensee MDPI, Basel, Switzerland. This article is an open access article distributed under the terms and conditions of the Creative Commons Attribution (CC BY) license (https://creativecommons.org/licenses/by/4.0/).
Share and Cite
Yu, J.; Chen, S.; Zeng, Z.; Xing, S.; Chen, D.; Yu, B.; He, J.; Huang, Z.; Luo, Y.; Zheng, P.; et al. Effects of Cold Exposure on Performance and Skeletal Muscle Fiber in Weaned Piglets. Animals 2021, 11, 2148. https://doi.org/10.3390/ani11072148
Yu J, Chen S, Zeng Z, Xing S, Chen D, Yu B, He J, Huang Z, Luo Y, Zheng P, et al. Effects of Cold Exposure on Performance and Skeletal Muscle Fiber in Weaned Piglets. Animals. 2021; 11(7):2148. https://doi.org/10.3390/ani11072148
Chicago/Turabian StyleYu, Jie, Shuai Chen, Ziyou Zeng, Shuaibing Xing, Daiwen Chen, Bing Yu, Jun He, Zhiqing Huang, Yuheng Luo, Ping Zheng, and et al. 2021. "Effects of Cold Exposure on Performance and Skeletal Muscle Fiber in Weaned Piglets" Animals 11, no. 7: 2148. https://doi.org/10.3390/ani11072148